1. Introduction
Thyroid cancer is the most common endocrine
malignancy, and its incidence continues to rise worldwide (1,2).
Thyroid cancer is divided into four subtypes: 94% of cases are
well-differentiated thyroid cancer that is derived from follicular
epithelial cells and is categorized as either papillary thyroid
carcinoma (PTC) or follicular thyroid carcinoma (FTC). The other 5%
of cases are medullary thyroid cancer (MTC), a neuroendocrine
tumor. The remaining 1% is anaplastic thyroid carcinoma (ATC),
which is generally derived from de-differentiation of the
differentiated thyroid cancer. Treatment of thyroid cancer is
multifactorial, including surgery, radioiodine therapy and
chemotherapy (1). Despite the
widespread use of multimodal treatment, survival rates have not
significantly improved in the past few decades (2). This suggests that some types of
thyroid cancer are resistant to the current therapeutic
options.
Recent progress in thyroid cancer biology has
revealed that the histological appearance and biological behaviors
of thyroid carcinoma often exhibit heterogeneity, with cells
exhibiting distinct proliferative and differential capacities
(3–5). Emerging evidence indicates that a rare
subpopulation of cells purported to be cancer stem cells (CSCs) or
tumor-initiating cells (TICs) drive thyroid cancer heterogeneity
and contribute to the resistance to cancer therapy as observed in
various cases (6–10). Moreover, clinical observations have
revealed that TIC marker frequency in thyroid cancer is related to
adverse outcomes (11). Thus, TICs
have been indicated to play a crucial role in the malignant
progression and therapeutic resistance of thyroid cancer. The
present review summarizes and evaluates the recent evidence for the
existence of TICs in thyroid cancer and the therapeutic strategies
for targeting TICs based on the increasing biological knowledge of
thyroid TICs.
2. Cellular origin and operational concept
of thyroid tumor-initiating cells
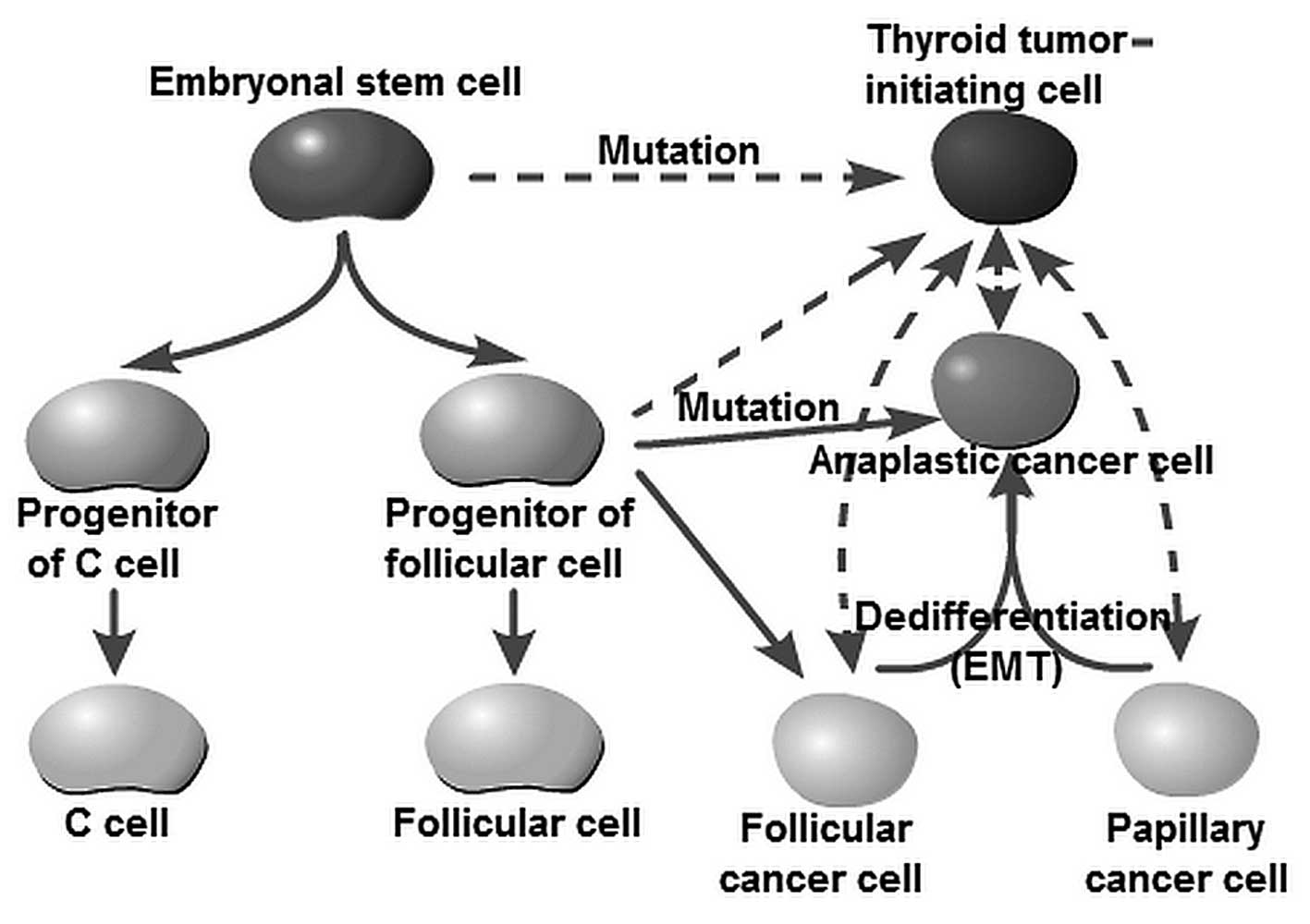
The cellular origins of thyroid cancer are still
largely unknown. Yet, it has been speculated that different
subtypes reflect cells of distinct origin at the time of tumor
initiation. For decades, tumor initiation and development have been
regarded as a multistep process, as reflected by the progressive
genetic alterations that drive the transformation of normal fetal
cells into highly malignant derivatives. The fetal cell
carcinogenesis hypothesis postulated by Takano (12) and Takano and Amino (13) suggests that thyroid cancer cells are
developed from remnant fetal thyroid cells. This model is partly
similar to the current prevailing CSC hypothesis. The ‘CSC’ (often
referred to as the TIC) theory is used to denote a cancer cell
subpopulation with characteristics similar to those of normal stem
cells (14,15). CSCs are characterized by their
capacity to initiate tumor growth, self-renewal and differentiation
by symmetric and asymmetric division (6,7,15). Zhu
et al (16) and Barker et
al (17) observed that CSCs
were derived from normal stem cells if oncogenes were activated. It
should be noted that the concept of a ‘CSC’ does not imply that the
cell is derived from a normal stem cell. Chaffer et al
(18) reported that basal-like
epithelial cells could spontaneously de-differentiate into
stem-like cells. Similar results showed that well-differentiated
PTC or FTC cells could transform into undifferentiated ATC cells
(5,19,20).
Lan et al and Yasui et al as well as other research
groups observed that more differentiated thyroid cancer cell
populations acquire CSC properties through
epithelial-to-mesenchymal transition (EMT) (21–24).
These studies, therefore, raise the possibility that thyroid TICs
may arise from restricted progenitors or more differentiated cells
that have acquired self-renewing capacity. Thus, the originating
tumor cells can be stem cells, progenitor cells or differentiated
cells (Fig. 1). The American
Association for Cancer Research (AACR) workshop stated that ‘cancer
stem cells can only be defined experimentally by their ability to
recapitulate the generation of a continuously growing tumor’
(25). Furthermore, the most widely
accepted assay with which to validate a candidate CSC population is
tumor initiation and serial transplantation in immunocompromised
mice (26). Based on the
consideration of cellular origin and functional identification, we
herein recommend the use of the term ‘TIC’ instead of ‘CSC’ to
describe this stem cell-like subpopulation within thyroid
cancer.
3. Isolation and identification of thyroid
tumor-initiating cells
Although the method is debated, putative TICs
derived from thyroid carcinoma cell lines and tumor biopsy
specimens were recently isolated using current prevailing cancer
stem cell methods (27–30).
The approaches used to identify and isolate these
putative TICs were based on the notion that TICs should have
conserved stem and progenitor cell functions and phenotypes. TICs
have been frequently isolated using specific markers from normal
stem cells of the same organ (14,15).
According to the literature, TICs can be isolated by the following
four methodologies: flow cytometry-based cell sorting according to:
i) TIC-specific cell surface markers; ii) aldehyde dehydrogenase
activity; iii) ABCG2 efflux-pump-mediated Hoechst 33342 dye
exclusion; and iiii) generation of spherical colonies in an
ultra-low-attachment and serum-free culture. The following sections
will discuss the various techniques that have to date been used in
an attempt to isolate TICs in thyroid cancer as well as the
limitations of these techniques.
Surface markers
Surface markers have been used to identify and
isolate adult stem/progenitor cells in human thyroid glands
(31). Recent studies have been
successful in using normal antigenic stem cell markers for the
isolation of putative TICs in thyroid cancer. A promising marker
that has been gaining popularity for the identification of thyroid
TICs is CD133, also known as prominin-1 (32). CD133 is a 120-kDa cell surface
protein comprising five transmembrane domains and was first
described as a hematopoietic stem cell marker (33). Zito et al (34) found that CD133-positive cells
comprised 64 and 57% of the cells within the undifferentiated ATC
cell lines ARO and KAT-4, respectively. Sorted ARO
CD133+ cells exhibit stem cell-like features such as
increased proliferation, self-renewal ability, clone formation and
resistance to chemotherapeutic agents (doxorubicin, cisplatin and
etoposide). Friedman et al (35) detected CD133+ cells in
two ATC cell lines, namely ARO (7.02%) and FRO (6.32%), but not in
well-differentiated PTC cell lines (NPA and WRO). CD133+
cells harbor stem cell features characterized by high expression of
stem cell gene Oct-4 and rapid long-term tumorigenesis. Ke et
al (8) recently reported that
CD133+ cells were present not only in the ATC cell line
ARO (61.3%), but also in the PTC cell line CG3 (5%) and FTC cell
line WRO (1.5%). A further study demonstrated that these
CD133+ cells showed higher radioresistance and an
undifferentiated status. Additionally, Zhu et al (36) suggested that the CD133+
subpopulation from MTC cell lines (MZ-CRC-1 and TT) exhibited the
TIC features of self-renewal and multiple lineage
differentiation.
Strikingly, the ATC cell line ARO used in the
above-mentioned studies showed a significantly different
CD133+ fraction ranging from 7.02 to 64% (8,34,35).
In contrast, Friedman et al (35) did not detect CD133+ cells
while Ke et al (8) found a
1.5% CD133-expressing fraction within the WRO PTC cell line. The
frequency of CD133-positive cells appears to be highly variable
among thyroid cancers of the same type. This issue may be related
to the different cell culture environments and method standards in
individual laboratories. In addition, a recent study provided
evidence that the presence of TICs was frequently correlated with
the tumor grade and clinical outcome (11). Thus, primary thyroid cancer cells
should be used to verify the value of CD133 markers in the
isolation of thyroid TICs. Furthermore, various surface markers
such as Oct-4, GATA binding protein 4 (GATA4) and hepatocyte
nuclear factor 4α (HNF4α) have been used to identify human thyroid
stem/progenitor cells (31,37). Based on the notion that TICs
frequently express specific markers from normal stem cells of a
certain organ, it would be valuable to identify these markers in
thyroid cancer.
Side population assay
The side population (SP) assay is based on the
differential potential of cells to efflux Hoechst dye (e.g.,
Hoechst 33342) via the ATP-binding cassette (ABC) family of
transporter proteins expressed within the cell membrane (27). The ability of ABC transporters to
rapidly efflux lipophilic fluorescent dyes in vitro serves
as the basis of the SP assay and was originally identified in mouse
bone marrow cells (38). SP cells
were found, not only to have stem cell characteristics, but also to
be enriched in a stem cell population (39). Hoshi et al (40) indicated that mouse thyroid SP cells
are less differentiated and have characteristics of stem/progenitor
cells. Thus, the SP assay has now emerged as a promising method for
identifying stem-like cells in thyroid cancer. Mitsutake et
al (41) reported that
well-differentiated PTC (NPA) and FTC (WRO) cells or
undifferentiated ATC (ARO and FRO) cell lines contained SP cells
(excluding TPC-1 PTC cells) ranging from 0.02 to 0.25% of the total
viable cell population. In another study by Zheng et al
(9), ATC cell lines (SW1736, C643
and HTh74) were found to comprise 0.43 to 0.83% of SP cells,
indicating that the SP fraction varies among thyroid cancer cell
lines. The SP cells within thyroid cancer cell lines displayed TIC
features characterized by overexpression of embryonic stem cell
genes, high tumorigenic potential following transplantation into
immunocompromised mice, increased self-renewal capacity
(thyrospheres), generation of non-SP cells and resistance to
chemotherapeutic agents (doxorubicin).
Theoretically, this method may be more suitable for
enrichment of potential TICs because it is less restricted by
tissue specificity than the cell surface makers discussed above.
However, there are some notable limitations and concerns regarding
the use of this technique to isolate putative TICs. First,
Mitsutake et al (41) were
not able to detect the SP fraction in the PTC cell line TPC-1,
indicating that SP cells may represent only one of the putative
thyroid TIC populations. Furthermore, Steuer et al (42) observed that Hoechst 33342 could
induce the differentiation of mouse embryonal carcinoma cell lines
(PCC3, P19 and PCC4); this effect may cause confounding errors when
introduced to stem cell sorting. Adhikari et al (43) found that Hoechst 33342 arrests human
glioma and squamous carcinoma cell growth and the cell cycle (late
S and G2 phases), and increases cytogenetic damage (micronucleus
formation) under ionizing radiation exposure. Therefore, Hoechst
dye may limit the functional analysis of the enriched SP
fraction.
Sphere formation assay
The sphere-forming assay is widely used in stem cell
biology as this is a relatively simple yet robust method for
isolating and expanding stem cell populations (28). Since the original sphere formation
assay was used to isolate neural stem cells, cells with stem-like
characteristics have been isolated from human goiters and thyroid
cancer cells by culturing them as non-adherent spheres and
characterizing them as stem/progenitor cells. Pillai et al
(44) and Malaguarnera et al
(45) reported that thyrospheres
isolated from PTC-derived cells have stem-like properties
characterized by the expression of stem cell markers and low/absent
thyroid-specific markers. Furthermore, Malaguarnera et al
found that thyrospheres overexpress insulin receptor (IR) isoforms
and the IGF receptor (IGF-IR) and that treatment with IGF increases
the stemness features of thyrospheres. Li et al (46) observed that ATC cells from thyroid
cancer patients also formed thyrospheres. These thyrospheres
expressed stem cell markers and exhibited tumorigenic and
metastatic features in an orthotopic mouse model.
However, there is little definitive information
regarding which cancer cells are propagated under these
non-adherent and serum-free conditions. Visvader and Lindeman
(47)observed that the breast
cancer cells from the sphere were no more tumorigenic in
vivo, and that cells from malignant pleural effusions (from
patients with breast cancer) did not give rise to tumors after more
than 10 months following fat-pad implantation, despite generating
spheres in culture. Therefore, the sphere-formation ability is not
restricted to TICs and self-renewal cannot solely be defined in the
context of a sphere assay.
Aldehyde dehydrogenase activity
Aldehyde dehydrogenase (ALDH) is an
NAD(P)+-dependent enzyme involved in the detoxification
of intracellular aldehydes to weak carboxylic acids and is
significantly more highly expressed in stem/progenitor cells
(29,48,49).
In several types of cancer, ALDH, which plays a crucial role in
stem cell biology, has emerged as a valuable functional marker for
the isolation of TICs (48–51). The basis of the ALDH assay is that
intracellular ALDH converts the uncharged ALDH substrate
BODIPY-aminoacetaldehyde (BAAA) into the negatively charged
BAA−, which is retained intracellularly and causes the
cell to become highly fluorescent. Thus, the ALDH-positive and
ALDH-negative cells can be separated based on intracellular ALDH
activity (29). Todaro et al
(10) found that FTC, PTC and UTC
cells derived from thyroid cancer tissue contain a small population
of tumorigenic cells that can be prospectively identified through
ALDH activity. Thyroid cells with high ALDH expression
(ALDHhigh) possess the ability to self-renew and
re-initiate serial transplantable tumors that recapitulate the
phenotype and metastatic behavior of the parental tumors promoted
by the activation of Met and Akt. In another study, Klonisch et
al (52) identified a 17–38%
ALDH1+ cell subpopulation with potential stem-like
characteristics in the ATC cell line UTC-8505C. Additionally,
Carina et al (53) recently
observed that the ATC cell line SW 1736 contained a high percentage
of ALDH-positive cells (10.4±2.1%) and exhibited high expression of
several TIC markers (Sox-2, Oct4, Nanog, c-myc and SSEA4).
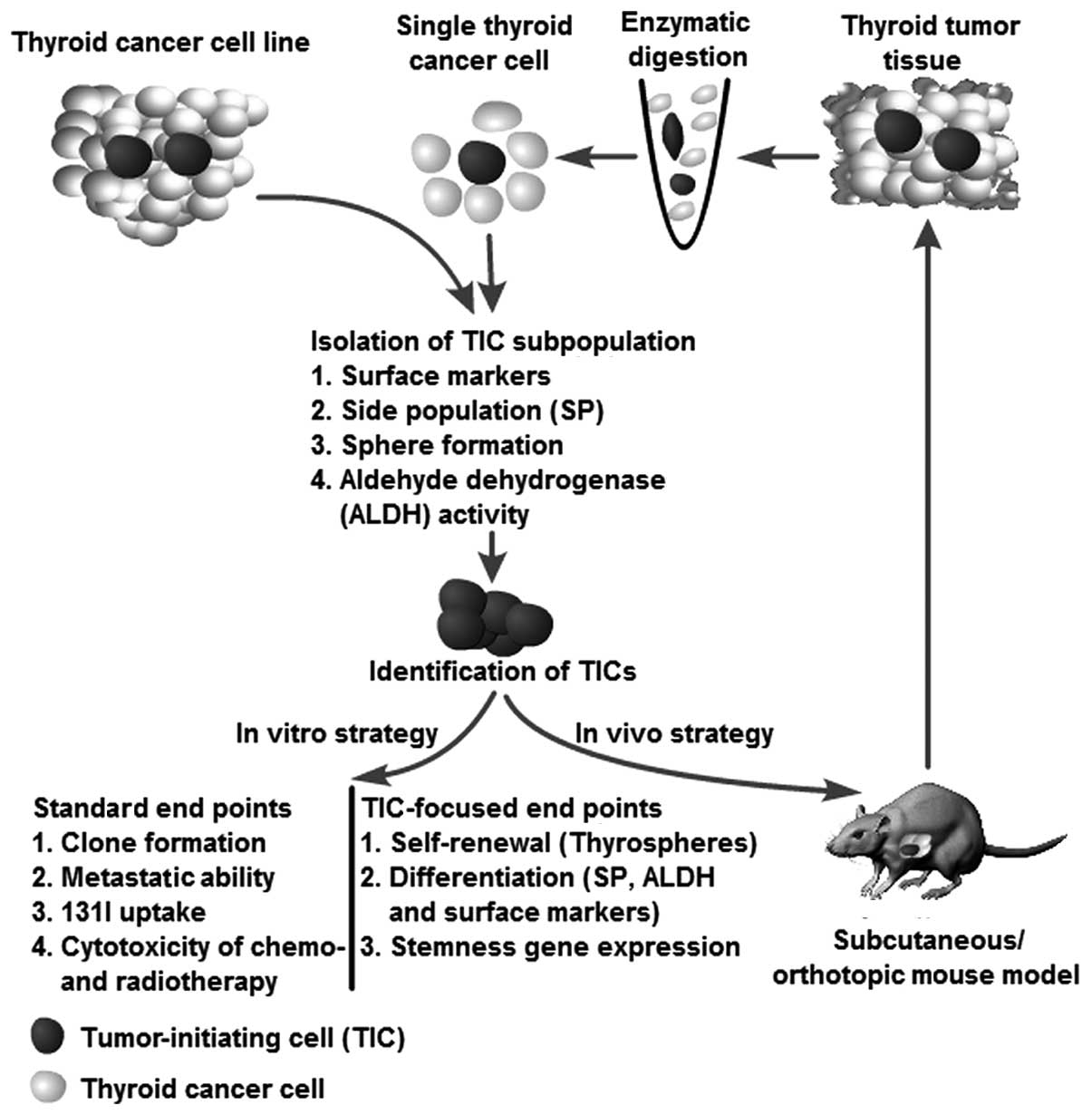
Identification of thyroid
tumor-initiating cells
A series of strategies have been used to verify
thyroid TICs isolated from patients with thyroid cancer and from
cell lines using the methods mentioned above. The methods used to
isolate TICs can also be used as strategies to identify putative
TICs. For instance, the putative TICs isolated from an SP assay can
validate their TIC characteristics through a sphere assay, ALDH
activity, and the expression of surface markers (e.g., CD133) and
vice versa. The strategies for identification of thyroid TICs are
illustrated in Fig. 2.
Generally, putative TICs should demonstrate the
ability to generate spheres in serum-free medium, even after serial
passage, thus indicating that the cells have an extensive capacity
for self-renewal. Furthermore, the cells should be able, not only
to generate a heterogeneous tumor cell population, indicating
multipotent potential, but also should have an unlimited capacity
to reconstitute the primary tumor morphology, thus, demonstrating
that heterogeneity is not simply a result of genetic instability.
Moreover, surface markers can be used to identify their cellular
origin, stemness and differentiation status. As an in vivo
strategy, serial transplantation is the golden standard recommended
by the AACR workshop to determine stem cell properties (25). The serial transplantation assay
reflects the tumor-initiating ability of a certain cell population.
If the putative TICs demonstrate the characteristics mentioned
above, they can be recognized as TICs (Fig. 2).
It should be noted that not all TICs harbor these
features. The different methods for the isolation of TICs may not
overlap for the same tumor type or may do so to a limited degree.
For instance, Todaro et al (10) observed that sphere-derived cells but
not the ALDHhigh thyroid cells from the same FTC5 tumors
could initiate tumors in NOD/SCID mice. The most reasonable
explanation for this observation may be related to the xenograft
model itself. Species-specific differences and residual immune
effects impair the tumor-initiating frequency. Furthermore, the
ability of tumor initiation may be more accurately evaluated using
an orthotopic transplantation to mimic the tumor environment as
closely as possible. Finally, most studies of thyroid TICs have
used thyroid cancer cell lines. Schweppe et al (54) reported that up to 42% of 40 types of
thyroid cancer cells used in different laboratories were
misidentified, redundant or cross-contaminated. In addition,
repeated passaging of cell lines was found to lead to changes in
characteristics and surface markers and the acquisition of genetic
aberrations. Thus, observations made in thyroid cancer cell lines
must be extended to primary tumors to validate their
significance.
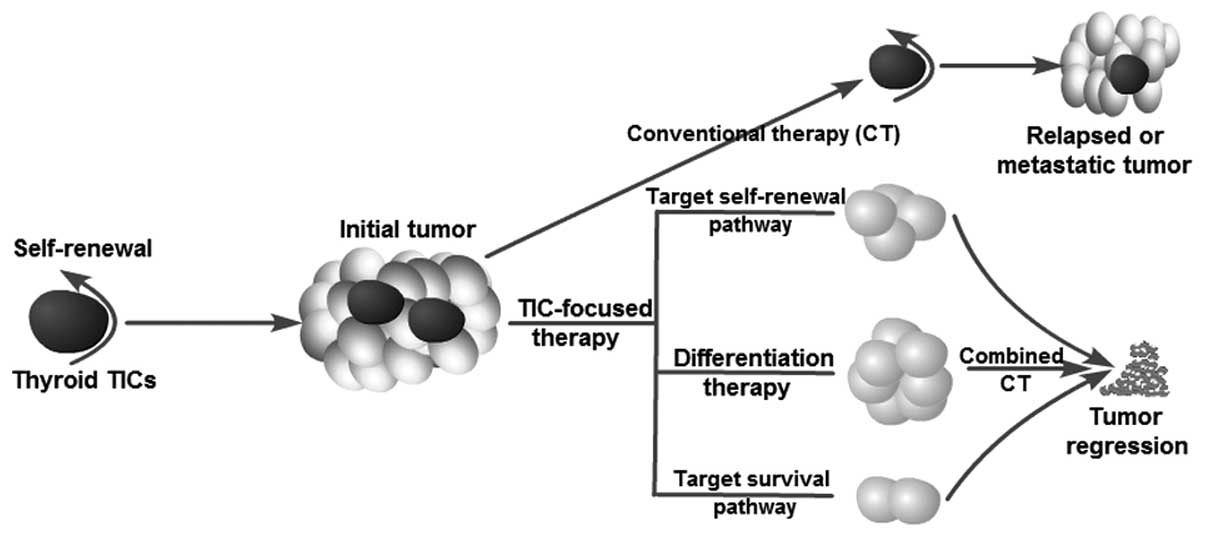
4. Therapeutic implications
Current anticancer treatments are often able to
destroy the bulk of a tumor but spare TICs. Thus, relapse or tumor
recurrence is not rare after a primary therapeutic response or
initial induction of tumor remission. The existence of TICs
explains the phenomenon observed in clinical therapy. Zheng et
al (9) and Ke et al
(8) recently found that the failure
of chemotherapy (doxorubicin) or radiotherapy (10 Gy) to eradicate
thyroid cancer cells may be due to the ineffective targeting of
thyroid TICs. An ideal therapeutic strategy should not only kill
differentiated cancer cells (as conventional therapy does) but
should also specifically destroy thyroid TICs simultaneously. Thus,
the focus on thyroid TICs has significant clinical implications for
the development of novel therapeutical strategies for refractory
thyroid cancer (such as lack of sufficient radioiodine uptake and
resistance to current chemotherapy) by targeting TICs. In fact,
several studies have investigated possible strategies by which to
target thyroid TICs and overcome resistance to current therapeutic
options (Fig. 3). Potential
approaches to killing TICs include blocking self-renewal signaling
pathways, inducing tumor cell differentiation and inhibiting
survival mechanisms, thus overcoming chemoresistance and
radioresistance.
Targeting self-renewal signaling
pathways
Approaches to blocking the self-renewal of TICs
represent rational therapeutic strategies for cancer prevention and
treatment. The signal transducer and activator of transcription 3
(STAT3) pathway has been shown to be required for self-renewal of
TICs in several types of cancers, including hepatoma and
glioblastoma (55,56). Microarray bioinformatic analysis
from ATC-derived cells recently revealed that the STAT3 pathway
plays an essential role for maintaining the self-renewal of
ATC-CD133+ cells, since blockage of STAT3 activity using
cucurbitacin I was found to significantly diminish TIC features
such as stemness gene expression, and induce their differentiation.
Cucurbitacin I plus radiochemotherapy suppressed CD133+
cell survival and tumorigenesis in vitro and in vivo,
respectively (57). Sox-2 is a
well-established regulator in the maintenance of adult tissue
homeostasis and regeneration (58).
Accumulating data indicate that Sox-2 expression is abnormally
elevated in thyroid TICs, thus, serving as a therapeutic target.
Carina et al (53) found
that the silencing of Sox-2 decreased the expression level of key
stemness genes (Nanog and Oct4) and increased the sensitivity to
chemotherapeutic agent (cisplatin and doxorubicin)-induced cell
death in the ATC SW1736 cell line. In addition, Malaguarnera et
al (45) reported that
overexpression of the insulin receptor and IGF-I receptor in
thyrospheres may sustain their self-renewal capacity and promote
TIC survival. Metformin suppression of the self-renewal of derived
thyroid TICs appears to be related to inhibition of the insulin/IGF
and AMPK signaling pathways, but this finding still requires
further investigation (59). The
thyrotropin receptor, which has been found to be highly expressed
in CD133+ cells, also plays a crucial role in
maintaining stemness, thus serving as another potential target
(35). Finally, Zhu et al
(36) found that the RET tyrosine
kinase receptor gene (RET) signaling pathway, which plays a
fundamental role in stem cell maintenance and development of MTC
contributes to tumor progression through facilitation of TIC
(thyrosphere) renewal. Knockdown of RET expression reduced the
sphere formation ability (a cardinal features of TICs) of MTC
cells. This suggests that targeting of the RET receptor offer
another opportunity to eradicate TIC populations (60,61).
Differentiation therapy
TICs facilitate tumor growth through enhanced
self-renewal and limited differentiation capacity. Therefore, an
alternative approach to the eradication of TICs may be achievable
by overcoming the blockage of differentiation by TICs. In the
1970s, this approach led to the identification and clinical use of
all-trans-retinoic acid for patients with acute
promyelocytic leukemia (62) and
metastatic thyroid cancer (63).
Tseng et al (57) found that
cucurbitacin I suppressed the proliferation and TIC properties of
CD133+ cells through induction of their differentiation
into CD133− cells characterized by upregulation of
thyroid-specific genes (NIS, TPO and Tg) and enhanced radioiodine
uptake. The effects of cucurbitacin I on normal thyroid
stem/progenitor cells remain unclear. Of note, differentiation
therapy should involve selective targeting of TICs while having no
effect on normal stem cells.
Targeting of other implicated molecular
pathways
The cell surface molecule CD44 may also be an
important target. CD44 is a transmembrane glycoprotein involved in
tumor cell proliferation and metastasis (64). De Falco et al (65) revealed that the CD44-CERB signaling
pathway is required to sustain thyroid cancer cells, as evidenced
by the finding that knockdown of CD44 obstructed cell
proliferation. CD44 was recently recognized as a promising marker
of TICs in many cancer types, including thyroid cancer (10,66).
Thus, CD44 serves as a potential cell surface marker for targeting
TICs. Other described targets include ABCG2 (breast
cancer-resistance protein, BRCP1) and ABCB1 (P-glycoprotein, MDR1),
which were found to be overexpressed in doxorubicin-resistant ATC
cancer cells (9). ABCG2 and MDR1
are the best-characterized drug transporter proteins belonging to
the ABC family that function to establish the SP phenotype
(27,67,68).
TICs express high levels of ABC drug pumps, which enable the efflux
of drugs from cells, thus serving to protect them from
chemotherapeutic agents (68,69).
Silencing of ABCG2 expression increases ATC cell sensitivity to
cisplatin and doxorubicin treatment (53). Finally, Todaro et al
(10) reported that enhanced cell
survival and migratory ability are associated with increased Met
and Akt expression in ALDHhigh thyroid cancer cells,
whereas silencing of these genes was found to decrease the
tumorigenic and metastatic capacities.
131I therapy and thyroid
tumor-initiating cells
131I therapy is regularly used following
surgery as a part of thyroid cancer management. Feng et al
(70) observed that
undifferentiated thyroid cancer cell populations were enriched
after 131I treatment, which indicates that TICs involved
in thyroid cancer may be resistant to 131I therapy.
Failure to accumulate iodide and resistance to ionizing radiation
(IR) may be the two main reasons for TIC insensitivity to
131I therapy. Undifferentiated thyroid carcinomas have
an absent or decreased ability for iodide uptake due to decreased
or undetected sodium-iodide symporter (NIS) expression (71). As mentioned in the section,
‘Differentiation therapy’, cucurbitacin I induces CD133+
cell differentiation by upregulation of NIS expression and thus
enhances radioiodine uptake (57).
Therefore, differentiation therapy may be a promising method for
restoration of the 131I uptake by TICs. A phase II study
used retinoids as redifferentiation agents to increase the iodine
uptake for metastatic thyroid cancer (63). Furthermore, accumulating evidence
suggests that TIC resistance to ionizing radiation results from
quiescence propensity, enhanced DNA repair, upregulated cell cycle
control mechanisms, mechanisms of free-radical scavenging, and
specific interaction with the stromal microenvironment (72,73).
Tseng et al (57) and Ke
et al (8) reported that
CD133+ thyroid cancer cells exhibit reduced IR-induced
cell death, particularly apoptosis. The critical mechanisms
determining the resistance of thyroid TICs to radiation remain
elusive and warrant further investigation. TICs can be effectively
eradicated by combined methods that overcome IR resistance and
restore radioiodine uptake with 131I therapy.
5. Future direction
Although accumulating evidence supports the
existence of TICs in thyroid cancer, and potential therapeutic
strategies for targeting thyroid TICs are currently underway, some
unanswered questions remain. First, the most basic cellular origins
of thyroid TICs are transplantations from other malignancies. Thus,
the origin of thyroid TICs should be explored in further studies to
prevent tumor initiation. Second, the TIC fractions in thyroid
cancer are highly impure, and the reported frequencies in the same
tumor types have varied enormously among different research groups.
This reflects the fact that TICs themselves are heterogeneous. This
highlights the importance to identify additional specific markers
or to use combinations of markers. Finally, the existence of TICs
may depend on the specific niche. Accumulating evidence suggests
that the tumor microenvironment is necessary for sustaining the
stem-like properties of TICs (74,75).
Thus, it is desirable to establish more definitive methods by which
the tumor microenvironment can be mimicked when isolating and
identifying TICs. On the other hand, the TIC niche could,
therefore, be a therapeutic target for the design of therapies
aimed at eradicating thyroid TICs. Studies concerning the thyroid
TIC niche are rare, and thus this subject should be investigated
in-depth. An increased understanding of thyroid TIC biology will
provide an important framework for drug discovery and cancer
treatment with the potential to identify novel antitumor
activities, impact cancers with undifferentiated phenotypes, and
yield long-term benefits for patients with thyroid cancer.
References
|
1
|
Sherman SI: Thyroid carcinoma. Lancet.
361:501–511. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar
|
|
3
|
Pilotti S, Collini P, Manzari A, Marubini
E and Rilke F: Poorly differentiated forms of papillary thyroid
carcinoma: distinctive entities or morphological patterns? Semin
Diagn Pathol. 12:249–255. 1995.PubMed/NCBI
|
|
4
|
Nishida T, Katayama S, Tsujimoto M,
Nakamura J and Matsuda H: Clinicopathological significance of
poorly differentiated thyroid carcinoma. Am J Surg Pathol.
23:205–211. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Aratake Y, Nomura H, Kotani T, et al:
Coexistent anaplastic and differentiated thyroid carcinoma: an
immunohistochemical study. Am J Clin Pathol. 125:399–406. 2006.
View Article : Google Scholar
|
|
6
|
Rosen JM and Jordan CT: The increasing
complexity of the cancer stem cell paradigm. Science.
324:1670–1673. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gupta PB, Chaffer CL and Weinberg RA:
Cancer stem cells: mirage or reality? Nat Med. 15:1010–1012. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ke CC, Liu RS, Yang AH, et al:
CD133-expressing thyroid cancer cells are undifferentiated,
radioresistant and survive radioiodide therapy. Eur J Nucl Med Mol
Imaging. 40:61–71. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zheng X, Cui D, Xu S, Brabant G and
Derwahl M: Doxorubicin fails to eradicate cancer stem cells derived
from anaplastic thyroid carcinoma cells: characterization of
resistant cells. Int J Oncol. 37:307–315. 2010.PubMed/NCBI
|
|
10
|
Todaro M, Iovino F, Eterno V, et al:
Tumorigenic and metastatic activity of human thyroid cancer stem
cells. Cancer Res. 70:8874–8885. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yun JY, Kim YA, Choe JY, et al: Expression
of cancer stem cell markers is more frequent in anaplastic thyroid
carcinoma compared to papillary thyroid carcinoma and is related to
adverse clinical outcome. J Clin Pathol. Aug 28–2013.(Epub ahead of
print). View Article : Google Scholar
|
|
12
|
Takano T: Fetal cell carcinogenesis of the
thyroid: theory and practice. Semin Cancer Biol. 17:233–240. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Takano T and Amino N: Fetal cell
carcinogenesis: a new hypothesis for better understanding of
thyroid carcinoma. Thyroid. 15:432–438. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ratajczak MZ: Cancer stem cells - normal
stem cells ‘Jedi’ that went over to the ‘dark side’. Folia
Histochem Cytobiol. 43:175–181. 2005.
|
|
15
|
Dalerba P, Cho RW and Clarke MF: Cancer
stem cells: models and concepts. Annu Rev Med. 58:267–284. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhu L, Gibson P, Currle DS, et al:
Prominin 1 marks intestinal stem cells that are susceptible to
neoplastic transformation. Nature. 457:603–607. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Barker N, Ridgway RA, van Es JH, et al:
Crypt stem cells as the cells-of-origin of intestinal cancer.
Nature. 457:608–611. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chaffer CL, Brueckmann I, Scheel C, et al:
Normal and neoplastic nonstem cells can spontaneously convert to a
stem-like state. Proc Natl Acad Sci USA. 108:7950–7955. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kondo T, Ezzat S and Asa SL: Pathogenetic
mechanisms in thyroid follicular-cell neoplasia. Nat Rev Cancer.
6:292–306. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Albores-Saavedra J, Hernandez M,
Sanchez-Sosa S, Simpson K, Angeles A and Henson DE: Histologic
variants of papillary and follicular carcinomas associated with
anaplastic spindle and giant cell carcinomas of the thyroid: an
analysis of rhabdoid and thyroglobulin inclusions. Am J Surg
Pathol. 31:729–736. 2007. View Article : Google Scholar
|
|
21
|
Lan L, Luo Y, Cui D, et al:
Epithelial-mesenchymal transition induces cancer stem cell
generation in human thyroid cancer cells in vitro. Zhonghua Yi Xue
Za Zhi. 93:1261–1265. 2013.(In Chinese).
|
|
22
|
Yasui K, Shimamura M, Mitsutake N and
Nagayama Y: SNAIL induces epithelial-to-mesenchymal transition and
cancer stem cell-like properties in aldehyde
dehydroghenase-negative thyroid cancer cells. Thyroid. 23:989–996.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mani SA, Guo W, Liao MJ, et al: The
epithelial-mesenchymal transition generates cells with properties
of stem cells. Cell. 133:704–715. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Polyak K and Weinberg RA: Transitions
between epithelial and mesenchymal states: acquisition of malignant
and stem cell traits. Nat Rev Cancer. 9:265–273. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Clarke MF, Dick JE, Dirks PB, et al:
Cancer stem cells - perspectives on current status and future
directions: AACR Workshop on Cancer Stem Cells. Cancer Res.
66:9339–9344. 2006. View Article : Google Scholar
|
|
26
|
Ailles LE and Weissman IL: Cancer stem
cells in solid tumors. Curr Opin Biotechnol. 18:460–466. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Golebiewska A, Brons NH, Bjerkvig R and
Niclou SP: Critical appraisal of the side population assay in stem
cell and cancer stem cell research. Cell Stem Cell. 8:136–147.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pastrana E, Silva-Vargas V and Doetsch F:
Eyes wide open: a critical review of sphere-formation as an assay
for stem cells. Cell Stem Cell. 8:486–498. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Douville J, Beaulieu R and Balicki D:
ALDH1 as a functional marker of cancer stem and progenitor cells.
Stem Cells Dev. 18:17–26. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Greve B, Kelsch R, Spaniol K, Eich HT and
Gotte M: Flow cytometry in cancer stem cell analysis and
separation. Cytometry A. 81:284–293. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Thomas T, Nowka K, Lan L and Derwahl M:
Expression of endoderm stem cell markers: evidence for the presence
of adult stem cells in human thyroid glands. Thyroid. 16:537–544.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Grosse-Gehling P, Fargeas CA, Dittfeld C,
et al: CD133 as a biomarker for putative cancer stem cells in solid
tumours: limitations, problems and challenges. J Pathol.
229:355–378. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Miraglia S, Godfrey W, Yin AH, et al: A
novel five-transmembrane hematopoietic stem cell antigen:
isolation, characterization, and molecular cloning. Blood.
90:5013–5021. 1997.PubMed/NCBI
|
|
34
|
Zito G, Richiusa P, Bommarito A, et al:
In vitro identification and characterization of
CD133poscancer stem-like cells in anaplastic thyroid
carcinoma cell lines. PLoS One. 3:e35442008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Friedman S, Lu M, Schultz A, Thomas D and
Lin RY: CD133+anaplastic thyroid cancer cells initiate
tumors in immunodeficient mice and are regulated by thyrotropin.
PLoS One. 4:e53952009.
|
|
36
|
Zhu W, Hai T, Ye L and Cote GJ: Medullary
thyroid carcinoma cell lines contain a self-renewing
CD133+population that is dependent on Ret proto-oncogene
activity. J Clin Endocrinol Metab. 95:439–444. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lin RY: New insights into thyroid stem
cells. Thyroid. 17:1019–1023. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Goodell MA, Brose K, Paradis G, Conner AS
and Mulligan RC: Isolation and functional properties of murine
hematopoietic stem cells that are replicating in vivo. J Exp Med.
183:1797–1806. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Robinson SN, Seina SM, Gohr JC, Kuszynski
CA and Sharp JG: Evidence for a qualitative hierarchy within the
Hoechst-33342 ‘side population’ (SP) of murine bone marrow cells.
Bone Marrow Transplant. 35:807–818. 2005.PubMed/NCBI
|
|
40
|
Hoshi N, Kusakabe T, Taylor BJ and Kimura
S: Side population cells in the mouse thyroid exhibit
stem/progenitor cell-like characteristics. Endocrinology.
148:4251–4258. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Mitsutake N, Iwao A, Nagai K, et al:
Characterization of side population in thyroid cancer cell lines:
cancer stem-like cells are enriched partly but not exclusively.
Endocrinology. 148:1797–1803. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Steuer B, Breuer B and Alonso A:
Differentiation of EC cells in vitro by the fluorescent dye
Hoechst 33342. Exp Cell Res. 186:149–157. 1990.PubMed/NCBI
|
|
43
|
Adhikari JS, Khaitan D, Arya MB and
Dwarakanath BS: Heterogeneity in the radiosensitizing effects of
the DNA ligand Hoechst-33342 in human tumor cell lines. J Cancer
Res Ther. 1:151–161. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Pillai R, Caria P, Cabras S, Saba F, Vanni
R and Sogos V: Thyrospheres enriched in stem-like cells from B-CPAP
thyroid cancer cell line: morphomolecular characterization. Italian
J Anat Embryol. 116:1432011.
|
|
45
|
Malaguarnera R, Frasca F, Garozzo A, et
al: Insulin receptor isoforms and insulin-like growth factor
receptor in human follicular cell precursors from papillary thyroid
cancer and normal thyroid. J Clin Endocrinol Metab. 96:766–774.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Li W, Reeb AN, Sewell WA, Elhomsy G and
Lin RY: Phenotypic characterization of metastatic anaplastic
thyroid cancer stem cells. PLoS One. 8:e650952013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Visvader JE and Lindeman GJ: Cancer stem
cells in solid tumours: accumulating evidence and unresolved
questions. Nat Rev Cancer. 8:755–768. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ginestier C, Hur MH, Charafe-Jauffret E,
et al: ALDH1 is a marker of normal and malignant human mammary stem
cells and a predictor of poor clinical outcome. Cell Stem Cell.
1:555–567. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Han M and Wu C: Advancement of studies on
ALDH1 as a universal marker of stem cells. Sheng Wu Yi Xue Gong
Cheng Xue Za Zhi. 27:1183–1186. 2010.(In Chinese).
|
|
50
|
Feng JQ, Xu ZY, Shi LJ, Wu L, Liu W and
Zhou ZT: Expression of cancer stem cell markers ALDH1 and Bmi1 in
oral erythroplakia and the risk of oral cancer. J Oral Pathol Med.
42:148–153. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Shenoy A, Butterworth E and Huang EH: ALDH
as a marker for enriching tumorigenic human colonic stem cells.
Methods Mol Biol. 916:373–385. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Klonisch T, Hoang-Vu C and
Hombach-Klonisch S: Thyroid stem cells and cancer. Thyroid.
19:1303–1315. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Carina V, Zito G, Pizzolanti G, et al:
Multiple pluripotent stem cell markers in human anaplastic thyroid
cancer: the putative upstream role of SOX2. Thyroid. 23:829–837.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Schweppe RE, Klopper JP, Korch C, et al:
Deoxyribonucleic acid profiling analysis of 40 human thyroid cancer
cell lines reveals cross-contamination resulting in cell line
redundancy and misidentification. J Clin Endocrinol Metab.
93:4331–4341. 2008. View Article : Google Scholar
|
|
55
|
Lee TK, Castilho A, Cheung VC, Tang KH, Ma
S and Ng IO: CD24+liver tumor-initiating cells drive
self-renewal and tumor initiation through STAT3-mediated NANOG
regulation. Cell Stem Cell. 9:50–63. 2011.
|
|
56
|
Kim E, Kim M, Woo DH, et al:
Phosphorylation of EZH2 activates STAT3 signaling via STAT3
methylation and promotes tumorigenicity of glioblastoma stem-like
cells. Cancer Cell. 23:839–852. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Tseng LM, Huang PI, Chen YR, et al:
Targeting signal transducer and activator of transcription 3
pathway by cucurbitacin I diminishes self-renewing and
radiochemoresistant abilities in thyroid cancer-derived
CD133+cells. J Pharmacol Exp Ther. 341:410–423. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Sarkar A and Hochedlinger K: The sox
family of transcription factors: versatile regulators of stem and
progenitor cell fate. Cell Stem Cell. 12:15–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Chen G, Xu S, Renko K and Derwahl M:
Metformin inhibits growth of thyroid carcinoma cells, suppresses
self-renewal of derived cancer stem cells, and potentiates the
effect of chemotherapeutic agents. J Clin Endocrinol Metab.
97:E510–E520. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Messina M and Robinson BG: Technology
insight: gene therapy and its potential role in the treatment of
medullary thyroid carcinoma. Nat Clin Pract Endocrinol Metab.
3:290–301. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Naughton CK, Jain S, Strickland AM, Gupta
A and Milbrandt J: Glial cell-line derived neurotrophic
factor-mediated RET signaling regulates spermatogonial stem cell
fate. Biol Reprod. 74:314–321. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wang ZY and Chen Z: Acute promyelocytic
leukemia: from highly fatal to highly curable. Blood.
111:2505–2515. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Short SC, Suovuori A, Cook G, Vivian G and
Harmer C: A phase II study using retinoids as redifferentiation
agents to increase iodine uptake in metastatic thyroid cancer. Clin
Oncol (R Coll Radiol). 16:569–574. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Naor D, Nedvetzki S, Golan I, Melnik L and
Faitelson Y: CD44 in cancer. Crit Rev Clin Lab Sci. 39:527–579.
2002. View Article : Google Scholar
|
|
65
|
De Falco V, Tamburrino A, Ventre S, et al:
CD44 proteolysis increases CREB phosphorylation and sustains
proliferation of thyroid cancer cells. Cancer Res. 72:1449–1458.
2012.PubMed/NCBI
|
|
66
|
Liu J and Brown RE: Immunohistochemical
detection of epithelial mesenchymal transition associated with
stemness phenotype in anaplastic thyroid carcinoma. Int J Clin Exp
Pathol. 3:755–762. 2010.PubMed/NCBI
|
|
67
|
Gottesman MM, Fojo T and Bates SE:
Multidrug resistance in cancer: role of ATP-dependent transporters.
Nat Rev Cancer. 2:48–58. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
68
|
Dean M, Fojo T and Bates S: Tumour stem
cells and drug resistance. Nat Rev Cancer. 5:275–284. 2005.
View Article : Google Scholar
|
|
69
|
Hirschmann-Jax C, Foster A, Wulf G, et al:
A distinct ‘side population’ of cells with high drug efflux
capacity in human tumor cells. Proc Natl Acad Sci USA.
101:14228–14233. 2004.
|
|
70
|
Feng F, Wang H, Fu H, et al:
Dedifferentiation of differentiated thyroid carcinoma cell line
FTC-133 is enhanced by 131I pretreatment. Nucl Med Biol.
38:1053–1058. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Robbins RJ and Schlumberger MJ: The
evolving role of 131I for the treatment of
differentiated thyroid carcinoma. J Nucl Med. 46:28S–37S. 2005.
|
|
72
|
Pajonk F, Vlashi E and McBride WH:
Radiation resistance of cancer stem cells: the 4 R’s of
radiobiology revisited. Stem Cells. 28:639–648. 2010.
|
|
73
|
Milas L and Hittelman WN: Cancer stem
cells and tumor response to therapy: current problems and future
prospects. Semin Radiat Oncol. 19:96–105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Takakura N: Formation and regulation of
the cancer stem cell niche. Cancer Sci. 103:1177–1181. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Nie D: Cancer stem cell and niche. Front
Biosci. 2:184–193. 2010. View
Article : Google Scholar
|