Introduction
Endometrial cancer (EC) is one of the most common
gynecological malignancies, with an escalating number of new cases
and an increasing mortality rate. In the US, ~49,500 new cases of
EC will be diagnosed in 2013, and 8,200 deaths are expected
(1). Accurate regulation of complex
transcriptional programs is central to the progression of EC.
Forkhead-box A1 (FOXA1) a member of the FOX family of transcription
factors (TFs) that comprises at least 40 members (2), was originally identified for its
transcriptional role in early liver and pancreas development
(3). At present, FOXA1 is mainly
known as a ‘pioneer factor’, which can occupy distal regulatory
enhancers and alter the chromatin accessibility for subsequent
recruitment of collaborating TFs, instead of directly promoting
transcription activation (4,5).
However, the functional role of FOXA1 in EC remains unclear.
Estrogen receptor α (ERα) is a ligand-activated
transcription factor that belongs to the nuclear receptor
superfamily (6), and regulates
estrogenic action in the female reproductive tract. Upon estrogen
stimulation, ERα binds at numerous genomic loci to promote the
transcriptional program (7), while
several coregulators are simultaneously recruited to the
chromosomal loops to facilitate the estrogenic transcriptional
response (8–11). Thus, ERα is an ideal target for
etiology-specific therapy in EC. Decreased or absent expression of
ERα is regarded to be accompanied by disease progression, but the
underlying mechanism is still unclear (12).
A number of studies have reported FOXA1 upregulation
in several types of human cancers, including breast cancer, in
which the expression patterns of FOXA1 are strikingly similar to
that of ERα (13). We speculated
that in endometrial cancer, a disease also closely related to
hormone and hormone receptors, FOXA1 may play a potential role in
cancer progression and may regulate the transcriptional activation
of ERα. In the present study, we investigated the level of FOXA1 in
multiple human EC cell lines and clinical samples. The expression
of FOXA1 in normal endometrial and EC tissues was analyzed and
correlated with clinicopathological parameters. The regulatory
roles of FOXA1 were investigated both in vitro and in
vivo. These experiments suggest that FOXA1 is a tumor
suppressor in EC. Furthermore, our results increased our
understanding of the ERα/FOXA1 relationship, and we propose that
FOXA1 may act as a regulator of the ER signaling pathway in EC cell
lines.
Materials and methods
Tissue selection and patient
information
Paraffin sections from 74 EC tissues were obtained
from patients who underwent initial hysterectomy at the
International Peace Maternity and Child Health Hospital of the
China Welfare Institute, which is affiliated to Shanghai Jiao Tong
University School of Medicine, from December 2009 to November 2012.
The stages and histological grades of these tumors were established
according to the criteria of the International Federation of
Gynecology and Obstetrics (FIGO) (14). Sixty-four normal endometrial samples
were obtained from patients who underwent hysterectomy to treat
other diseases such as adenomyosis or myoma. Twelve atypical
hyperplasia tissues were prepared from patients who underwent
hysteroscopic examination for the reason of irregular bleeding.
Following excision, tissue samples were immediately frozen in
liquid nitrogen and stored at −80°C until RNA extraction. Formalin
fixed paraffin-embedded (FFPE) tissues, matching the frozen cases,
were retrieved for IHC analysis. Pathological diagnoses of
endometrial lesions were carried out by two gynecologic
pathologists based on World Health Organization
classifications.
This study was approved by the Human Investigation
Ethical Committee of International Peace Maternity and Child
Hospital Affiliated Shanghai Jiao Tong University. The samples of
normal endometrial tissues, endometrial carcinoma and breast cancer
were collected after written informed consent from the
patients.
Immunohistochemical analysis
All samples were prepared and analyzed with the
Histostain-Plus kit (rabbit) (MRBiotech, Emeryville, CA, USA)
according to the manufacturer’s protocol. After dewaxing and
hydration, 4-μm sections from FFPE tissue were treated by boiling
in sodium citrate (pH 6.0) for 20 min for antigen retrieval, and
endogenous peroxidase activity was blocked by incubation in 0.3%
H2O2 in methanol for 10 min. Nonspecific
binding of antibodies was blocked with serum for 15 min. The slides
were then incubated with rabbit polyclonal anti-FOXA1 antibody
(1:200; Abcam, Cambridge, MA, USA) for 24 h at 4°C. The sections
were incubated with a biotinylated secondary antibody (MRBiotech).
Then the sections were treated by an horseradish
peroxidase-conjugated avidin-biotin complex. Following the
manufacturer’s instructions, samples were detected by DAB to ensure
the intensity of FOXA1 expression. We used the methods of Badve
et al to score the staining intensity (15). Briefly, the percentage of staining
was classified as ‘0’ when there was no nuclear expression, ‘1’ for
up to 10% positive tumor nuclei, ‘2’ for 11–20% and continuing in
the same manner until a maximum score of ‘10’. Intensity was scored
as ‘+,’ ‘++’, and ‘+++’ for weak, moderate, and strong staining,
respectively. The percentage (P) and intensity (I) of nuclear
expression were multiplied to generate a numerical score (S = P ×
I). As a positive control, a section of breast cancer tissue was
immunostained with anti-FOXA1 in the same manner. As a negative
control, phosphate-buffered saline (PBS) was used to replace the
primary antibody. ERα expression status was confirmed in the same
manner. Clinical and pathological data relating to the clinical
samples are presented in Table
I.
 | Table ICorrelation of FOXA1 expression with
clinicopathological parameters of the endometrial carcinoma
cases. |
Table I
Correlation of FOXA1 expression with
clinicopathological parameters of the endometrial carcinoma
cases.
| | FOXA1
histoscores | |
|---|
| |
| |
|---|
|
Characteristics | Case (n) | Means ± SD | p-value |
|---|
| Age (years) |
| ≤55 | 31 | 11.03±4.658 |
0.411b |
| >55 | 43 | 10.26±5.741 | |
| Histology |
| Endometrioid | 62 | 11.76±4.827 |
<0.001b |
| Non-endometrioid
(serous/clear) | 12 | 4.50±2.939 | |
| FIGO stage |
| Early (I–II) | 65 | 11.35±5.094 |
0.001b |
| Late (III–IV) | 9 | 5.00±2.872 | |
| Histological
grade |
| Grade 1 | 37 | 14.27±3.437 |
<0.0001a |
| Grade 2 | 18 | 8.67±4.498 | |
| Grade 3 | 19 | 5.21±2.974 | |
| Lymph node
metastasis |
| Positive | 7 | 4.43±2.992 |
0.002b |
| Negative | 67 | 11.22±5.075 | |
| Myometrial
invasion |
| ≤1/2 | 47 | 12.23±5.117 |
0.001b |
| >1/2 | 27 | 7.70±4.340 | |
| ER expression |
| Positive | 58 | 12.00±4.761 |
<0.0001b |
| Negative | 16 | 5.44±3.777 | |
Cell culture
Human endometrial cell lines (Ishikawa, RL95-2,
HEC-1B and AN3CA) were obtained from the Chinese Academy of
Sciences Committee Type Culture Collection cell bank. The cells
were grown in Dulbecco’s modified Eagle’s medium: Nutrient Mixture
F-12 (DMEM/F12) supplemented with 10% fetal bovine serum (FBS) and
1% penicillin/streptomycin (Gibco, Auckland, New Zealand) in a
humidified atmosphere of 5%CO2/95% air at 37°C.
RNA extraction and quantitative real-time
PCR
Total RNA was prepared from the EC cell lines using
TRIzol RNA isolation reagents (Invitrogen, Carlsbad, CA, USA). The
cDNA was generated with the Prime Script RT reagent kit (Takara
Inc., Otsu, Japan). A 50-μl PCR amplification of single-strand cDNA
was performed with 40 cycles of denaturation (94°C) for 60 sec,
annealing (55°C) for 30 sec, and elongation (72°C) for 30 sec using
SYBR Premix Ex Taq (Takara Inc.). The primer sequences are listed
in Table II. For all the data,
values on the y-axis employed the 2−ΔCt method. Data
were obtained in triplicate from three independent experiments.
| Table IIPrimer sequences for real-time PCR
analysis. |
Table II
Primer sequences for real-time PCR
analysis.
| mRNA | Primer
sequence |
|---|
| FOXA1 | Forward
5′-AGGTGTGTATTCCAGACCCG
Reverse 5′-TTGACGGTTTGGTTTGTGTG |
| ERα | Forward
5′-TGATTGGTCTCGTCTGGCG
Reverse 5′-CATGCCCTCTACACATTTTCCC |
| pS2 | Forward
5′-GTGTCACGCCCTCCCAGT
Reverse 5′-GGACCCCACGAACGGTG |
| GREB1 | Forward
5′-CAAAGAATAACCTGTTGGCCCTGC
Reverse 5′-GACATGCCTGCGCTCTCATACTTA |
| ACTB | Forward
5′-CAGCCATGTACGTTGCTATCCAGG
Reverse 5′-AGGTCCAGACGCAGGATGGCATG |
Western blot analysis
Cells were lysed for total protein extraction using
ProteoJET Mammalian Cell Lysis Reagent (MBI Fermentas, Ontario,
Canada) including a protease inhibitor cocktail (Roche Diagnostics,
Basel, Switzerland). Protein (80 μg) was loaded onto precast 4%
stacking, 10% Tris-glycine gels and separated by gel
electrophoresis. Proteins in the gels were then transferred to a
polyvinylidene difluoride (PVDF) membrane. After transfer,
membranes were blocked with 5% bovine serum albumin
(BSA)/phosphate-buffered saline (PBS) for 3 h. The membranes were
incubated with primary antibodies at 4°C overnight. The membranes
were then incubated with peroxidase-linked secondary antibody
(1:10,000; Epitomics, Burlingame, CA, USA) for 2 h at room
temperature. The blotted proteins were visualized using an ECL kit
(Beyotime, China), scanned and analyzed with TotalLab software.
Primary antibodies included: rabbit anti-FOXA1
(1:1000; Abcam), rabbit anti-ERα (1:2000; Epitomics) and rabbit
anti-β-actin (1:7500; Epitomics)
Plasmid and transfection
To stably express FOXA1, HEC-1B cells were washed
with PBS and switched to antibiotic-free growth medium for 24 h
before transfection. All transfections used Lipofectamine 2000
reagent (Invitrogen) according to the manufacturer’s instructions.
The plasmid pCMV6/GFP/Neo-FOXA1 (Genechem, Shanghai, China)
containing transfection-ready FOXA1 cDNA (GenBank: NM_004496) was
transfected into HEC-1B cells and then the cells were selected with
G418 (800 μg/ml; Gibco, Carlsbad, CA, USA) in the growth medium and
resistant clones were chosen.
To stably silence FOXA1, Ishikawa cells were
transfected with shFOXA1 (pGLV/h1/GFP/puro-FOXA1; Shanghai
Genepharma Ltd., China) (5′-GAGAGAAAAAAUCAAC AGC-3′) and were then
selected with puromycin (0.5 μg/ml; Sigma Chemical, St. Louis, MO,
USA). RL95-2 cells were transiently transfected with shFOXA1 in the
absence of selection pressure.
For the negative controls, HEC-1B cells were
transfected with a pure pCMV6/GFP/Neo vector, and Ishikawa and
RL95-2 cells were transfected with the pGLV/h1/GFP/puro vector,
respectively, under the same culture conditions.
To evaluate the effects of ERα expression and
silencing, Ishikawa and HEC-1B cells were transiently transfected
with ERα and shERα plasmids as described above, the ERα-expressing
vector (RG213277) and control vector (PS10010), the plasmid
encoding ERα silencing short hairpin RNA (shRNA) (sh-ERα,
GI378604), and the controls (sh-NC, TR3008) (all purchased from
OriGene Technologies, Beijing, China). The transfection efficiency
was verified by real-time quantitative reverse-transcription PCR
(qRT-PCR) and western blotting.
Cell migration assay
Cell migration activity was accomplished using
Boyden chambers containing polycarbonate filters with an 8-μm pore
size (Millipore). Cells (5×104 cells/well) were
resuspended with serum-free medium in the upper chamber. Medium
containing 10% fetal bovine serum was added to the lower chamber.
After incubation at 37°C for 24 h, cells on the upper side of the
membrane were removed using sterile cotton swabs. Cells adhering to
the lower surface were fixed with 100% methanol and stained with
hematoxylin, scanned and digital images were obtained with an
Aperio Scanscope System (Aperio Technologies, USA) at a
magnification of ×200. Five random fields were selected for each
membrane, and results are expressed in terms of the number of
migratory cells per field. Each experiment was conducted in
triplicate and repeated at least three times.
Cell invasion assay
Cell invasive activity was assessed using a BD
BioCoat Matrigel Invasion Chamber (BD Biosciences, USA) according
to the manufacturer’s instructions. Cells (1×105
cells/well) were resuspended with serum-free medium in the upper
chamber with a thin layer of Matrigel matrix. Medium containing 10%
FBS was added to the lower chamber. After incubation at 37°C for 72
h, cells that had migrated through the membrane were fixed, stained
and counted as described above.
Proliferation assay
Cells (3×103 cells/well) were plated in
96-well plates. Cell number was measured every 24 h via a
colorimetric assay with 3-(4,5-dimethylthiazol-2-yl)-2,
5-diphenyltetrazolium bromide (MTT; Sigma). Absorbance at 490 nm
was evaluated with a microplate reader (Model 680, Bio-Rad, USA).
Medium was changed every other day. For the plate colony formation
assay, 800 cells/well were seeded into 6-well plates. The cells
were fixed and stained with crystal violet when clearly
identifiable cell clones had formed. Results were detected under a
light microscope. Each experiment was repeated at least three
times, and assessed in triplicate.
Co-immunoprecipitation of FOXA1 and
ERα
Nuclear protein lysates were prepared at a
concentration of 1 μg/μl from Ishikawa and RL95-2 EC cell lines.
These lysates were incubated with 10 μg anti-FOXA1 (Abcam). The
immunoprecipitated proteins were collected using protein A/protein
G agarose beads (Beyotime), washed with PBS and resuspended in
loading buffer. The boiled samples were separated by SDS-PAGE,
transferred to a PVDF membrane and probed with anti-ERα
(Epitomics). Results were detected using an ECL kit (Beyotime),
scanned, and analyzed with TotalLab software.
Tumorigenicity assays in nude mice
All experimental protocols were approved by the
Ethics Committee for Animal Experimentation of Shanghai Jiao Tong
University. Female BALB/C athymic nude mice 4–6 weeks old were
divided into groups of six mice per group and housed with free
access to food and water. To verify the effect of the silencing of
FOXA1 on EC cells in vivo, we subcutaneously injected
1×107 Ishikawa cells suspended in 200 μl 1X PBS into the
unilateral foreleg of nude mice. Tumor measurement began 1 week
after injection and was conducted weekly using digital calipers.
Tumor volume (mm3) was calculated using the following
standard formula: Tumor volume (mm3) = (the longest
diameter) × (the shortest diameter)2 × 0.5. Mice were
sacrificed 5 weeks post-injection, and tumors were carefully
removed and their volumes and weights were determined prior to
further histological evaluation.
Statistics
Each experiment was completed at least three times,
and all tests were carried out with Statistical Package for the
Social Science (SPSS) software version 17.0 (Chicago, IL, USA).
Data represent the mean with standard deviation (SD). Data were
compared using the two-tailed Student’s t-test or Mann-Whitney
U-test for multiple comparisons. Differences having a probability
of p<0.05 were regarded as statistically significant.
Results
Expression of FOXA1 in tissues and its
association with clinicopathological parameters
To assess whether FOXA1 is commonly upregulated in
tissues, we compared its level in normal endometrial samples,
atypical complex hyperplasia, and EC tissues using
immunohistochemistry. Breast cancer tissues were immunostained as a
positive control. The results showed that FOXA1 expression was
restricted to the nucleus with little or no cytoplasmic staining
(Fig. 1A). We found strong staining
in normal endometrium. In contrast, staining was moderate or weak
in atypical complex hyperplasia and EC tissues.
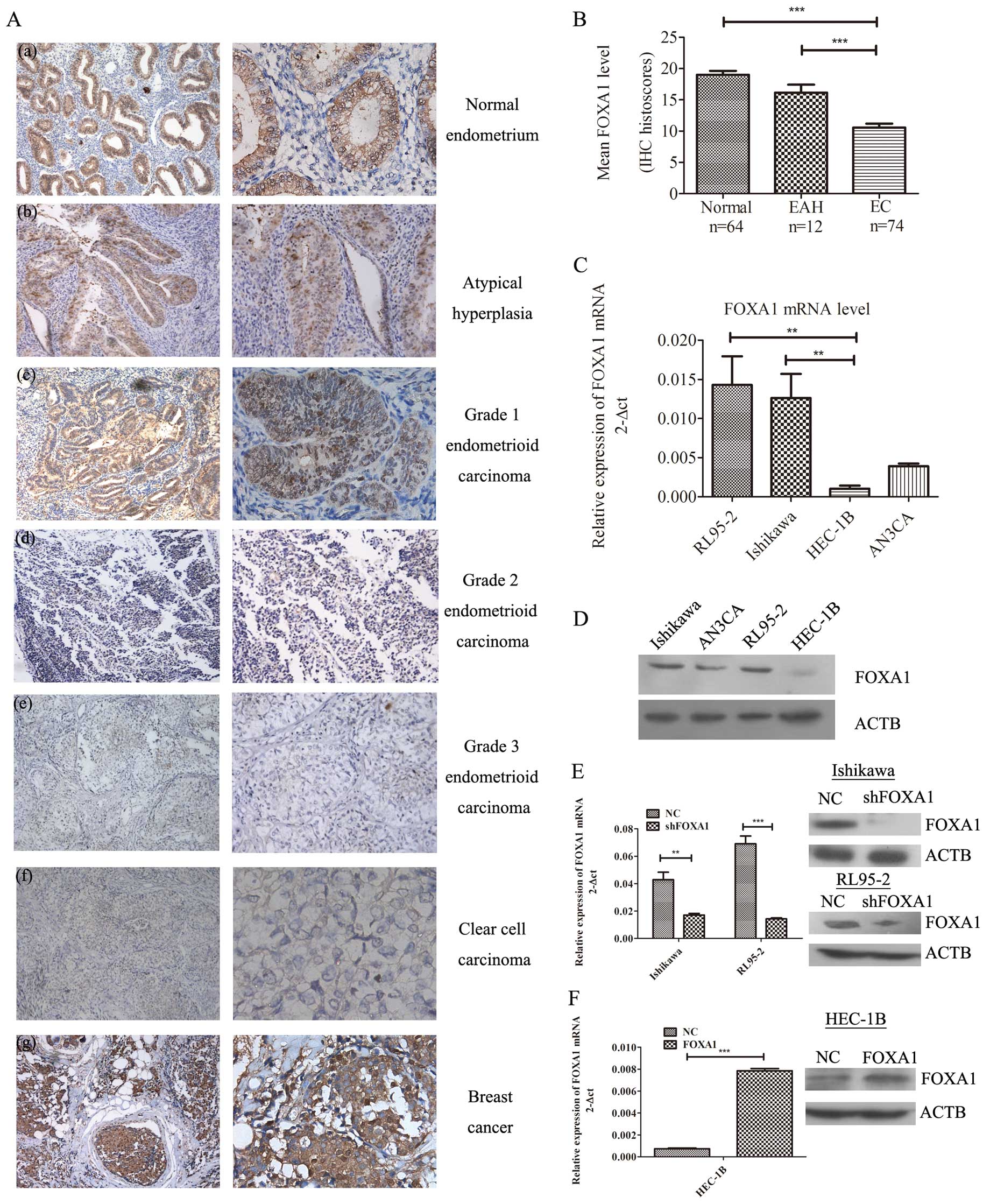 | Figure 1Overexpression of FOXA1 in normal
endometrium, endometrial cancer (EC) tissues and EC cell lines (A)
FOXA1 immunohistochemical staining in (a) normal endometrium with
strong staining, (b) atypical hyperplasia with moderate staining,
(c) grade 1 endometrioid endometrial carcinoma with moderate
staining, (d) grade 2 endometrioid endometrial carcinoma with weak
staining, (e) grade 3 endometrioid endometrial carcinoma with weak
staining, (f) clear cell carcinoma with no staining and (g) breast
cancer with strong staining. Original magnification ×100 (left) and
×200 (right). (B) Statistical summary of the immunostaining scores
in normal endometrium, endometrial atypical hyperplasia (EAH) and
endometrial cancer (EC), Statistics (U-test):
***p<0.001. (C) mRNA and (D) protein expression of
FOXA1 in EC cell lines as determined by qRT-PCR
(**p<0.01) and western blot analysis, respectively.
(E) mRNA and protein expression of FOXA1 were assessed in Ishikawa
and RL95-2 cells transiently transfected with shRNA against FOXA1
and its corresponding negative control. FOXA1 was downregulated
after transfection. (F) mRNA and protein expression of FOXA1 was
assessed in FOXA1 stable overexpressing HEC-1B cells. FOXA1 level
was altered 10-fold when compared to the negative control. Data for
each bar reflect triplicate measurements in each of three
independent experiments. Means (bars) and SD (error bars) are
shown. **p<0.01, ***p<0.001. |
To investigate whether the change in FOXA1
expression of EC was associated with any of the clinical
characteristics, we compared the association of FOXA1 expression
levels with the clinicopathological parameters of the EC cases
(Table I). We calculated a
composite histoscore to account for both staining intensity and
uniformity. Compared to the atypical hyperplasia, and EC tissues,
normal endometrial tissues expressed the highest levels of FOXA1
(Fig. 1B; H-score=19.02;
p<0.001). In EC tissues, strong staining of FOXA1 was observed
in early-stage EC (Fig. 1A;
H-score=11.35; p=0.001), whereas advanced-stage cancers had a
weaker FOXA1 expression (Fig. 1A;
H-score=5.00; p=0.001). FOXA1 expression was also correlated with
histologic type (p<0.001), with serous tumors (H-score=4.50)
showing lower FOXA1 scores when compared with the endometrial
endometrioid carcinomas (H-score=11.76). Furthermore, the status of
FOXA1 was positively associated with ER levels, suggesting that
FOXA1 expression has prognostic significance within the context of
ER expression (p<0.0001). All of these results indicate that
FOXA1 is a tumor suppressor in EC, and its expression level is a
favorable prognostic and diagnostic marker.
FOXA1 expression in human EC cell
lines
To confirm the expression of FOXA1 in EC, 4 human EC
cell lines (Ishikawa, RL95-2, HEC-1B and AN3CA) were used. Analysis
of FOXA1 expression by qRT-PCR (Fig.
1C) and western blotting (Fig.
1D) revealed that there was a high endogenous FOXA1 expression
in Ishikawa and RL95-2 cells, which are derived from
well-differentiated local endometrial adenocarcinoma (16,17).
In contrast, HEC-1B cells, which are derived from moderately
well-differentiated endometrial adenocarcinoma (18) had relatively low FOXA1 expression
among the cancer cell lines (p=0.0048).
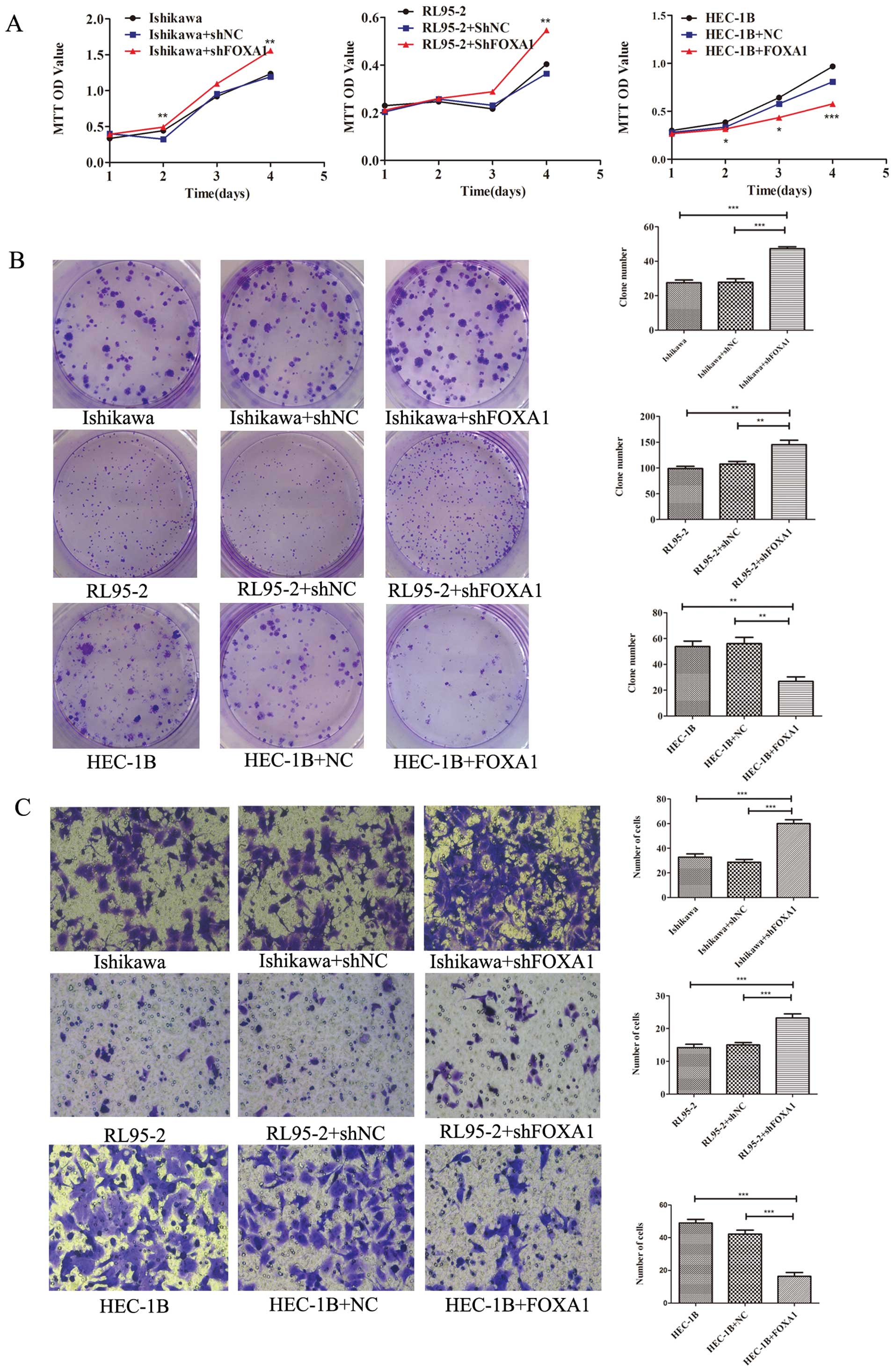
Suppression of EC cell proliferation,
migration and invasion by FOXA1
To determine whether loss of FOXA1 has an effect on
EC cells, Ishikawa and RL95-2 cells were transiently transfected
with a vector encoding a short hairpin RNA (shRNA) against FOXA1
and its corresponding negative control. After 48 h, RNA was
extracted and analyzed by qRT-PCR, and the protein expression was
examined after 72 h. As shown in Fig.
1E, the inhibitory efficiency of FOXA1 was ~50–70% in Ishikawa
and RL95-2 cells (p<0.01 and p<0.001, respectively).
We used the MTT assay to determine whether depletion
of FOXA1 induced cell proliferation; a statistically significant
induction of cell proliferation was detected in two EC cell lines
(Fig. 2A). The results revealed
that FOXA1 depletion promoted cell proliferation in Ishikawa and
RL95-2 cells in a time-dependent manner. To further explore the
role of FOXA1 in cell growth, we carried out plate colony formation
assays. Ishikawa and RL95-2 cells formed more colonies when
transfected with shFOXA1 than with the control vector (p<0.01;
Fig. 2B). Additionally, Transwell
migration (data not shown)and invasion assays (Fig. 2C) were performed to investigate the
migratory and invasive potential of EC cells. FOXA1 depletion
markedly promoted cell migration and invasion in Ishikawa and
RL95-2 cells (p<0.001).
In comparison with the effect of FOXA1 knockdown,
restoring FOXA1 expression in HEC-1B cells had an opposite effect.
Successful reestablishment of FOXA1 expression in HEC-1B cells was
confirmed by qRT-PCR and western blotting (p<0.001; Fig. 1F). After 72 h of incubation, HEC-1B
cells stably transfected with the FOXA1 expression plasmid showed
significant growth inhibition when compared with the negative
control cells (p<0.01; Fig. 2A and
B). Moreover, FOXA1 plasmid transfection decreased the
migratory and invasive potential of HEC-1B cells (p<0.001,
Fig. 2C). All of the in
vitro experiments suggest that FOXA1 plays an essential role in
inhibiting EC progression.
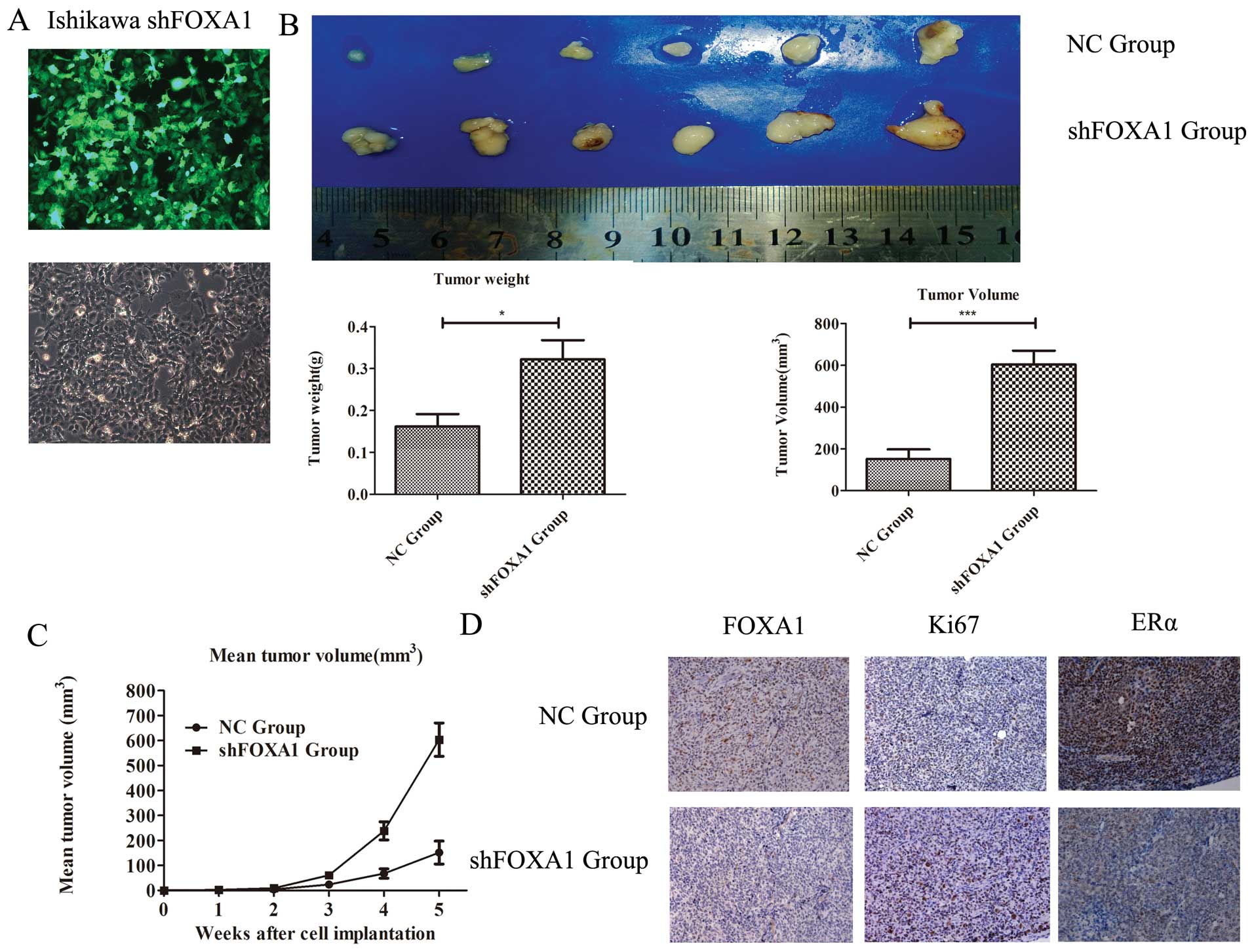
Oncogenic role of FOXA1 in an in vivo
tumor xenograft model
To further assess the role of FOXA1 in the
progression of EC, we performed tumorigenicity assays in nude mice.
We used shRNA-mediated stable knockdown of FOXA1 in Ishikawa cells
(Fig. 3A), and the cells were
injected subcutaneously. Tumors formed 1 week after injection. Over
a 5-week period, we observed higher tumor growth rates in the mice
injected with the cells transfected with shFOXA1 compared with the
tumor growth in the negative control group (p<0.01, Fig. 3C). Five weeks after injection,
tumors were completely removed from the mice. The final mean weight
and volume of the tumors were markedly higher in the shFOXA1 group
than in the negative control group (p<0.05 and p<0.001;
Fig. 3B). To explore whether
shFOXA1 influences the proliferative capacity of Ishikawa cells, we
compared the levels of Ki-67 in the two groups. Previous studies
revealed that expression of Ki-67 is associated with cell
proliferation (19). As shown in
Fig. 3D, Ki-67 was upregulated in
the shFOXA1 group (p<0.01). Furthermore, ERα expression was
downregulated in the shFOXA1 group (p<0.01). Taken together,
these results suggest an important role for FOXA1 in regulating
tumor viability in EC.
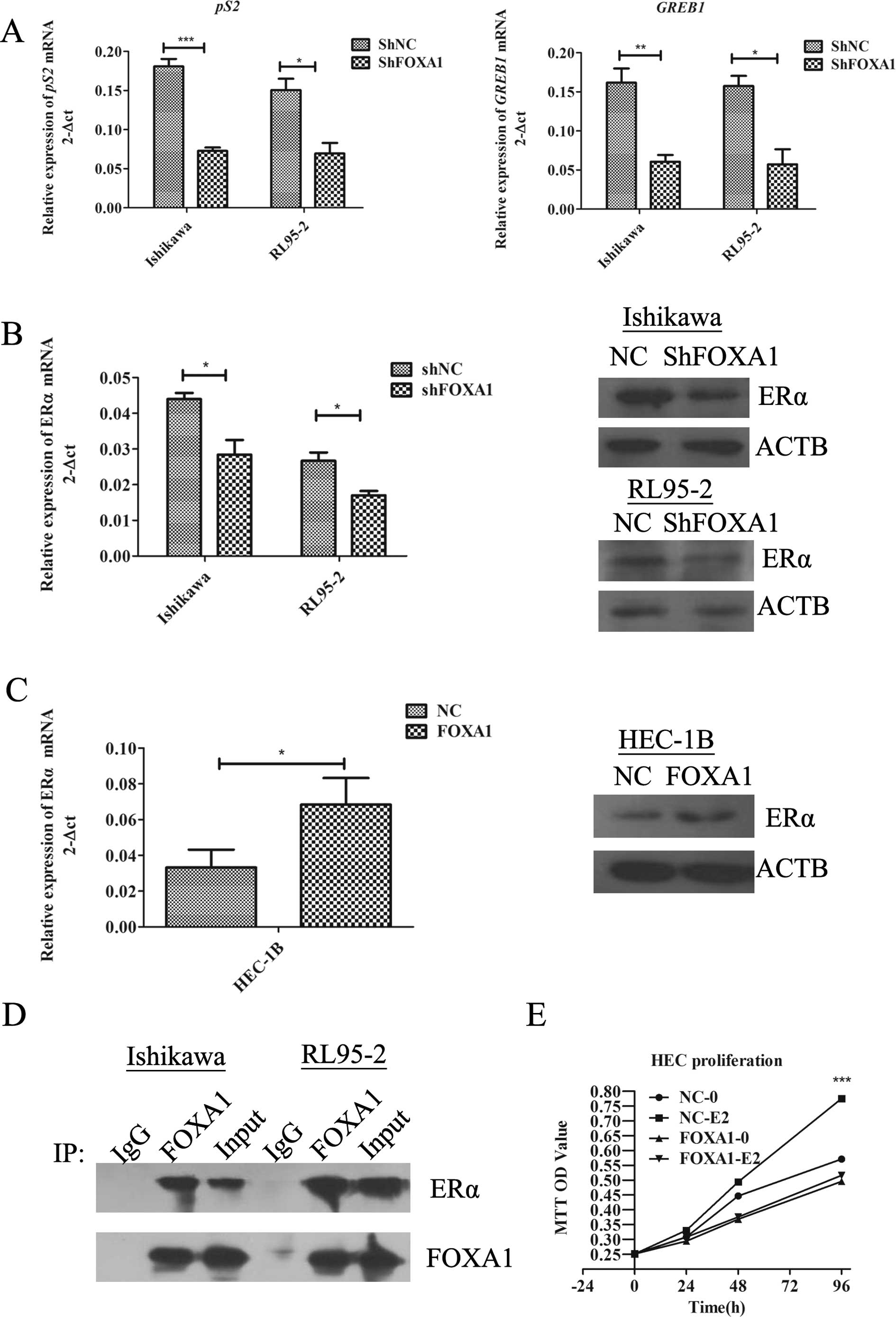
FOXA1 regulates the ER signaling
pathway
Consistent with several reports that describe FOXA1
as a regulator of ER activity (20), our experiments used Ishikawa and
RL95-2 cells as they express easily detectable levels of both FOXA1
and ERα (data not shown). FOXA1 knockdown by shRNA revealed a
significant decrease in the expression of ERα-dependent pS2
and GREB1 genes in the Ishikawa and RL95-2 cells in response
to 10 nM estradiol (Fig. 4A).
Additionally, we found that FOXA1 depletion abolished the
expression of ERα at the mRNA and protein levels (p<0.05,
Fig. 4B). We also examined whether
FOXA1 overexpression promotes ERα expression in
FOXA1-overexpressing HEC-1B cells, an EC cell line with relatively
low ERα expression. As shown in Fig.
4C, an increased FOXA1 level was correlated with higher ER mRNA
and protein levels (p<0.05). These results revealed that FOXA1
regulated endogenous ERα expression in the three EC cell lines at
the post-transcriptional level. To ascertain whether ERα influences
the FOXA1 level, we performed a reciprocal experiment. After
inhibition or upregulation of the expression of ERα, we found no
significant difference in FOXA1 expression (data not shown),
confirming that FOXA1 acts upstream of ERα.
We next sought to determine whether FOXA1 and ERα
interact with each other by exploring endogenous FOXA1 and ERα
immunoprecipitates from Ishikawa cells with the reciprocal
antibodies. As shown in Fig. 4D, we
found an interaction between the two proteins, which demonstrated
that there was a cell-specific correlation between these two genes,
supporting an involvement of FOXA1 in the ER signaling pathway.
To further assess the role of FOXA1 in the
estrogen-induced growth in EC cells, we transfected HEC-1B cells
with a FOXA1 expression plasmid, and the proliferation assay was
completed in phenol red-free medium containing 10%
charcoal/dextran-treated FBS. The results showed that the EC cells
expressing FOXA1 no longer proliferated following estrogen
stimulation (p<0.001, Fig. 4E),
suggesting a functional role for FOXA1 in mediating the response to
estrogen in EC.
Discussion
Endometrial cancer involves a heterogeneous group of
tumors. Early-stage type I endometrioid tumors are often treated
using adjuvant radiotherapy (21),
whereas similarly staged type II serous tumors are treated with
chemotherapy. Therefore, an accurate classification determines the
appropriate adjuvant therapy. However, high-grade serous and
endometrioid endometrial carcinomas are difficult to accurately
subtype, and intraobserver concordance among specialized
pathologists is low (22–24). Thus, the molecular classification of
endometrial carcinomas may be a novel method by which to carry out
informed treatment decisions.
The FOX family, particularly FOXA1, plays an
important role in organ morphogenesis and disease progression
(25). However, the role of FOXA1
expression or activity in EC is still unclear. A previous study
investigated the expression pattern of FOXA1 in EC tissues, yet the
samples of normal endometrium and the precise mechanism remain to
be further explored (26). In the
present study, we found that FOXA1 was present in almost all normal
endometrial tissues, whereas differential expression was observed
among the EC tumor types. We confirmed that FOXA1 can be detected
in early-satge endometrial endometrioid carcinomas more frequently
than in advanced-stage cancers, including endometrioid carcinomas
and serous tumors. Furthermore, the FOXA1 level was correlated with
the depth of myometrial invasion and lymph node metastasis of EC,
which are important prognostic factors (27). More importantly, we found a
significantly positive correlation between FOXA1 and ERα in EC
tissues, and the promotive effect of FOXA1 on ERα was proven in EC
cell lines. This expression analysis revealed that FOXA1 was more
highly expressed in well-differentiated tumors, which can provide a
clinically effective prognostic factor particularly in low-risk EC
patients, among ERα-positive cases.
Over the past decade, FOXA1 expression has been
reported in several types of human cancers, and the results suggest
it has a dual function. Briefly, the FOXA1 level is increased in
acute myeloid leukemia, esophageal cancer and lung cancer, and acts
as an oncogene (28,29). In contrast, in human hepatocellular
carcinoma, bladder cancer, pancreatic and luminal subtype A breast
cancer, FOXA1 inhibits metastasis and is a favorable prognostic
factor (30–33). In the present study, we confirmed
that FOXA1 overexpression in HEC-1B cells decreased cellular
invasion and migration, and completely prevented estrogen-induced
proliferation, whereas its depletion in Ishikawa and RL95-2 cells
promoted cell viability and was associated with tumorigenicity.
FOXA1 was first reported to act as a growth inhibitor through
activation of the transcription of p27 in breast cancer (34). Subsequent studies revealed that
FOXA1 is associated with cyclin-dependent kinase inhibitors
(13,35), which may explain the
anti-proliferative capacity of FOXA1 in EC. In addition, our
xenograft experiments showed that ablation of FOXA1 expression
promoted the proliferative ability of Ishikawa cell. For several
diseases, loss of FOXA1 expression promotes tumorigenesis or
increases the level of tumor aggressiveness (32). All of these findings were confirmed
in our study and suggest that FOXA1 acts as a tumor suppressor in
EC.
An increasing number of studies suggest that
crosstalk between ER and signal transduction pathways is a
potential factor in the proliferation of ERα-positive EC cells and
endocrine resistance (36,37). It has been well established that
FOXA1 binding parallels ERα binding in both accessible and
inaccessible regions, revealing that FOXA1 is a key determinant of
ER function and endocrine response in breast cancer (20). Critically, one important issue
raised by our experiments is that the expression pattern of FOXA1
in EC are strikingly similar to that of ERα. Hence, we speculated
that decreased ERα expression (or its absence) during disease
progression may be due to loss of FOXA1 function. FOXA1 depletion
in vitro significantly decreased pS2 and GREB1
expression, which indicates that FOXA1 is a key regulator of ERα
gene transcription. Furthermore, specific silencing of FOXA1 was
found to lead to the discrepancy of ERα mRNA and protein levels in
the multiple EC cell lines tested. To further understand the role
of FOXA1 in the ERα signaling pathway of EC, co-immunoprecipitation
was performed and we found an interaction between the two proteins.
The comparable effects of FOXA1 and ERα suggest that the two
proteins may cooperatively regulate the transcriptional networks
previously ascribed to ERα alone. These findings are consistent
with previous studies concerning breast cancer and confirm the
involvement of FOXA1 in the ERα signaling pathway.
A number of studies have reported that FOXA1 acts as
a ‘pioneer factor’. It can mimic a linker histone and can bind
directly to condensed chromatin (38,39).
This raises the possibility that FOXA1 may provide the opportunity
for other TFs to associate with chromatin. Several lines of
evidence have provided an indication of the mechanistic complexity
required for acute transcriptional control of steroid hormone
receptors (8,40). Of note, imbalanced activity of
several co-regulator proteins has been proven to cause endocrine
therapy-resistance, suggesting that future therapies for
hormone-related diseases may target additional components of the
steroid hormone receptors (41).
Although our present understanding of whether FOXA1 is involved in
ERα-regulated gene transcription in endometrial cancer is limited,
emerging evidence implicates an important role for FOXA1 in disease
therapy (42). However, there are
many TFs which may regulate the progression of endometrial cancer;
therefore our team is investigating whether FOXA1 affects the
function of other steroid receptors, including progesterone
receptor, androgen receptor and glucocorticoid receptor, which may
establish a foundation for future application of FOXA1 in the
treatment of EC.
In summary, our findings confirm that FOXA1 is a
valuable prognostic factor in EC, and a positive relationship
exists between FOXA1 and ERα. Further reasearch concerning FOXA1 in
the steroid receptor signaling pathway for the treatment of EC is
warranted.
Acknowledgements
We thank Qin Huang, Yifei Chen, and Fangyuan Wang
(Centre of Research Laboratory, International Peace Maternity and
Child Health Hospital Affiliated to Shanghai Jiao Tong University
School of Medicine, Shanghai, China) for assisting in the
preparation of this manuscript. This study was supported by the
National Natural Science Foundation of China (nos. 81272885,
81172476, 81072139), the Foundation Project of Shanghai Municipal
Science and Technology Commission (no. 13JC1404500) and the Ph.D.
Programs Foundation of the Ministry of Education of China (no.
20120073110090).
Abbreviations:
|
EC
|
endometrial cancer
|
|
FOXA1
|
Forkhead-box A1
|
|
ERα
|
estrogen receptor α
|
|
TFs
|
transcription factors
|
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar
|
|
2
|
Hannenhalli S and Kaestner KH: The
evolution of Fox genes and their role in development and disease.
Nat Rev Genet. 10:233–240. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Costa RH, Grayson DR and Darnell JE Jr:
Multiple hepatocyte-enriched nuclear factors function in the
regulation of transthyretin and alpha 1-antitrypsin genes. Mol Cell
Biol. 9:1415–1425. 1989.PubMed/NCBI
|
|
4
|
Carroll JS, Liu XS, Brodsky AS, et al:
Chromosome-wide mapping of estrogen receptor binding reveals
long-range regulation requiring the forkhead protein FoxA1. Cell.
122:33–43. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang Q, Li W, Zhang Y, et al: Androgen
receptor regulates a distinct transcription program in
androgen-independent prostate cancer. Cell. 138:245–256. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Evans RM: The steroid and thyroid hormone
receptor superfamily. Science. 240:889–895. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Vilquin P, Villedieu M, Grisard E, et al:
Molecular characterization of anastrozole resistance in breast
cancer: pivotal role of the Akt/mTOR pathway in the emergence of de
novo or acquired resistance and importance of combining the
allosteric Akt inhibitor MK-2206 with an aromatase inhibitor. Int J
Cancer. 133:1589–1602. 2013. View Article : Google Scholar
|
|
8
|
Holmes KA, Hurtado A, Brown GD, et al:
Transducin-like enhancer protein 1 mediates estrogen receptor
binding and transcriptional activity in breast cancer cells. Proc
Natl Acad Sci USA. 109:2748–2753. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gaughan L, Stockley J, Coffey K, et al:
KDM4B is a master regulator of the estrogen receptor signalling
cascade. Nucleic Acids Res. 41:6892–6904. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rahimi M, George J and Tang C: EGFR
variant-mediated invasion by enhanced CXCR4 expression through
transcriptional and post-translational mechanisms. Int J Cancer.
126:1850–1860. 2010.PubMed/NCBI
|
|
11
|
Theodorou V, Stark R, Menon S and Carrol
JS: GATA3 acts upstream of FOXA1 in mediating ESR1 binding by
shaping enhancer accessibility. Genome Res. 23:12–22. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Srijaipracharoen S, Tangjitgamol S,
Tanvanich S, et al: Expression of ER, PR, and Her-2/neu in
endometrial cancer: a clinicopathological study. Asian Pac J Cancer
Prev. 11:215–220. 2010.PubMed/NCBI
|
|
13
|
Wolf I, Bose S, Williamson EA, et al:
FOXA1: Growth inhibitor and a favorable prognostic factor in human
breast cancer. Int J Cancer. 120:1013–1022. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Creasman W: Revised FIGO staging for
carcinoma of the endometrium. Int J Gynaecol Obstet. 105:1092009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Badve S, Turbin D, Thorat MA, et al: FOXA1
expression in breast cancer - correlation with luminal subtype A
and survival. Clin Cancer Res. 13:4415–4421. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nishida M: The Ishikawa cells from birth
to the present. Hum Cell. 15:104–117. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Way DL, Grosso DS, Davis JR, et al:
Characterization of a new human endometrial carcinoma (RL95-2)
established in tissue culture. In Vitro. 19:147–158. 1983.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kuramoto H: Studies of the growth and
cytogenetic properties of human endometrial adenocarcinoma in
culture and its development into an established line. Acta Obstet
Gynaecol Jpn. 19:47–58. 1972.PubMed/NCBI
|
|
19
|
Kawase M, Toyama T, Takahashi S, et al:
FOXA1 expression after neoadjuvant chemotherapy is a prognostic
marker in estrogen receptor-positive breast cancer. Breast Cancer.
Jun 16–2013.(Epub ahead of print).
|
|
20
|
Hurtado A, Holmes KA, Ross-Innes CS, et
al: FOXA1 is a key determinant of estrogen receptor function and
endocrine response. Nat Genet. 43:27–33. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bokhman JV: Two pathogenetic types of
endometrial carcinoma. Gynecol Oncol. 15:10–17. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
McConechy MK, Ding J, Cheang MC, et al:
Use of mutation profiles to refine the classification of
endometrial carcinomas. J Pathol. 228:20–30. 2012.PubMed/NCBI
|
|
23
|
Alvarez T, Miller E, Duska L and Oliva E:
Molecular profile of grade 3 endometrioid endometrial carcinoma: is
it a type I or type II endometrial carcinoma? Am J Surg Pathol.
36:753–761. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Clarke BA and Gilks CB: Endometrial
carcinoma: controversies in histopathological assessment of grade
and tumour cell type. J Clin Pathol. 63:410–415. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bernardo GM and Keri RA: FOXA1: a
transcription factor with parallel functions in development and
cancer. Biosci Rep. 32:113–130. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Abe Y, Ijichi N, Ikeda K, et al: Forkhead
box transcription factor, forkhead box A1, shows negative
association with lymph node status in endometrial cancer, and
represses cell proliferation and migration of endometrial cancer
cells. Cancer Sci. 103:806–812. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Uharcek P: Prognostic factors in
endometrial carcinoma. J Obstet Gynaecol Res. 34:776–783. 2008.
View Article : Google Scholar
|
|
28
|
Neben K, Schnittger S, Brors B, et al:
Distinct gene expression patterns associated with FLT3- and
NRAS-activating mutations in acute myeloid leukemia with normal
karyotype. Oncogene. 24:1580–1588. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lin L, Miller CT, Contreras JI, et al: The
hepatocyte nuclear factor 3 alpha gene, HNF3alpha (FOXA1), on
chromosome band 14q13 is amplified and overexpressed in esophageal
and lung adenocarcinomas. Cancer Res. 62:5273–5279. 2002.PubMed/NCBI
|
|
30
|
Coulouarn C, Factor VM, Andersen JB, et
al: Loss of miR-122 expression in liver cancer correlates with
suppression of the hepatic phenotype and gain of metastatic
properties. Oncogene. 28:3526–3536. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
DeGraff DJ, Clark PE, Cates JM, et al:
Loss of the urothelial differentiation marker FOXA1 is associated
with high grade, late stage bladder cancer and increased tumor
proliferation. PLoS One. 7:e366692012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Song Y, Washington MK and Crawford HC:
Loss of FOXA1/2 is essential for the epithelial-to-mesenchymal
transition in pancreatic cancer. Cancer Res. 70:2115–2125. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Early Breast Cancer Trialists’
Collaborative Group (EBCTCG). Effects of chemotherapy and hormonal
therapy for early breast cancer on recurrence and 15-year survival:
an overview of the randomised trials. Lancet. 365:1687–1717.
2005.PubMed/NCBI
|
|
34
|
Williamson EA, Wolf I, O’Kelly J, et al:
BRCA1 and FOXA1 proteins coregulate the expression of the cell
cycle-dependent kinase inhibitor p27(Kip1). Oncogene. 25:1391–1399.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li Q, Zhang Y, Fu J, et al: FOXA1 mediates
p16(INK4a) activation during cellular senescence. EMBO J.
32:858–873. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Prat J, Gallardo A, Cuatrecasas M and
Catasus L: Endometrial carcinoma: pathology and genetics.
Pathology. 39:72–87. 2007. View Article : Google Scholar
|
|
37
|
Gururaj AE, Rayala SK, Vadlamudi RK and
Kumar R: Novel mechanisms of resistance to endocrine therapy:
genomic and nongenomic considerations. Clin Cancer Res.
12:S1001–S1007. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cirillo LA, Lin FR, Cuesta I, et al:
Opening of compacted chromatin by early developmental transcription
factors HNF3 (FoxA) and GATA-4. Mol Cell. 9:279–289. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lupien M, Eeckhoute J, Meyer CA, et al:
FoxA1 translates epigenetic signatures into enhancer-driven
lineage-specific transcription. Cell. 132:958–970. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kong SL, Li G, Loh SL, et al: Cellular
reprogramming by the conjoint action of ERalpha, FOXA1, and GATA3
to a ligand-inducible growth state. Mol Syst Biol. 7:5262011.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Green KA and Carroll JS:
Oestrogen-receptor-mediated transcription and the influence of
co-factors and chromatin state. Nat Rev Cancer. 7:713–722. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ross-Innes CS, Stark R, Teschendorff AE,
et al: Differential oestrogen receptor binding is associated with
clinical outcome in breast cancer. Nature. 481:389–393.
2012.PubMed/NCBI
|