Introduction
Although osteosarcoma (OS) is a relatively rare
cancer, it is the most common type of primary malignant bone cancer
(1). OS mainly arises in
adolescents and young adults (between the ages of 10 to 25). Almost
6 per million adolescents and 2 per million adults will develop OS.
By using modern OS treatment strategies that combine surgery,
chemotherapy, sometimes radiotherapy, the 5-year survival rate for
OS patients is still only 60–70% (2). Thus, there is an urgent need to
explore new cancer management methods to treat OS.
Currently, gene therapy, as a novel cancer therapy,
is gaining increasing attention. This therapy works by transferring
a nucleic acid regulator, which affects expression of a specific
tumor-associated gene to cancer cells, such as short hairpin RNA
(shRNA) (3). shRNA expression
vector system has been established to generate small interfering
RNA (siRNA) in mammalian cells. siRNA plays many important roles in
the cellular regulatory mechanism, especially in the RNA
interference (RNAi) pathway, where siRNA utilises the cell’s
inherent machinery (Dicer) to degrade target mRNA and thereby
silencing specific gene expression (4). Lentivirus vector has been demonstrated
to be a highly effective and stable gene transfer tool (5). Therefore, lentivirus-delivered shRNA
is considered to be a hopeful tool for gene therapy, which has been
used in clinical trials without prominent side-effects (6).
Aurora kinase A (AURKA), a member of
serine/threonine kinase family, is involved in correct centrosome
duplication, mauration and separation, as well as in chromosomal
segregation during cellular mitosis (7). Overexpression of AURKA gene is evident
in various human cancers, including esophageal squamous cell
carcinoma, pancreatic, and ovarian cancer, and is implicated in
promoting cell proliferation and suppressing cell apoptosis
(8–11). Some studies also show that altered
activity of AURKA leads to genomic instability and thus to
tumorigenesis (12,13).
In the present study, we used lentivirus-mediated
shRNA to silence AURKA expression in human OS cells. Then we
examined the effect of AURKA silence on tumor growth and the
mechanisms.
Materials and methods
Cell culture
Human OS cell lines SAOS-2, U2OS, MG-63 and HOS U251
were purchased from the American Type Culture Collection (ATCC;
Manassas, VA, USA) and cultured in Dulbecco’s modified Eagle’s
medium (DMEM) containing 10% fetal bovine serum (FBS), 100 U/ml
penicillin and 100 μg/ml streptomycin at 37°C under humidified air
containing 5% CO2.
RNA extraction and quantitative real-time
RT-PCR
Total RNA was extracted from cells with TRIzol
reagent (Invitrogen, Carlsbad, CA, USA) according to the
manufacturer’s instructions. The cDNA was synthesized using the
RevertAid First Strand cDNA Synthesis kit (Fermentas, Vilnius,
Lithuania) according to the manufacturer’s protocol. Genes
expression were detected by quantitative real-time RT-PCR (qRT-PCR)
using the standard SYBR-Green RT-PCR kit (Takara, Kyoto, Japan) as
recommended by the manufacturer. The specific primer pairs and the
amplified products are as follows: AURKA (286 bp), sense: 5′-GCC
CTG TCT TAC TGT CAT TCG-3′; antisense: 5′-AGG TCT CTT GGT ATG TGT
TTG C-3′; GAPDH (121 bp), sense: 5′-TGA CTT CAA CAG CGA CAC CCA-3′;
antisense: 5′-CAC CCT GTT GCT GTA GCC AAA-3′. The relative levels
of AURKA gene mRNA transcripts were normalized to the control
GAPDH. Relative gene expression was quantified using the GraphPad
Prism 5.0 software (GraphPad Software, Inc., San Diego, CA,
USA).
AURKA gene silence mediated by
lentivirus-delivered shRNA
shRNA candidate target sequences to AURKA is
5′-GAAAG CTCCACATCAATAA-3′. The oligonucleotides encoding the shRNA
sequence or non-silencing control (NC) sequence
(5′-TTCTCCGAACGTGTCACGT-3′) were inserted into the GFP express
vector pGCL-GFP (Shanghai GeneChem Co., Ltd., Shanghai, China). NC
RNAi was used as a negative RNAi control. The recombinant virus was
packaged using the Lentivector Expression systems (Shanghai
GeneChem). SAOS-2 and U2OS cells were infected. After 3 days,
GFP-positive cells were counted under fluorescence microscope
(Olympus, Tokyo, Japan). AURKA expression after infection of shRNA
expression construct was revealed by real-time PCR on day 4.
Western blotting
The whole-cell lysates were subjected to SDS-PAGE.
The blots were incubated with AURKA antibodies (Cell Signaling
Technology, Beverly, MA, USA) and then with anti-rabbit IgG
peroxidase-conjugated secondary antibody (Santa Cruz Biotechnology,
Santa Cruz, CA, USA) and chemiluminescent substrates. Hybridization
with anti-GAPDH (Santa Cruz Biotechnology) was used to confirm
equal protein loading.
Colony-forming assay
Exponentially growing cells were suspended in
complete growth medium and seeded in 6-well plates with 200
cells/well. The plates were maintained at 37°C in a humidified
incubator with 5% CO2 for two weeks. The visible
colonies were subsequently recorded by a fluorescence microscope.
After fixation in paraformaldehyde, the colonies were stained with
Giemsa for 10 min followed by taking images with a digital
camera.
Tumorigenesis assay
Male nude mice (7–8 weeks old, weighing 18–20 g)
were obtained from the Shanghai Experimental Center, Chinese
Science Academy, Shanghai, and maintained at an animal facility
under pathogen-free conditions. All animals received humane care
according to the National Research Council’s guidelines. The study
was approved by the Animal Experimentation Committee of Soochow
University.
SAOS-2 cells infected with control lentivirus or
AURKA-specific siRNA lentivirus were resuspended in PBS at a final
density of 1×107 cells/ml. Each group of nude mice was
injected into the axilla with 100 μl of the corresponding cell
suspension (equivalent to 1×106 cells). The mice were
sacrificed 3 weeks after injection and examined for tumor growth.
The effect of AURKA knockdown on tumor growth was assessed by
measuring tumor size and tumor weight. Measurement of tumor size
was used to calculate the tumor volume using the formula: 1/2
*a*b2, where a is the major diameter and b is the minor
diameter.
Cell viability assay
Cell viability was evaluated by
3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT)
(Sigma Chemical, St. Louis, MO, USA) assay. The test cells in
exponential growth were plated at a final concentration of
2×103 cells/well in 96-well culture plates for different
culture time. Then MTT (10 μl, 5 mg/ml) was added to each well.
Incubation was continued at 37°C for 4 h. The reaction was
terminated by replacing the MTT-containing medium with 100 μl DMSO
and the formazan salts were dissolved by gentle shaking for about
10 min at room temperature. Optical density (OD) of each well was
measured at 490 nm using ELISA reader (ELx808; Bio-Tek Instruments,
Winooski, VT USA).
Detection of apoptosis by flow
cytometry
Cells were stained with APC labeled Annexin V
(eBioscience, San Diego, CA, USA) to detect apoptotic cells
(Annexin V positive). A total of 1.0×106 cells were
washed twice with ice-cold PBS and incubated for 15 min in 100 μl
staining buffer including 5 μl APC labeled Annexin V.
Fluorescence-activated cell sorting (FACS) analysis for Annexin V
staining was performed by flow cytometer (FACSCalibur; BD
Biosciences, San Jose, CA, USA). All experiments were performed in
triplicate.
Cell cycle assay
Cells were seeded into a 6-cm dish, harvested at
specific times by centrifuging at 1,200 rpm for 5 min. Following
washing with pre-cooled PBS (pH 7.4) twice, cells were fixed in 70%
alcohol. Percentage of cells in each stage of the cell cycle was
determined by staining with propidium iodide (PI) (Sigma). The
analysis of cell cycle distribution was performed by a flow
cytometer (FACSCalibur; BD Biosciences) in accordance with the
manufacturer’s guidelines.
Statistical methods
Data are expressed as the mean ± SEM. Statistical
significance was determined by the Student’s t-test and one-way
ANOVA using GraphPad Prism 5.0 software (GraphPad software). A
value of P<0.05 was considered to indicate a statistically
significant difference.
Results
Lentivirus-delivered shRNA mediated
downregulation of AURKA expression in OS cells
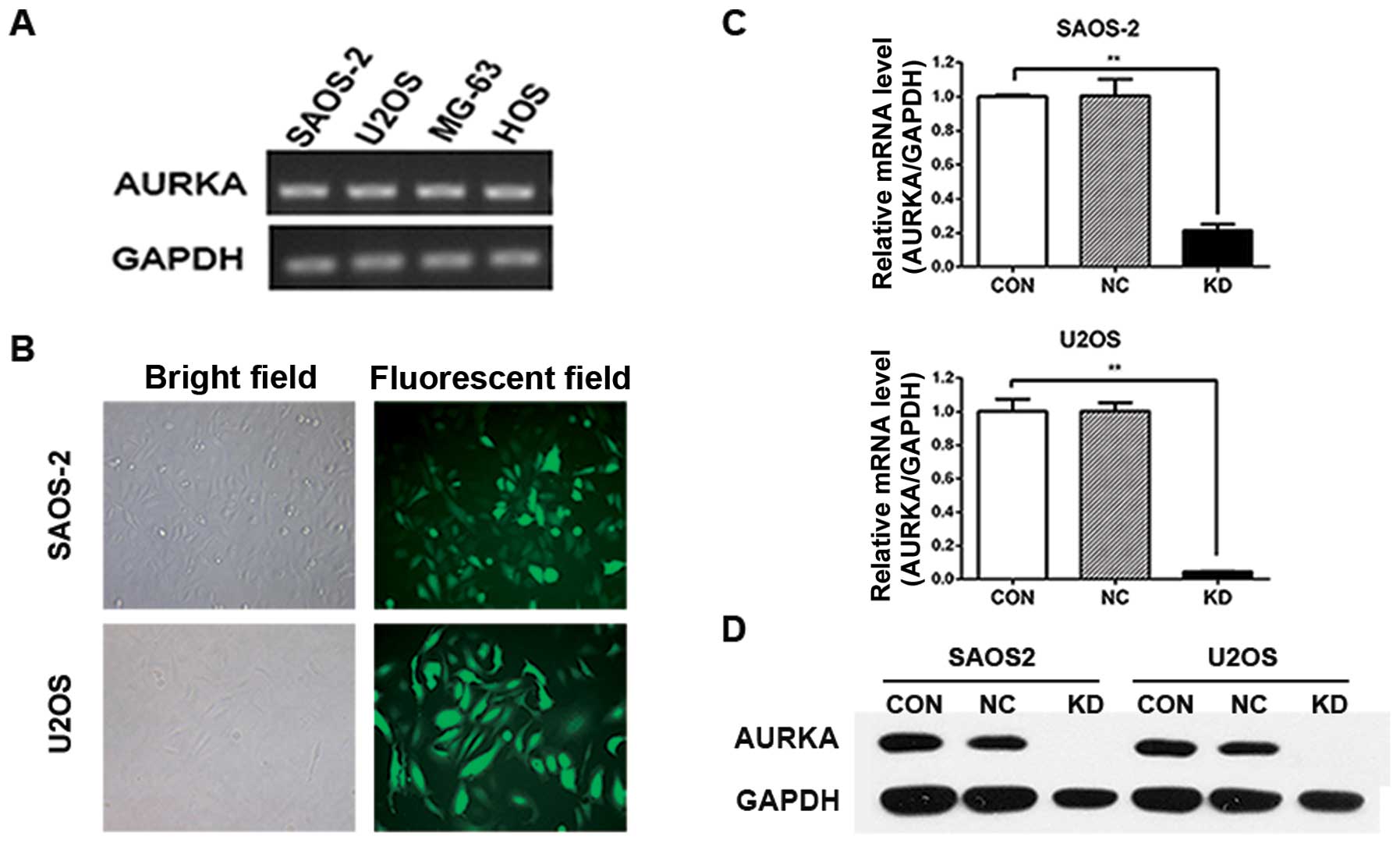
We used RT-PCR to detect AURKA expression in the OS
cell line SAOS-2, U2OS, MG-63 and HOS. The results showed that
AURKA gene is expressed in these human OS cell lines (Fig. 1A). To study the role of AURKA in OS
cells, we used recombinant lentivirus delivered shRNA to silence
AURKA expression in SAOS-2 and U2OS cells. The infection efficiency
of AURKA-shRNA lentivirus was >80% in both SAOS-2 and U2OS
cells. Most of the infected cells expressed GFP protein 3 days
after infection (Fig. 1B).
To evaluate the effectiveness of lentivirus-mediated
infection on the silencing of AURKA expression, lentivirus infected
OS cells were assayed by real-time PCR. The data demonstrated that
AURKA mRNA expression was efficiently suppressed in AURKA-shRNA
lentivirus infected cells (KD) compared to uninfected cells (CON).
There was no significant difference in AURKA mRNA expression
between the CON group and control lentivirus infected cells (NC)
group (Fig. 1C). Western blot assay
also showed that lentivirus-delivered AURKA-shRNA silenced AURKA
protein expression in SAOS-2 and U2OS cells (Fig. 1D). These results showed that AURKA
gene could be significantly and sustainably downregulated by
lentivirus-mediated AURKA-specific shRNA in OS cells.
AURKA silencing in OS cells leads to the
decrease of colony formation ability in vitro and tumorigenicity in
vivo
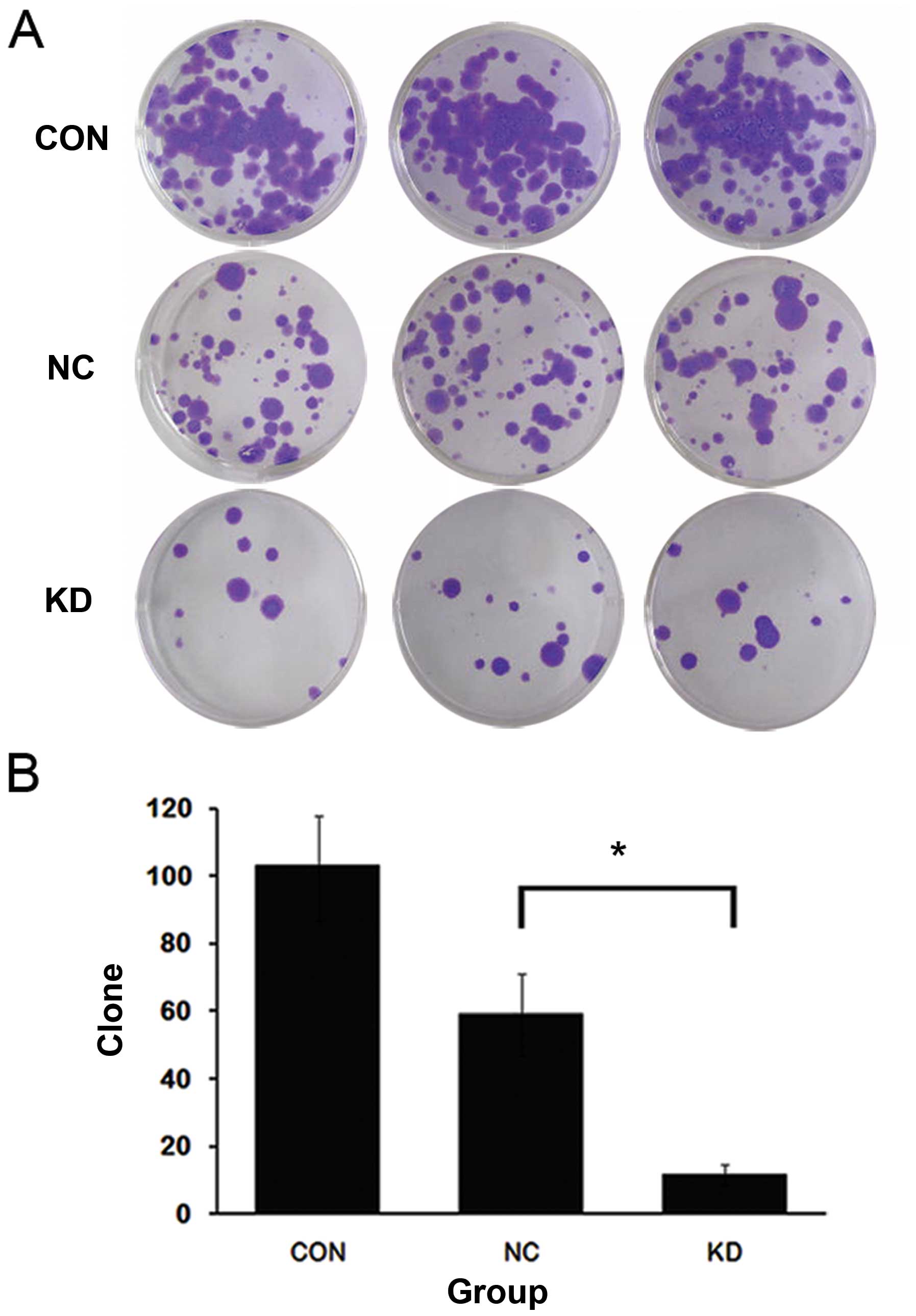
Then we explored how AURKA silencing affects
tumorigenesis of OS cells in vitro and in vivo. The
colony formation assay showed that compared to CON and NC groups,
KD showed a marked decrease (~80%) of the colony number in SAOS-2
cells (Fig. 2). These data
suggested that AURKA silencing suppressed the tumorigenic ability
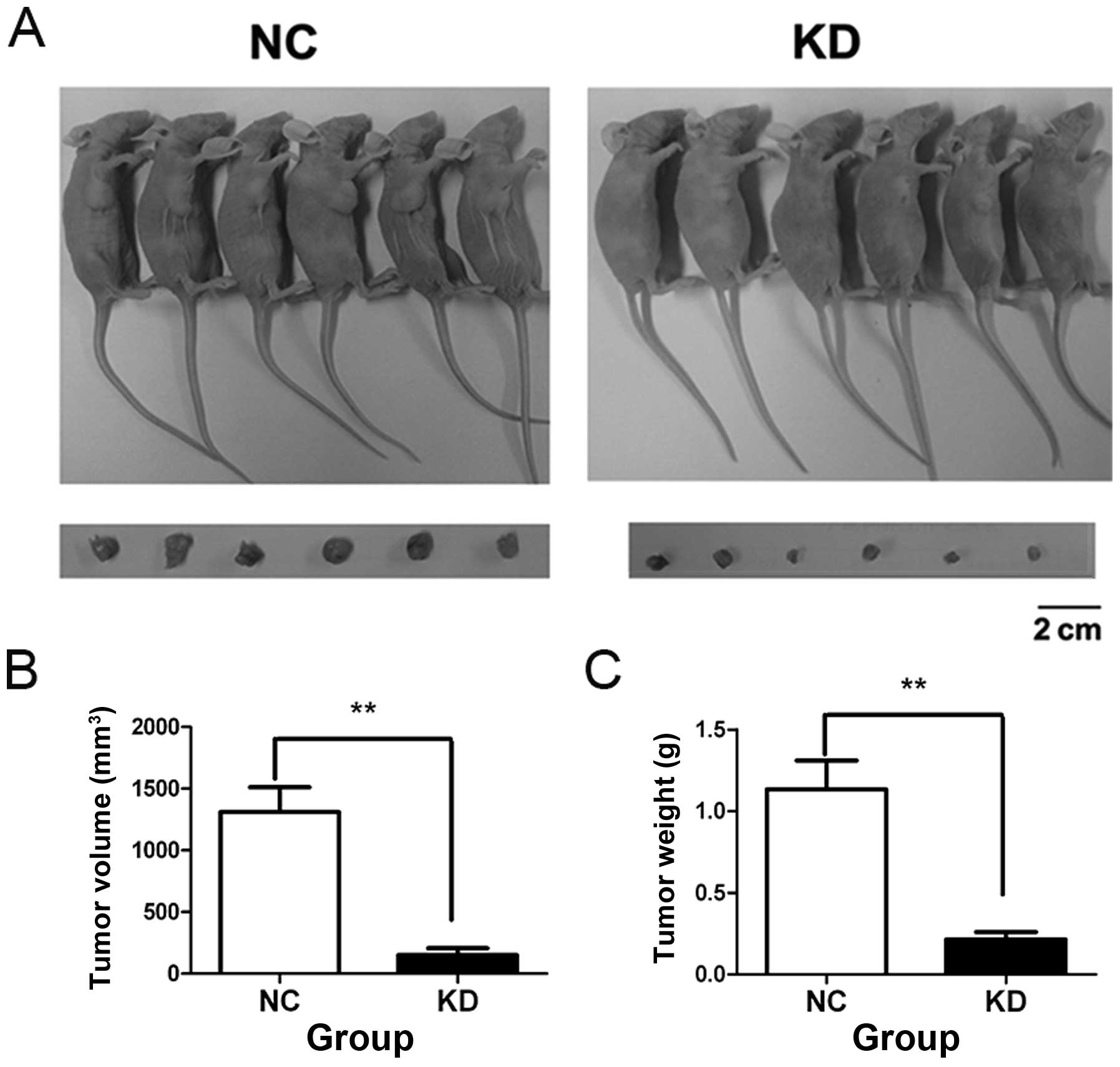
of SAOS-2 cells in vitro. We injected the SAOS-2 cells of NC
and KD groups into axilla of the nude mice, respectively.
Representative pictures of tumors at 3 weeks after cell injection
are shown in Fig. 3A. Compared to
NC group, the mean tumor volume and weight of KD group were reduced
by 90 and 80%, respectively (Fig. 3B
and C). These data demonstrated that AURKA silence
significantly inhibits colony formation ability and tumorigenicity
of SAOS-2 cells.
AURKA silence induces cell apoptosis and
cell cycle arrest at G2/M phase in OS cells
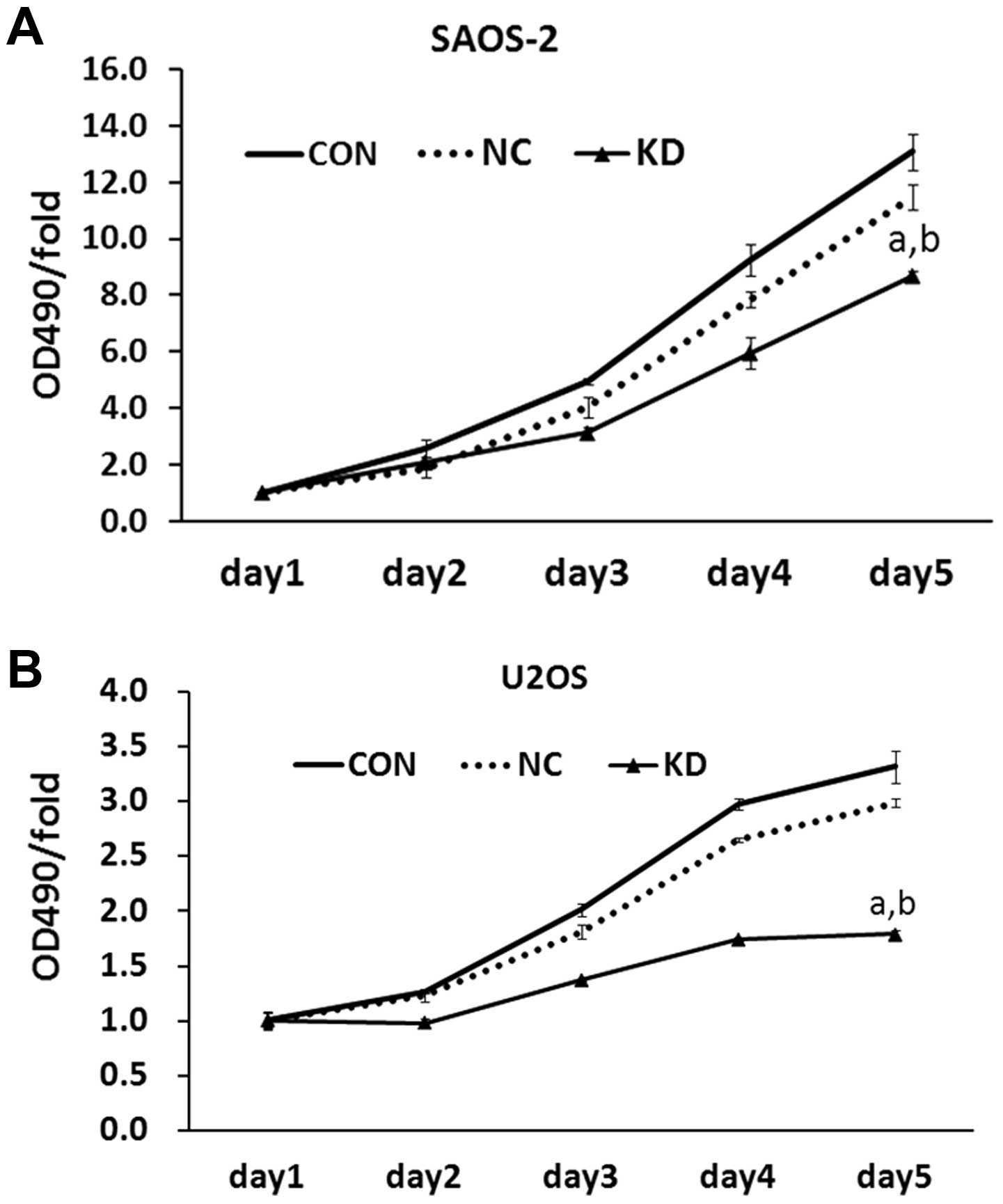
We attempted to explore why AURKA silence affected
tumorigenesis of human OS cells. MTT assay showed that AURKA
silence decreased cell viability of SAOS-2 cells in a
time-dependent manner. Following a 5-day period, cell viability of
KD group was only 66% of that of CON group, whereas control
lentivirus had no effect on the viability of SAOS-2 cells (Fig. 4A). The data in U2OS cells
demonstrated a similar result (Fig.
4B). This showed that AURKA silencing remarkably inhibits cell
viability in OS cells.
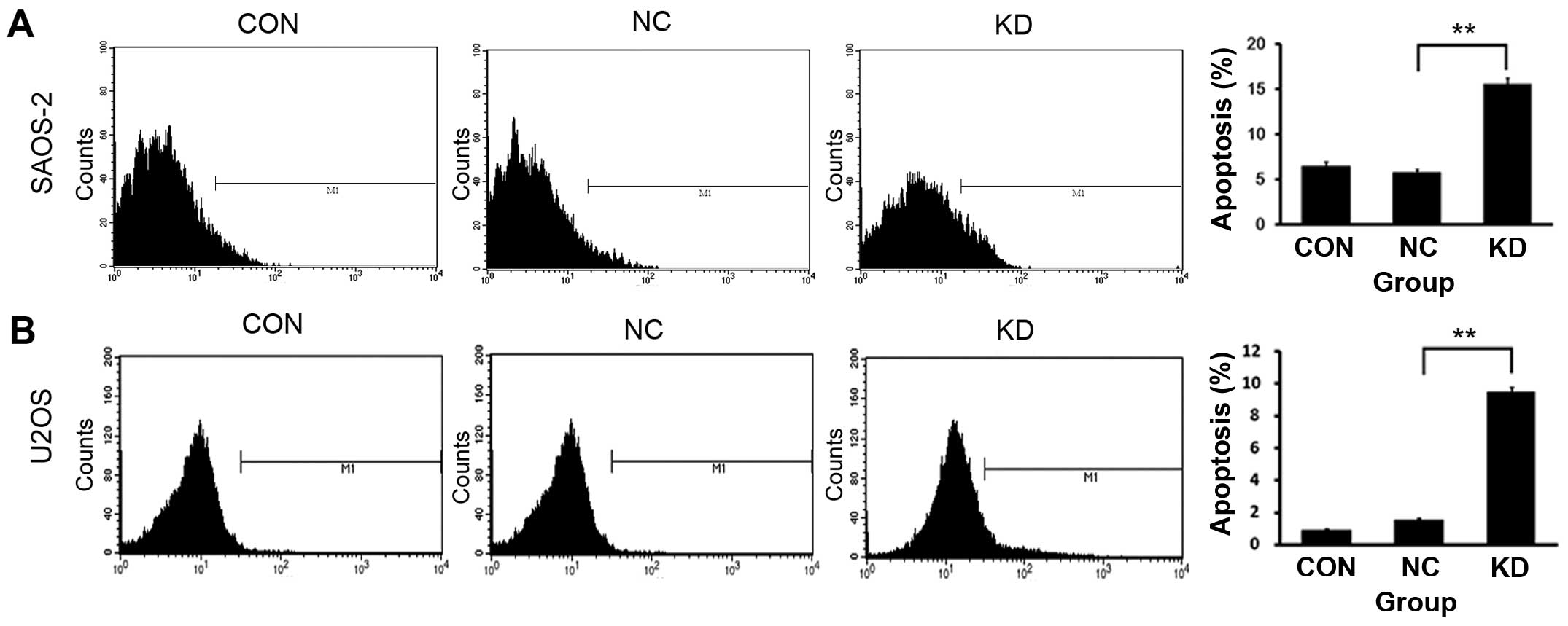
We further investigated how AURKA silence inhibits
the viability of OS cells. We used Annexin V-APC staining and FACS
analysis to determine whether the downregulation of AURKA gene
induces OS cell apoptosis. The data indicated that KD group had
more apoptotic cells (Annexin V positive) compared to those of CON
and NC group (Fig. 5A and B). We
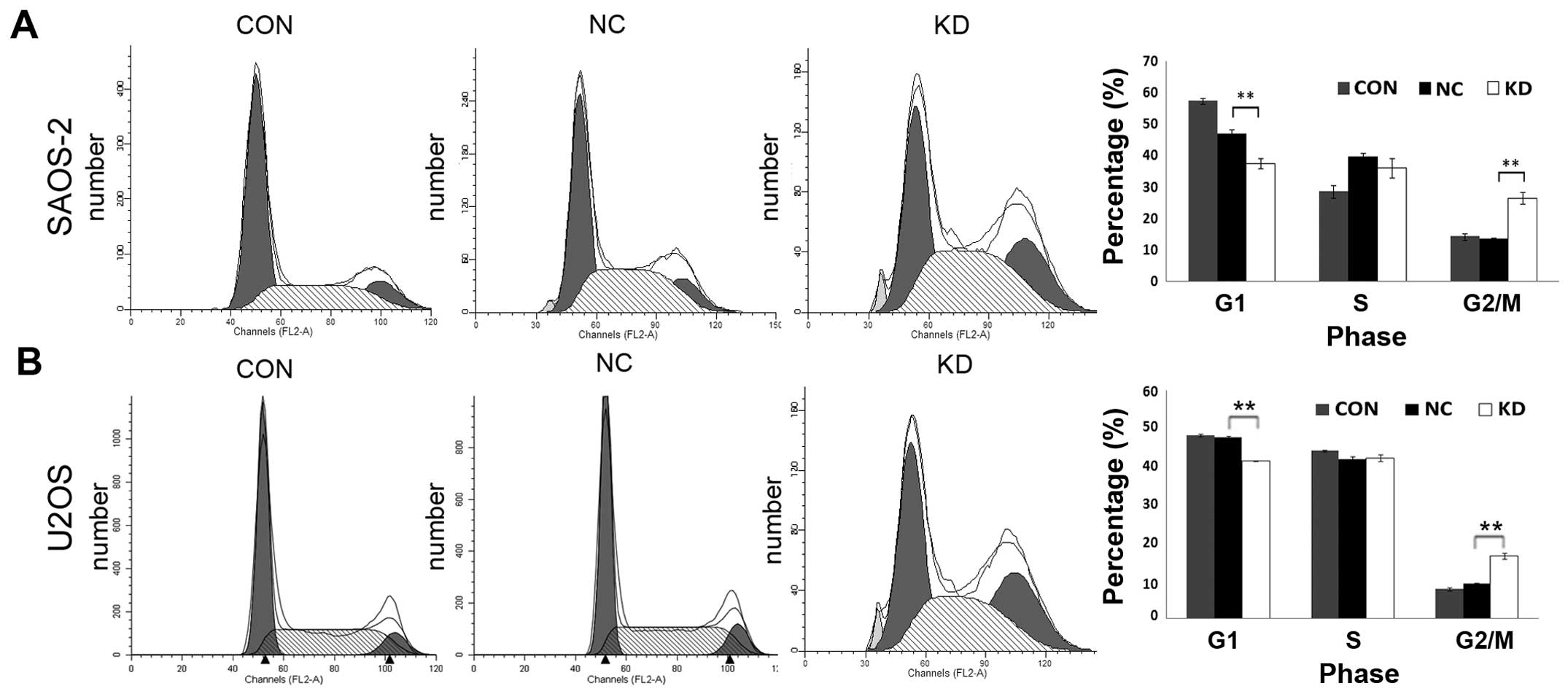
used PI staining and FACS analysis to identify AURKA-shRNA
lentivirus-induced changes in the cell cycle of OS cells. The data
showed that downregulation of AURKA expression led to the
percentage of SAOS-2 cells in G0/G1 phase significantly decreased
from 46.85% in NC group to 37.5% in KD group, and the percentage of
SAOS-2 cells in G2/M phase prominently increased from 13.48% in NC
group to 26.49% in KD group. There were no significant difference
in cell cycle distribution between CON group and NC group (Fig. 6A). The study in U2OS cells revealed
similar results (Fig. 6B). In view
of the decrease of cell viability, these data suggested that AURKA
silence caused an arrest in G2/M phase, but no more proliferative
activity. The above results showed that AURKA silence suppressed
cell viability by inducing cell apoptosis and cell cycle arrest at
G2/M phase in OS cells.
Discussion
Surgery remains the main option for OS patients.
However, even with the advanced techniques available at present,
the rate of local recurrence in non-metastatic OS patients is still
as high as 46% (14). Other
conventional OS treatments, including chemotherapy and
radiotherapy, are mainly palliative alternatives for cases where
surgery is not feasible. Both chemotherapy and radiotherapy have
disadvantages, as it is not possible to differentiate between
normal and cancerous cells, leading to destruction of cancerous as
well as normal cells. Therefore, novel therapeutic approaches are
urgently required.
Overexpression of oncogenes and upregulation of
their associated signaling transduction pathways are known as the
driving forces in OS progression (15). RNAi is an effective method for
silencing specific genes (16).
Compared to conventional targeting gene technologies including
chemical inhibitors, ribozymes and deoxyribozymes, which often
cause side-effects, shRNA are more specific inhibitors of gene
expression with less toxicity, and can be relatively easily
infected into cancer cells by lentivirus delivery (17). Hence, lentivirus-delivered shRNA
mediated gene silencing has significant therapeutic potential for
cancer.
In the present study, our data suggested that
AURKA-shRNA lentivirus leads to efficient and specific inhibition
of endogenous AURKA mRNA expression in human OS cells. AURKA-shRNA
lentivirus infected SAOS-2 cells prominently decreased
colony-forming ability in vitro and tumorigenicity in
vivo. AURKA silence showed a significant decrease of cell
viability and a cell cycle arrest in G2/M phase in OS cells. As
reported previously, AURKA is localized mainly at spindle poles and
the mitotic spindle during mitosis, where it regulates the
functions of centrosomes, spindles and kinetochores required for
proper mitotic progression (18).
Overexpression of AURKA contributes to genetic instability and
tumorigenesis by disrupting the proper assembly of the mitotic
checkpoint complex at the level of the Cdc20-BubR1 interaction
(19). Moreover, AURKA is a key
regulatory component of the p53 pathway, as its overexpression
leads to increased p53 degradation, thus facilitating oncogenic
transformation (20). These
characteristics provide an explanation for the decrease of cell
viability and tumorigenicity of AURKA-shRNA infected cells.
Moreover, although AURKA is also expressed in normal cells during
mitosis, its expression level in normal tissue is quite low.
Therefore, AURKA is an ideal target for shRNA-mediated gene cancer
therapy.
In summary, our studies investigated and confirmed
the significant effect of downregulation of AURKA expression in
human OS cells, showing that AURKA silence can induce cell
apoptosis and cell arrest in G2/M phase of the cell cycle, and,
thus, inhibits colony formation ability and tumorigenicity of OS
cells. Furthermore, the present study exhibited the feasibility of
AURKA-targeting gene therapy for OS, which provided an important
theoretical direction for clinical practice of OS.
Acknowledgements
The present study was supported by a grant from the
National Natural Science Foundation of China (no. 81172347). The
authors thank Dr Wenxiang Wei (Soochow University, Suzhou, China)
for his sincere help and technical support.
References
|
1
|
Dorfman HD and Czerniak B: Bone cancers.
Cancer. 75:203–210. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bielack SS, Kempf-Bielack B, Delling G, et
al: Prognostic factors in high-grade osteosarcoma of the
extremities or trunk: an analysis of 1,702 patients treated on
neoadjuvant cooperative osteosarcoma study group protocols. J Clin
Oncol. 20:776–790. 2002. View Article : Google Scholar
|
|
3
|
Culver KW, Anderson WF and Blaese RM:
Lymphocyte gene therapy. Hum Gene Ther. 2:107–109. 1991. View Article : Google Scholar
|
|
4
|
Abdelrahim M, Safe S, Baker C and
Abudayyeh A: RNAi and cancer: implications and applications. J RNAi
Gene Silencing. 2:136–145. 2006.PubMed/NCBI
|
|
5
|
Rubinson DA, Dillon CP, Kwiatkowski AV, et
al: A lentivirus-based system to functionally silence genes in
primary mammalian cells, stem cells and transgenic mice by RNA
interference. Nat Genet. 33:401–406. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Manilla P, Rebello T, Afable C, et al:
Regulatory considerations for novel gene therapy products: a review
of the process leading to the first clinical lentiviral vector. Hum
Gene Ther. 16:17–25. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dutertre S, Descamps S and Prigent C: On
the role of aurora-A in centrosome function. Oncogene.
21:6175–6183. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tong T, Zhong Y, Kong J, et al:
Overexpression of Aurora-A contributes to malignant development of
human esophageal squamous cell carcinoma. Clin Cancer Res.
10:7304–7310. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tanaka E, Hashimoto Y, Ito T, et al: The
clinical significance of Aurora-A/STK15/BTAK expression in human
esophageal squamous cell carcinoma. Clin Cancer Res. 11:1827–1834.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li D, Zhu J, Firozi PF, et al:
Overexpression of oncogenic STK15/BTAK/Aurora A kinase in human
pancreatic cancer. Clin Cancer Res. 9:991–997. 2003.PubMed/NCBI
|
|
11
|
Gritsko TM, Coppola D, Paciga JE, et al:
Activation and overexpression of centrosome kinase BTAK/Aurora-A in
human ovarian cancer. Clin Cancer Res. 9:1420–1426. 2003.PubMed/NCBI
|
|
12
|
Miyoshi Y, Iwao K, Egawa C and Noguchi S:
Association of centrosomal kinase STK15/BTAK mRNA expression with
chromosomal instability in human breast cancers. Int J Cancer.
92:370–373. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sen S, Zhou H, Zhang RD, et al:
Amplification/overexpression of a mitotic kinase gene in human
bladder cancer. J Natl Cancer Inst. 94:1320–1329. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bacci G, Ferrari S, Mercuri M, et al:
Neoadjuvant chemotherapy for osteosarcoma of the extremities in
patients aged 41–60 years: outcome in 34 cases treated with
adriamycin, cisplatinum and ifosfamide between 1984 and 1999. Acta
Orthop. 78:377–384. 2007.
|
|
15
|
Murdoch D and Sager J: Will targeted
therapy hold its promise? an evidence-based review. Curr Opin
Oncol. 20:104–111. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li CX, Parker A, Menocal E, Xiang S,
Borodyansky L and Fruehauf JH: Delivery of RNA interference. Cell
Cycle. 5:2103–2109. 2006. View Article : Google Scholar
|
|
17
|
Dave RS and Pomerantz RJ: RNA
interference: on the road to an alternate therapeutic strategy! Rev
Med Virol. 13:373–385. 2003.PubMed/NCBI
|
|
18
|
Marumoto T, Zhang D and Saya H: Aurora-A -
a guardian of poles. Nat Rev Cancer. 5:42–50. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ke YW, Dou Z, Zhang J and Yao XB: Function
and regulation of Aurora/Ipl1p kinase family in cell division. Cell
Res. 13:69–81. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Katayama H, Sasai K, Kawai H, et al:
Phosphorylation by Aurora kinase A induces Mdm2-mediated
destabilization and inhibition of p53. Nat Genet. 36:55–62. 2004.
View Article : Google Scholar : PubMed/NCBI
|