Introduction
As known, a hypoxic microenvironment can induce the
resistance of cells to radiotherapy in solid tumors (1,2).
Several factors are considered to be responsible for the refractory
effect of cancer cells to radiation, such as a high level of
reduced glutathione (GSH), expression of hypoxia inducible factor-1
(HIF-1), upregulation of the EGFR pathway and the alteration of
cell metabolism (3–6). In the development of therapies against
tumors, utilizing chemical and natural drugs is a promising method
for improving therapeutic response and outcome (2,7).
GSH, the tripeptide thiol
L-α-glutamyl-L-cysteinyl-glycine, is the most abundant non-protein
thiol in mammalian cells, and is regarded as one of the most
important factors for maintaining cellular redox homeostasis and
attenuating the injury of oxidative stress. It is synthesized in
the cytosol via an enzymatic reaction consisting of two steps: the
formation of α-glutamylcysteine from L-glutamate and L-cysteine,
and the formation of GSH from α-glutamylcysteine and glycine. The
first step in the enzymatic reaction is catalyzed by
α-glutamylcystine synthetase (α-GCS), followed by glutathione
synthetase (GSS) catalyzing to form GSH. α-GCS is also regarded as
the key rate-limiting enzyme in de novo GSH synthesis
(8,9). Data have been reported indicating that
hypoxia may enhance the intracellular GSH content to cause an
adaptive response to a hypoxic environment (10–12).
Research on the biology of tumors has found that intracellular GSH
contents are increased in various types of tumor cells (13,14).
Since a high level of GSH often promotes cancer cell survival and
resistance to radiotherapy by scavenging reactive oxygen species
(ROS) and free radicals (FRs), a GSH-depletion strategy may be used
as an effective tool to enhance the radiosensitivity of hypoxic
cancer cells (15,16).
HIF-1 is a heterodimer composed of an
oxygen-sensitive α subunit and a constitutively expressed β
subunit. Hydroxylated HIF-1α forms a complex with the von
Hippel-Lindau protein (VHL) resulting in HIF-1α ubiquitination by
the E3 ubiquitin protein ligase and subsequent proteosomal
degradation. It has been demonstrated that expression of HIF-1α in
tumor cells targets the transcription of many genes involved in DNA
repair and cellular resistance to various physical and chemical
injuries. High expression of HIF-1α in tumors indicates
ubiquitously poor prognosis and failure of treatment (17,18).
The strong relevance of HIF-1α in the refractory of hypoxic tumor
cells to ionizing radiation has been elucidated by numerous
experimental data, indicating that the inhibition of HIF-1α leads
to the enhancement of cancer cell radiosensitivity (19–21).
Oleanolic acid (3b-hydroxy-olea-12-en-28-oic acid,
OA) extracted from plants belongs to the triterpenoid family. It is
commonly used in various diseases, as an anti-inflammatory, for
hepatotoxicity protection, and the recovery of the hematopoietic
system after irradiation (22–24).
In addition, other data have shown that OA and its derivatives
potentiated antitumor activity via cell cycle arrest, generating
reactive oxygen species (ROS), and loss of mitochondrial membrane
potential (25,26). Thereby, OA is considered as a new
anticancer drug in combination with other conventional therapeutics
(27). Our previous results showed
that the monomer OA may increase the lethal effect on aerobic tumor
cells exposed to irradiation via the attenuation of intracellular
GSH content (28). However, it
remains unclear whether OA regulates the radiosensitivity of
hypoxic tumor cells. In the present study, cobalt chloride
(CoCl2) was used to generate a hypoxic microenvironment
in lung cancer and glioma cells. Subsequently, the alteration in
the radiosensitivity of these hypoxic cells was investigated
following OA treatment. Intracellular GSH content and the level of
HIF-1α expression were simultaneously observed under the same
conditions.
Materials and methods
Cell culture and treatment
The rat glioma C6 and human lung cancer A549 cell
lines (Cell Bank, Chinese Academy of Sciences) were cultured in
RPMI-1640 medium (BAL Gibco, Grand Island, NY, USA) supplemented
with 10% FBS (Gibco), penicillin (100 U/ml), streptomycin (100
μg/ml) (Sigma-Aldrich) at 37°C in an incubator containing a humid
atmosphere of 95% air and 5% CO2 and propagated
according to the protocol supplied by the American Type Culture
Collection. The cells in the exponential phase of growth were
incubated in culture media with 100 μm CoCl2
(Sigma-Aldrich), a common mimetic hypoxia reagent. OA was purchased
from Nanjing Zelang Medical Technology Co., Ltd., (Jiangsu, China)
and was dissolved in dimethyl sulfoxide (DMSO; Sigma) at a stock
concentration of 250 μg/ml and stored at −20°C. The cells were
treated with OA at different concentrations for 24 h prior to
exposure to irradiation.
Cell viability assay
The influence of OA on cell growth was determined
using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
bromide (MTT; Sigma) method. C6 and A549 cells were seeded in
96-well plates at a density of 5×103 cells/well. They
were then treated with OA at different concentrations for 24 h.
Furthermore, the medium was replaced with fresh medium allowing
cells to undergo continuous growth up to 72 h. MTT dye was added to
a final concentration of 50 mg/ml, and cells were subsequently
incubated for another 4 h at 37°C. The medium containing residual
MTT dye was carefully aspirated from each of the wells, and 200 μl
DMSO (Sigma-Aldrich) was added to each well to dissolve the reduced
formazan dye. The fraction of viable cells was calculated by
comparing the optical absorbance of the culture exposed to OA
treatment with that of the untreated control.
Irradiation
Irradiation was emitted using a 6 MV X-ray linear
accelerator (Varian Medical Systems, Inc., Palo Alto, CA, USA) at a
dose rate of 250 cGy/min.
Clonogenic assay
The radiosensitivity of tumor cells was determined
using the clonogenic assay. Both tumor cell lines were seeded and
cultured overnight at an appropriate density in T25 flasks, and
subsequently the drugs at different concentrations were added into
the medium for 24 h. After being pretreated with control and OA,
cells were subjected to 0, 1, 2, 3, 5 or 7 Gy X-ray irradiation.
The medium was then replaced with fresh medium allowing cells to
continuously grow for colony formation for 9 to 12 days. Cell
colonies were fixed by absolute methanol and stained with Giemsa
(Sigma-Aldrich) for counting. The clonogenic survival fraction (SF)
was calculated as the number of colonies/(the number of seeded
cells × plating efficiency). Plating efficiency was defined as the
number of colonies/the number of seeded cells of the untreated
control. Survival curve was fitted with the single target
multi-model of an equation: S=1–1(1−e−D/D0)N.
The oxygen enhancement ratio (OER) was calculated accordingly,
comparing the hypoxic D0 with the corresponding aerobic
D0.
Micronucleus assay
Micronucleus (MN) frequencies were tested with the
cytokinesis-block technique as a biological end point for the
response of cells under mimetic hypoxia to irradiation. Briefly,
the cells were exposed to 0.83 μg/ml cytochalasin B (Sigma-Aldrich)
for 19–20 h followed by 75 mM KCl hypotonic treatment for 1–3 min
and then fixed in situ with methanol:acetic acid (9:1 v/v)
for 30 min. Air-dried cells were stained with 5% Giemsa for 10 min.
Micronuclei were scored in binucleated cells, and the formation of
binucleated cells was measured as the percentage of the total
number of cells scored. For each sample, at least 1,000 binucleated
cells were counted. The MN yield, YMN, is the ratio of
the number of micronuclei to the number of binucleated cells
scored.
Intracellular GSH assay
After triplicate samples of 106 cells
were treated with different reagents, the intracellular GSH content
was measured with the glutathione
reductase/5,5′-dithiobis-(2-nitrobenzoic acid) (DTNB) recycling
assay kit (Beyotime Institute of Biotechnology, Shanghai, China)
following the methods recommended by the manufacturer. Briefly, GSH
was determined using a reaction mixture, containing 50 μl of cell
lysates, 50 μl of 2.4 mM DTNB, and 50 μl of 10.64 mU/μl glutathione
reductase in the assay buffer (pH 7.5) containing 153 mM sodium
phosphate and 8.4 mM EDTA. After a 5-min incubation at 25°C, the
reaction was started by the addition of 50 μl NADPH solution (0.16
mg/ml) in assay buffer. The standard sample and checking sample
cuvettes were placed into a dual-beam spectrophotometer, and the
increases in absorbance at 412 nm were followed as a function of
time.
Intracellular α-GCS activity assay
Cells (106) were homogenized in 50 mM
potassium phosphate (pH 7.5) containing TES/SB buffer (20 mM Tris,
1 mM EDTA, 250 mM sucrose, 20 mM sodium borate, 2 mM serine) for
the α-GCS assay. Homogenates were centrifuged at 12,000 rpm (15
min, 4°C), and the supernatants were maintained on ice for
determination of enzyme activity. The protein concentration of the
cell supernatants was measured using the Bio-Rad DC protein assay
kit (Bio-Rad Laboratories, Hertfordshire UK) and enzyme activity
was reported as units/mg protein, where a unit of activity is the
amount of enzyme required to convert 1 μmole of substrate to
product per minute at 25°C. The α-GCS assay is an adaptation of the
method previously described, in which α-GCS in cell extracts
synthesizes α-glutamylcysteine which is then reacted with
2,3-naphthalenedicarboxaldehyde (NDA) to form a highly fluorescent
product that can be measured fluorimetrically at 520 nm (29).
Intracellular GSS activity assay
Tumor cells were plated in 60-mm culture dishes at a
density of 106 cells/dish then divided into different
groups for various methods of pretreatment. The intracellular GSS
content was measured using the GSS assay kit (Hefei Lanxu Biotech
Co., Ltd., Hefei, China). Briefly, the cells were subjected to
repeated freeze-thaw cycles to lyse these cells for release of
intracellular components. Cell lysates were centrifuged at 3,000
rpm (20 min, 4°C), and the supernatants were maintained on ice for
determination of enzyme activity. All procedures were performed
according to the protocol of the kit. After the reaction was
terminated, the absorbance was measured at 450 nm on an ELISA
reader. The activity of GSS in the sample was then determined by
comparing the OD of the samples to the standard curve.
Western blot analysis of HIF-1α
expression
The cells in the different treatment groups were
scraped off from the culture flasks and lysed in lysis buffer
containing 10% glycerol, 10 mM Tris-HCl (pH 6.8), 1% SDS, 5 mM
dithiothreitol (DTT) and 1X complete protease inhibitor cocktail
(Sigma, St. Louis, MO, USA). The method of Bradford was used to
detect the concentrations of protein in the diverse samples. The
protein concentration was measured using an auto multifunction
microplate reader. Fifty micrograms of proteins was separated by 8%
polyacrylamide-SDS in consecutive gel electrophoresis. The
separated proteins were electrophoretically transferred to a
polyvinylidene difluoride membrane. Membranes were blocked with 5%
skim milk in Tris-buffered saline (TBS) containing 0.1% Tween-20 at
room temperature for 1 h and then incubated with mouse HIF-1α
antibody (Abcam, Cambridge, MA, USA) at a 1:500 dilution overnight
at 4°C, followed by goat anti-mouse IgG for 1 h at room
temperature. Signals were detected with enhanced chemiluminescence
(ECL Plus; Amersham, Pittsburgh, PA, USA). Microtubule protein
(Tubulin; Abcam) at a 1:1,000 dilution was used as an internal
control to observe the changes in the HIF-1α bands.
Statistical analysis
Data are reported as the means ± SEM of three
separate experiments. Statistical significance was measured by the
independent sample t-test and analysis of variance. A value of
P<0.05 was considered to indicate a statistically significant
result.
Results
Selection of the experimental
concentration of OA
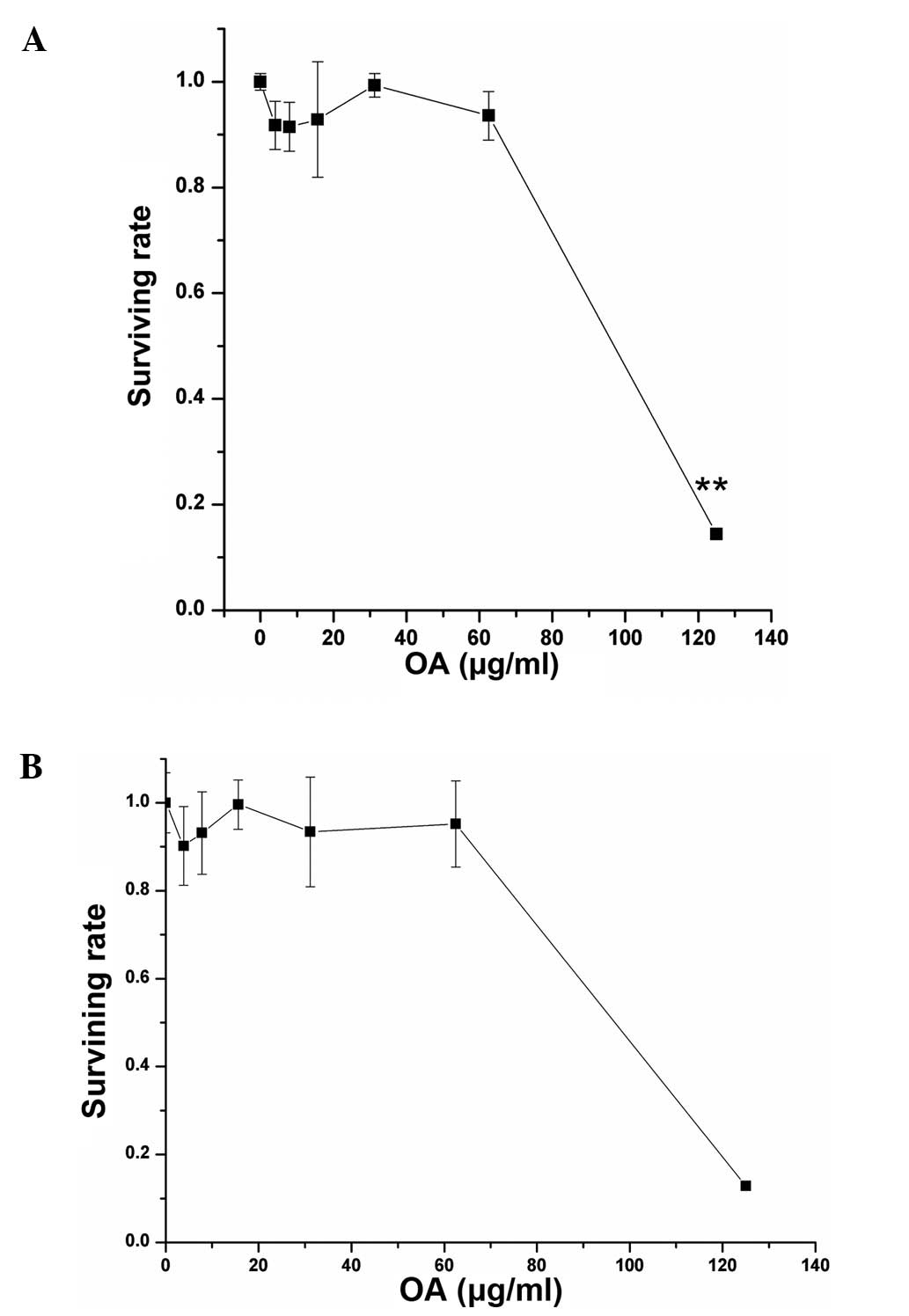
The cytotoxicity test showed that the half maximal
inhibitory concentrations (IC50) of OA in the hypoxic C6
and A549 cells were 80 and 81 μg/ml, respectively (Fig. 1). The concentrations of OA at 16
μg/ml (20% IC50) and 24 μg/ml (30% IC50) were
used to pretreat the cells in order to observe the alteration of
radiosensitivity in both hypoxic tumor cell lines.
Alteration of tumor cell
radiosensitivity
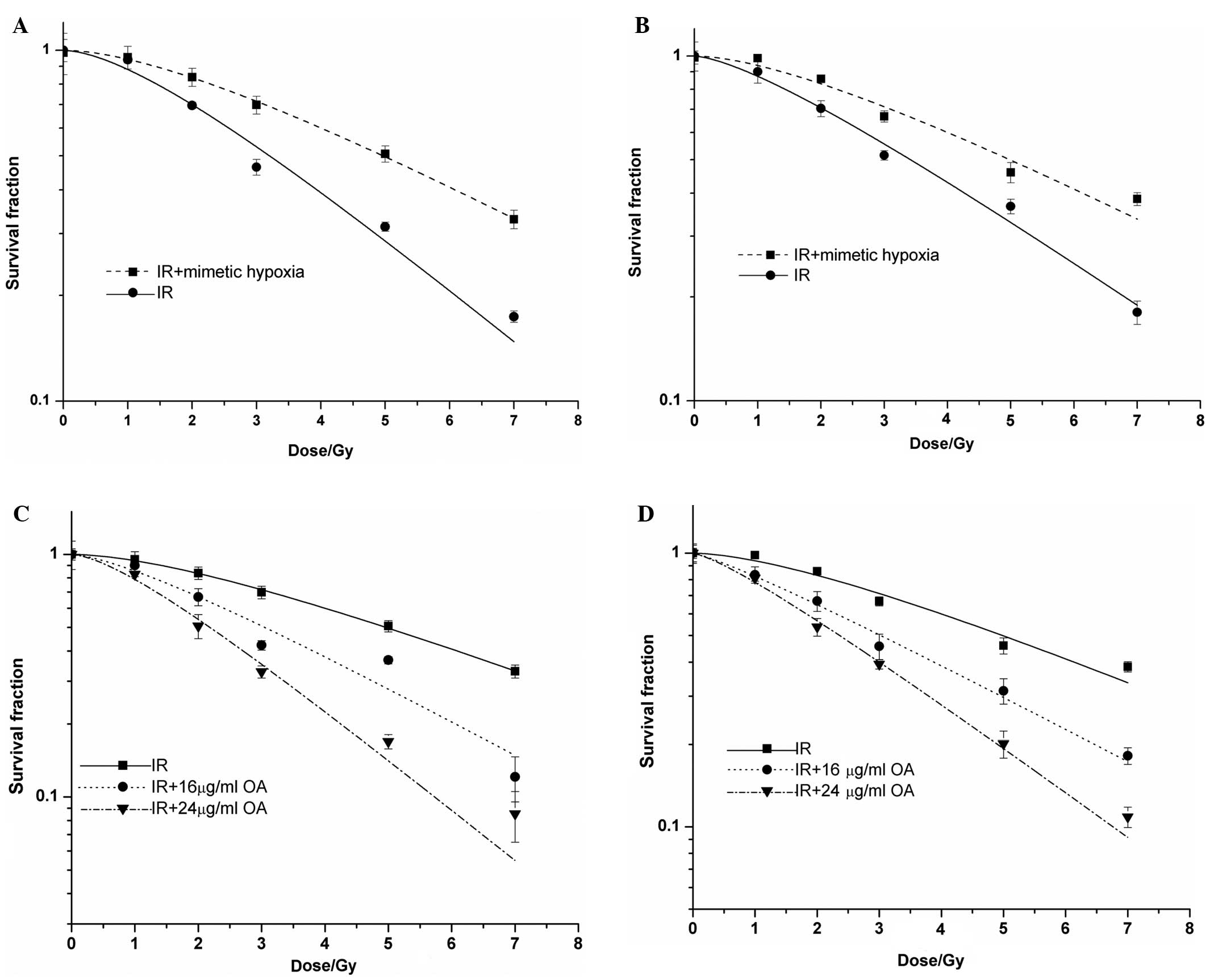
No statistical differences were observed in the
numbers of colonies formed in the C6 and A549 cells with
CoCl2 treatment and those without CoCl2
treatment. However, after the cells were exposed to radiation, both
cell lines in the mimetic hypoxic microenvironment had a higher
resistance to irradiation. The OER values of the C6 and A549 cells
were 1.52 and 1.31, respectively (Fig.
2A and B). Subsequently, we observed the alteration in
radiosensitivity of the C6 and A549 cells following treatment with
OA at the different concentrations. After the hypoxic cells were
exposed to irradiation, the SF of the cells treated with OA was
lower than that of the cells without OA treatment. Following
calculation of the sensitive enhancement ratio (SER), the
SER of the irradiated cells was elevated concomitant with the
increase in OA concentrations. The SERs of the hypoxic C6 and A549
cells treated with OA at 30% IC50 were 2.09 and 1.71,
respectively (Fig. 2C and D).
Changes in the intracellular micronucleus
frequencies
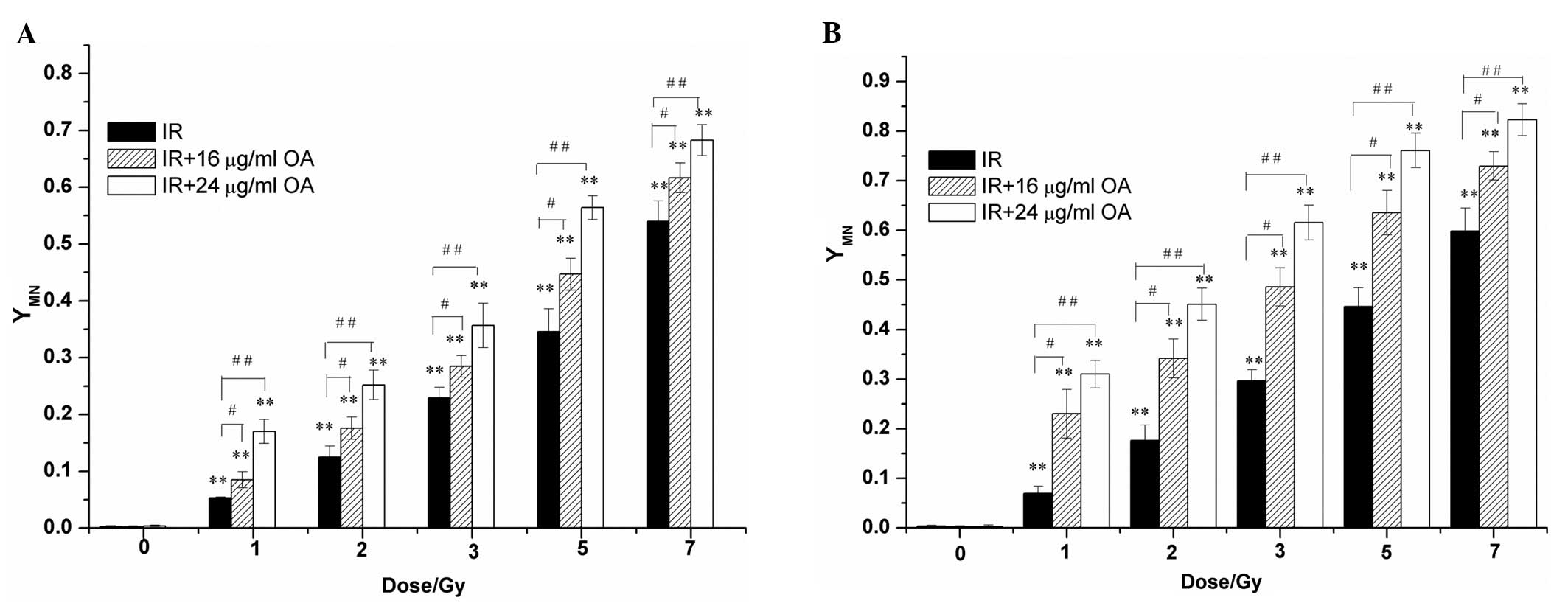
MN assay showed that there was no obvious influence
of OA on the frequencies of MN in both hypoxic cell lines unexposed
to irradiation. Subsequently, the numbers of intracellular MN were
significantly increased concomitant with the irradiation doses.
When both irradiated cell lines were pretreated with OA at
different concentrations, further enhancement in the numbers of
intracellular MN was noted. Compared with the irradiated cells
without OA treatment, the irradiated cells pretreated with 16 and
24 μg/ml OA displayed a statistically significant increase in
intracellular MN frequencies (Fig.
3).
Influence of OA on GSH level, the
activity of α-GCS and GSS
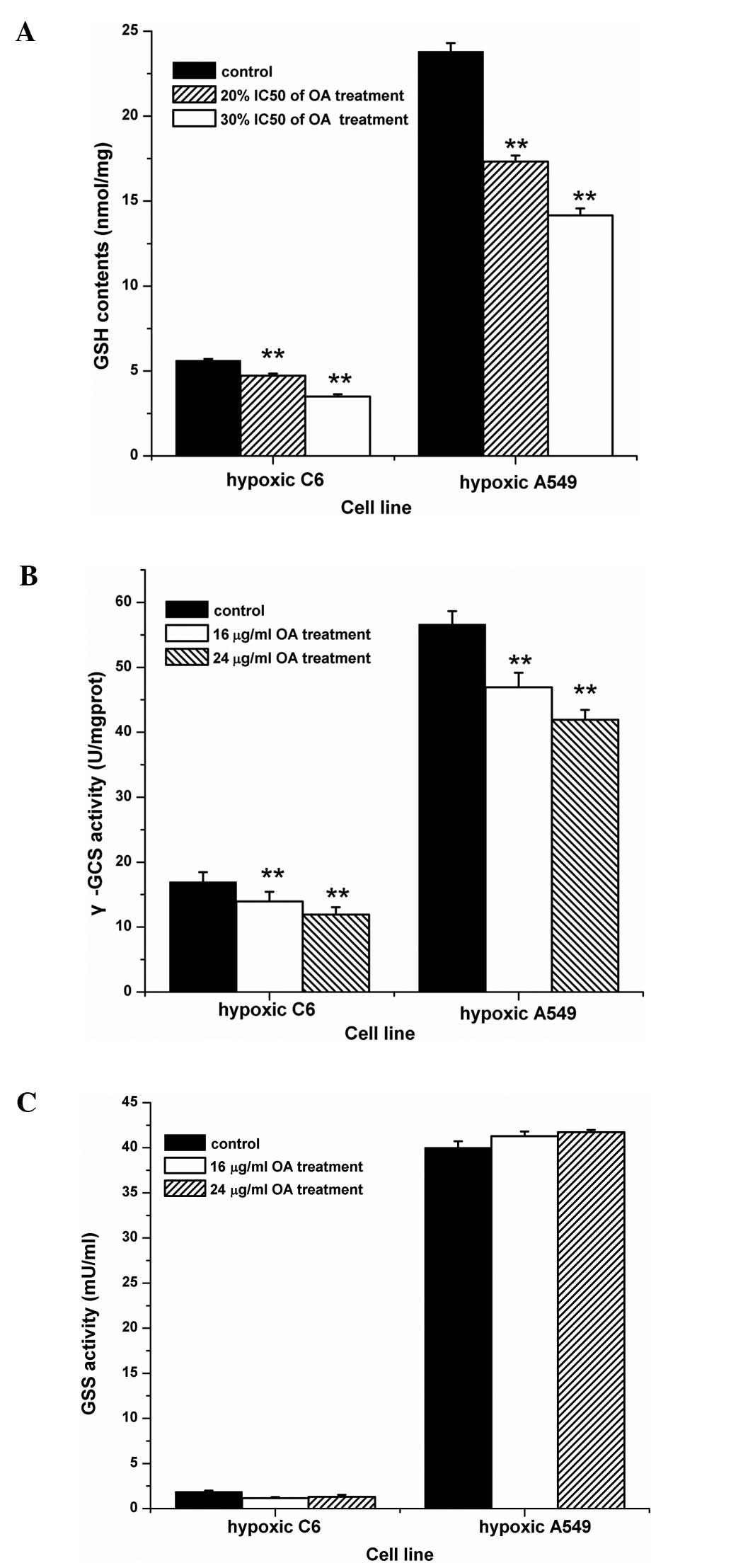
To further observe the mechanism of the influence of
OA on the radiosensitivity of hypoxic cells, intracellular GSH
levels were measured following treatment of OA at different
concentrations for 24 h. Significant decreases in the GSH levels of
hypoxic C6 cells were noted in the presence of OA, when compared
with levels in the absence of OA. Moreover, a similar phenomenon
occurred in hypoxic A549 cells, when intracellular GSH levels
showed a gradually declining tendency concomitant with increases in
OA concentrations (Fig. 4A). Since
α-GCS is the key limiting-enzyme in the synthesis of intracellular
GSH, its activity was further measured. As shown in Fig. 4B, the different concentrations of OA
significantly decreased α-GCS activity in both hypoxic cell lines.
Notably, the activity of GSS, another synthetic enzyme, did not
exhibit a statistically significant change in the tumor cells with
the same treatments (Fig. 4C).
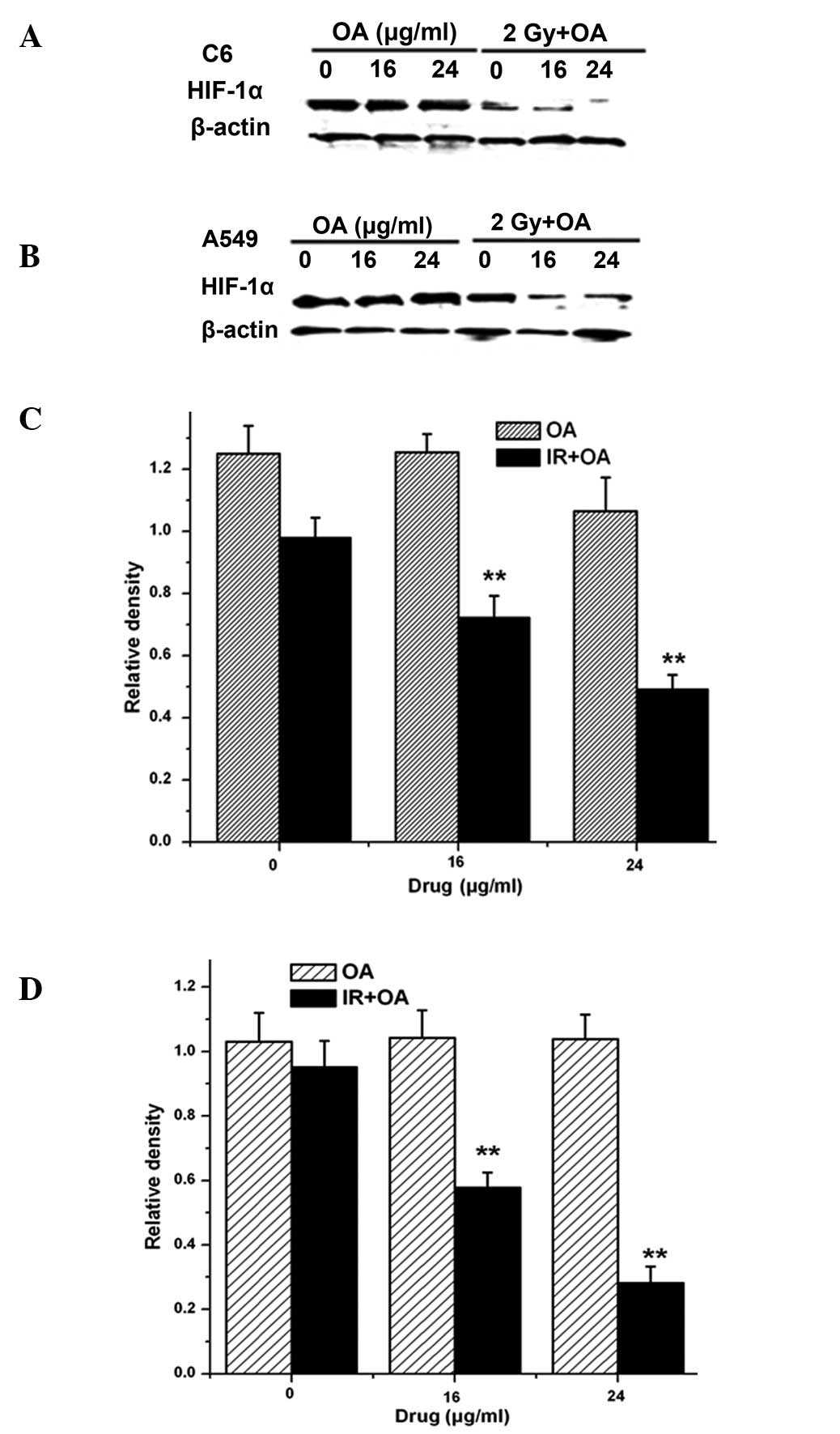
Change in the intracellular HIF-1α level
by OA pretreatment
High expression of intracellular HIF-1α was induced
by CoCl2 treatment. The hypoxic cells exposed to
irradiation still exhibited high levels of HIF-1α. Meanwhile,
HIF-1α expression in the hypoxic cells without irradiation did not
exhibit a statistically significant change following OA treatment.
However, the combination of OA treatment with irradiation
suppressed the high levels of HIF-1α expression in the hypoxic
cells (Fig. 5).
Discussion
Tumor hypoxia, which is generally attributed to an
imbalance between the demand and supply of oxygen and poorly
organized vasculature, is observed in many tumor types particularly
glioma and lung cancer (30).
Hypoxia appears to be the most important factor in the development
of radioresistance, invasiveness and more aggressive tumor
phenotypes (17,31). Therefore, regarding glioma and lung
cancer, enhancement of the efficacy of radiotherapy by hypoxic
radiosensitizers is favorable for the improvement of the
radiotherapeutic effectiveness in these two invariably fatal
diseases. Numerous natural compounds from the extracts of plants
such as curcumin and soy isoflavones have been developed and
screened (32,33). In a previous study, we found that OA
enhanced the radiosensitizing effect on aerobic C6 and A549 cells
(28). Subsequently, the effect of
OA on the radiosensitivity of tumor cells with mimetic hypoxia
treatment was observed in the present study.
Based on a large number of experimental data,
preliminary exposure to mimetic hypoxia with CoCl2
inducing a similar to real hypoxic condition may increase the
tolerance against subsequent injury of other biological and
physicochemical factors including chemotherapeutic drugs, ionizing
radiation and tert-butyl-hydroperoxide-induced oxidative stress
(34–36). Therefore, CoCl2 can be
universally used as a chemical reagent that induces biochemical and
molecular responses similar to those observed under a hypoxic
condition (37,38). Our previous results also showed that
similar to hypoxia, CoCl2 enhanced cellular
radioresistance and increased the levels of HIF-1α (36). We selected two different
concentrations of OA, with no obvious influence on cell viability,
to carry out the present experiment. It was further observed that
different doses of radiation combined with OA significant inhibited
the cell growth and an additive lethal effect was noted. According
to the calculation of D0 and SERs, the sensitivity of
tumor cells to radiation was significantly enhanced by OA
treatment.
The protective effect of GSH is very important for
the resistance of cancer cells against radiotherapeutics (15). We, therefore, investigated the
influence of OA on the level of GSH and the capability of
regulatory enzymes in hypoxic cells. The results of the present
study clearly showed that the levels of GSH in the two hypoxic cell
lines were downregulated following pretreatment of OA at 20 and 30%
IC50. Regarding the biosynthesis of GSH, the inhibition
of α-GCS activity was subsequently observed in the hypoxic cells in
the presence of OA. However, the phenomenon, the alteration of GSS
activity, was not found under the same treatment condition. It was
demonstrated that OA might decrease the level of GSH via the
inhibition of α-GCS activity in hypoxic tumor cells. According to
our previous study and other data, there was a higher level of
cellular GSH in hypoxic cells or mimetic hypoxic cells compared
with that in aerobic cells, resulting in the refractoriness of
cells to irradiation (36,39,40).
Therefore, the combination of OA and radiation to effectively
destroy hypoxic tumor cells is strongly correlated with the
inhibition of intracellular GSH biosynthesis.
As one of the biomarkers, MN assay is usually used
in the observation of DNA damage by radiation leading to the
formation of single and double strand breaks. The MN frequency in
binucleated cells indicates the degree of radiation damage
(41–43). Data have demonstrated that increased
GSH participates non-enzymatically in the protection against DNA
damage by irradiation following the significant reduction in MN
frequency (44,45). On the contrary, the attenuation of
GSH increased the formation of MN in irradiated cells (46). Our results showed that, compared to
the irradiated cells without OA treatment, there was a statistical
elevation in the MN frequency in the hypoxic cells treated with OA
at different concentrations after irradiation. It was further found
that OA increased the radiosensitivity of hypoxic cells by the
depletion of intracellular GSH.
HIF-1α has attracted attention in the field of
cancer radio-chemotherapy. The expression of HIF-1α induced by
hypoxia or CoCl2 imitating hypoxia enhances the
refractoriness of cells to irradiation (47,48).
On the other hand, increased radiosensitivity of hypoxic cells is
associated with the inhibition of intracellular HIF-1α (4). The results from our experiment were
not due to the statistical alteration of HIF-1α expression by OA or
radiation treatment alone. Notably, the findings showed that, after
the hypoxic cells were exposed to irradiation, there were
significantly decreased levels of HIF-1α following pretreatment of
OA at 20 and 30% IC50. Since HIF-1α is involved in the
restoration of DNA damage in irradiated cells, OA effectively
attenuated the repair potential in both hypoxic cell lines
following irradiation by inhibition of HIF-1α expression. Other
data indicate that the inhibition of intracellular HIF-1α by
natural products extracted from plants such as soy isoflavones and
honokiol reduced the resistance of hypoxic tumor cells to
irradiation (33,49). In addition, research revealed that
three triterpenic acids extracted from natural products, including
oleanolic acid, ursolic acid and maslinic acid, suppressed the
expression of HIF-1α in human liver cancer cells exposed to a
hypoxic microenvironment as observed in a previous study similarly
to our observation (50).
Therefore, the downregulation of HIF-1α level is another target, by
which OA modulated the sensitivity of the two hypoxic tumor cell
lines to radiation.
In conclusion, the present study demonstrated that,
under chemical hypoxia, OA increased the sensitivity of rat glioma
C6 cells and human lung cancer A549 cells to radiation. The primary
mechanism of the effect of OA may involve the downregulation of
intracellular GSH synthesis by inhibiting the activity of α-GCS and
the decreased expression of HIF-1α after irradiation. Collectively,
the present findings and our previous report suggest that OA has
the potential to improve the radiation response for tumor
treatment.
Acknowledgements
The present study was financially supported by the
Anhui Provincial Natural Science Foundation, China (grant no.
11040606M210).
References
|
1
|
Toustrup K, Sørensen BS, Alsner J and
Overgaard J: Hypoxia gene expression signatures as prognostic and
predictive markers in head and neck radiotherapy. Semin Radiat
Oncol. 22:119–127. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bischoff P, Altmeyer A and Dumont F:
Radiosensitising agents for the radiotherapy of cancer: advances in
traditional and hypoxia targeted radiosensitisers. Expert Opin Ther
Pat. 19:643–662. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tian J, Peehl DM, Zheng W and Knox SJ:
Anti-tumor and radiosensitization activities of the iron chelator
HDp44mT are mediated by effects on intracellular redox status.
Cancer Lett. 298:231–237. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Meijer TW, Kaanders JH, Span PN and
Bussink J: Targeting hypoxia, HIF-1, and tumor glucose metabolism
to improve radiotherapy efficacy. Clin Cancer Res. 18:5585–5594.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Karar J and Maity A: Modulating the tumor
microenvironment to increase radiation responsiveness. Cancer Biol
Ther. 8:1994–2001. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Min JW, Kim KI, Kim HA, et al:
INPP4B-mediated tumor resistance is associated with modulation of
glucose metabolism via hexokinase 2 regulation in laryngeal cancer
cells. Biochem Biophys Res Commun. 440:137–142. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Raffoul JJ, Banerjee S, Che M, et al: Soy
isoflavones enhance radiotherapy in a metastatic prostate cancer
model. Int J Cancer. 120:2491–2498. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Forman HJ, Zhang H and Rinna A:
Glutathione: overview of its protective roles, measurement, and
biosynthesis. Mol Aspects Med. 30:1–12. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Botta D, White CC, Vliet-Gregg P, et al:
Modulating GSH synthesis using glutamate cysteine ligase transgenic
and gene-targeted mice. Drug Metab Rev. 40:465–477. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Catalano V, Turdo A, Di Franco S, Dieli F,
Todaro M and Stassi G: Tumor and its microenvironment: a
synergistic interplay. Semin Cancer Biol. 23:522–532. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Saxena S, Shukla D, Saxena S, et al:
Hypoxia preconditioning by cobalt chloride enhances endurance
performance and protects skeletal muscles from exercise-induced
oxidative damage in rats. Acta Physiol. 200:249–263. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Oh C, Dong Y, Harman C, Mighty HE,
Kopelman J and Thompson LP: Chronic hypoxia differentially
increases glutathione content and α-glutamyl cysteine synthetase
expression in fetal guinea pig organs. Early Hum Dev. 84:121–127.
2008.PubMed/NCBI
|
|
13
|
Ruiz-Gómez MJ, Souviron A,
Martínez-Morillo M and Gil L: P-glycoprotein, glutathione and
glutathione S-transferase increase in a colon carcinoma cell line
by colchicine. J Physiol Biochem. 56:307–312. 2000.PubMed/NCBI
|
|
14
|
Honda T, Coppola S, Ghibelli L, et al: GSH
depletion enhances adenoviral bax-induced apoptosis in lung cancer
cells. Cancer Gene Ther. 11:249–255. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Simons AL, Parsons AD, Foster KA, Orcutt
KP, Fath MA and Spitz DR: Inhibition of glutathione and thioredoxin
metabolism enhances sensitivity to perifosine in head and neck
cancer cells. J Oncol. 2009:5195632009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Boivin A, Hanot M, Malesys C, Maalouf M,
Rousson R, Rodriguez-Lafrasse C and Ardail D: Transient alteration
of cellular redox buffering before irradiation triggers apoptosis
in head and neck carcinoma stem and non-stem cells. PLoS One.
6:e145582011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang Y, Sun M, Wang L and Jiao B: HIFs,
angiogenesis, and cancer. J Cell Biochem. 114:967–974. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Semenza GL: Hypoxia-inducible factors:
mediators of cancer progression and targets for cancer therapy.
Trends Pharmacol Sci. 33:207–214. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pires IM, Olcina MM, Anbalagan S, et al:
Targeting radiation-resistant hypoxic tumour cells through ATR
inhibition. Br J Cancer. 107:291–299. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lu H, Liang K, Lu Y and Fan Z: The
anti-EGFR antibody cetuximab sensitizes human head and neck
squamous cell carcinoma cells to radiation in part through
inhibiting radiation-induced upregulation of HIF-1α. Cancer Lett.
322:78–85. 2012.PubMed/NCBI
|
|
21
|
Staab A, Fleischer M, Loeffler J, et al:
Small interfering RNA targeting HIF-1α reduces hypoxia-dependent
transcription and radiosensitizes hypoxic HT 1080 human
fibrosarcoma cells in vitro. Strahlenther Onkol. 187:252–259.
2011.
|
|
22
|
Reisman SA, Aleksunes LM and Klaassen CD:
Oleanolic acid activates Nrf2 and protects from acetaminophen
hepatotoxicity via Nrf2-dependent and Nrf2-independent processes.
Biochem Pharmacol. 77:1273–1282. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hsu HY, Yang JJ and Lin CC: Effects of
oleanolic acid and ursolic acid on inhibiting tumor growth and
enhancing the recovery of hematopoietic system postirradiation in
mice. Cancer Lett. 111:7–13. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang EJ, Lee W, Ku SK, Song KS and Bae JS:
Anti-inflammatory activities of oleanolic acid on HMGB1 activated
HUVECs. Food Chem Toxicol. 50:1288–1294. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fujiwara Y, Komohara Y, Kudo R, Tsurushima
K, Ohnishi K, Ikeda T and Takeya M: Oleanolic acid inhibits
macrophage differentiation into the M2 phenotype and glioblastoma
cell proliferation by suppressing the activation of STAT3. Oncol
Rep. 26:1533–1537. 2011.PubMed/NCBI
|
|
26
|
Chakravarti B, Maurya R, Siddiqui JA, Bid
HK, Rajendran SM, Yadav PP and Konwar R: In vitro anti-breast
cancer activity of ethanolic extract of Wrightia tomentosa:
role of pro-apoptotic effects of oleanolic acid and urosolic acid.
J Ethnopharmacol. 142:72–79. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yamai H, Sawada N, Yoshida T, et al:
Triterpenes augment the inhibitory effects of anticancer drugs on
growth of human esophageal carcinoma cells in vitro and
suppress experimental metastasis in vivo. Int J Cancer.
125:952–960. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang J, Yu M, Xiao L, Xu S, Yi Q and Jin
W: Radiosensitizing effect of oleanolic acid on tumor cells through
the inhibition of GSH synthesis in vitro. Oncol Rep.
30:917–924. 2013.PubMed/NCBI
|
|
29
|
White CC, Viernes H, Krejsa CM, Botta D
and Kavanagh TJ: Fluorescence-based microtiter plate assay for
glutamate-cysteine ligase activity. Anal Biochem. 318:175–180.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lee ST and Scott AM: Hypoxia positron
emission tomography imaging with 18f-fluoromisonidazole.
Semin Nucl Med. 37:451–461. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bulnes S, Bengoetxea H, Ortuzar N,
Argandoña EG, Garcia-Blanco A, Rico-Barrio I and Lafuente JV:
Angiogenic signalling pathways altered in gliomas: selection
mechanisms for more aggressive neoplastic subpopulations with
invasive phenotype. J Signal Transduct. 2012:5979152012. View Article : Google Scholar
|
|
32
|
Aravindan S, Natarajan M, Herman TS,
Awasthi V and Aravindan N: Molecular basis of ‘hypoxic’
breast cancer cell radio-sensitization: phytochemicals converge on
radiation induced Rel signaling. Radiat Oncol. 8:462013.
|
|
33
|
Singh-Gupta V, Zhang H, Banerjee S, Kong
D, Raffoul JJ, Sarkar FH and Hillman GG: Radiation-induced HIF-1α
cell survival pathway is inhibited by soy isoflavones in prostate
cancer cells. Int J Cancer. 124:1675–1684. 2009.
|
|
34
|
Ji Z, Yang G, Shahzidi S, Tkacz-Stachowska
K, Suo Z, Nesland JM and Peng Q: Induction of hypoxia-inducible
factor-1α overexpression by cobalt chloride enhances cellular
resistance to photodynamic therapy. Cancer Lett. 244:182–189.
2006.
|
|
35
|
Piret JP, Lecocq C, Toffoli S, Ninane N,
Raes M and Michiels C: Hypoxia and CoCl2 protect HepG2
cells against serum deprivation- and t-BHP-induced apoptosis: a
possible anti-apoptotic role for HIF-1. Exp Cell Res. 295:340–349.
2004.PubMed/NCBI
|
|
36
|
Jin W, Wang J, Xu S, Xiao L, Chen G, Zhang
W and Li J: Radioprotective effect on HepG2 cells of low
concentrations of cobalt chloride: induction of hypoxia-inducible
factor-1 alpha and clearance of reactive oxygen species. J Radiat
Res. 54:203–209. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Law PC, Auyeung KK, Chan LY and Ko JK:
Astragalus saponins downregulate vascular endothelial growth factor
under cobalt chloride-stimulated hypoxia in colon cancer cells. BMC
Complement Altern Med. 12:1602012. View Article : Google Scholar
|
|
38
|
Brusevold IJ, Husvik C, Schreurs O,
Schenck K, Bryne M and Søland TM: Induction of invasion in an
organotypic oral cancer model by CoCl2, a hypoxia
mimetic. Eur J Oral Sci. 118:168–176. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Donnelly ET, Liu Y, Paul TK and Rockwell
S: Effects of motexafin gadolinium on DNA damage and X-ray-induced
DNA damage repair, as assessed by the Comet assay. Int J Radiat
Oncol Biol Phys. 62:1176–1186. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Vukovic V, Nicklee T and Hedley DW:
Differential effects of buthionine sulphoximine in hypoxic and
non-hypoxic regions of human cervical carcinoma xenografts.
Radiother Oncol. 60:69–73. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kashino G, Prise KM, Suzuki K, et al:
Effective suppression of bystander effects by DMSO treatment of
irradiated CHO cells. J Radiat Res. 48:327–333. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Thierens H and Vral A: The micronucleus
assay in radiation accidents. Ann Ist Super Sanita. 45:260–264.
2009.PubMed/NCBI
|
|
43
|
Romm H, Barnard S, Boulay-Greene H, et al:
Laboratory intercomparison of the cytokinesis-block micronucleus
assay. Radiat Res. 180:120–128. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tokuzumi S, Hori M, Monobe M, Hosoi Y and
Kojima S: Effect of nitric oxide on α-ray-induced micronucleus
frequency in RAW264.7 cells. Radiat Res. 164:723–732. 2005.
|
|
45
|
Kumar M, Meena P, Verma S, Kumar M and
Kumar A: Anti-tumour, anti-mutagenic and chemomodulatory potential
of Chlorophytum borivilianum. Asian Pac J Cancer Prev.
11:327–334. 2010.PubMed/NCBI
|
|
46
|
Hanot M, Boivin A, Malésys C, et al:
Glutathione depletion and carbon ion radiation potentiate clustered
DNA lesions, cell death and prevent chromosomal changes in cancer
cells progeny. PLoS One. 7:e443672012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Sendoel A, Kohler I, Fellmann C, Lowe SW
and Hengartner MO: HIF-1 antagonizes p53-mediated apoptosis through
a secreted neuronal tyrosinase. Nature. 465:577–583. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Indovina P, Ferrante A, Rainaldi G and
Santini MT: Hypoxia and ionizing radiation: changes in adhesive
properties and cell adhesion molecule expression in MG-63
three-dimensional tumor spheroids. Cell Commun Adhes. 13:185–198.
2006. View Article : Google Scholar
|
|
49
|
Lan KL, Lan KH, Sheu ML, et al: Honokiol
inhibits hypoxia-inducible factor-1 pathway. Int J Radiat Biol.
187:579–590. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Lin CC, Huang CY, Mong MC, Chan CY and Yin
MC: Antiangiogenic potential of three triterpenic acids in human
liver cancer cells. J Agric Food Chem. 59:755–762. 2011. View Article : Google Scholar : PubMed/NCBI
|