Introduction
Adenovirus (Ad) has shown promise in cancer therapy.
Ad has a high viral titer and high transduction efficiency for both
proliferating and quiescent cells; it permits the insertion of
large fragments of foreign DNA, and the viral DNA does not
integrate into the host (1). These
characteristics make it an ideal choice compared to other vectors.
However, the progress of the application of Ad in cancer gene
therapy is hindered by the lack of expression of native adenovirus
receptor on a variety of cancer types (2–4). As
such, strategies are needed to retarget the Ad vector to non-native
cellular surface receptors.
Most of the commonly used adenoviral vectors are
based on the serotype 5 (Ad5). Host cell entry of Ad5 is a two-step
process with two major interactions between Ad coat proteins and
cell surface receptors. First, high-affinity interaction occurs
when the fiber knob domain binds to its cognate primary receptor,
the coxsackie and Ad receptor (CAR) (5–8).
Second, cellular integrins interact with Arg-Gly-Asp (RGD) motifs
in the penton base to promote the viral internalization (9,10).
Strategies have been utilized to transductionally
target Ad-based vectors, which include adapter-based approach
(11,12), creation of fiber-chimeric Ad
(13–16) and direct incorporation of peptides
into viral capsid (17–21). The latter has been preferred for
cell-specific Ad retargeting. Fiber proteins (either C terminus or
HI-loop) and hexon are commonly used capsid proteins for the
incorporation of targeting ligands.
In the present study, a new peptide capable of
selectively targeting a human glioblastoma cell line A172 was
identified by direct biopanning of phage-display peptide libraries.
The peptide was then utilized to modify adenoviral hexon, which was
shown to be able to mediate specific adenoviral binding to A172
glioblastoma cells and improve the adenoviral transduction
efficiency.
Materials and methods
Cells and phage libraries
The human glioblastoma cell lines A172, U87 and
human glioma cell line U251 were purchased from the Cell Bank of
the Chinese Academy of Sciences (Shanghai, China); the human
embryonic kidney cell line HEK 293 and lung carcinoma cell line
A549 were obtained from the American Type Culture Collection (ATCC;
Manassas, VA, USA). All the cells were grown in high glucose DMEM
(Gibco, Carlsbad, CA, USA) containing 10% (v/v) newborn calf serum
with 100 U/ml penicillin and 0.1 mg/ml streptomycin. PhD-12 library
displaying 12 random amino acids were obtained from New England
Biolabs (Ipswich, MA, USA).
In vitro selections
BRASIL protocol (22) was optimized and used for our phage
selections. In the subtraction step, HEK 293 cells were detached
and collected with 5 mM EDTA in PBS, then washed with serum-free
DMEM, and re-suspended in 300 μl DMEM containing 1% BSA with
5×106 cells and incubated with 2×1010 pfu
phage within 1.5 ml Eppendorf tubes at 4°C for 3 h with slow
shaking. Then, 300 μl of the cell-phage suspension was centrifuged
at 500 × g for 10 min and the supernatant was incubated with 50 μl
RPMI-1640 medium containing 1% BSA with 5×106 A172 cells
for 3 h at 4°C. Then, the 350 μl cell-phage suspension was gently
transferred to the top of 800 μl non-miscible organic lower phase
[dibutyl phthalate: cyclohexane =9:1 (v:v)] in 1.5 ml Eppendorf
tube and centrifuged at 10,000 × g for 10 min at 4°C. The tube was
snap frozen in liquid nitrogen and the cell-phage pellet at the
bottom of the tube was sliced off and transferred to a new tube
incubated with the host E. coli strain ER2738 to be
recovered, amplified and tittered following the Ph.D. Phage Display
Libraries instruction manual from New England Biolabs. The entire
subtraction/selection cycle was repeated four times.
Immunofluorescence staining
A172 cells were placed in 6-well plates at the
density of 2×105 cells/well. Twenty-four hours later,
culture medium was removed and the cells were rinsed with PBS
twice. Then 2×1011 pfu phages diluted in blocking buffer
[3% (m/v) BSA in serum-free DMEM] were added into every well and
incubated with the cells for 90 min at 4°C. After the incubation,
cells were washed with PBST five times and then fixed by 4%
paraformaldehyde at room temperature for 20 min. Then, the cells
were rinsed three times in PBS for 5 min each. The antibody against
M13 phage was used as the primary antibody followed by the SABC-Cy3
solution. DAPI was used for nuclear counter staining.
Determination of phage binding
efficiency
Phages of 5×109 pfu were incubated with
1×106 cells at 4°C. After elution and titration, the
phage binding efficiency was calculated as number of output
phage/number of input phage.
Specific binding of the phages and cells
by ELISA
A172, U87, A549 and U251 cells (1×104
cells) were plated on 96-well plates, respectively. Twenty-four
hours later, cells were fixed with 4% formaldehyde/PBS for 15 min
and washed three times with PBS. Before cells were incubated with
phages for 1.5 h at 4°C, cells were first blocked with PBS
containing 1% BSA for 30 min at room temperature to avoid
non-specific binding of anti-M13 antibody. Then, the phage bound to
cells was quantified following the normal direct ELISA protocol
using mouse monoclonal antibody to M13 Bacteriophage Coat Protein
g8p (Abcam, Cambridge, MA, USA) as primary antibody, horseradish
peroxidase conjugated-goat anti-mouse IgG antibody (ZSGB-Bio,
Beijing, China) as secondary antibody, ABTS (Shanghai Sangon
Biotechnology, Shanghai, China) as the substrate, and a Multiskan
MK3 microplate reader (Thermo Fisher Scientific, Waltham, MA, USA)
for visualization at 405 nm.
Construction and propagation of
adenoviral vector
The recombinant replication deficient Ad5 vector
used in the present study contains an eGFP expression cassette
driven by human cytomegalovirus intermediate-early promoter in E1
region. The peptide SWDI selected from phage library was inserted
into the HVR5 of hexon protein in the Ad5 capsid by one step
ligation as previously described (21). The nucleotide sequence encoding SWDI
peptide (SWDIAWPPLKVP), flanked by BamHI and SfuI,
was ligated with BamHI and SfuI digested pRGHMAd5.
The obtained plasmid, termed pRGHMAd5-SWDI, was then confirmed by
enzyme digestion and DNA sequencing. The recombinant vector was
linearized by PacI and then transfected into HEK 293 to
package the modified Adenoviral vector Ad5 Hexon-SWDI. The
adenovirus was further propagated in HEK 293 cells and was purified
by cesium chloride gradient methods. The control unmodified
Ad5-eGFP expressing eGFP in E1 region under the control of CMV
promoter was prepared as previously described (17). The virus particle titers were
detected by spectrophotometry at an absorbance (A) of 260 nm.
Adenovirus transduction in vitro
A172, A549, U87 or U251 cells were seeded into a
24-well plate at the density of 5×104 cells/well the day
prior to the infection with Ad5-Hexon-SWDI or Ad5-eGFP at 5,000
vp/cell for A172 cells and 1,000 vp/cell for the other 3 cell
lines. Representative areas were photographed using the inverted
fluorescence microscope at ×200 magnification.
Statistical analysis
Statistical analysis for all experimental conditions
was performed using the Statistical Package for the Social Sciences
(SPSS) software (version 13; SPSS Inc., Chicago, IL, USA) by using
one-way analysis of variance between groups (ANOVA/LSD).
Results
In vitro selection of phages
The level of CAR expression in some glioma cell
lines, such as A172 cells, is low, which is consistent with limited
adenoviral infection observed in our preliminary studies. For the
identification of an alternative binding ligand to improve viral
transduction of A172 cells, we performed panning with PhD-12
library, which consists of randomized linear 12-mer peptides. Phage
clones that could specifically bind to A172 cells were isolated by
4 subtraction/selection rounds. We used A172 for selections, and
HEK 293 cells for subtractions. In each round, the bound phages
were rescued and amplified in ER2738 for the following round of
panning. After four subtraction/selection rounds, an aliquot of the
phage pool was plated out, and plaques were picked for purification
and sequencing. A total of 12 phage clones were chosen randomly
from the final round of biopanning and subjected to DNA sequencing.
The sequences of the displayed peptides are shown in Table I. The phage SWDI that appeared most
frequently was selected as candidate.
 | Table ISequences displayed in individual
phage clones. |
Table I
Sequences displayed in individual
phage clones.
| Phage clone | Consensus
sequences | Frequency |
|---|
| 12-2, 12-4, 12-5,
12-9, 12-11, 12-12 | SWDIAWPPLKVP | 6 |
| 12-1, 12-6,
12-13 | NILPYNKTMLVK | 3 |
| 12-3 | SSSNTTTKHVFT | 1 |
| 12-7 | DSYYTKTERNTH | 1 |
| 12-8 | YEFPRSWDMETN | 1 |
| 12-10 | NRPDSAQFWLHH | 1 |
| 12–14 | GWEVTWPASYAF | 1 |
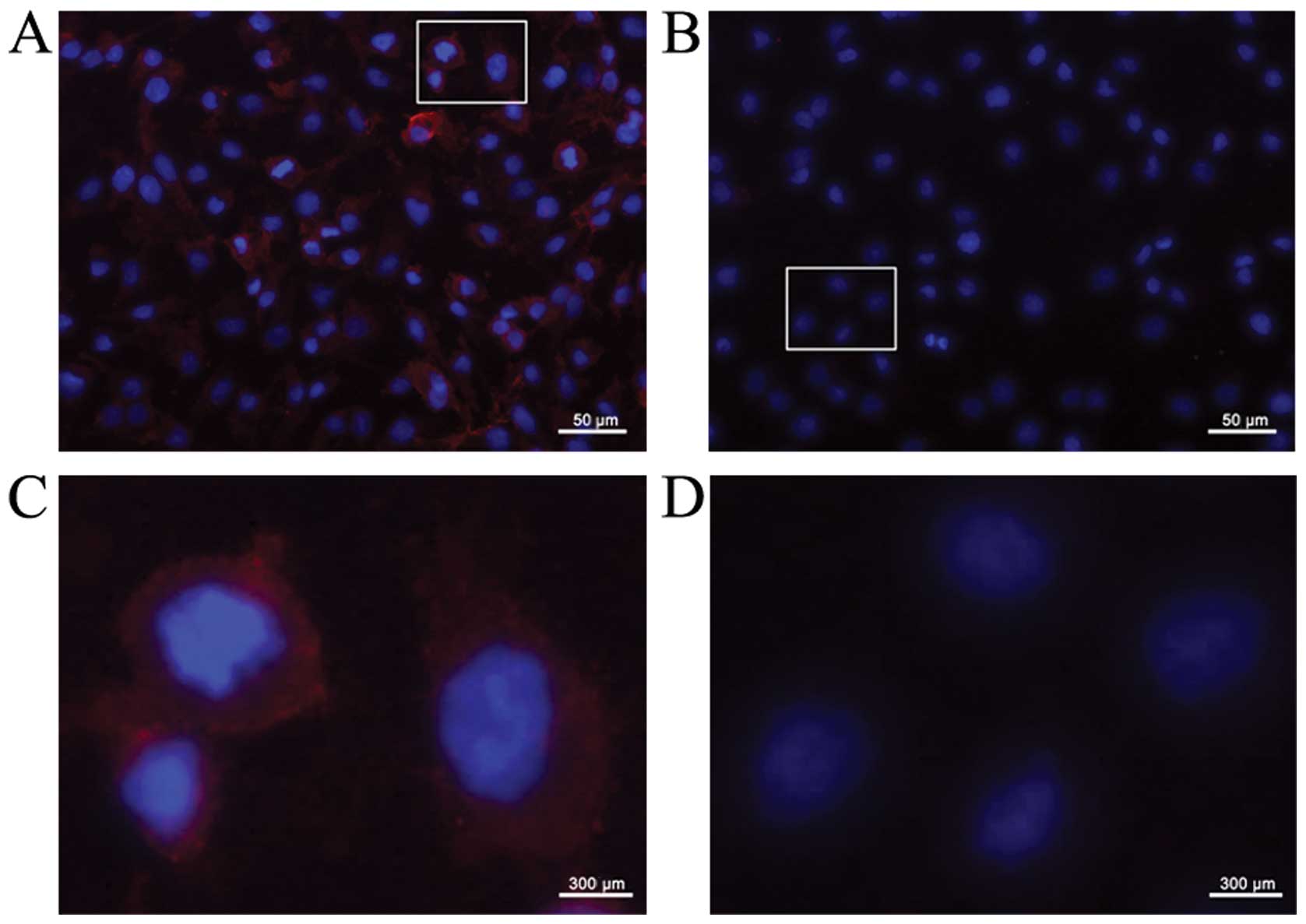
Specific binding of phage SWDI to
A172
Immunofluorescence staining with anti-M13 phage
antibody was performed to observe the specific binding of phage
SWDI to A172 cells. SWDI phages revealed by the red fluorescent
foci were seen on the cell surface of A172 (Fig. 1A and C); however, no fluorescent
signals were present when A172 cells were incubated with the
negative control, phage ASNL, which was picked randomly from phage
library (Fig. 1B and D).
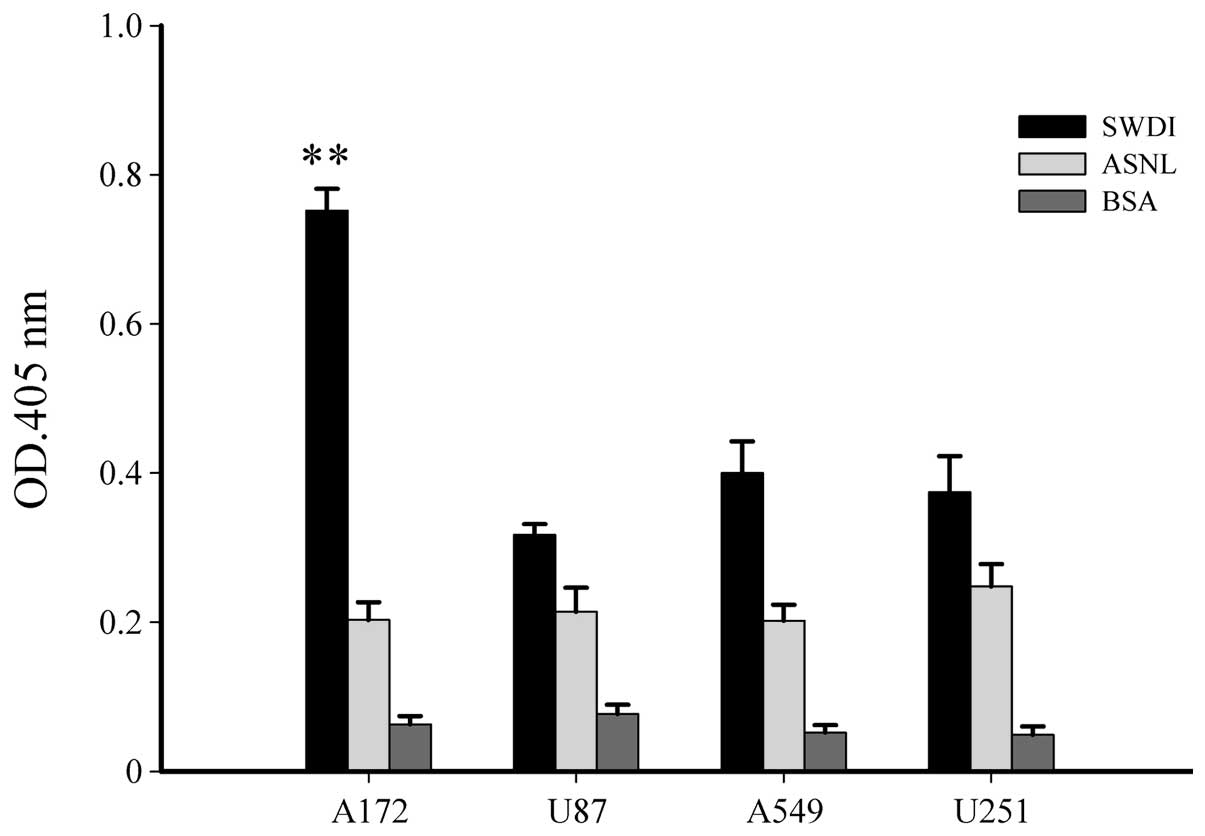
To confirm the specific binding of phage SWDI to
A172 cells, ELISA was performed using anti-M13 antibody. In
addition to A172 cells, two other human glioblastoma cell lines,
U87 and U251, and human lung carcinoma cell line A549 were used as
the control cell lines. As shown in Fig. 2, the binding activity of phage SWDI
to A172 cells was 2.7-fold higher than that of phage ASNL to A172
(P<0.01). In U87, U251 and A549 cells, the binding activity of
phage SWDI showed no significant difference when compared to that
of ASNL.
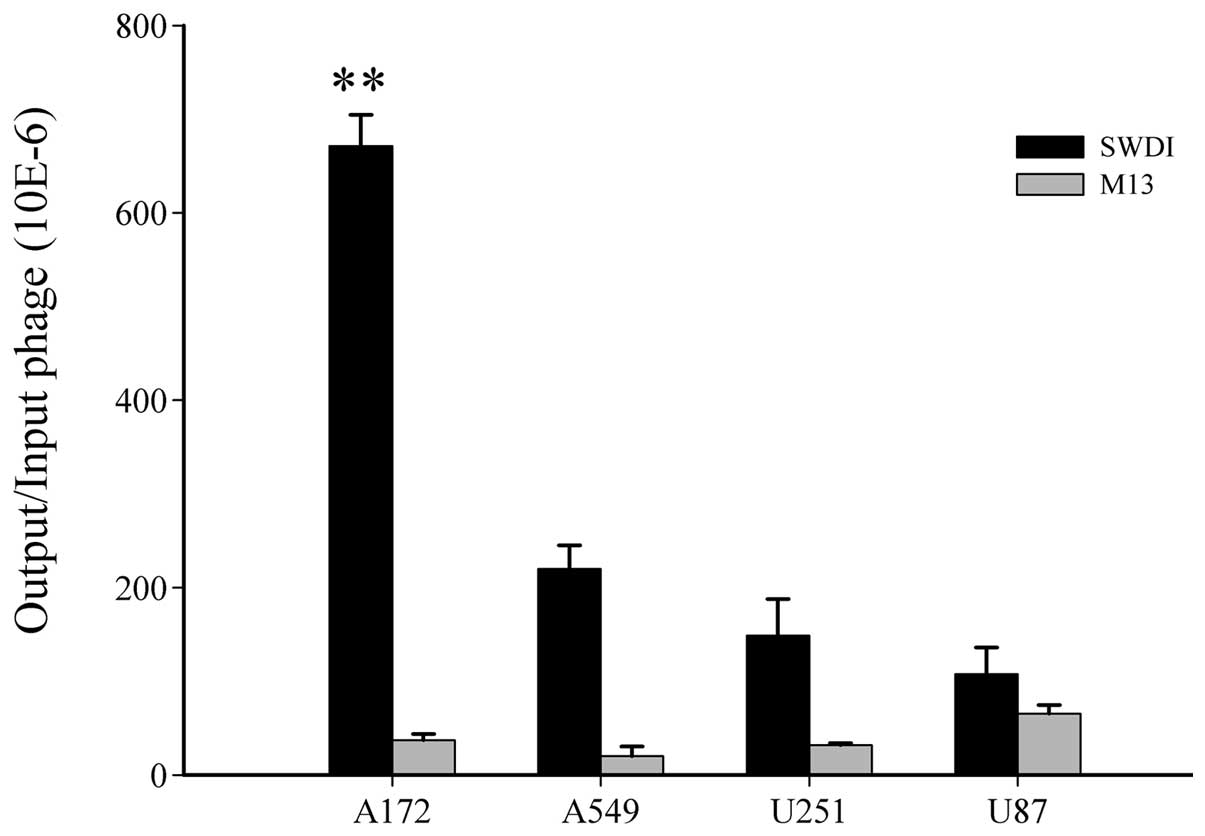
Then, phage SWDI was incubated with A172 cells
(5×103 pfu/cell) at 4°C to further evaluate the phage
binding efficiency, which was calculated as number of output
phage/number of input phage (Fig.
3). In A172 cells, the binding efficiency of phage SWDI was
~13-fold higher than that of the negative control wild-type M13
phage (P<0.01). There was no significant difference in binding
efficiency between phage SWDI and M13 in other cell lines.
Generation of adenovirus with SWDI
modified hexon
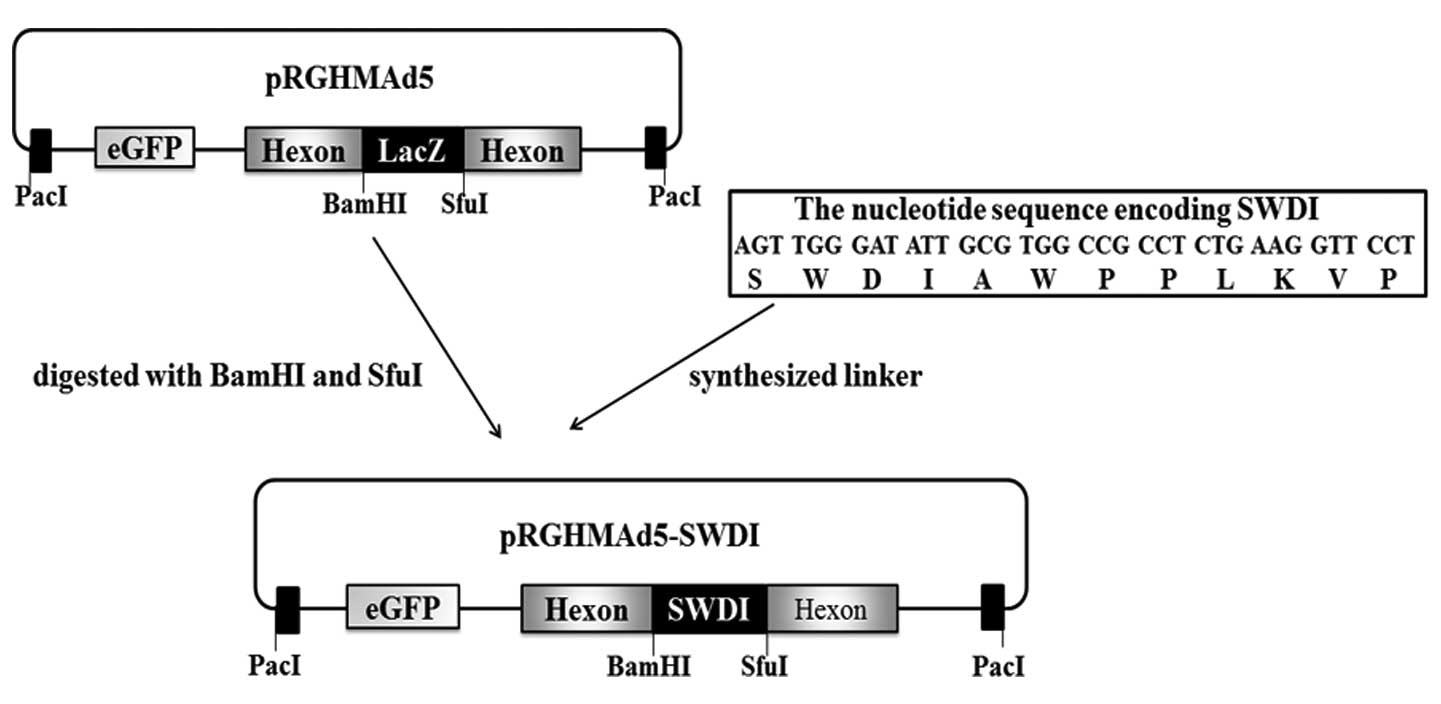
We utilized our previously constructed shuttle
vector pRGHMAd5 (21), which
contains a lacZ expression cassette in the hexon HVR5 region and an
eGFP expression cassette driven by the CMV promoter in E1 region
(Fig. 4), to incorporate peptide
SWDI into the HVR5 of the Ad5 hexon. Peptide SWDI was ligated to
the BamHI and SfuI digested pRGHMAd5. The obtained
plasmid, termed pRGHMAd5-SWDI (Fig.
4), was confirmed by the restriction endonuclease analysis and
DNA sequencing. Then, adenovirus with SWDI modified-hexon was
successfully generated by transfecting PacI-linearized
pRGHMAd5-SWDI into HEK 293 cells. The obtained virus was termed
Ad5-Hexon-SWDI. The virus was produced with a titer of
2.8×1011 vp/ml and the titer of the control virus
Ad5-eGFP is 2.2×1012 vp/ml.
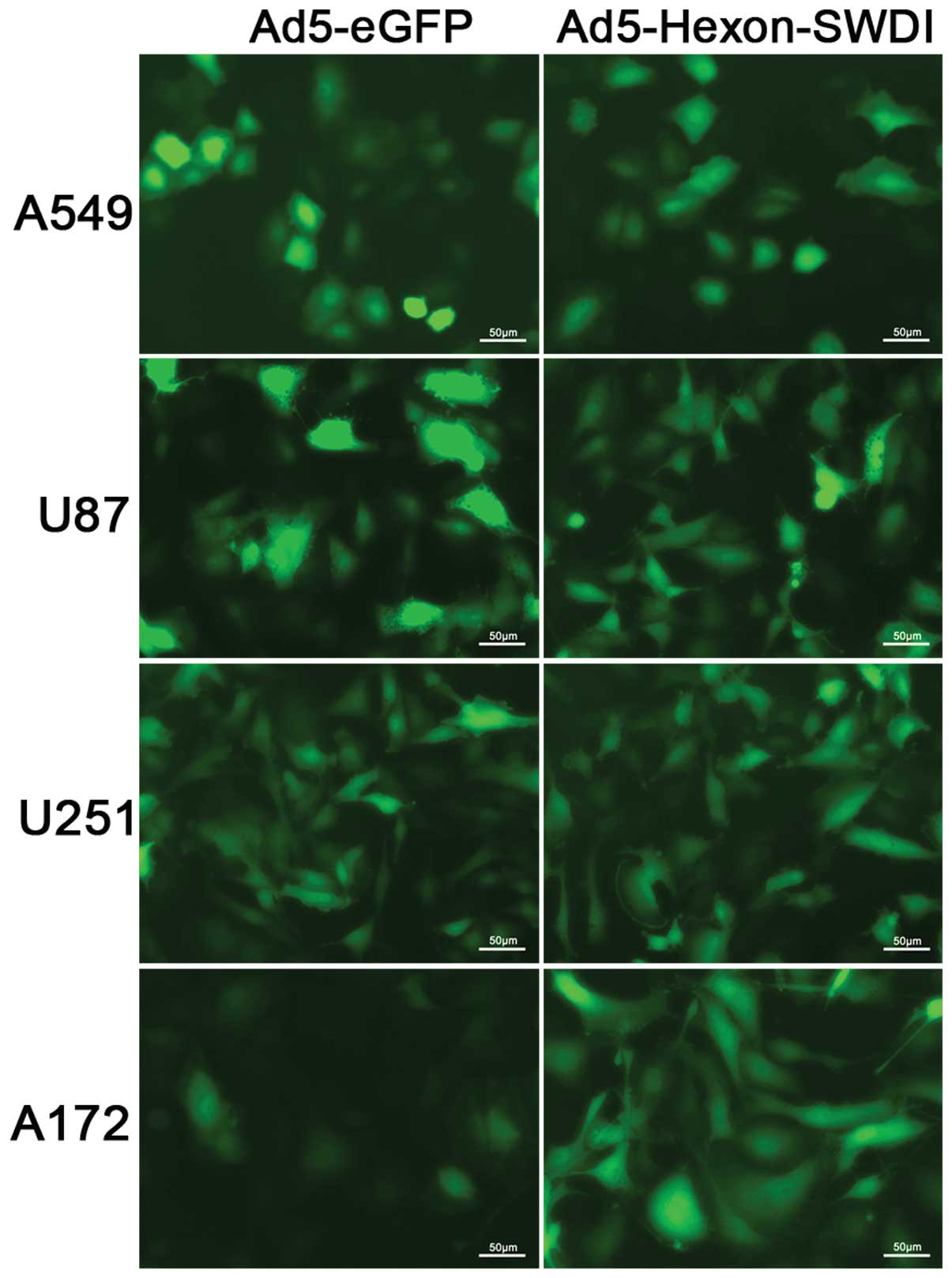
Transduction efficiency of adenovirus
with SWDI-modified hexon
To examine the transduction efficiency of
SWDI-modified Ad, A172 and control cell lines including A549, U251
and U87 were infected with Ad5-Hexon-SWDI and control virus,
Ad5-eGFP. Fig. 5 shows that the
eGFP expression level in A172 infected with Ad5-Hexon-SWDI was
higher than that in A172 infected with Ad5-eGFP. In each control
cell line, no apparent difference in eGFP expression was observed
between cells infected with Ad5-Hexon-SWDI and Ad5-eGFP. The
results indicated that SWDI modification of hexon protein could
improve the adenoviral transduction in CAR-deficient A172 cells
specifically. GFP expression quantification by FACS revealed that
GFP positivity ratio was higher in A172 treated by Ad5-Hexon-SWDI
(86.8±1.3%) than that in cells treated by Ad5-eGFP (34.3±1.7%)
(P<0.01).
Discussion
In the present study, a new peptide SWDIAWPPLKVP,
capable of selectively targeting a human glioblastoma cell line
A172, was identified by direct biopanning of phage-display peptide
libraries. The binding activity of SWDIAWPPLKVP peptide in A172 was
more than 10-fold higher than that of the control phage. In
addition, modification of hexon protein with this peptide could
improve adenoviral infectivity in A172 cells.
Gene therapy is being studied in clinical trials for
different types of cancer (23,24).
The features of Ad, such as high viral titer, high transduction
efficiency for both proliferating and quiescent cells, large
insertional capacity for foreign genes and high safety profile,
make it well suited for cancer gene therapy (1). However, the progress of the
application of Ad in cancer gene therapy is hindered by the lack of
the expression of native adenovirus receptor on a variety of cancer
types. To enable both efficient transduction of target cells and to
avoid ectopic infection of non-target cells, strategies are needed
to retarget the Ad vector to non-native cellular surface
receptors.
A number of strategies have been developed,
primarily based on the use of bispecific antibodies (25,26),
creation of fiber-chimeric Ad (13–16)
and direct incorporation into viral capsid of peptide analogues
specific for a tumor-associated receptor (17–21).
The latter has been preferred for cancer cell-specific Ad
retargeting, in which finding new cancer targeting peptides is
crucial. On this path, phage display has been shown to be a
powerful tool in identifying specific ligands on target cancer
cells, such as glycine-serine-leucine (GSL) of breast carcinomas
(27), asparagine-glycine-arginine
(NGR) of tumor neovasculature (28,29),
and HEWSYLAPYPWF of colorectal cancer (30).
The fiber is the most commonly used capsid protein
for retargeting Ad. Studies have shown that this location can be
modified without interfering with capsid assembly (31). Several groups have successfully
inserted cellular-specific targeting peptides into the HI loop of
adenovirus fiber protein (17–19).
Notably, in the present study, the Ad failed to propagate when
peptide SWDIAWPPLKVP was inserted into the fiber HI-loop.
Alternatively, we modified another capsid protein, hexon, within
which the hypervariable regions (HVR) have solvent exposed loops,
an ideal location for modification. Previous studies showed that
genetic modification of HVR with integrin-binding RGD motif
(32) had no effect on hexon
structure or capsid stability but increased CAR-independent viral
transduction. We took advantage of the adenoviral vector system for
rapid modification of hexon protein, which was described in our
previous study (21), and
successfully generated a modified Ad with SWDI peptide inserted in
hexon. The SWDI peptide-modified Ad was produced with decent titer,
indicating that the viral assembly was not interfered by this
modification. In addition, this peptide modification of Ad improved
cell-specific adenoviral transduction.
In conclusion, these findings demonstrated that a
peptide acquired through phage display can mediate cell-specific Ad
retargeting when inserted into Ad hexon, suggesting an approach for
targeting adenoviral infection to specific cancer cells. In the
future, in vivo studies will be carried out to confirm the
retargeting ability of the modified Ad in a glioblastoma animal
model.
Acknowledgements
This study was supported by the Fundamental Research
Funds for the Central Universities (GK201104004 and GK201301010),
the Innovation Funds of Graduate Programs, Shaanxi Normal
University (2011CXB006) and the National Natural Science Foundation
of China (no. 81272543).
References
|
1
|
Breyer B, Jiang W, Cheng H, et al:
Adenoviral vector-mediated gene transfer for human gene therapy.
Curr Gene Ther. 1:149–162. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kim M, Zinn KRB, Barnett G, et al: The
therapeutic efficacy of adenoviral vectors for cancer gene therapy
is limited by a low level of primary adenovirus receptors on tumour
cells. Eur J Cancer. 38:1917–1926. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wickham TJ, Roelvink PW, Brough, et al:
Adenovirus targeted to heparan-containing receptors increases its
gene delivery efficiency to multiple cell types. Nat Biotechnol.
14:1570–1573. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Krasnykh VN, Mikheeva GV, Douglas JT, et
al: Generation of recombinant adenovirus vectors with modified
fibers for altering viral tropism. J Virol. 70:6839–6846.
1996.PubMed/NCBI
|
|
5
|
Tomko RP, Xu R and Philipson L: HCAR and
MCAR: the human and mouse cellular receptors for subgroup C
adenoviruses and group B coxsackieviruses. Proc Natl Acad Sci USA.
94:3352–3356. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bergelson JM, Cunningham JA, Droguett G,
et al: Isolation of a common receptor for coxsackie B viruses and
adenoviruses 2 and 5. Science. 275:1320–1323. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bergelson JM, Krithivas A, Celi L, et al:
The murine CAR homolog is a receptor for coxsackie B viruses and
adenoviruses. J Virol. 72:415–419. 1998.PubMed/NCBI
|
|
8
|
Roelvink PW, Lizonova A, Lee JG, et al:
The coxsackievirus-adenovirus receptor protein can function as a
cellular attachment protein for adenovirus serotypes from subgroups
A, C, D, E and F. J Virol. 72:7909–7915. 1998.PubMed/NCBI
|
|
9
|
Wickham TJ, Matthias P, Cheresh DA, et al:
Integrins αvβ3 and αvβ5
promote adenovirus internalization but not virus attachment. Cell.
73:309–319. 1993.
|
|
10
|
Nemerow GR, Pache L, Reddy V and Stewart
PL: Insights into adenovirus host cell interactions from structural
studies. Virology. 384:380–388. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Eto Y, Gao JQ, Sekiguchi F, et al:
PEGylated adenovirus vectors containing RGD peptides on the tip of
PEG show high transduction efficiency and antibody evasion ability.
J Gene Med. 7:604–612. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wortmann A, Vöhringer S, Engler T, et al:
Fully detargeted polyethylene glycol-coated adenovirus vectors are
potent genetic vaccines and escape from pre-existing
anti-adenovirus antibodies. Mol Ther. 16:154–162. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sakurai F, Mizuguchi H and Hayakawa T:
Efficient gene transfer into human CD34+ cells by an
adenovirus type 35 vector. Gene Ther. 10:1041–1048. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wohlfahrt ME, Beard BC, Lieber A and Kiem
HP: A capsid-modified, conditionally replicating oncolytic
adenovirus vector expressing TRAIL leads to enhanced cancer cell
killing in human glioblastoma models. Cancer Res. 67:8783–8790.
2007. View Article : Google Scholar
|
|
15
|
Havenga MJ, Lemckert AA, Grimbergen JM, et
al: Improved adenovirus vectors for infection of cardiovascular
tissues. J Virol. 75:3335–3342. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Goossens PH, Havenga MJ, Pieterman E, et
al: Infection efficiency of type 5 adenoviral vectors in synovial
tissue can be enhanced with a type 16 fiber. Arthritis Rheum.
44:570–577. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xia H, Anderson B, Mao Q and Davidson BL:
Recombinant human adenovirus: targeting to the human transferrin
receptor improves gene transfer to brain microcapillary
endothelium. J Virol. 74:11359–11366. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rittner K, Schreiber V, Erbs P and Lusky
M: Targeting of adenovirus vectors carrying a tumor cell-specific
peptide: in vitro and in vivo studies. Cancer Gene
Ther. 14:509–518. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jullienne B, Vigant F, Muth E, et al:
Efficient delivery of angiostatin K1–5 into tumors following
insertion of an NGR peptide into adenovirus capsid. Gene Ther.
16:1405–1415. 2009.PubMed/NCBI
|
|
20
|
Liu S, Mao Q, Zhang W, et al: Genetically
modified adenoviral vector with the protein transduction domain of
Tat improves gene transfer to CAR-deficient cells. Biosci Rep.
29:103–109. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Di B, Mao Q, Zhao J, et al: A rapid
generation of adenovirus vector with a genetic modification in
hexon protein. J Biotechnol. 157:373–378. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Giordano RJ, Cardó-Vila M, Lahdenranta J,
et al: Biopanning and rapid analysis of selective interactive
ligands. Nat Med. 7:1249–1253. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Alexander BL, Ali RR, Alton EW, et al:
Progress and prospects: gene therapy clinical trials (part 1). Gene
Ther. 14:1439–1447. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Aiuti A, Bachoud-Lévi AC, Blesch A, et al:
Progress and prospects: gene therapy clinical trials (part 2). Gene
Ther. 14:1555–1563. 2007. View Article : Google Scholar
|
|
25
|
Volpers C, Thirion C, Biermann V, et al:
Antibody-mediated targeting of an adenovirus vector modified to
contain a synthetic immunoglobulin g-binding domain in the capsid.
J Virol. 77:2093–2104. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Haisma HJ, Grill J, Curiel DT, et al:
Targeting of adenoviral vectors through a bispecific single-chain
antibody. Cancer Gene Ther. 7:901–904. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Arap W, Pasqualini R and Ruoslahti E:
Cancer treatment by targeted drug delivery to tumor vasculature in
a mouse model. Science. 279:377–380. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Curnis F, Arrigoni G, Sacchi A, et al:
Differential binding of drugs containing the NGR motif to CD13
isoforms in tumor vessels, epithelia, and myeloid cells. Cancer
Res. 62:867–874. 2002.PubMed/NCBI
|
|
29
|
Pasqualini R, Koivunen E, Kain R, et al:
Aminopeptidase N is a receptor for tumor-homing peptides and a
target for inhibiting angiogenesis. Cancer Res. 60:722–727.
2000.PubMed/NCBI
|
|
30
|
Rasmussen UB, Schreiber V, Schultz H, et
al: Tumor cell-targeting by phage-displayed peptides. Cancer Gene
Ther. 9:606–612. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Belousova N, Krendelchtchikova V, Curiel
DT and Krasnykh V: Modulation of adenovirus vector tropism via
incorporation of polypeptide ligands into the fiber protein. J
Virol. 76:8621–8631. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Vigne E, Mahfouz I, Dedieu JF, et al: RGD
inclusion in the hexon monomer provides adenovirus type 5-based
vectors with a fiber knob-independent pathway for infection. J
Virol. 73:5156–5161. 1999.PubMed/NCBI
|