Introduction
Epidermal growth factor receptor (EGFR) is a
membrane tyrosine kinase receptor that belongs to the EGF tyrosine
kinase receptor family consisting of EGFR (ErbB1), Her2 (ErbB2),
ErbB3 and ErbB4 (1,2). EGFR monomer is composed of four
domains (3), namely, extracellular,
transmembrane, intracellular domain (including kinase domain) and
cytoplasmic tail (4,5). According to the literature, EGF or
other growth factors can cause EGFR to change its extracellular
domain structure and then trigger dimerization of intracellular
domain (6–13). In response to growth factors, EGFR
forms either homodimer or heterodimer with other family members,
therefore it subsequently changes its protein structure and protein
interaction with numerous intracellular proteins (14–17).
EGFR is subject to autophosphorylation and/or transphosphorylation
that are essential processes for EGFR to become an active molecule
and play important roles in regulating several signaling pathways,
such as MAPK, PI3-Akt and JAK-Stat 3/5 pathways (18–20).
There are several critical tyrosine phosphorylation sites in the
intracellular domain of EGFR. For example, Y845, 992, 1045, 1068,
1086 and 1173. These differential phosphorylation sites correspond
to different signal pathways and functions (19). Generally, Y845 is associated with
the Stat signaling pathway. Y992, 1068, 1086 and 1173 are generally
linked to MAPK and Akt signaling pathways (19), while Y1045 phosphorylation is
associated with the interaction with cbl that regulates EGFR
ubiquitination and degradation (21). However, what determines these
phosphorylation sites and how an individual site is regulated over
the entire process remain unclear.
Although EGFR is one of the most intensively studied
receptor tyrosine kinases (RTKs), it remains elusive as to whether
or not its intra-tyrosine phosphorylations are interrelated. As a
cell surface molecule, EGFR plays an essential and fundamental role
in dictating cell proliferation and differentiation, cell cycle
control, biological development, tumorigenesis and malignant
development (22–24), therefore it is necessary to
extensively elucidate its activation and interplay among EGFR
intracellular tyrosine phosphorylations in various cellular
environments. As a traditionally accepted concept, upon ligand
stimulation, EGFR forms homodimer or heterodimer with one of the
other three family members in physiological conditions, which
subsequently results in phosphorylation of EGFR by either Src
kinase or autophosphorylation. However, previous studies
demonstrated that high local EGFR concentration may force EGFR to
form dimers even in the absence of ligand stimulation (14). The phosphorylated EGFR provides
docking sites for binding downstream adaptor proteins and
thereafter activates several downstream signaling pathways. Upon
Src-induced phosphorylation at Y845 on EGFR, the phosphorylated
Y845 serves as docking site to recruit Stat3/5 and result in
phosphorylation of Stat3 and/or Stat5, which are transcription
factors and can form homo- or heterodimers. The dimerized Stat3 or
Stat5 translocates into the nucleus and regulates cell
proliferation, differentiation, cell cycle and migration.
Src-activated signal pathway through Y845 is typically considered a
transphosphorylation pathway. EGF ligand stimulation also causes an
autophosphorylation of EGFR. Several tyrosine residues in
intracellular domain of EGFR are involved in this type of
autophosphorylation such as Y992, 1068, 1086 and 1173 and provide
docking sites for adaptor proteins such as Shc, Grb2 and Gab that
result in the activation of PI3K/Akt and Ras/MAPK signaling
pathways (19). The activation of
PI3K/Akt and/or Ras/MAPK pathways has been linked to various types
of cancer (22,25,26).
Depending on specific cell types or environments, one of these
pathways may dominate or all of these pathways equally contribute
to cellular processes. Despite these advances, the interplay
between transphosphorylation and autophosphorylation of EGFR
remains elusive.
Here, we report that transphosphorylation of EGFR at
Y845 is linked to its autophosphorylation and kinase activity. The
substitution of Y845 to phenylalanine (F) significantly reduced its
biological function in response to ligand stimulation, suggesting
the importance of this transphosphorylation site. Taken together,
our results provide insights into the activation of EGFR and may
indicate a potential therapeutic target for treating various types
of cancer related to aberrant EFGR expression and activation.
Materials and methods
Cell lines, antibodies and chemicals
HEK293 and MCF7 cells were cultured with Dulbecco’s
modified Eagle’s medium (DMEM) containing 10% fetal bovine serum
(FBS) and appropriate antibiotics in an incubator with 37°C and 5%
CO2. Antibodies were purchased as follows: anti-EFGR
(Santa Cruz Biotechnology, Santa Cruz, CA, USA);
anti-phospho-tyrosine (4G10; Upstate); anti-phospho-EGFR-Y845 and
phospho-EGFR-Y1068 (Cell Signaling Technology Inc.). All chemicals
were purchased from Sigma-Aldrich Corporation and are of analytical
grade except where noted otherwise.
Immunoprecipitation and
immunoblotting
The cells were lysed in RIPA buffer containing 2 mM
PMSF, 2 mM Na3VO4, 2 mM NaF, 1 μg/ml
aprotinin, leupeptin and pepstatin, respectively. For
immunoprecipitation, a total of 500 μg of cell lysate was used and
diluted in 500 μl of RIPA buffer with the corresponding antibodies.
After addition of 2 μg EGFR or flag antibody, the lysate was
incubated with gentle rotation for 2 h at 4°C, and then 80 μl of
protein-A and 20 μl of protein-G beads were added into the mixture
and incubated for another 2 h. For immunoblotting, a total of 20 μg
of cell lysate was directly loaded and resolved onto 8%
SDS-polyacrylamide gels and then probed with antibodies as
indicated in the images.
In vitro kinase assay
After immunoprecipitation, the beads were washed
twice using RIPA buffer containing appropriate inhibitors. After
removing wash buffer from last step wash, we added 50 μl 2X kinase
buffer containing 100 mM HEPES (pH 7.2), 20 mM MgCl2, 20
mM MnCl2, 6 mM Na3VO4, and 2.5 mM
DTT, and 0.5 μl γ-32p-ATP, mixed the reaction mixture
well with beads and incubated in a 30°C water bath for 25 min, then
added 20 μl 6X loading buffer followed by heating of the samples
for 5 min at 100°C. We briefly centrifuged the samples and the
supernatants were used for analysis.
For in vitro transphosphorylation assay,
after immunoprecipitating flag tagged EGFR-K721A with an anti-flag
antibody from 1,000 μg HEK293 cell lysate and washing twice with
RIPA buffer, we added 50 μl 2X kinase buffer containing 100 mM
HEPES (pH 7.2), 20 mM MgCl2, 20 mM MnCl2, 6
mM Na3VO4 and 2.5 mM DTT and 0.5 mM ATP and
then mixed with 10, 30 and 100 μg cell lysates from HEK293 cells
expressing myc-tagged either wt-EGFR or EGFR-Y845F or K721A mutant,
respectively. After incubation in a 30°C water bath for 25 min, we
washed the beads twice using RIPA buffer, then added 20 μl 6X
loading buffer followed by heating of the samples for 5 min at
100°C. The final elutants were resolved onto 8% SDS-ployacrylamide
gels and then probed with anti-phospho antibodies as indicated in
the images.
Transfection and establishment of stably
expressing EGFR cell clones
Transfection of wild-type EGFR (wt-EGFR) or
EGFR-Y845F or EGFR-Y721A was performed using Lipofectamine 2000
(Life Technologies Corporation) according to the standard
procedures and the manufacturer’s instructions. For establishing
stable clones expressing EGFR or mutants, after 48 h transfection,
the cells were selected with 6 μg/ml blasticidin (Life Technologies
Corporation) for 3 weeks and single clones were isolated, and
continued to culture for another 2–3 weeks in the presence of
blasticidin. The cloned cells were analyzed for EGFR expression and
other analysis.
Cell growth assay
Cells (5×104) in 1 ml DMEM were seeded in
6-well plates. The cells were cultured in the absence or presence
of EGF and cell number was counted by addition of trypan blue at
time points indicated in the image. All results are in triplicate
from three independent experiments.
In vitro DNA incorporation assay
For measuring cell DNA synthesis, 1×105
MCF7 cells stably expressing either wt-EGFR or EGFR-Y845F mutant
were seeded in 6-well plates. After 30 h serum starvation, the
cells were treated with 30 μM BrdU (BD transduction) in 10% FBS
fresh medium with or without 25 ng/ml EGF for 16 h, then an
anti-BrdU monoclonal antibody (Upstate) was used to detect
incorporated BrdU substrate in the cells. An FITC-conjugated goat
anti-mouse IgG antibody (Johnson Laboratory, USA) was used to
amplify signals. Then, DNA incorporated cells were counted by using
a fluorescence microscope. At least three independent experiments
were performed as each experiment was set up in triplicate.
In vivo mammary fat pad tumor cell
inoculation
MCF7 (5×106) cells stably expressing
either wt-EGFR or EGFR-Y845F mutant were injected into the fat pads
of nude mice. The tumor size was measured once per week. All animal
handling procedures were performed in accordance with the
institutional policies and regulations.
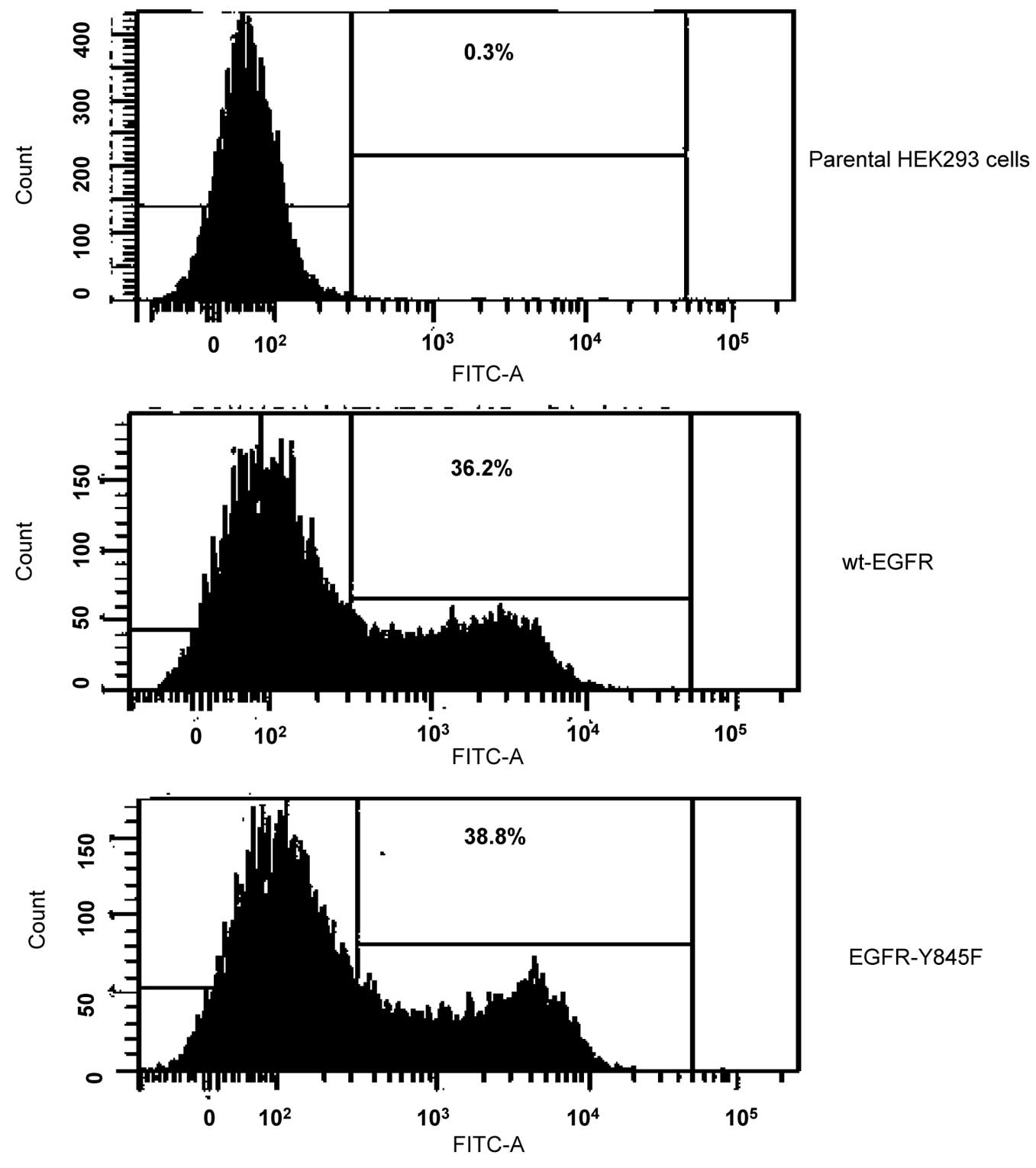
Flow cytometry analysis
Either wt-EGFR or EGFR-Y845F was transiently
expressed in HEK293 cells. The cells were incubated with an
anti-EGFR antibody for 1 h on ice, followed by incubation with an
FITC labeled anti-IgG antibody for 30 min on ice, and then
subjected to flow cytometry analysis.
Statistical analysis
All data are described as means ± standard
deviation. Statistical analyses were performed by Student’s t-test
analysis. P<0.05 was considered to indicate a statistically
significant difference.
Results
Transphosphorylation of EGFR at Y845
affects its autophosphorylation
To determine if there is a correlation between EGFR
transphosphorylation and autophosphorylation, we established stable
cell clones that express either wild-type EGFR (wt-EGFR) or
EGFR-Y845F mutant in an MCF7 breast carcinoma and an HEK293 cell
line. Expression of target protein in both cell lines was confirmed
by western blotting (Fig. 1A and
B). To test if transphosphorylation deficient EGFR-Y845F has an
impact on its autophosphorylation, the cloned cells were serum
starved for 24 h and then stimulated by addition of EGF. The
results demonstrated that in the absence of EGF the levels of
autophosphorylation at Y1068 and total phosphorylation were close
to each other in both cell clones. However, in the presence of EGF,
the phosphorylation at Y1068 and total phosphorylation were
significantly reduced by EGFR-Y845F mutant, suggesting that
EGFR-Y845 phosphorylation is important for its EGF-induced
autophosphorylation (Fig. 1C and
D).
The transphosphorylation deficient
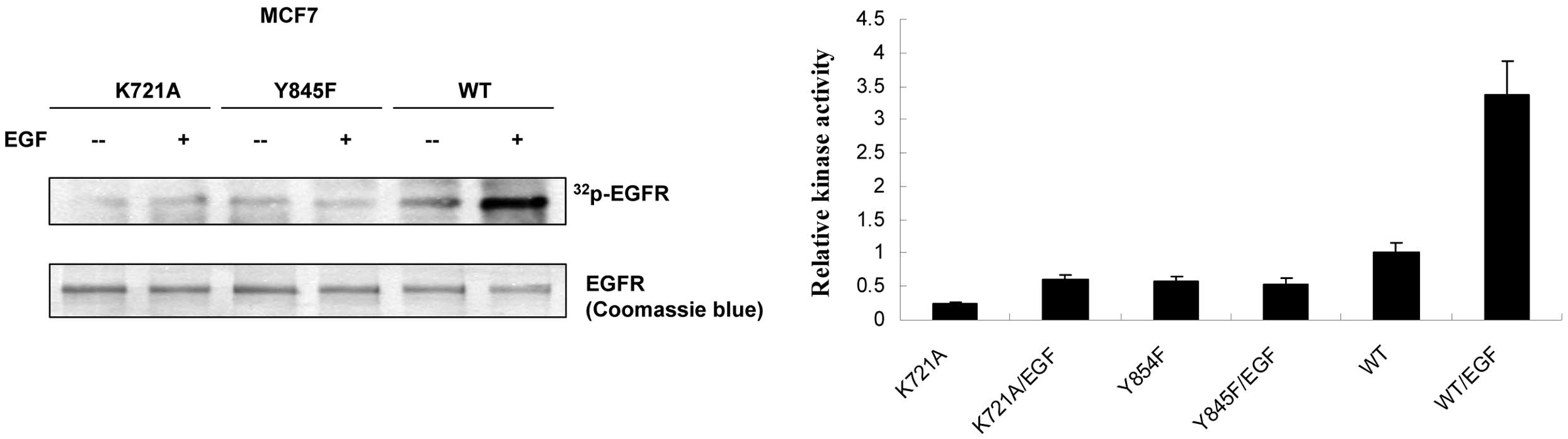
EGFR-Y845F mutant reduces its ligand-induced kinase activity
To determine if hindrance of EGF-induced
phosphorylation by EGFR-Y845F mutant links to mitigation of its
kinase activity, in vitro kinase assay was performed as
described in Materials and methods. MCF7 clone cells were treated
with EGF before harvesting. In comparison to wt-EGFR, EGFR-Y845F
significantly reduced its EGF-induced kinase activity (Fig. 2), suggesting that loss of
EGF-induced transphosphorylation indeed reduced its kinase activity
in EGFR-Y845F mutant.
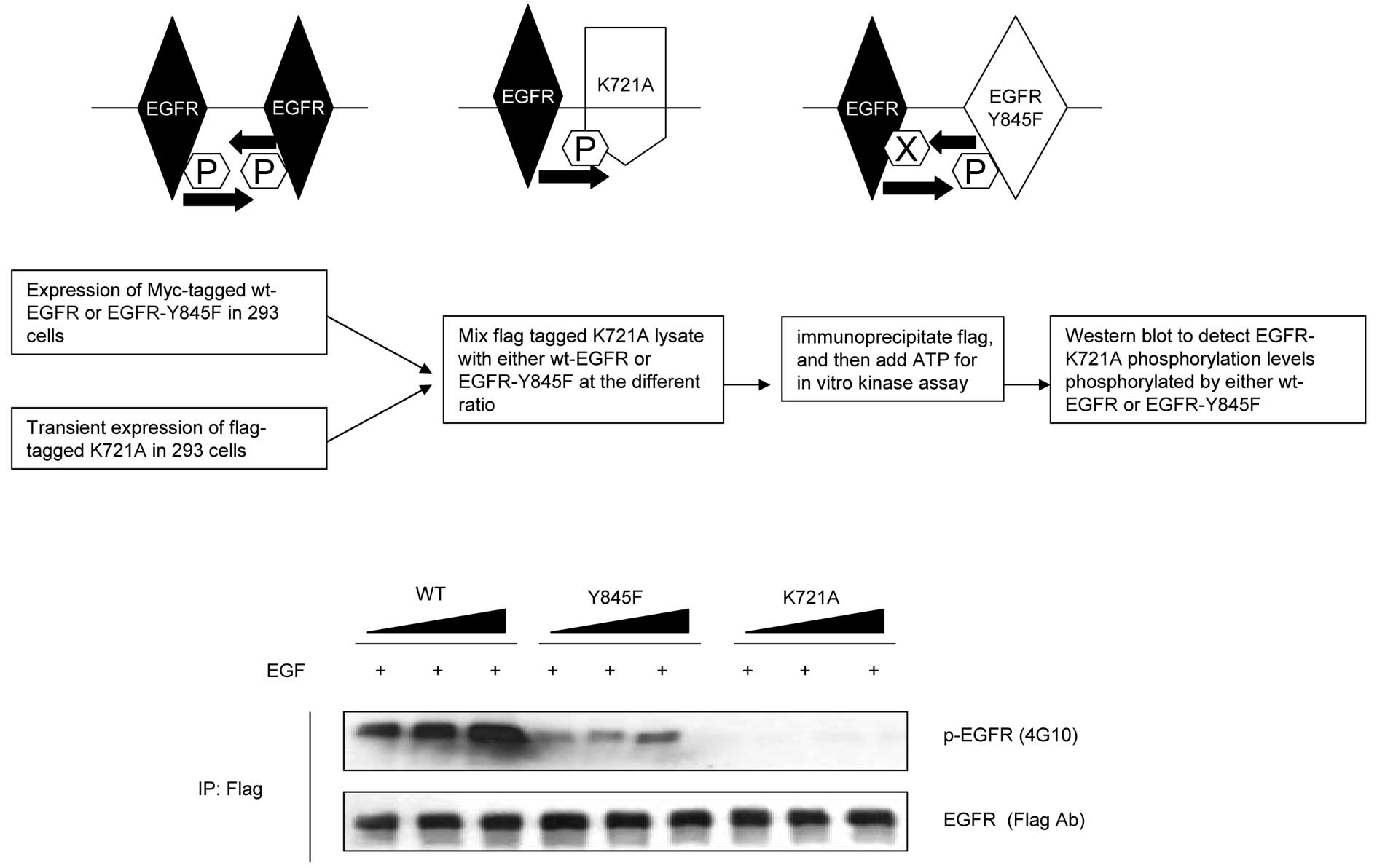
The transphosphorylation deficient
EGFR-Y845F mutant reduces its ability to phosphorylate its dimeric
partners
Upon the initiation of its activity, EGFR
autophosphorylation occurs by cross phosphorylating its dimeric
counterparts. To further distinguish if the loss of EGFR
autophosphorylation in EGFR-Y845F mutant was the consequence of the
loss of its transphosphorylation, we performed in vitro
kinase assay by mixing different amounts of either wt-EGFR or
EGFR-Y845F mutant with flag tagged EGFR-K721A, which is a kinase
dead mutant and loses its autophosphorylation ability to its
dimeric counterparts (Fig. 3). We
were thus able to discriminate if EGFR-Y845F mutant may reduce its
auto phosphorylation potential. Our results showed that EGFR-Y845F
mutant considerably reduced its ability of EGF-induced
phosphorylation to its dimeric counterparts, suggesting the
importance of Y845 transphosphorylation in this process.
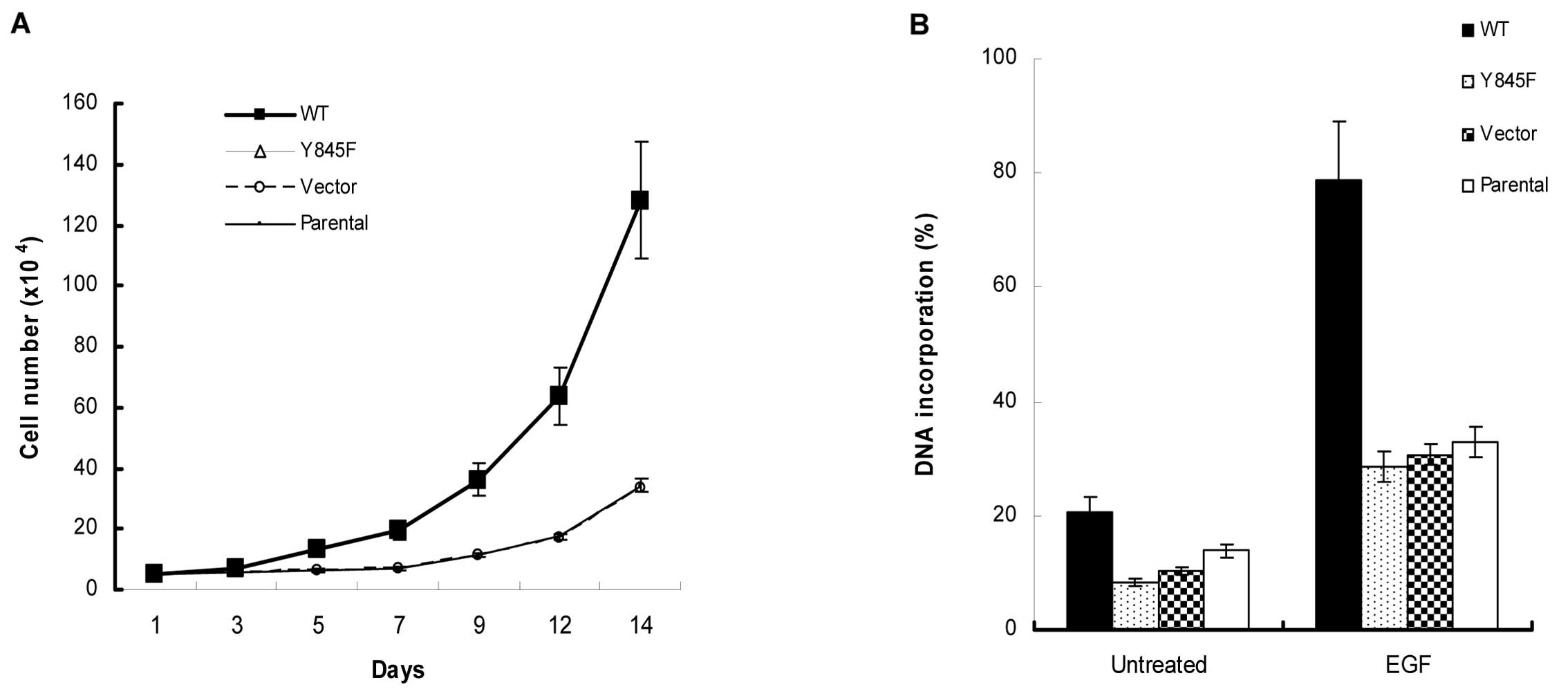
The EGFR-Y845F mutant reduces its ability
to stimulate cell growth, DNA synthesis and EGFR-mediated tumor
growth
To determine the functional consequences of the loss
of Y845 transphosphorylation, we used MCF7 stable clone cells to
perform functional assays. The results showed that EGFR-Y845F
mutant reduced its ability to respond to EFG-stimulated cell growth
and DNA synthesis (Fig. 4).
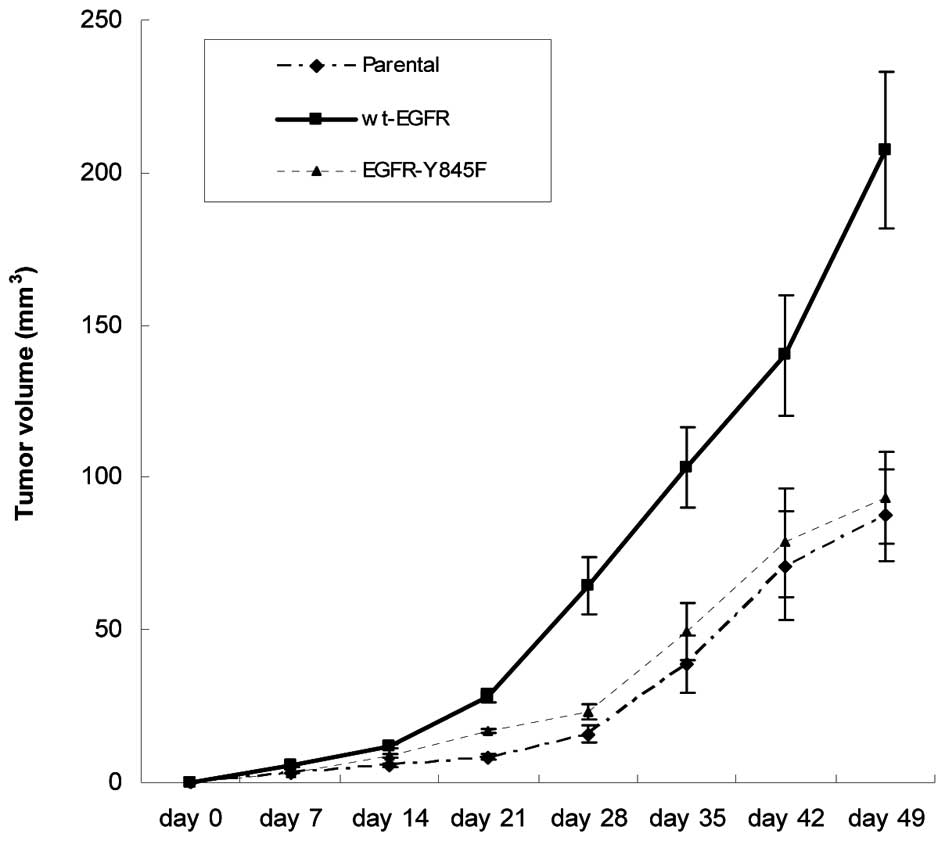
Furthermore, EGFR-Y845 mutant significantly reduced EGFR-mediated
tumor growth in nude mice (Fig. 5),
suggesting that transphosphorylation of EGFR-Y845 is critical to
its biological function.
Discussion
In the present study, we clearly demonstrated the
intrinsic correlation between EGFR transphosphorylation and
autophosphorylation. Our results support that transphosphorylation
of EGFR at Y845, which is located in its activation loop and a
critical tyrosine transphosphorylation site, potentially influences
its kinase activity and biological function. Upon the replacement
of tyrosine to phosphorylation incapable phenylalanine, we observed
that EGFR autophosphorylation and ligand-induced kinase activity
were significantly reduced. Meanwhile, the ability of the
EGFR-Y845F mutant responding to EGF ligand stimulation was severely
impaired (Figs. 3 and 4). The functional results are also
consistent with this alteration as to which EGF-induced cell
growth, DNA synthesis and tumor growth in nude mice were impaired
as well (Figs. 4 and 5). A previous study demonstrated that
EGFR-Y845 phosphorylation was essential for ligand-stimulated DNA
synthesis. However, EGFR autophosphorylation seemed independent of
its transphosphorylation at Y845 (27). In contrast to this report, one study
demonstrated that EGFR-Y845F mutant actually augmented both
ligand-stimulated EGFR tyrosine phosphorylation and DNA synthesis
(28). We consider that these
inconsistent observations may result from different cell types and
EGFR expression levels within the underlying cell lines. In
addition, we observed that massively transient expression of EGFR
in HEK293 cells, EGFR could be overphosphorylated in the absence of
EGF ligand. Although the cells were subjected to 24 h serum
starvation, both wt-EGFR and EGFR-Y845F tyrosine phosphorylation
were still detectable (data not shown), suggesting that excessive
EGFR molecules on the cell surface may be sufficient for triggering
its tyrosine phosphorylation even without participation of its
ligands. Notably, we observed that cell surface expression levels
and distribution between wt-EGFR and EGFR-Y845 are very similar to
each other (Fig. 6), suggesting the
EGFR-Y845F mutant might not change its protein folding and
structure. What caused EGFR phosphorylation and activation remains
elusive as this phenomenon seemingly mimics some types of cancer in
the clinical settings such as glioblastoma. Therefore, it is of
interest to understand how this ligand independent EGFR
phosphorylation occurs in the first place under the conditions of
EGFR overexpression. To avoid this issue, we carefully selected
cell lines with undetectable levels of endogenous EGFR and
generated stable clones expressing appropriate amounts of either
wt-EGFR or mutants as confirmed by flow cytometry analysis (data
not shown). The final selected clones expressing EGFR or mutants
with appropriate levels of EGFR that lacks self phosphorylation
were employed in the present study. These clones allowed us to
mimic EGFR cellular responses to its ligands in the conditions
close to the physiological environment. Therefore, the results may
possibly reflect real situations of EGFR phosphorylation and
functions. Collectively, our results from these studies suggested
that transphosphorylation of EGFR plays a critical role in its
ligand-induced kinase activity and functions as demonstrated in
Figs. 4 and 5.
As an important target therapeutic molecule in
various types of cancer such as non-small cell lung, breast, colon
and brain cancer, EGFR inhibition is dominant in the field of new
drug development and clinical applications. The mutations of EGFR
involved in tumor response or resistance to target therapeutic
reagents in various types of cancer have been documented as well.
These advances are helpful and critical to basic research and
clinical settings. However, the mutation and activation of EGFR
eventually have to link to its phosphorylation and kinase activity,
especially the interplay of its intrinsic phosphorylation.
Therefore, elucidation of the interaction of EGFR
transphosphorylation and autophosphorylation not only provides
insights into EGFR activation, but may also open up a novel venue
towards EGFR-associated target therapy.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China NSFC 31271495 (to H.S.), and the
Priority Academic Program Development of Jiangsu Higher Education
Institutions (to L.Y.).
References
|
1
|
Herbst RS, Fukuoka M and Baselga J:
Gefitinib - a novel targeted approach to treating cancer. Nat Rev
Cancer. 4:956–965. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hynes NE and Lane HA: ERBB receptors and
cancer: the complexity of targeted inhibitors. Nat Rev Cancer.
5:341–354. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Carlin CR and Knowles BB: Identity of
human epidermal growth factor (EGF) receptor with glycoprotein
SA-7: evidence for differential phosphorylation of the two
components of the EGF receptor from A431 cells. Proc Natl Acad Sci
USA. 79:5026–5030. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lynch TJ, Bell DW, Sordella R, et al:
Activating mutations in the epidermal growth factor receptor
underlying responsiveness of non-small-cell lung cancer to
gefitinib. N Engl J Med. 350:2129–2139. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sharma SV, Bell DW, Settleman J and Haber
DA: Epidermal growth factor receptor mutations in lung cancer. Nat
Rev Cancer. 7:169–181. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Verveer PJ, Wouters FS, Reynolds AR and
Bastiaens PI: Quantitative imaging of lateral ErbB1 receptor signal
propagation in the plasma membrane. Science. 290:1567–1570. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lax I, Fischer R, Ng C, Segre J, Ullrich
A, Givol D and Schlessinger J: Noncontiguous regions in the
extracellular domain of EGF receptor define ligand-binding
specificity. Cell Regul. 2:337–345. 1991.PubMed/NCBI
|
|
8
|
Zhou M, Felder S, Rubinstein M, Hurwitz
DR, Ullrich A, Lax I and Schlessinger J: Real-time measurements of
kinetics of EGF binding to soluble EGF receptor monomers and dimers
support the dimerization model for receptor activation.
Biochemistry. 32:8193–8198. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sorokin A, Lemmon MA, Ullrich A and
Schlessinger J: Stabilization of an active dimeric form of the
epidermal growth factor receptor by introduction of an
inter-receptor disulfide bond. J Biol Chem. 269:9752–9759.
1994.PubMed/NCBI
|
|
10
|
Lemmon MA, Bu Z, Ladbury JE, et al: Two
EGF molecules contribute additively to stabilization of the EGFR
dimer. EMBO J. 16:281–294. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Klein P, Mattoon D, Lemmon MA and
Schlessinger J: A structure-based model for ligand binding and
dimerization of EGF receptors. Proc Natl Acad Sci USA. 101:929–934.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dawson JP, Berger MB, Lin CC, Schlessinger
J, Lemmon MA and Ferguson KM: Epidermal growth factor receptor
dimerization and activation require ligand-induced conformational
changes in the dimer interface. Mol Cell Biol. 25:7734–7742. 2005.
View Article : Google Scholar
|
|
13
|
Burgess AW, Cho HS, Eigenbrot C, et al: An
open-and-shut case? Recent insights into the activation of EGF/ErbB
receptors. Mol Cell. 12:541–552. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang X, Gureasko J, Shen K, Cole PA and
Kuriyan J: An allosteric mechanism for activation of the kinase
domain of epidermal growth factor receptor. Cell. 125:1137–1149.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang X, Pickin KA, Bose R, Jura N, Cole
PA and Kuriyan J: Inhibition of the EGF receptor by binding of MIG6
to an activating kinase domain interface. Nature. 450:741–744.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jura N, Endres NF, Engel K, et al:
Mechanism for activation of the EGF receptor catalytic domain by
the juxtamembrane segment. Cell. 137:1293–1307. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Honegger AM, Kris RM, Ullrich A and
Schlessinger J: Evidence that autophosphorylation of solubilized
receptors for epidermal growth factor is mediated by intermolecular
cross-phosphorylation. Proc Natl Acad Sci USA. 86:925–929. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Skolnik EY, Margolis B, Mohammadi M, et
al: Cloning of PI3 kinase-associated p85 utilizing a novel method
for expression/cloning of target proteins for receptor tyrosine
kinases. Cell. 65:83–90. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sordella R, Bell DW, Haber DA and
Settleman J: Gefitinib-sensitizing EGFR mutations in lung
cancer activate anti-apoptotic pathways. Science. 305:1163–1167.
2004.PubMed/NCBI
|
|
20
|
Nyati MK, Morgan MA, Feng FY and Lawrence
TS: Integration of EGFR inhibitors with radiochemotherapy. Nat Rev
Cancer. 6:876–885. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sorkin A and Goh LK: Endocytosis and
intracellular trafficking of ErbBs. Exp Cell Res. 315:683–696.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Citri A and Yarden Y: EGF-ERBB signalling:
towards the systems level. Nat Rev Mol Cell Biol. 7:505–516. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yarden Y and Sliwkowski MX: Untangling the
ErbB signalling network. Nat Rev Mol Cell Biol. 2:127–137. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yarden Y: The EGFR family and its ligands
in human cancer: signalling mechanisms and therapeutic
opportunities. Eur J Cancer. 37(Suppl 4): S3–S8. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Morgan S and Grandis JR: ErbB receptors in
the biology and pathology of the aerodigestive tract. Exp Cell Res.
315:572–582. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Goel S, Hidalgo M and Perez-Soler R: EGFR
inhibitor-mediated apoptosis in solid tumors. J Exp Ther Oncol.
6:305–320. 2007.PubMed/NCBI
|
|
27
|
Tice DA, Biscardi JS, Nickles AL and
Parsons SJ: Mechanism of biological synergy between cellular Src
and epidermal growth factor receptor. Proc Natl Acad Sci USA.
96:1415–1420. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gotoh N, Tojo A, Hino M, Yazaki Y and
Shibuya M: A highly conserved tyrosine residue at codon 845 within
the kinase domain is not required for the transforming activity of
human epidermal growth factor receptor. Biochem Biophys Res Commun.
186:768–774. 1992. View Article : Google Scholar
|