Introduction
Luteolin is a flavonoid compound extracted from
several medicinal plants, including celery, green pepper and
perilla leaf (1). Flavonoids are
secondary plant metabolites, defined as substances that contain a
phenylchromanone structure (C6-C3-C6, with one or more hydroxyl
groups or other substituents) (2).
They are known to exert biological effects such as anticancer,
anti-inflammatory and anti-oxidative effects. They are also
associated with inhibition of angiogenic processes and modulation
of multidrug resistance (2–4). Although the incidence of cervical
cancer has declined following the introduction of human
papillomavirus (HPV) vaccines, it remains the leading cause of
cancer-related death in women (5).
Infection with HPV subtypes such as HPV-16, -18, -31 and -33
greatly increases the risk of cancer and plays a central role in
the development of ~99.5% of cervical cancers (6). HPVs are involved not only in cervical
cancer but also in several other cancer types, including neoplasms
of the head and neck and anal and penile cancers (7). The HPV genome consists of 6 early open
reading frames (ORFs) that encode early proteins such as E1, E2,
E4, E5, E6 and E7, as well as 2 late ORFs that encode late proteins
such as L1 and L2 (8). The E6 and
E7 oncoproteins have been shown to be key mediators of the
development of HPV-induced cervical carcinoma. HPV-related
transcription factors are involved in the G1/S and G2/M transition
within the cell cycle (9). The E6
protein, which associates with E6-associated protein (E6AP),
induces p53 degradation via ubiquitin-proteasome pathways. E7
protein inactivates the E7-associated factor pRb. As recently
discovered, E2F5 activates the cell cycle through direct
transcriptional activation of E7 in HPV-18-infected HeLa cells
(9,10). Additionally, HPV escapes the immune
system of the infected host. For example, E6 and E7 mediate
protection against interferons and elicit inappropriate or reduced
activation of antigen-presenting cells (7). E6- and E7-mediated inhibition of the
host’s immune system underlies the effects of the related disease
(11–14). Under conditions in which E6 and E7
gene expression is inhibited, tumor-suppressor proteins such as pRb
and p53 are activated at normal levels.
Cell death can occur as one of two distinct
processes: apoptosis and necrosis. Necrosis is a degenerative
pattern that follows cell injury. Conversely, apoptosis is an
active process, requiring protein synthesis for its execution.
Apoptosis can be distinguished from necrosis by a number of unique
features, such as chromatin condensation, DNA fragmentation, cell
membrane flip-flop, cytoplasmic shrinkage, and the formation of
apoptotic bodies (15). Apoptosis
commonly occurs via two distinct pathways, intrinsic and extrinsic
pathways, which are connected at a number of intermediate steps
(16). Caspase-8 activation plays a
critical role in the extrinsic apoptotic pathway. However,
caspase-8 and caspase-3 are activated by each other via caspase-3
and -8 cross-activation responses (17,18).
The intrinsic apoptotic pathway involves a cascade of molecular
events that occur entirely within the cell. Mitochondria are
central to the intrinsic apoptotic pathway. On receipt of an
apoptotic signal, a pro-apoptotic member of the Bcl-2 family, such
as Bax, oligomerizes and inserts into the mitochondrial membrane,
permeabilizing it and allowing cytochrome c to redistribute
into the cytoplasm (19). Intrinsic
pathway-related p53 protein is degraded by the HPV E6 oncoprotein
(20).
Luteolin has been well known to exhibit an
anticancer effect in several cancer cell lines. However, the exact
mechanism behind this effect remains to be elucidated. In the
present study, we focused on the intrinsic and extrinsic pathways
related to E6 and E7 oncogenes. We demonstrated that luteolin
exerts anticancer effects targeted to the E6 and E7 oncogenes and
caspase-8 and caspase-3 are activated by each other through
caspase-3 and -8 cross-activation responses.
Materials and methods
Cell culture and treatment
HPV-18-positive HeLa cervical cancer cells,
HPV-16-positive SiHa and CaSki cervical carcinoma cells, and
HPV-negative C33A cervical cancer cell line were obtained from the
American Type Culture Collection (ATCC; Rockville, MD, USA). Cells
were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Hyclone
Laboratories, Logan, UT, USA) supplemented with 2 mM/L-glutamine
and 10% fetal bovine serum (FBS; Hyclone Laboratories) and
incubated under humidified conditions at 37°C with 5%
CO2. Treatment was performed by adding 5, 10 and 20 μM
luteolin (Sigma, St. Louis, MO, USA) directly to the culture media
for 48 h.
Cell viability test
Cell viability was quantified using
3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-
(4-sulfophenyl)-2H-tetrazolium (MTS) (Promega, Madison, WI, USA)
reagent as previously described (21). Cervical cancer cells
(1×105) were seeded in 100 μl of medium in 96-well
plates and incubated overnight. After 20 h, the cells were treated
with various doses of luteolin for 24 and 48 h. Samples of the
media were removed, and the cells were incubated with 20 μl of MTS
(2 mg/ml) and phenazine methosulfate (PMS) (Sigma) mixed solution
in serum-free DMEM for 1 h at 37°C. Optical absorbance was measured
at 492 nm using a spectrophotometer (Apollo LB 9110; Berthold
Technologies GmbH, Germany).
DAPI staining
Characteristic apoptotic nuclear morphologic changes
can be detected in cells stained with 4′,6-diamidino-2-phenylindole
(DAPI) (Sigma) stain solution and by using fluorescence microscopy
at ×100 magnification (22).
Cervical cancer cells were seeded on coverslips in 6-well plates
and treated with luteolin for 48 h. After washing with
phosphate-buffered saline (PBS), the HeLa cells were fixed with
para-formaldehyde and stained with Hoechst staining solution at
37°C. The coverslips were then washed with PBS, dried completely,
and mounted on microscope slides with mounting solution. These
slides were examined using fluorescence microscopy.
Flow cytometric analysis using Annexin V
and PI staining
Annexin V and propidium iodide (PI) stains were used
to observe the progression of apoptosis (23). Cells were seeded in 6-well plates
and treated with luteolin for 48 h. After treatment, the cells were
harvested using trypsin-EDTA and washed twice by centrifugation
(300 × g). Annexin V and PI staining were performed using the
FITC-Annexin V Apoptosis Detection Kit I (BD Biosciences, San
Diego, CA, USA) in accordance with the manufacturer’s instructions.
The percentages of early and late apoptotic cells were calculated
using Annexin-V-positive/PI-negative and
Annexin-V-positive/PI-positive signals, respectively.
Western blot analysis
To identify protein expression levels, HeLa cells
were seeded on 6-well plates after 48 h of luteolin treatment.
Harvested cells were lysed in a radioimmunoprecipitation assay
(RIPA) buffer (0.1% sodium dodecyl sulfate-SDS), 0.1% sodium
deoxycholate, 1% Triton X-100, 1 mM EDTA, 0.5 mM ETDA, 140 mM NaCl
and 10 mM Tris-HCl, pH 8.0) containing phosphate and protease
inhibitors. Cell lysates were centrifuged at maximum speed for 30
min at 4°C. The protein concentration of the resulting supernatant
was quantified. Component proteins were separated by sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and then
transferred to polyvinylidene difluoride (PVDF) membranes. The
membranes were blocked by incubation in 5% non-fat milk solution in
Tris-buffered saline containing Tween-20 (TBST: 2.7 M NaCl, 53.65
mM KCl, 1 M Tris-HCl, pH 7.4, 0.1% Tween-20) for 1 h at room
temperature. After blocking, these membranes were incubated in 1%
milk solution containing the primary antibody in TBST for 2–4 h.
After three consecutive washes in TBST, the membranes were
incubated with the secondary antibodies [horseradish peroxidase
(HRP)-conjugated α-rabbit or α-mouse IgG] for 1 h at room
temperature. After washing the membranes 3 times, the signal was
visualized using the Westzol Plus Western Blotting detection system
(iNtRON Biotechnology, SungNam, Korea). Antibodies specific to
cyclin D were purchased from BD Biosciences (San Diego, CA, USA).
Antibodies specific to PARP, caspase-3, caspase-9, caspase-8,
Bcl-2, Bcl-xL, p-pRb, p53, p-p53 and cytochrome c, and
anti-mouse IgG-horseradish peroxidase were purchased from Cell
Signaling Technology (Beverly, MA, USA). Antibodies specific to
p21, GAPDH and anti-goat IgG-HRP were from Santa Cruz Biotechnology
(Santa Cruz, CA, USA). Pan-caspase inhibitor (Z-VAD-fmk), caspase-3
inhibitor (Z-DEVD-fmk), caspase-9 inhibitor (Z-LEHD-fmk) and
caspase-8 inhibitor (Z-IETD-fmk) were from R&D Systems
(Minneapolis, MN, USA).
Reverse-transcription polymerase chain
reaction (RT-PCR) and real-time qPCR
The cells were harvested and lysed using the
easy-BLUE™ Total RNA Extraction kit (iNtRon Biotechnology, Seoul,
Korea) according to the manufacturer’s instructions. The
Oligo-primed RNAs (5 μg) were reverse transcribed using M-MuLV
reverse transcriptase (New England Biolabs, Beverly, MA, USA).
RT-PCR analysis was performed using a PCR Thermal Cycler Dice
instrument (Takara, Otsu, Shiga, Japan) with the following primer
sets: HPV-18 E6, 5′-GCG ACC CTA CAA GCT ACC TG-3′ (forward) and
5′-GTT GGA GTC GTT CCT GTC GT-3′ (reverse); HPV-18 E7, 5′-GCA TGG
ACC TAA GGC AAC AT-3′ (forward) and 5′-TGT TGC TTA CTG CTG GGA
TG-3′ (reverse); E2F5, 5′-ACC TAT CCA TGT GCT GCT TA-3′ (forward)
and 5′-AGA TTT TGA GTT GCC ATG CT-3′ (reverse); DR5, 5′-GTC TGC TCT
GAT CAC CCA AC-3′ (forward) and 5′-CTG CAA ACT GTG ACT CCT ATG-3′
(reverse); FasL, 5′-CAA GAT TGA CCC CGG AAG TA-3′ (forward); GAPDH,
5′-TGA TGA CAT CAA CAA GGT GGT-3′ (forward) and 5′-TCC TTG GAG GCC
ATG TAG GCC-3′ (reverse). GAPDH was used as an internal control.
Real-time quantitative PCR was performed using a relative
quantification protocol using the Chromo 4 Real-Time PCR system and
iQ SYBR-Green Supermix (both from Bio-Rad, Hercules, CA, USA). All
the target genes were normalized to the expression of the
housekeeping gene GAPDH. Each sample was run with the following
primer sets: HPV-18 E6 qPCR, 5′-TAT TTG TGG TGT ATA GAG AC-3′
(forward) and 5′-CAG TGT TAG TTA GTT TTT CC-3′ (reverse); HPV-18 E7
qPCR, 5′-CTC AGA GGA AGA AAA CGA TG-3′ (forward) and 5′-GGC TTC ACA
CTT ACA ACA CA-3′ (reverse); FADD, 5′-ACC TCT TCT CCA TGC TG-3′
(forward) and 5′-CAC ACA GGT CTT CCC CA-3′ (reverse); Fas, 5′-TGA
AGG ACA TGG CTT AGA AGT G-3′ (forward) and 5′-GGT GCA AGG GTC ACA
GTG TT-3′ (reverse); TRAIL, 5′-AAG TTT GTC GTC GTC GGG GT-3′
(forward) and 5′-TGG TGC AGG GAC TTC TCT CT-3′ (reverse); GAPDH
qPCR, 5′-GGC TGC TTT TAA CTC TGG TA-3′ (forward) and 5′-TGG AAG ATG
GTG ATG GGA TT-3′ (reverse). The fold changes in expression
represent the ratio of E6 and E7 expression in the luteolin-treated
cervical cancer cells compared to the untreated control.
Nuclear and cytoplasmic
fractionation
HeLa cells were treated with luteolin for 48 h prior
to harvesting and fractionation with NE-PER nuclear and cytoplasmic
extraction reagents (Thermo Fisher Scientific Inc, Rockford, IL,
USA) according to the manufacturer’s instructions. Briefly, cells
were collected by centrifugation at 1,980 × g and 4°C for 5 min,
washed with PBS, and re-centrifuged. The cell pellet was suspended
with buffer I, vortexed, and incubated on ice for 10 min. Buffer II
was added and the cells were incubated for 1 min prior to
centrifugation at 16,000 × g and 4°C for 5 min, yielding a
cytoplasmic extract as the supernatant. The insoluble pellet was
then suspended in buffer III, incubated on ice for 40 min, and
centrifuged at 16,000 × g for 10 min, yielding the nuclear extract
as the supernatant. Equal quantities of protein from these extracts
(50 μg) were separated by SDS-PAGE.
Analysis of mitochondrial transmembrane
potential by JC-1
JC-1 (Enzo, Farmingdale, NY, USA) stain can detect
differences in mitochondrial membrane potential (MMP) (24). Cells were harvested with
trypsin-EDTA, transferred to 1.5-ml tubes containing JC-1 stain
solution, and incubated for 10 min in the dark. The samples were
centrifuged at 300 × g and 4°C for 3 min, washed twice with PBS,
and resuspended in PBS for fluorescence-activated cell sorting
(FACS) analysis.
Statistical analysis
Data are presented as the mean ± SEM of results from
at least 3 independent experiments. Statistical significance was
assessed using the Student’s t-test, with P<0.05 considered
statistically significant. P<0.05, P<0.01 and P<0.001 are
indicated in the figure legends.
Results
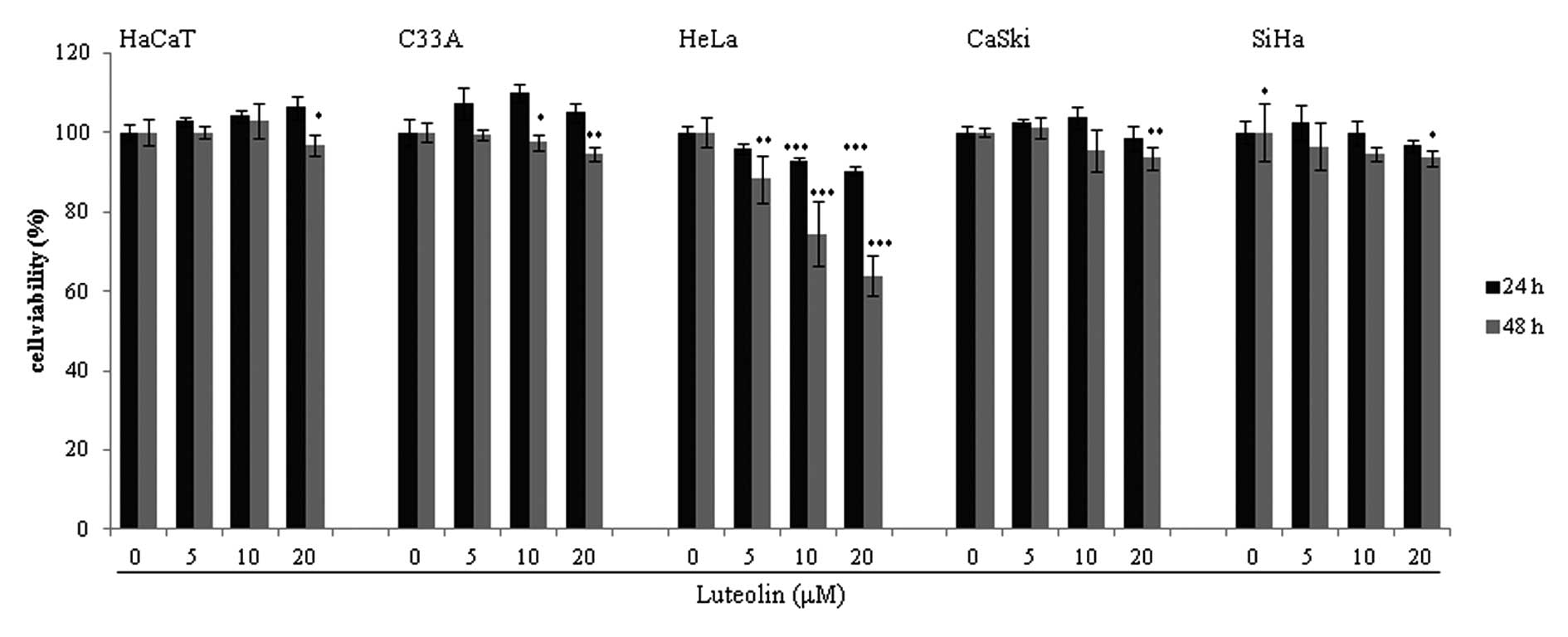
Luteolin treatment reduces the viability
of HeLa cells
The cervical cancer cell lines and the normal
keratinocyte HaCaT cell line were treated with various
concentrations of luteolin (Fig.
1). Mild cytotoxicity was observed in the HPV-16-positive SiHa
and CaSki cell lines at a high concentration of luteolin (>40
μM, data not shown). Luteolin showed a significant dose-dependent
cytotoxic effect in the HeLa HPV-18-positive cervical cancer cells.
However, there was no cytotoxic effect of luteolin in the
HPV-negative C33A cervical cancer cell line and in the HaCaT human
normal keratinocytes. Therefore, we focused on HeLa cells to
further investigate the apoptotic effect of luteolin.
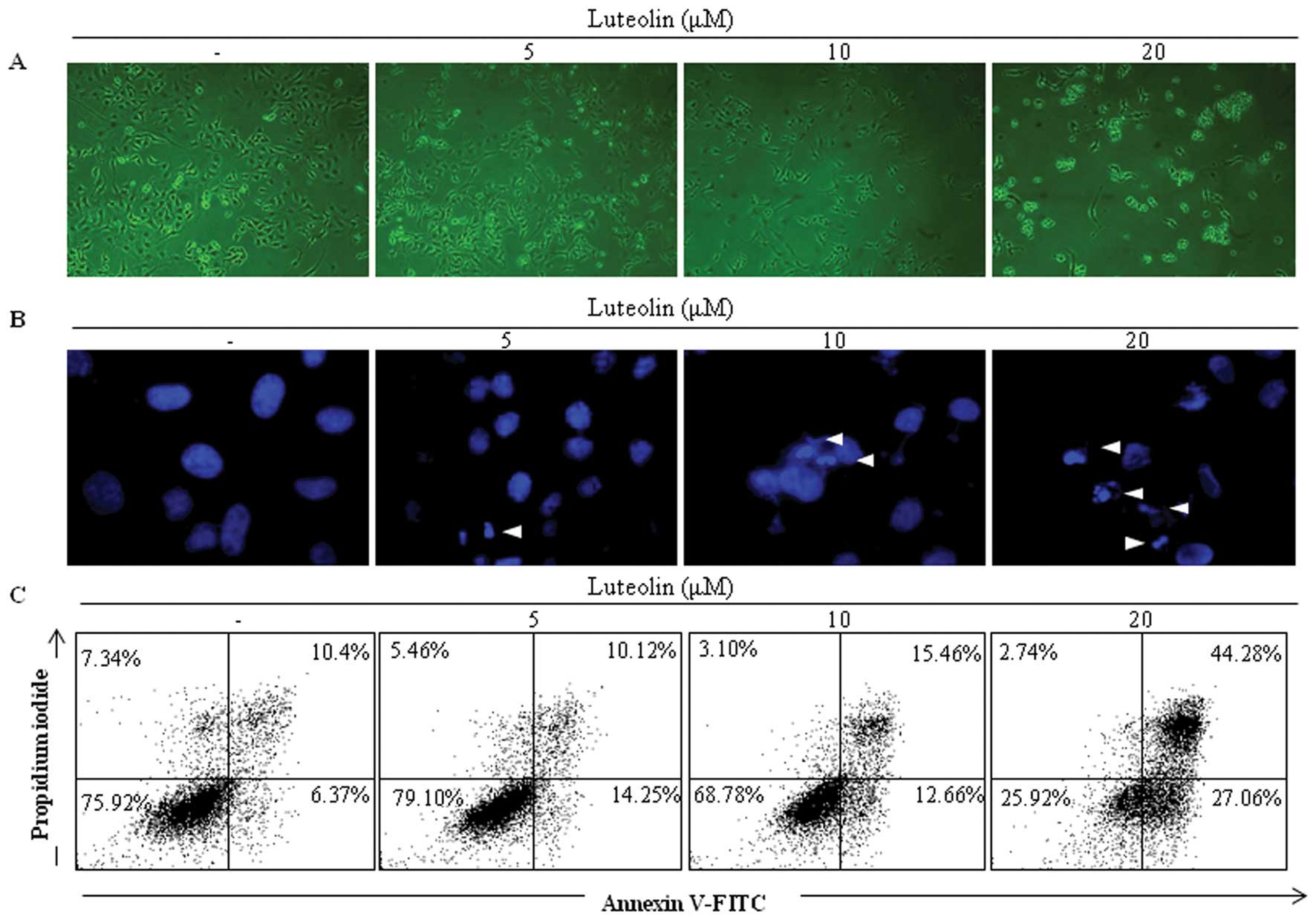
Luteolin induces apoptosis in HeLa
cells
Phase-contrast microscopy revealed that luteolin
induced cell death in the HeLa cells in a dose-dependent manner
after 48 h of treatment (Fig. 2A).
Nuclear condensation detected by DAPI staining is one of the
commonly used markers of apoptosis. HeLa cells treated with
luteolin exhibited notable nuclear condensation (Fig. 2B). Annexin V-FITC/PI staining is
commonly used to detect apoptosis and necrosis. Positive Annexin
V-FITC staining suggests cell exposure to phosphatidyl serine.
Positive PI staining is associated with cell membrane disruption.
Double positive staining with both Annexin V-FITC and PI reflects
the flip-flop of phosphatidyl serine and membrane disruption
(23). Luteolin-treated HeLa cells
exhibited Annexin V-FITC single positive staining and Annexin
V-FITC/ PI double-positive staining, indicating that luteolin
induced apoptosis in the HeLa cells (Fig. 2C).
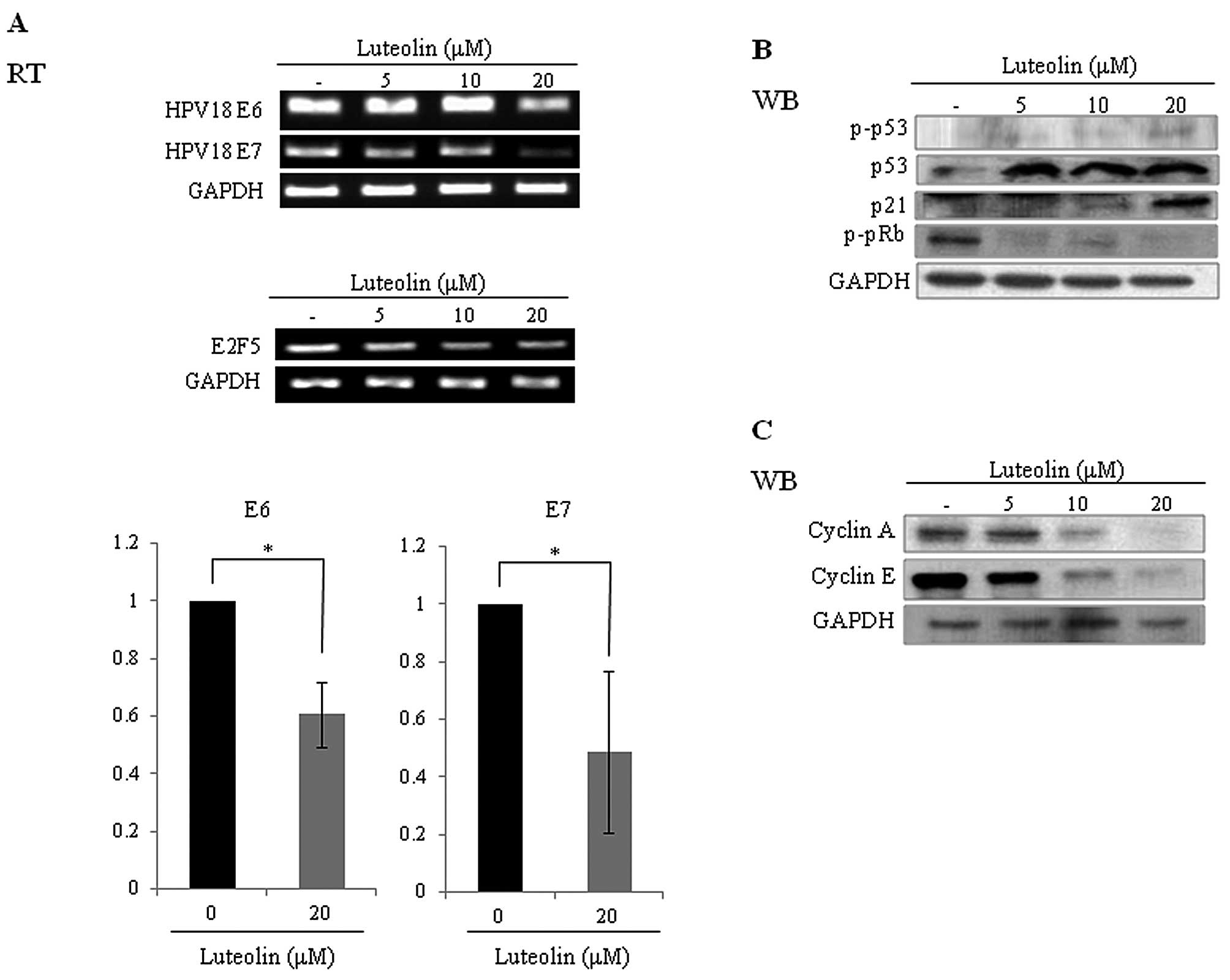
Effects of luteolin on mRNA levels of
E6/E7 and on expression levels of cyclins, pRb and p53 in HeLa
cells
The E6 protein binds, ubiquitinates and degrades the
tumor-suppressor protein p53, while the E7 protein binds and
interferes with the Rb/E2F complex (20). Levels of E6 and E7 mRNA were reduced
by luteolin treatment in the HeLa cells (Fig. 3A). Following incubation of HeLa
cells with luteolin, the p53 level was increased in a
dose-dependent manner, while levels of phosphorylated pRb (p-pRb)
and E7 activator E2F5 were decreased. These results indicated that
activities of p53 and pRb in the HeLa cells were recovered by
luteolin treatment. The p53 downstream factor p21 level was also
increased (Fig. 3B). Cyclins are
differentially regulated in every step of the cell cycle. Cyclin A
and E are modulated by E6 and E7 proteins, as E7 induces cyclin E
expression (25). Cyclin A
overexpression has been reported in an HPV-infected cancer cell
line (25). E6 abrogates the
repression of cyclin A transcription by degradation of p53
(26). Expression levels of cyclin
A and E were suppressed by luteolin treatment in the HeLa cells as
expected (Fig. 3C).
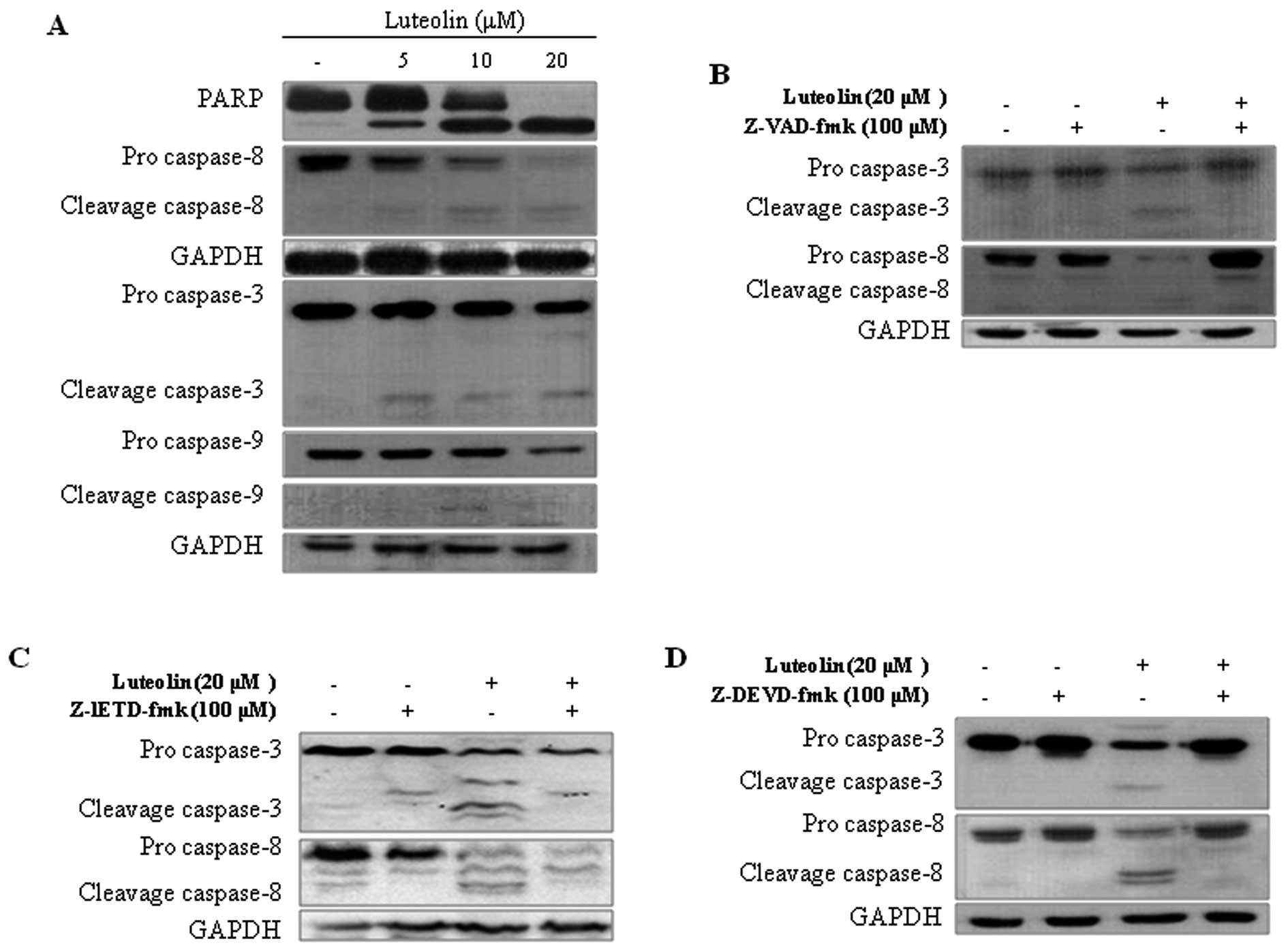
Effect of luteolin on apoptosis-related
factors, caspase-3 and -8 in HeLa cells
Caspases and polyADP ribose polymerase (PARP) are
important mediators of apoptosis and are known to contribute to the
overall apoptotic morphologic changes by the cleavage of a number
of cellular substrates (27).
Western blot analysis was used to further detect protein expression
of caspase-3, -8, and -9 and PARP in HeLa cells after luteolin
treatment for 48 h. As shown in Fig.
4, caspase-3, -8, and -9 and PARP proteins were cleaved to the
corresponding active forms. These results indicate that luteolin
treatment induces apoptotic death in HeLa cells through a
caspase-dependent pathway. HeLa cells were pretreated with caspase
inhibitors to identify the specific caspases involved in the
apoptotic mechanism. A pan-caspase inhibitor Z-VAD-fmk and
caspase-3 inhibitor Z-DEVD-fmk fully alleviated luteolin-induced
cleavage of caspase-3 and -8 in the HeLa cells (Fig. 4B and D). However, the caspase-8
inhibitor Z-IETD-fmk only partially inhibited the cleavage of
caspase-3 and -8 induced by luteolin (Fig. 4C). Taking all the results into
consideration, our findings suggest that luteolin activates
caspase-3, which in turn induces caspase-8 processing and vice
versa in HeLa cells. Overall, these analyses revealed that caspases
play pivotal roles in luteolin-induced apoptosis.
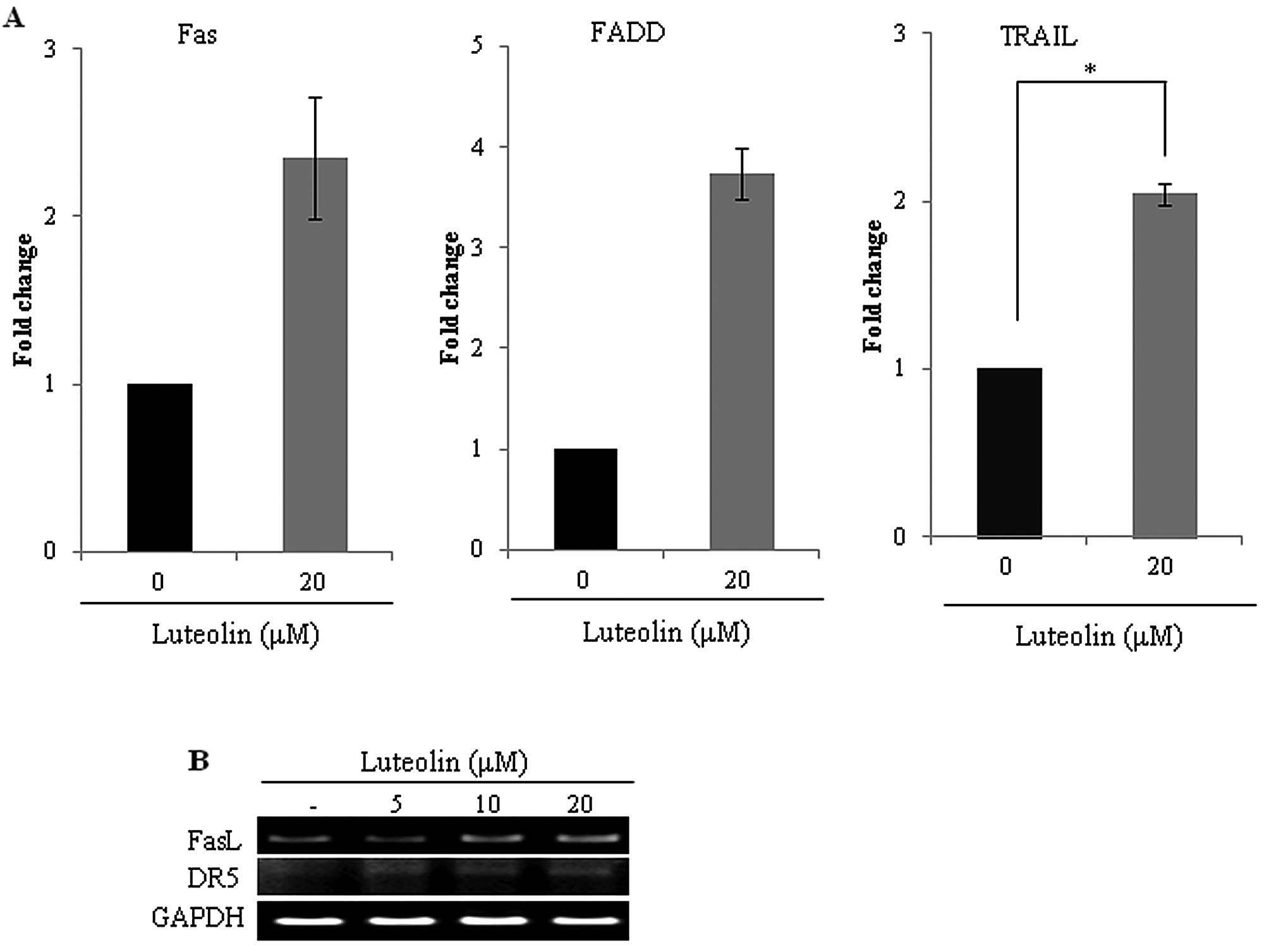
Luteolin enhances Fas death receptor
expression in HeLa cells
Cervical cancer cells harboring HPV express E6
oncoprotein, which inhibits apoptotic signaling in response to Fas
associated protein with death domain (FADD) (28). Since luteolin induced caspase-8
processing and inhibited E6 protein in HeLa cells, we further
investigated whether luteolin modulates death receptor expression
in HeLa cells. Luteolin enhanced death receptor (Fas/FasL, DR5,
FADD) expression in the HeLa cells (Fig. 5), demonstrating that luteolin
induced extrinsic apoptosis via death receptor-caspase-8/3
pathways.
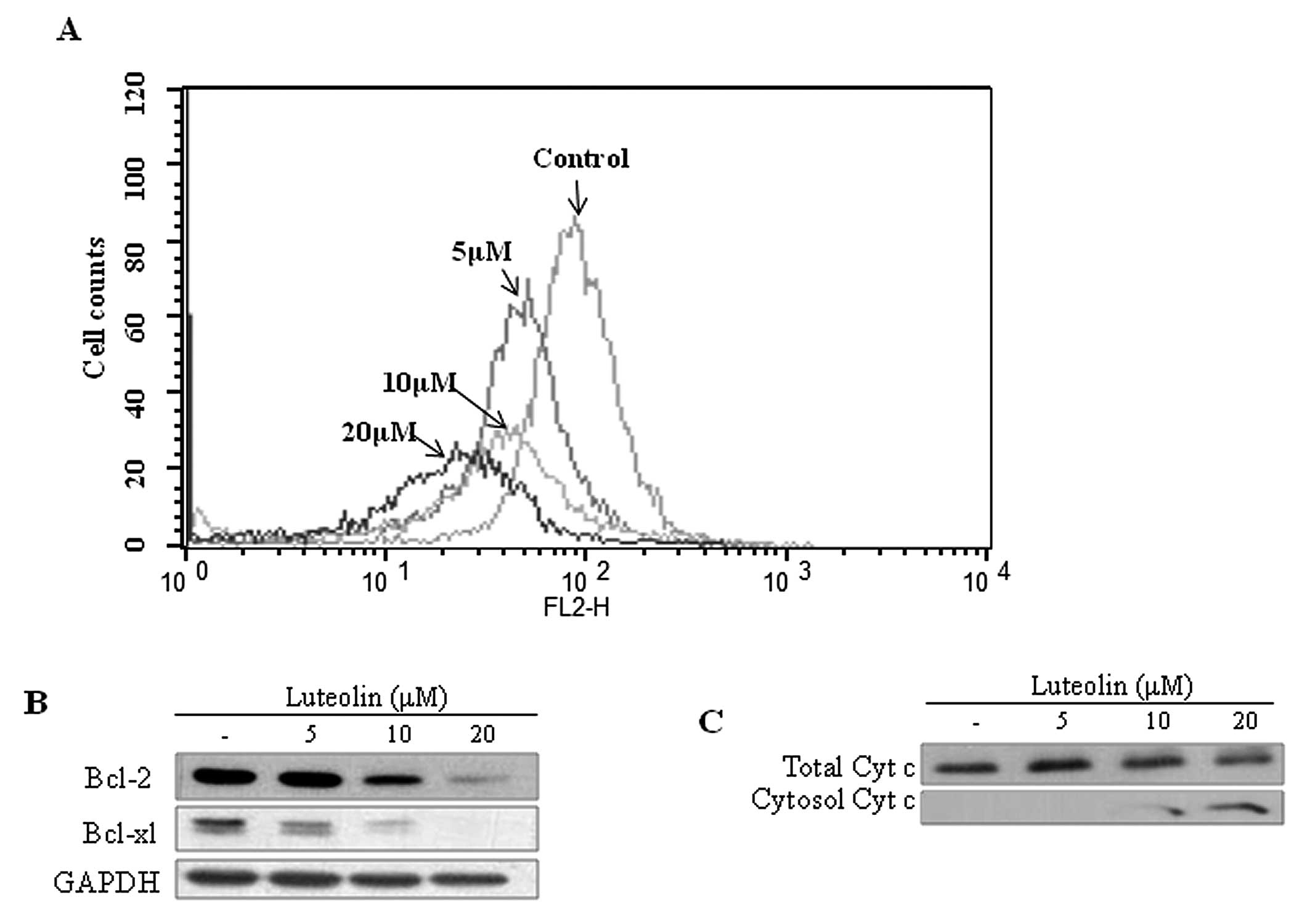
Luteolin disrupts MMP in HeLa cells
Disruption of MMP is one of the critical steps in
the intrinsic apoptotic pathway (5). When apoptosis progresses, JC-1-stained
cells are altered from orange to green as MMP decreases (24,29).
MMP was measured by JC-1 staining in the luteolin-treated HeLa
cells. As shown in Fig. 6A,
JC-1-stained HeLa cells exhibited a dose-dependent left shift. This
result indicates that the mitochondrial membrane was disrupted by
luteolin treatment. Bcl-2 and Bcl-xL expression levels were also
decreased following luteolin treatment (Fig. 6B). Cytochrome c was detected
by fractionation and western blot analysis to confirm its release
into the cytosol from the mitochondria. Cytochrome c was
released into the cytosol in the luteolin-treated HeLa cells
(Fig. 6C).
Discussion
Luteolin, a polyphenolic compound present in celery,
green peppers, perilla leaf and chamomile tea, belongs to the
flavone subclass of flavonoids (1).
This compound effectively suppresses the development and
progression of stomach, cervical, lung, and bladder cancers.
However, the molecular mechanisms underlying its anticancer effects
in cervical cancers are poorly understood. The precise relationship
between luteolin and HPV infection in particular has thus far never
been reported. The cytotoxic effects against HPV-positive cell
lines were assessed following a 48-h luteolin treatment. HeLa cells
in particular showed effective cytotoxicity at 48 h after luteolin
treatment. The cytotoxic effect and markers of apoptotic activity
were induced by luteolin treatment in HeLa cells.
HeLa cells harbor the HPV-18 genome, which encodes
the E6 and E7 oncogenes (10). The
E6 protein degrades p53, one of the tumor-suppressor proteins. The
E7 protein interferes with the pRb/E2F complex, freeing E2F to
serve as a transcription factor (30). Reduction of E6 and E7 protein
expression in cervical cancer cells results in the reactivation of
p53 and pRb, and can lead to apoptosis and regulation of the cell
cycle. Moreover, overexpression of cyclin A and cyclin E has been
found in cells expressing E6 and E7 (25). As shown in Fig. 4, E6 and E7 mRNA levels were reduced
by luteolin treatment in HeLa cells. Consequently, E6 and E7
inhibition, reactivation of p53 and pRb, and suppression of cyclin
A and cyclin E expression were induced by treatment with luteolin,
resulting in the induction of apoptosis and re-establishment of the
cell cycle control in HeLa cells. It has also been reported that
HPV can modulate or evade the immune system through the E6 and E7
oncogenes (7). These results
suggest that luteolin may be used as a controlling agent against
HPV-positive cancers and cervical cancer through targeting E6/E7
oncogenes in vitro and in vivo.
In the present study, we confirmed that
luteolin-induced apoptosis is mediated by the death receptors. PCR
and Western blot analyses revealed that luteolin enhanced DR5 death
receptor expression (data not shown), caspase-8 activity, and TRAIL
expression, as expected from previous research (31). Research also revealed that E6
protein inhibits Fas and Fas-associated proteins (28). Luteolin inhibited E6 expression and
enhanced Fas and FADD in HeLa cells, suggesting that death
receptors were upregulated via inhibition of E6 expression.
However, caspase-8 activity was also upregulated by the activity of
caspase-3, as well as by death receptors such as DR5 and Fas. All
specific inhibitors of caspase-3 and -8, as well as pan-caspase
inhibitors, alleviated luteolin-induced activation of caspase-3 and
-8. In the present study, we used caspase inhibitors to show that
caspase-8 is activated by caspase-3 and that caspase-8 in turn
activates caspase-3. It was reported in a previous report that
certain anticancer drugs, such as paclitaxel, induce apoptosis via
the caspase-8 and caspase-3 amplification loop (17). It was also shown that
luteolin-mediated apoptosis might be mediated via the caspase-8 and
caspase-3 amplification loop in HeLa cells.
The mitochondrial-dependent pathway is the most
recognized intrinsic apoptosis pathway. Disruption of mitochondrial
membrane potential is a key commitment step in the induction of the
intrinsic pathway. This signaling pathway leads to the release of
apoptotic proteins from the mitochondrial inter-membrane space
(32). As shown in Fig. 7, our results clearly demonstrated
that luteolin treatment led to a disruption of the mitochondrial
membrane potential. Moreover, luteolin reduced expression levels of
Bcl-2 and Bcl-xL, which inhibit the mitochondrial-mediated
apoptosis pathway. This result indicates that luteolin induces
apoptosis via the mitochondrial-dependent pathway.
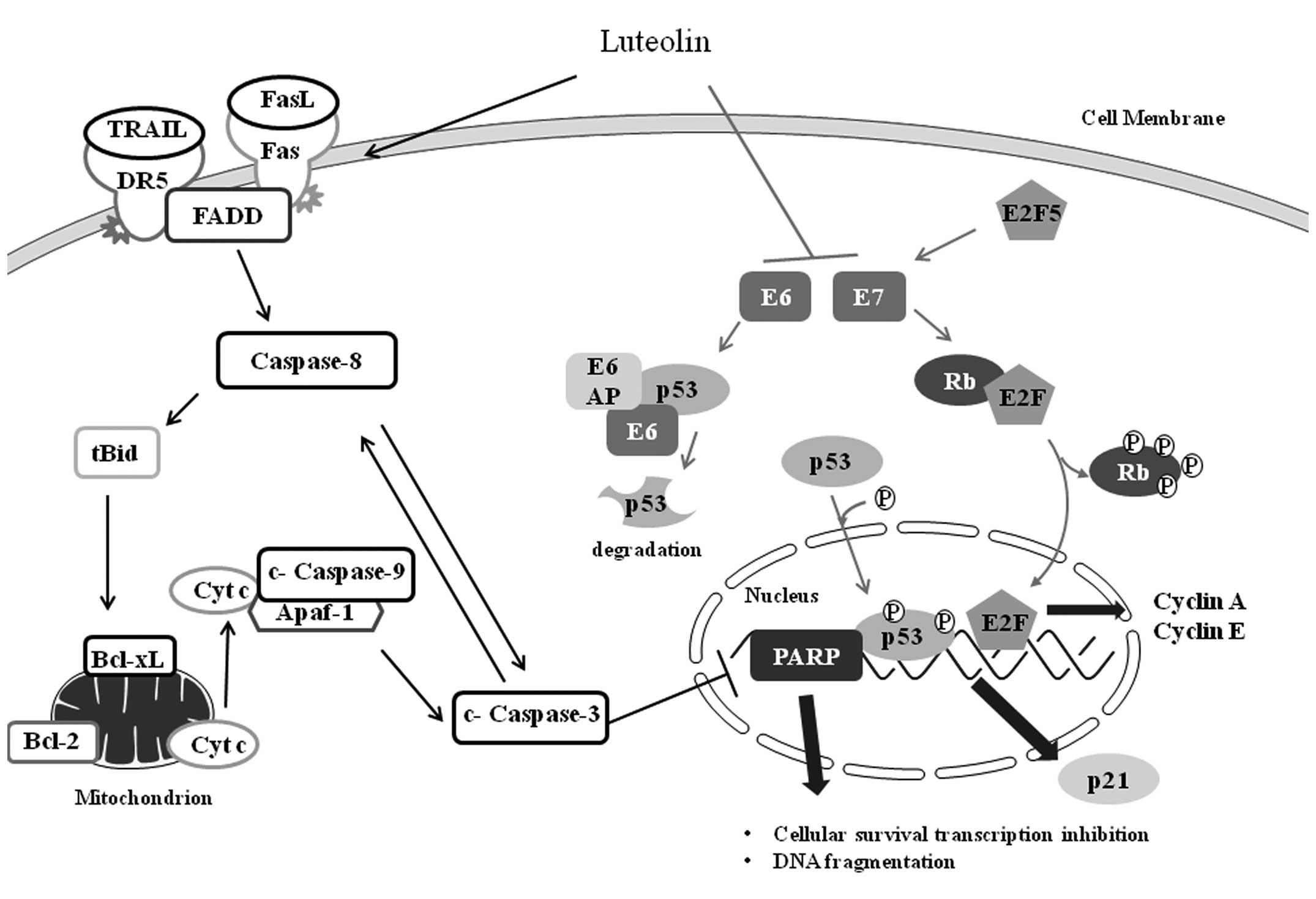 | Figure 7Schematic diagram illustrating the
effect of luteolin-induced apoptosis in HeLa cells. Luteolin
triggers progression of the death receptor (DR5/TRAIL,
Fas/FasL)-mediated apoptotic signaling pathway, by inducing
caspase-8 processing, resulting in cytochrome c (Cyt c)
release. Moreover, luteolin activates caspase-3, which in turn
induces caspase-8 processing and vice versa in HeLa cells,
indicating that caspases play pivotal roles in luteolin-induced
apoptosis. In addition, luteolin suppressed the expression of E6
and E7 oncogenes, resulting in the recovery of tumor suppressors
such as p53 and Rb. Tumor suppressors p53 and Rb can induce
cyclin-dependent kinase inhibitor p21 and inhibit E2F,
respectively. Luteolin also inhibits expression of E2F5, which
activates the cell cycle through direct transcriptional activation
of E7. |
In conclusion, luteolin was shown to target the
E6/E7 pathways and the caspase-8/-3 amplification loop (17). Luteolin inhibited HeLa
HPV-18-positive cancer cell proliferation, while it did not affect
the HPV-negative cancer cells and the normal HaCaT keratinocytes.
The expression levels of E6 and E7 oncogenes were suppressed by
treatment with luteolin. Notably, inhibitors of caspase-3 and -8
confirmed that caspase-8 activity was activated by active
caspase-3, suggesting that caspase-8 is activated by the DR5 and
Fas death receptors and that activation signals of caspase-8 and
caspase-3 interact by cross-activation.
Acknowledgements
This work was supported by the Basic Program
(2013-A423-0061) of the National Research Foundation of Korea (NRF)
and by the Ministry for Health, Welfare and Family affairs, Korea
(A120833). D-Y.Y. was supported partially by the Priority Research
Centres Program (2012-0006686).
References
|
1
|
Seelinger G, Merfort I, Woelfle U and
Schempp CM: Anti-carcinogenic effects of the flavonoid luteolin.
Molecules. 13:2628–2651. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Birt DF, Hendrich S and Wang W: Dietary
agents in cancer prevention: flavonoids and isoflavonoids.
Pharmacol Ther. 90:157–177. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ren W, Qiao Z, Wang H, Zhu L and Zhang L:
Flavonoids: promising anticancer agents. Med Res Rev. 23:519–534.
2003. View Article : Google Scholar
|
|
4
|
Hoensch H and Oertel R: Anti-inflammatory
effects of tea-flavonoids. Dtsch Med Wochenschr. 137:2738–2740.
2012.(In German).
|
|
5
|
Ju HK, Lee HW, Chung KS, et al:
Standardized flavonoid-rich fraction of Artemisia princeps
Pampanini cv. Sajabal induces apoptosis via mitochondrial pathway
in human cervical cancer HeLa cells. J Ethnopharmacol. 141:460–468.
2012.PubMed/NCBI
|
|
6
|
Munagala R, Kausar H, Munjal C and Gupta
RC: Withaferin A induces p53-dependent apoptosis by repression of
HPV oncogenes and upregulation of tumor suppressor proteins in
human cervical cancer cells. Carcinogenesis. 32:1697–1705. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Stern PL, van der Burg SH, Hampson IN, et
al: Therapy of human papillomavirus-related disease. Vaccine.
30(Suppl 5): F71–F82. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Grabowska AK and Riemer AB: The invisible
enemy - how human papillomaviruses avoid recognition and clearance
by the host immune system. Open Virol J. 6:249–256. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Teissier S, Pang CL and Thierry F: The
E2F5 repressor is an activator of E6/E7 transcription and of the
S-phase entry in HPV18-associated cells. Oncogene. 29:5061–5070.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Doorbar J, Quint W, Banks L, et al: The
biology and life-cycle of human papillomaviruses. Vaccine. 30(Suppl
5): F55–F70. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Adhim Z, Otsuki N, Kitamoto J, et al: Gene
silencing with siRNA targeting E6/E7 as a therapeutic intervention
against head and neck cancer-containing HPV16 cell lines. Acta
Otolaryngol. 133:761–771. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jonson AL, Rogers LM, Ramakrishnan S and
Downs LS Jr: Gene silencing with siRNA targeting E6/E7 as a
therapeutic intervention in a mouse model of cervical cancer.
Gynecol Oncol. 111:356–364. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Manzo-Merino J, Thomas M, Fuentes-Gonzalez
AM, Lizano M and Banks L: HPV E6 oncoprotein as a potential
therapeutic target in HPV related cancers. Expert Opin Ther
Targets. 17:1357–1368. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bosch FX, Broker TR, Forman D, et al:
Comprehensive control of human papillomavirus infections and
related diseases. Vaccine. 31(Suppl 7): H1–H31. 2013. View Article : Google Scholar
|
|
15
|
Walker NI, Harmon BV, Gobé GC and Kerr JF:
Patterns of cell death. Methods Achiev Exp Pathol. 13:18–54.
1988.
|
|
16
|
Bak YS, Kim HJ, Kang JW, et al: A
synthetic naringenin derivative,
5-hydroxy-7,4′-diacetyloxyflavanone-N-phenyl hydrazone (N101-43),
induces apoptosis through up-regulation of Fas/FasL expression and
inhibition of PI3K/Akt signaling pathways in non-small-cell lung
cancer cells. J Agric Food Chem. 59:10286–10297. 2011.PubMed/NCBI
|
|
17
|
von Haefen C, Wieder T, Essmann F,
Schulze-Osthoff K, Dorken B and Daniel PT: Paclitaxel-induced
apoptosis in BJAB cells proceeds via a death receptor-independent,
caspases-3/-8-driven mitochondrial amplification loop. Oncogene.
22:2236–2247. 2003.PubMed/NCBI
|
|
18
|
Chung KS, Choi JH, Back NI, et al:
Eupafolin, a flavonoid isolated from Artemisia princeps,
induced apoptosis in human cervical adenocarcinoma HeLa cells. Mol
Nutr Food Res. 54:1318–1328. 2010.PubMed/NCBI
|
|
19
|
Franklin JL: Redox regulation of the
intrinsic pathway in neuronal apoptosis. Antioxid Redox Signal.
14:1437–1448. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tan S, de Vries EG, van der Zee AG and de
Jong S: Anticancer drugs aimed at E6 and E7 activity in
HPV-positive cervical cancer. Curr Cancer Drug Targets. 12:170–184.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cory AH, Owen TC, Barltrop JA and Cory JG:
Use of an aqueous soluble tetrazolium/formazan assay for cell
growth assays in culture. Cancer Commun. 3:207–212. 1991.PubMed/NCBI
|
|
22
|
Daxhelet GA, Coene MM, Hoet PP and Cocito
CG: Spectrofluorometry of dyes with DNAs of different base
composition and conformation. Anal Biochem. 179:401–403. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vermes I, Haanen C, Steffens-Nakken H and
Reutelingsperger C: A novel assay for apoptosis. Flow cytometric
detection of phosphatidylserine expression on early apoptotic cells
using fluorescein labelled Annexin V. J Immunol Methods. 184:39–51.
1995. View Article : Google Scholar
|
|
24
|
Smiley ST, Reers M, Mottola-Hartshorn C,
et al: Intracellular heterogeneity in mitochondrial membrane
potentials revealed by a J-aggregate-forming lipophilic cation
JC-1. Proc Natl Acad Sci USA. 88:3671–3675. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Milde-Langosch K and Riethdorf S: Role of
cell-cycle regulatory proteins in gynecological cancer. J Cell
Physiol. 196:224–244. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yamamoto M, Yoshida M, Ono K, et al:
Effect of tumor suppressors on cell cycle-regulatory genes: RB
suppresses p34cdc2 expression and normal p53 suppresses cyclin A
expression. Exp Cell Res. 210:94–101. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jin CY, Moon DO, Lee JD, et al:
Sulforaphane sensitizes tumor necrosis factor-related
apoptosis-inducing ligand-mediated apoptosis through downregulation
of ERK and Akt in lung adenocarcinoma A549 cells. Carcinogenesis.
28:1058–1066. 2007. View Article : Google Scholar
|
|
28
|
Moody CA and Laimins LA: Human
papillomavirus oncoproteins: pathways to transformation. Nat Rev
Cancer. 10:550–560. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bak Y, Ham S, Baatartsogt O, et al: A1E
inhibits proliferation and induces apoptosis in NCI-H460 lung
cancer cells via extrinsic and intrinsic pathways. Mol Biol Rep.
40:4507–4519. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Doorbar J: Molecular biology of human
papillomavirus infection and cervical cancer. Clin Sci.
110:525–541. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Horinaka M, Yoshida T, Shiraishi T, et al:
Luteolin induces apoptosis via death receptor 5 upregulation in
human malignant tumor cells. Oncogene. 24:7180–7189. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhao J, Chen X, Lin W, et al: Total
alkaloids of Rubus aleaefolius Poir inhibit hepatocellular
carcinoma growth in vivo and in vitro via activation
of mitochondrial-dependent apoptosis. Int J Oncol. 42:971–978.
2013.
|