Introduction
Although considerable advances in scientific
knowledge and technology have been made, the use of animals in
research is considered essential for understanding the disease
mechanisms and evaluating the safety and efficacy of potential new
therapeutic strategies. For animal welfare, different strategies
for replacement, refinement and reduction of animals in scientific
experiments have been suggested. For reduction and replacement of
tumor transplantation on immunodeficient mice, we recently
established the method of tumor transplantation on fertilized
chicken eggs in our laboratory (1).
According to the German animal law, the chick embryo is not
considered a living animal, and thus is a unique model that
overcomes many limitations to studying the biology of cancer in
vivo. Similarly, the American Institutional Animal Care and Use
Committee (IACUC) allows the use of the chick embryo for research
experiments without the submission of an animal application until
day 18 of development. The chorioallantoic membrane (CAM), a
well-vascularized extra-embryonic tissue located underneath the
eggshell, has a successful history as a biological platform for the
molecular analysis of cancer (2,3),
although it is not yet generally accepted. The chick embryo is
naturally immunodeficient and the CAM readily supports the
engraftment of both normal and tumor tissues (4). According to our experience, human
freshly resected tumor tissue grows even faster on eggs compared to
immunodeficient mice. In addition to tumor growth, the CAM
successfully supports the development of a characteristic tumor
microenvironment and the detection of invasion, migration and
angiogenesis. Other advantages are the complete environment in
vivo, the possibility to easily study therapeutic effects to
tumor take, growth and expression of signaling molecules along with
side-effects, since the response of CAM tissue and chick embryo is
similar to the mammalian response. A further advantage is the price
and easy handling, since one fertilized chicken egg costs 25 cents
and an animal house or a special room are not necessary. Tumor
transplantation to eggs can be performed in a normal laboratory
room equipped with small-sized breeding incubators. Therefore, we
routinely use this animal replacement model for short-term in
vivo evaluation of new therapeutic options for pancreatic
ductal adenocarcinoma (PDA).
PDA is the most common type of pancreatic malignancy
and it is usually diagnosed at an advanced state due to its
untypical symptoms and aggressive behavior. PDA is characterized by
an intense tumor stroma with pronounced hypoxic microenvironment
(5), extensive local invasion,
early systemic dissemination and resistance to chemo- and
radiotherapy. The hypoxic microenvironment results in high
expression of the hypoxia marker CAIX and the hypoxia-inducible
factor HIF-1α in patient tissue with poor clinical outcome
(6). In experimental studies,
hypoxia indicates aggressive growth and spontaneous metastasis in
pancreatic cancer xenografts (7).
Most importantly, hypoxia is supposed to promote invasion and
metastasis and is involved in enrichment of cancer stem cells
(CSCs).
To study if new therapeutic options can overcome the
pronounced tumor hypoxia of PDA, a method for induction of hypoxia
in fertilized chicken eggs is urgently needed. Since our
preliminary results suggest that the conventional in vitro
method of gas hypoxia using 1% O2, 5% CO2 and
94% N2 is not suited for the chicken model, we employed
chemical induction of hypoxia by cobalt chloride
(CoCl2). Previous publications have shown that exposure
to cobalt promotes a response similar to hypoxia due to the
stabilization of HIF-1α (8). Thus,
CoCl2 is a hypoxia mimetic agent, which has been
frequently used for in vitro induction of hypoxia (9).
In the present study, we demonstrated that
CoCl2 is well suited for the induction of hypoxia in PDA
tumor xenografts growing on fertilized chicken eggs. To study
therapeutic intervention with tumor hypoxia in the animal
replacement model, we used the herb-derived triptolide, which is
the major active substance of the vine-like Tripterygium
wilfordii plant. In Chinese traditional medicine (TCM), this
herb is used for the treatment of rheumatoid arthritis and cancer
due to its anti-inflammatory and anti-carcinogenic properties
(10).
Materials and methods
Tumor cell line
The established PDA cell line BxPc-3 was obtained
from the American Type Culture Collection (Manassas, VA, USA). The
cells were recently authenticated by a commercial service
(Multiplexion, Heidelberg, Germany). Mycoplasma-negative cultures
were ensured by monthly mycoplasma tests. The cells were cultured
in DMEM medium (PAA, Pasching, Austria) and supplemented with 10%
heat-inactivated FCS (Sigma, Deisenhofen, Germany) and 25 mmol/l
HEPES (PAA).
Treatment of cells
Triptolide (PG-490) was obtained from Sigma-Aldrich
(St. Louis, MO, USA). The purity of triptolide was 98% (HPLC), and
a 10 mM stock solution was prepared in DMSO. CoCl2 was
obtained from Sigma-Aldrich. The 100 mM stock solution was prepared
in phosphate-buffered saline (PBS).
Transplantation of tumor cells to the CAM
of fertilized chicken eggs
This method was performed as previously described
with modifications (4). Fertilized
white Leghorn chicken eggs (Geflügelzucht Hockenberger, Eppingen,
Germany) were incubated in a digital motor breeders Type 168/D
(Siepmann GmbH, Herdecke, Germany) at a humidity of 55–60% at
37.8°C. The embryo with the CAM was detached at day 4 of embryonic
development by removing 2–3 ml albumen with a syringe. Then, a
small hole was cut into the eggshell, 1 ml albumen was restituted
and the hole was sealed with tape (Leukosilk; BSN Medical GmbH). At
day 9 of embryonic development the tumor cells were transplanted on
the CAM. Therefore small handmade rings from Thermanox™ cover discs
(Thermo Scientific, Schwerte, Germany) were placed on the CAM, the
CAM was scratched with a 27 gauge needle and 5×105
BxPc-3 were mixed 1:1 with Matrigel (BD Biosciences) and deposited
into the rings. Whatman filter paper was placed to the CAM directly
adjacent to the plastic ring containing the tumor cells.
CoCl2, triptolide or PBS were applied via dropping to
the filter paper until saturation. At day 18, the embryos were
humanely euthanized by injection of a barbiturate into the breast
muscle. Tumors and the liver of the chicken embryos were resected
and the tumor volumes were calculated with the formula: Volume =
4/3 × π × r3 (r = 1/2 × square root of diameter 1 ×
diameter 2) (11). Tumor tissue was
embedded in Tissue-Tek O.C.T. compound (Sakura, Zoeterwoude, The
Netherlands) on dry ice and stored at −20°C.
Immunofluorescence and H&E
staining
Frozen xenograft (6 μm) of embryonal liver sections
were fixed in ice-cold acetone at −20°C for 10 min and blocked with
10% normal goat serum in PBS with 0.2% Tween-20 for 30 min at room
temperature. Incubation with primary and secondary antibodies was
performed using standard procedures (1). The primary mouse monoclonal antibodies
directed toward Ki67, cytokeratin 19 (both from Abcam, Cambridge,
UK), EpCAM [kindly provided by G. Moldenhauer and published
(12)], CD24 [(SWA11; kindly
provided by P. Altevogt and published (13)], and the rabbit polyclonal antibodies
carbonic anhydrase IX (CAIX; sc-25599; Santa Cruz Biotechnology,
Heidelberg, Germany), CD44 (GeneTex, Nottingham, UK), Sox2 (Abcam),
and CD133 (Millipore, Darmstadt, Germany) were used. The secondary
antibodies were goat anti rabbit Alexa Fluor 594 IgG or goat
anti-mouse Alexa Fluor 594 IgG (Invitrogen, Camarillo, CA, USA).
Nuclei were counterstained with 4,6-diamidino-2-phenylindol (DAPI;
1 μg/ml). Primary and secondary antibody was repeated with the
respective antibodies. The samples were mounted in Fluoromount-G
(Southern Biotech, Birmingham, AL, USA) and stored at −20°C in the
dark. Liver sections of chick embryo were stained with hematoxylin
and eosin (H&E; Dako, Denmark), mounted in Pro Tags Mount Aqua
(Quartett, Berlin, Germany) and stored at room temperature.
Representative images were captured with a Leica DMRB fluorescence
microscope (Leica, Wetzlar, Germany) at ×400 magnification with a
SPOT™ Flex 15.2 64 Mp shifting pixel digital color camera
(Diagnostic Instruments; and analyzed with SPOT Basic Advanced
software 4.6).
Statistical analysis
The quantitative data of chick embryo weight are
presented as the means ± SD. The tumor growth data were analyzed
using Student’s t-test for statistical significance. P<0.05 was
considered statistically significant.
Results
Conventional gas hypoxia leads to rapid
death of chick embryos
To study if new therapeutic options can overcome the
pronounced tumor hypoxia of PDA, we tried to adapt the conventional
in vitro method of gas hypoxia to chicken eggs. The eggs
were placed into a hypoxia chamber, which was flushed with a gas
mixture of 1% O2, 5% CO2 and 94%
N2. However, after 24 h, even after only 30 min, all
chicken embryos were dead (data not shown). Therefore, this method
of hypoxia induction is not suited for the chicken egg model.
Cobalt chloride induces expression of
CAIX in PDA xenografts on fertilized chicken eggs
To study whether cobalt chloride may be more suited
for induction of hypoxia in PDA xenografts on fertilized chicken
eggs, we transplanted BxPc-3 cells to the CAM of fertilized chicken
eggs at day 9 of incubation. At day 11, eggs with a developed and
living embryo were selected and divided into 4 groups. Hypoxia was
induced in half of the eggs at days 11, 14, 15, 16 and 17 by
dropping CoCl2 until saturation to a small filter paper,
which was placed directly adjacent to the tumor xenograft (Fig. 1A). Similarly, triptolide was applied
via the filter paper at days 12 and 15 and the controls received
PBS only. At day 18, the embryos were humanely euthanized by
injection of a barbiturate into the blood vessels of CAM, followed
by resection of the tumor xenografts. CoCl2 reduced the
tumor volume from 0.2 cm3 in the control to 0.05
cm3 (Fig. 1B).
Triptolide further reduced the tumor growth under normoxic and
hypoxic conditions below 0.01 and 0.02 cm3,
respectively. Most importantly, CoCl2 induced the
expression of CAIX, which was measured by staining xenograft
sections with a specific antibody and fluorescence microscopy
(Fig. 1C). These results indicate
that CoCl2 induced hypoxia-induced signaling, which was
abolished by triptolide.
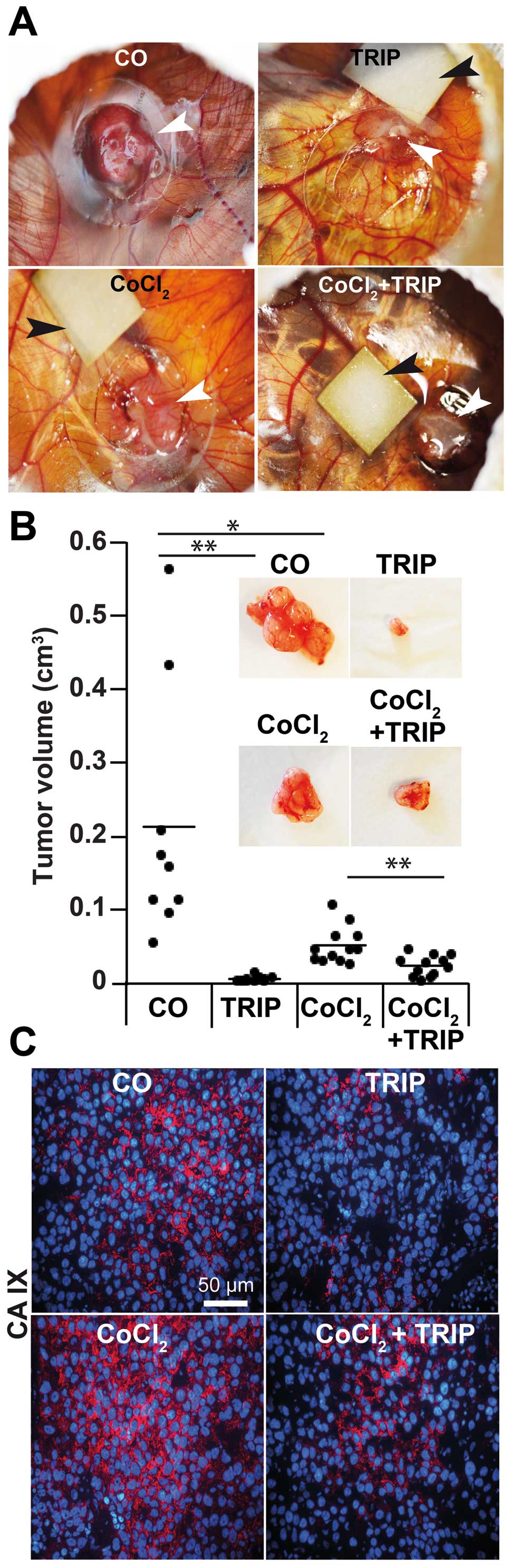 | Figure 1Cobalt chloride induces expression of
CAIX in PDA xenografts on fertilized chicken eggs. (A) BxPc-3 cells
(5×105) were transplanted in Matrigel on the CAM of
fertilized chicken eggs at day 9 of embryonic development. Hypoxia
was induced by dropping 10 μl CoCl2 (100 μM) to a
Whatman filter paper (0.5 cm2) placed directly adjacent
to the tumor xenografts at days 11, 14, 15, 16 and 17 until
saturation. For therapeutic treatment, 10 μl triptolide (50 nm,
TRIP) was applied to the filter paper at developmental days 12 and
15. PBS was applied as control instead of triptolide or
CoCl2. White arrows point to the tumor xenografts and
black bars point to the filter papers. (B) The xenografts were
resected at day 18 and the tumor diameters were measured with
calipers. The tumor volumes were calculated as described in
Materials and methods. The volumes of the individual tumors per
group are presented as black dots, and the bars indicate the
average tumor size of each group. Representative images of the
resected tumors are shown in the upper panel.
**P<0.001; *P<0.05. (C) Sections of the
xenograft tissue of each group were stained with the hypoxia marker
CAIX, followed by counterstaining of nuclei with DAPI. Positive
signals were detected by fluorescence microscopy. The bar indicates
50 μm. CAIX, carbonic anhydrase IX; PDA, pancreatic ductal
adenocarcinoma; CAM, chorioallantoic membrane; CoCl2,
cobalt chloride. |
Cobalt chloride is well tolerated by
chick embryos
In order to detect putative but unwanted
side-effects of CoCl2, we analyzed developmental
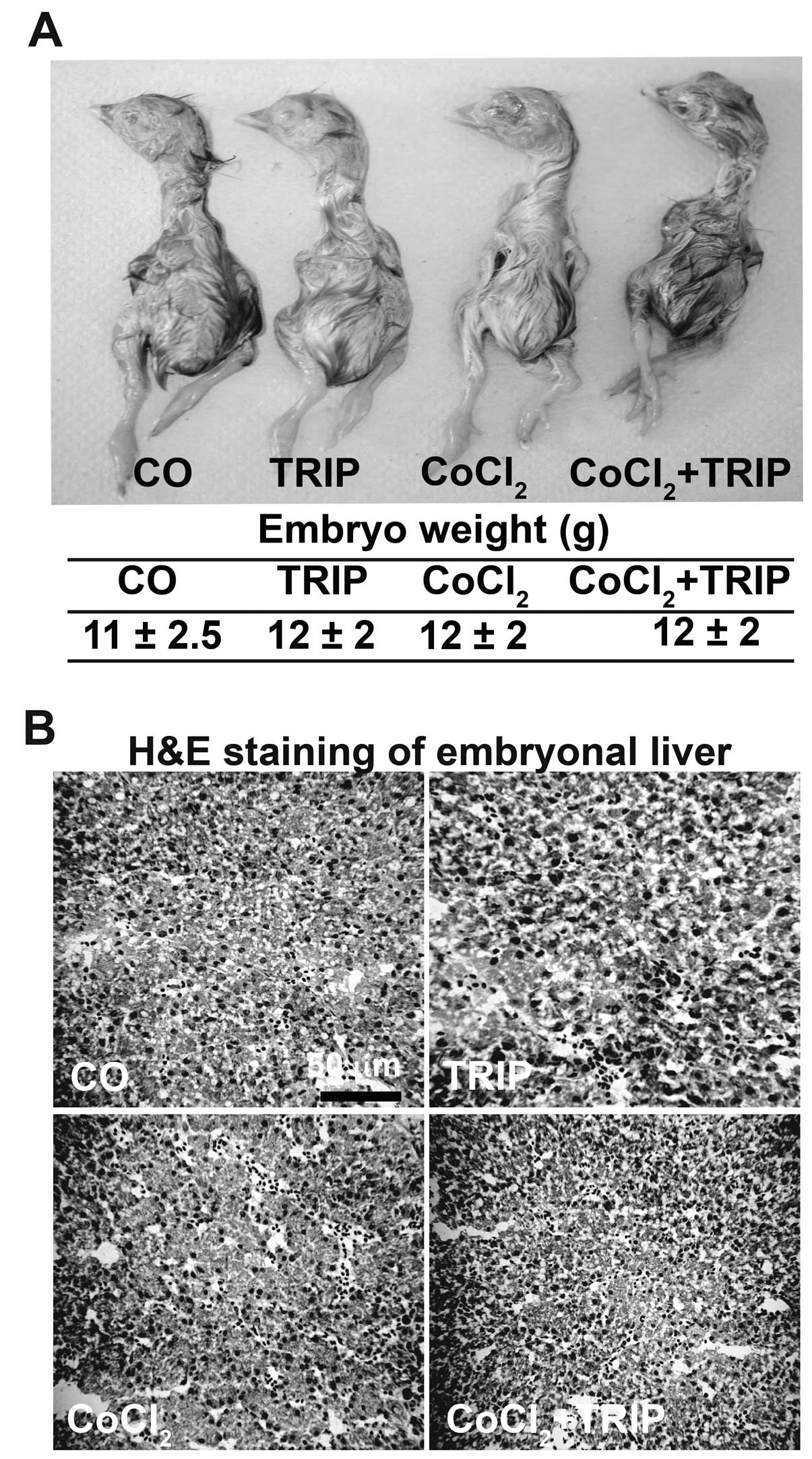
defects, morphology and the weight of embryos of each group. We did
not observe abnormality, the embryos looked well developed and had
a regular weight in each group (Fig.
2A). For detection of toxic side-effects to the liver, we
stained frozen liver tissue sections with H&E and examined the
tissue by microscopy (Fig. 2B). No
necrosis was detectable, suggesting that CoCl2 did not
induce toxic side-effects. Notably, triptolide alone or combined
with CoCl2 did not influence development, weight or
liver tissue either. Thus, CoCl2-induced hypoxia, alone
or in combination with triptolide, does not induce toxic
side-effects in chick embryos.
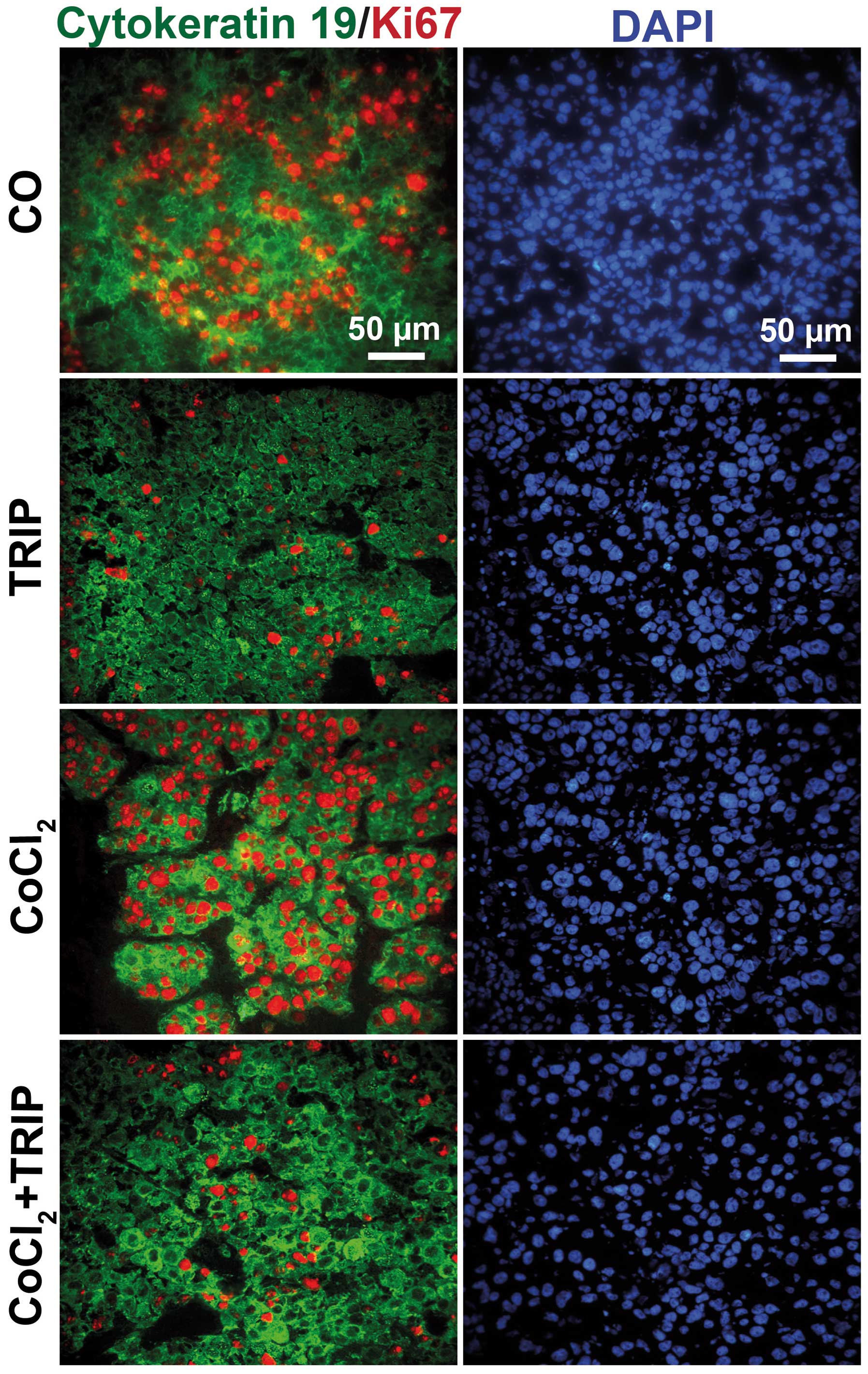
Xenograft tissue consists of human cells
and proliferation is inhibited by triptolide
To ensure that the xenograft tumors consisted of
human cells, we stained tumor sections with human-specific
antibodies to detect proliferation by Ki67 and the cytoskeletal
marker cytokeratin 19 (Fig. 3).
Whereas both expression of cytokeratin 19 and Ki67-positive
proliferating human tumor cells were detected in control and
CoCl2-treated xenografts, triptolide abolished Ki67
expression, but had no effect on cytokeratin expression. This
control experiment confirmed that the xenograft tissue consisted of
human tumor cells and that CoCl2 did not decrease the
proliferation, which was, however, strongly inhibited by
triptolide.
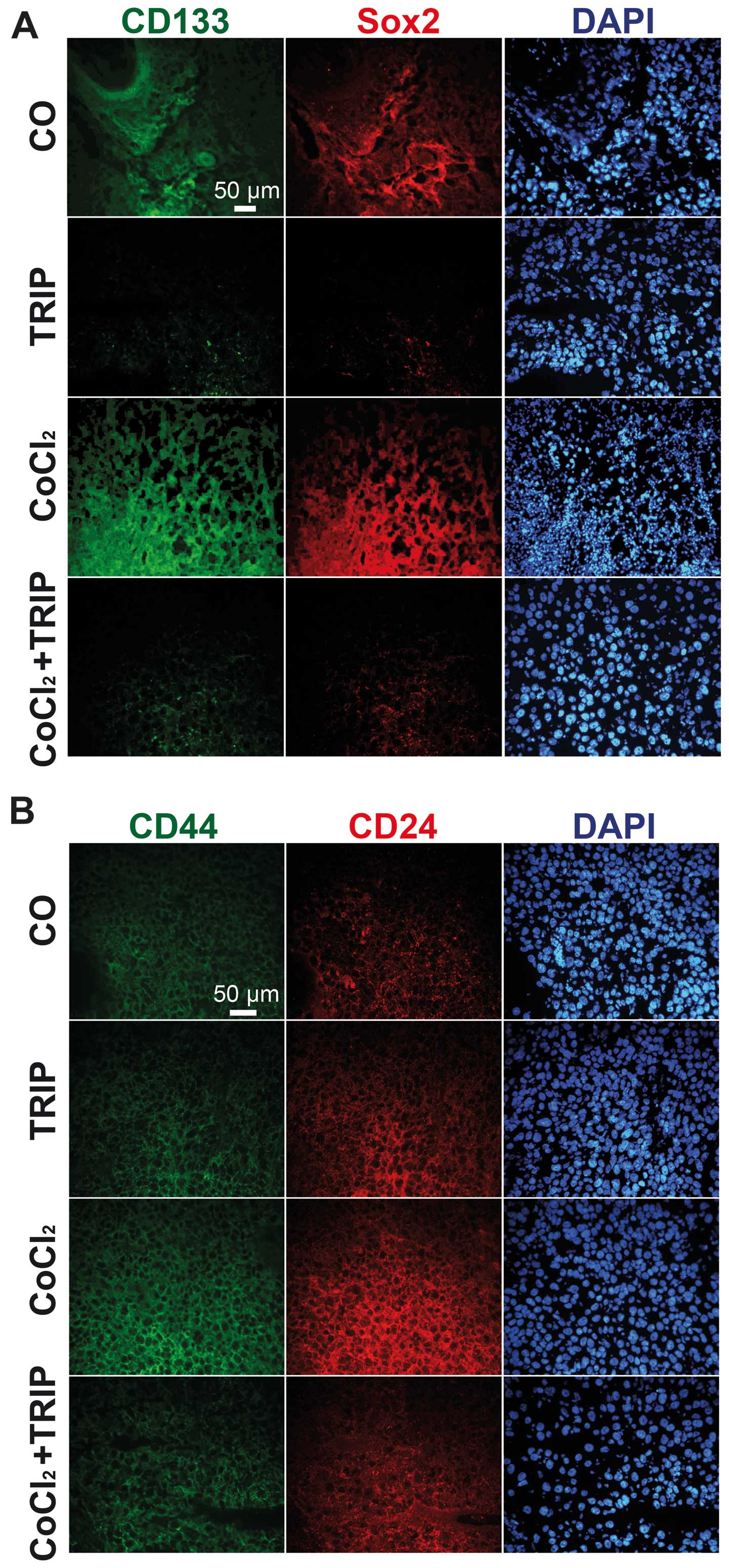
Cobalt chloride induces the expression of
the CSC markers CD133, Sox2, CD44 and CD24
Our recent data suggest that intrinsic tumor hypoxia
and the in vitro induction of gas hypoxia increases the
expression of CSC markers (1,14).
Therefore, we evaluated, by immunohistochemistry, whether
CoCl2 may have the same effect. Indeed, we detected
induction of CD133 and Sox2 (Fig.
4A) as well as induction of CD44 and CD24 in xenograft tissue
derived from eggs with CoCl2 treatment (Fig. 4B). In contrast, triptolide alone did
not induce the expression of CSC markers and triptolide
co-treatment of CoCl2-treated eggs completely abolished
the expression of CSC markers. These results confirm the successful
induction of hypoxia in tumor xenografts on fertilized chicken eggs
by application of CoCl2 to a Whatman filter paper
directly adjacent to the tumor xenograft.
Discussion
The aim of the present study was to establish a
method for induction of hypoxia in xenografted fertilized chicken
eggs without inducing toxic side-effects. However, our preliminary
results demonstrated that the application of gas hypoxia is not
suited as the chick embryo does not survive when eggs are incubated
in a hypoxia chamber. The question is whether the induction of
hypoxia in vivo is necessary, since due to the fast tumor
xenograft growth, hypoxia is automatically induced, as we recently
demonstrated by positive CAIX staining of PDA xenografts growing on
chicken eggs (1). Although these
results are confirmed by the present data, we were able to further
increase the expression of CAIX and thus hypoxic signaling by
CoCl2-treatment. Considering the pronounced hypoxic
tumor microenvironment in patient tumors of PDA (5), this situation may reflect the tumor
conditions in patients and thus may be considered as a suited model
for experimental evaluation of PDA. This suggestion is underlined
by our finding that CoCl2-induced hypoxia did not induce
toxic side-effects in the chick embryo.
Recent studies suggest that hypoxia increases the
aggressiveness of tumor growth (7).
This finding is, however, not confirmed by the present data, since
the tumor volume measured for CoCl2-treated cells was
smaller compared to untreated cells. This may be due to the fact
that the transplanted BxPc-3 cells are less aggressive and are
therefore sensitive to hypoxia, as we demonstrated in our recent
study (14). In the present study,
in vitro gas hypoxia led to increased self-renewal potential
in highly aggressive PDA cell lines, but not in the less aggressive
BxPc-3 cells. This may be due to a low percentage of CSCs, which
are present in BxPc-3 cells (15).
In this respect, our recent study demonstrated, that CSCs resist
tumor hypoxia and start to proliferate and invade, in contrast to
normal, more differentiated tumor cells (1,14,16).
Indeed, we observed the enhanced expression of the CSC markers
CD133, Sox2, CD44 and CD24 in our model, which suggests the
possibility that hypoxia induces dormant CSCs within the mixed
tumor population of BxPc-3 cells.
A previous study by Lester et al (17) already used the application of
CoCl2-induced hypoxia to fertilized chicken eggs
transplanted with breast cancer cells. However, in this former
study, the induction of hypoxia was not controlled and it was
unclear if CoCl2 was effective. Also, the conditions of
CoCl2 application in this former study were different
from ours, since Lester et al applied 25 μl of a 100 μM
CoCl2 solution daily between days 9 and 18. In contrast,
our data show that a less frequent application of CoCl2
(days 11, 14, 15, 16 and 17) is sufficient for a pronounced
induction of hypoxia. Our results clearly demonstrate the induction
of hypoxia since we found enhanced expression of CAIX and the CSC
markers Sox2, CD133, CD44 and CD24.
Finally, we demonstrated that triptolide, which was
used as a treatment control, did not induce hypoxic signaling, but
inhibited the CoCl2-mediated hypoxic effects. These
results are in line with our previous findings, which demonstrated
that triptolide strongly reverts tumor hypoxia-induced progression
of PDA without inducing side-effects (1).
Taken together, CoCl2 is suitable to
induce hypoxia in PDA xenografts growing on the chick embryo animal
replacement model, which is a promising tool for future in
vivo experimental evaluation of therapeutic strategies.
Acknowledgements
The authors thank Dr W. Gross for calculation of
statistics and J. Gladkich for the technical assistance. This study
was supported by grants from the German Cancer Aid (Deutsche
Krebshilfe 109362), the German Research Community (DFG HE
3186/11-1), the Federal Ministry of Education and Research (BMBF
031A213), the German-Israeli Foundation for Scientific Research and
Development (GIF 1058-7.11/2008), the Heidelberger Stiftung
Chirurgie, the Stiftung für Krebs und Scharlachforschung, the Hanns
A. Pielenz Stiftung and the Dietmar Hopp-Stiftung.
Abbreviations:
|
CAM
|
chorioallantoic membrane
|
|
CSC
|
cancer stem cell
|
|
EMT
|
epithelial-mesenchymal transition
|
|
CoCl2
|
cobalt chloride
|
|
PDA
|
pancreatic ductal adenocarcinoma
|
References
|
1
|
Liu L, Salnikov AV, Bauer N,
Aleksandrowicz E, Labsch S, Nwaeburu C, Mattern J, Gladkich J,
Schemmer P, Werner J and Herr I: Triptolide reverses
hypoxia-induced EMT and stem-like features in pancreatic cancer by
NF-κB downregulation. Int J Cancer. 134:2849–2503. 2014.PubMed/NCBI
|
|
2
|
Ribatti D, Nico B, Vacca A and Presta M:
The gelatin sponge-chorioallantoic membrane assay. Nat Protoc.
1:85–91. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Schneiderhan W, Scheler M, Holzmann KH,
Marx M, Gschwend JE, Bucholz M, Gress TM, Seufferlein T, Adler G
and Oswald F: CD147 silencing inhibits lactate transport and
reduces malignant potential of pancreatic cancer cells in in vivo
and in vitro models. Gut. 58:1391–1398. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Janse EM and Jeurissen SH: Ontogeny and
function of two non-lymphoid cell populations in the chicken
embryo. Immunobiology. 182:472–481. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Brown JM and Giaccia AJ: The unique
physiology of solid tumors: opportunities (and problems) for cancer
therapy. Cancer Res. 58:1408–1416. 1998.PubMed/NCBI
|
|
6
|
Hoffmann AC, Mori R, Vallbohmer D,
Brabender J, Klein E, Drebber U, Baldus SE, Cooc J, Azuma M,
Metzger R, Hoelscher AH, Danenberg KD, Prenzel KL and Danenberg PV:
High expression of HIF1α is a predictor of clinical outcome in
patients with pancreatic ductal adenocarcinomas and correlated to
PDGFA, VEGF, and bFGF. Neoplasia. 10:674–679.
2008.
|
|
7
|
Chang Q, Jurisica I, Do T and Hedley DW:
Hypoxia predicts aggressive growth and spontaneous metastasis
formation from orthotopically grown primary xenografts of human
pancreatic cancer. Cancer Res. 71:3110–3120. 2011. View Article : Google Scholar
|
|
8
|
Ho VT and Bunn HF: Effects of transition
metals on the expression of the erythropoietin gene: further
evidence that the oxygen sensor is a heme protein. Biochem Biophys
Res Commun. 223:175–180. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gray MJ, Zhang J, Ellis LM, Semenza GL,
Evans DB, Watowich SS and Gallick GE: HIF-1α, STAT3, CBP/p300 and
Ref-1/APE are components of a transcriptional complex that
regulates Src-dependent hypoxia-induced expression of VEGF in
pancreatic and prostate carcinomas. Oncogene. 24:3110–3120.
2005.
|
|
10
|
Brinker AM, Ma J, Lipsky PE and Raskin I:
Medicinal chemistry and pharmacology of genus Tripterygium
(Celastraceae). Phytochemistry. 68:732–766. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Balke M, Neumann A, Szuhai K, Agelopoulos
K, August C, Gosheger G, Hogendoorn PC, Athanasou N, Buerger H and
Hagedorn M: A short-term in vivo model for giant cell tumor of
bone. BMC Cancer. 11:2412011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Moldenhauer G, Momburg F, Möller P,
Schwartz R and Hämmerling GJ: Epithelium-specific surface
glycoprotein of Mr 34,000 is a widely distributed human carcinoma
marker. Br J Cancer. 56:714–721. 1987. View Article : Google Scholar
|
|
13
|
Weber E, Lehmann HP, Beck-Sickinger AG,
Wawrzynczak EJ, Waibel R, Folkers G and Stahel RA: Antibodies to
the protein core of the small cell lung cancer workshop antigen
cluster-w4 and to the leucocyte workshop antigen CD24 recognize the
same short protein sequence leucine-alanine-proline. Clin Exp
Immunol. 93:279–285. 1993. View Article : Google Scholar
|
|
14
|
Rausch V, Liu L, Apel A, Rettig T,
Gladkich J, Labsch S, Kallifatidis G, Kaczorowski A, Groth A, Gross
W, Gebhard MM, Schemmer P, Werner J, Salnikov AV, Zentgraf H,
Büchler MW and Herr I: Autophagy mediates survival of pancreatic
tumour-initiating cells in a hypoxic microenvironment. J Pathol.
227:325–335. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kallifatidis G, Rausch V, Baumann B, Apel
A, Beckermann BM, Groth A, Mattern J, Li Z, Kolb A, Moldenhauer G,
Altevogt P, Wirth T, Werner J, Schemmer P, Büchler MW, Salnikov A
and Herr I: Sulforaphane targets pancreatic tumour-initiating cells
by NF-κB-induced antiapoptotic signalling. Gut. 58:949–963.
2009.PubMed/NCBI
|
|
16
|
Salnikov AV, Liu L, Platen M, Gladkich J,
Salnikova O, Ryschich E, Mattern J, Moldenhauer G, Werner J,
Schemmer P, Büchler MW and Herr I: Hypoxia induces EMT in low and
highly aggressive pancreatic tumor cells but only cells with cancer
stem cell characteristics acquire pronounced migratory potential.
PLoS One. 7:e463912012. View Article : Google Scholar
|
|
17
|
Lester RD, Jo M, Montel V, Takimoto S and
Gonias SL: uPAR induces epithelial-mesenchymal transition in
hypoxic breast cancer cells. J Cell Biol. 178:425–436. 2007.
View Article : Google Scholar : PubMed/NCBI
|