Introduction
Nasopharyngeal carcinoma (NPC) is one of the most
common carcinomas in Southern China, particularly in Guangdong
Province; however, it is rare in most other regions of the world
(1,2). NPC cells are highly malignant, often
invading adjacent regions and metastasizing to regional lymph nodes
and distant organs. Despite radiation therapy or adjunctive
chemotherapy, 30–40% of NPC patients die from local recurrence and
distant metastasis (3). A better
understanding of the molecular mechanisms contributing to NPC
progression may provide new insights into possible therapeutic
targets.
Increasing evidence indicates that metastasis of
many epithelium-derived tumors is closely correlated with
epithelial-to-mesenchymal transition (EMT) (4–7). EMT
is characterized by cells losing their epithelial morphology, and
acquiring fibroblast-like properties. They also exhibit reduced
intercellular adhesion and increased motility (8). Loss of E-cadherin expression and
increased expression levels of mesenchymal cell markers such as
vimentin, N-cadherin, fibronectin and α smooth muscle actin (α-SMA)
appear to be strongly correlated with EMT. Suppression of
E-cadherin by transcriptional regulators such as Snail or Twist has
emerged as a critical step driving EMT (9). Previously, it was reported that cancer
cells could acquire epithelial-to-mesenchymal reverting transition
(EMrT) to adapt to metastatic microenvironments, with re-expression
of E-cadherin, an indicator of EMrT (10,11).
Under such conditions, cancer cells continue to display phenotypic
plasticity beyond the EMT that initiates metastasis. To date,
little is known about the precise mechanisms, biology, and clinical
significance of EMrT in NPC.
Previous studies revealed that several signaling
pathways are involved in EMT and EMrT (9,12,13).
The phosphatidylinositol-3 OH kinase/protein kinase B (PI3K/Akt)
signaling pathway is constitutively activated in various epithelial
cancers, and plays an important role in tumor formation and
metastasis (14,15). Phosphatidylinositol
3,4,5-triphosphate (PIP3), generated from PI3K, acts as a lipid
second messenger essential for the translocation of Akt to the
plasma membrane. Akt is phosphorylated at two sites, T308 in the
kinase domain and S473 in the regulatory tail. Phosphorylation at
T308 and S473 is essential for maximal Akt activation.
Phosphorylated Akt (p-Akt) regulates the function of a broad array
of intracellular proteins involved in fundamental processes,
including cell proliferation, cell death, cell motility/adhesion,
cell transformation, neovascularization and the inhibition of
apoptosis. It has been reported that Akt suppresses transcription
of the E-cadherin gene (10,16).
This transcriptional suppression induces responses leading to the
conversion of epithelial cells into invasive mesenchymal cells.
A chemical inhibitor of PI3K,
2–4-morpholinyl-8-phenylchromone (LY294002), has been extensively
used to study the role of the PI3K/AKT pathway in normal and
transformed cells. Inactivation of PI3K using LY294002 has been
shown to lead to the dephosphorylation of Akt at T308 and S473. A
consequence of this is induction of G1 arrest during cell growth,
which leads to apoptosis. Inhibitors of PI3K also possess antitumor
activity in vitro and in vivo in a variety of tumors.
It is possible that cells constitutively expressing active Akt
become dependent on its ability to promote survival. Although these
results have been observed in many human cancers, the effects of
LY294002 on EMrT and metastasis in human NPC remain unclear. In the
present study, we investigated the role of PI3K/Akt signaling
activation and inhibition in EMrT for human NPC cases.
Materials and methods
Sample collection and ethics
Human benign nasopharyngeal samples (chronic
nasopharyngitis, n=20) and NPC samples (n=130) were obtained from
the Affiliated Gaozhou Hospital, and the Affiliated Hospital of
Guangdong Medical College. All NPC patients were diagnosed with
non-keratinizing carcinoma following histological examination.
Tissues were paraffin-embedded and sectioned (4-μm thickness). Of
the 130 NPC cases, 53 involved local lymph node metastasis; of
these 53 cases, 23 were paired with primary and metastatic lesions.
The use of human tissues in this study was approved by the Ethics
Council of the Affiliated Gaozhou Hospital, and the Affiliated
Hospital of Guangdong Medical College.
Immunohistochemistry (IHC)
Paraffin-embedded sections were used for
immunohistochemical analysis of p-Akt (Ser473), E-cadherin,
vimentin and α-SMA expression. Sections were deparaffinized and
rehydrated, then heat-induced antigen retrieval at 95°C was
conducted in sodium citrate buffer (10 mM, pH 6.0). Endogenous
peroxidases were blocked using 0.3% (v/v)
H2O2, while binding of non-specific proteins
was blocked with 10% goat serum. Sections were then incubated with
primary antibodies against p-Akt (Ser473) (1/200 dilution; Cell
Signaling, Danvers, MA, USA), E-cadherin (1/200) and vimentin
(1/200) (both from Cell Signaling), and α-SMA (1/100; Maixin Bio,
Fujiang, China) at 4°C overnight. Non-immune immunoglobulin G (IgG)
was used as a negative control. Antigenic sites were visualized
using a streptavidin-peroxidase (SP) and a 3,3′-diaminobenzidine
(DAB) kit (ZSGB-BIO, Beijing, China). Expression of p-Akt (Ser473),
vimentin, and α-SMA was localized to the cytoplasm, while
E-cadherin signals were observed in the cell membrane or cytoplasm.
Scoring of sections was conducted by two experienced pathologists,
blinded to the identity of samples. Staining intensity was
classified as 0 (negative); 1 (weak); 2 (moderate); and 3 (strong).
The proportion of p-Akt (Ser473)-, E-cadherin-, vimentin- and
α-SMA-positive cells was scored as 1 (0–9% positive); 2 (10–50%);
and 3 (>50%). Samples with a sum immunoreactive score (IRS) ≥1
were considered to be positive for p-Akt (Ser473), E-cadherin,
vimentin and α-SMA.
Cell culture
The poorly differentiated human NPC cell line,
CNE2Z, was maintained as described previously (17–19).
Briefly, CNE2Z cells were cultured in Dulbecco’s modified Eagle’s
medium (DMEM; Life Technologies, Guangzhou, China) supplemented
with 10% fetal bovine serum (FBS; Gibco), 100 U/ml penicillin and
100 μg/ml streptomycin at 37°C/5% CO2. LY294002 (Cayman
Chemical Co., Ann Arbor, MI, USA) was dissolved in dimethyl
sulfoxide (DMSO), and the final concentration of DMSO in growth
medium was 0.5% (v/v). CNE2Z cells were incubated in DMEM
containing 0.5% FBS for 24 h; media were then replaced with
complete growth medium containing LY294002 at 10, 25, 50 or 75 μM,
or DMSO (0.5%, v/v) for 48 h; cells were then harvested for use in
relevant experiments.
In vitro cell invasion and migration
assays
Cell invasion assays were conducted using Transwell
chambers containing membranes with 8 μm pores. The upper surfaces
of the Transwell chambers were coated with Matrigel matrix (250
μl/ml) overnight at 4°C, then rehydrated with 0.1% (w/v) bovine
serum albumin (BSA) in DMEM for 1 h at 37°C. Transwell chambers
were then placed into 24-well culture plates. CNE2Z cells
(5×105) were suspended in 200 μl of complete growth
media supplemented with LY294002 (10, 25, 50 or 75 μM), or DMSO
(0.5%, v/v), and were then added to the upper chambers and allowed
to migrate toward the underside of the membrane. Membranes were
fixed with 3.5% paraformaldehyde 24 h later; cells on the upper
surface of the membrane were removed by wiping with a cotton swab,
and then membranes were mounted onto glass slides. Cells on the
lower faces of the membranes were counted, with 20 random fields of
view per membrane counted for each assay. For cell migration
assays, the protocol was similar to that used in cell invasion
assays, except that Matrigel was not added to the upper
chambers.
Quantitative polymerase chain reaction
(qPCR) assays
The CNE2Z cells were treated with LY294002 (10, 25,
50 and 75 μM) and DMSO (0.5%, v/v) for 24 h. Total RNA was
extracted using Takara RNAiso Plus reagent (Takara Biotechnology
Co., Ltd., China). We used the Promega RT System and oligo(dT)18,
along with total RNA (1 μg) as a template to generate first strand
cDNA. Specific primer pairs for the amplification of E-cadherin
(5′-TTG CTA CTG GAA CAG GGA CAC-3′ and 5′-CCC GTG TGT TAG TTC TGC
TGT-3′), vimentin (5′-TGC GTG AAA TGG AAG AGA ACT-3′ and 5′-TCA GGT
TCA GGG AGG AAA AGT-3′), α-SMA (5′-TCC CTT GAG AAG AGT TAC GAG
TTG-3′ and 5′-ATG ATG CTG TTG TAG GTG GTT TC-3′), and β-actin
(5′-TGA CGT GGA CAT CCG CAA AG-3′ and 5′-CTG GAA GGT GGA CAG CGA
GG-3′) were synthesized by Sangon Biotech Co., Ltd. (Shanghai,
China). The total reaction (20 μl) comprised 10 μl of SYBR-Green I
PCR Master Mix (Roche), 0.8 μl of each primer (10 μM), 2 μl of cDNA
and 6.4 μl of double-distilled water. Thermal cycling involved an
initial denaturation step at 95°C for 5 min, followed by 45 cycles
at 95°C for 25 sec, and 60°C for 60 sec, in a LightCycler 480 II
instrument (Roche China Ltd., Shanghai, China). The relative
abundances of target mRNAs were determined from Ct values and
plotted as fold-change compared with the control group. The
transcription levels of β-actin served as a loading control.
Western blot analysis
Cells were homogenized with lysis buffer (1% Triton
X-100, 50 mM Tris-HCl pH 7.5, 0.1% SDS, 150 mM NaCl, 10% glycerol,
1.5 mM MgCl2, 1 mM PMSF, 0.1 mM NaV04, 0.1 mM
benzamidine, 5 μl/ml leupeptin and 5 μl/ml aprotinin), and protein
homogenates were separated by sodium dodecyl sulfate-polyacrylamide
gel electrophoresis (SDS-PAGE), and then transferred to
nitrocellulose membranes. Membranes were incubated with primary
antibodies against p-Akt (Ser473) (1:1,000 dilution), E-cadherin
(1:500), vimentin (1:500) (all from Cell Signaling), α-SMA (1:500;
Maxin Bio), and β-actin (1:1000; Santa Cruz, Dallas, TX, USA) at
4°C overnight. Membranes were washed twice and incubated with
horseradish peroxidase-conjugated secondary antibodies for 2 h at
room temperature. Protein bands were visualized using enhanced
chemiluminescent reagents (Thermo Fisher, Rockford, IL, USA) and
analyzed using an InGenius LHR Gel Documentation System (Syngene,
Frederick, MD, USA).
Tumor xenograft experiments
Specific pathogen-free (SPF) Balb/c null mice (6–8
weeks old) were purchased from the Experimental Animal Center of
Guangdong Medical College. Mice were housed in a facility with
controlled temperature, humidity, and a 12 h light/dark cycle. All
animal procedures were conducted in accordance with protocols
approved by the Institutional Animal Care and Use Committee (IACUC)
of Guangdong Medical College. CNE2Z cells (1×106
cells/mouse) were subcutaneously injected (200 μl) into the right
flanks of mice. After 1 week, tumor-bearing mice were
intraperitoneally injected twice a week with 0.5% DMSO (n=4) or
LY294002 (10, 25, 50 or 75 mg/kg; n=4 for each group) for 4 weeks.
Mice were monitored, with body weight and tumor sizes measured
twice a week. All mice were euthanized at the end of the treatment
period. Portions of the xenografts were fixed in natural formalin,
and embedded in paraffin for immunohistochemical analysis of EMT
markers. Xenografts that were not fixed were stored at −80°C. Lungs
of each mouse were collected, the intermittent sections (10
sections/lung, 20 sections/mouse) were stained with H&E and the
tumor metastatic lesions were calculated.
Statistical analysis
Statistical analyses were conducted using GraphPad
Prism (GraphPad Software, CA, USA). We used χ2 tests to
analyze and compare expression of p-Akt (Ser473) and EMT markers in
clinical samples. Data for in vitro experiments are
expressed as the means ± SD. Differences between two groups were
analyzed using Student’s t-test. For comparison of multiple groups,
ANOVAs were used followed by the Student-Newman-Keuls test.
P-values <0.05 were considered to indicate a statistically
significant difference.
Results
Activation of PI3K/Akt signaling and
expression of EMT markers in NPC
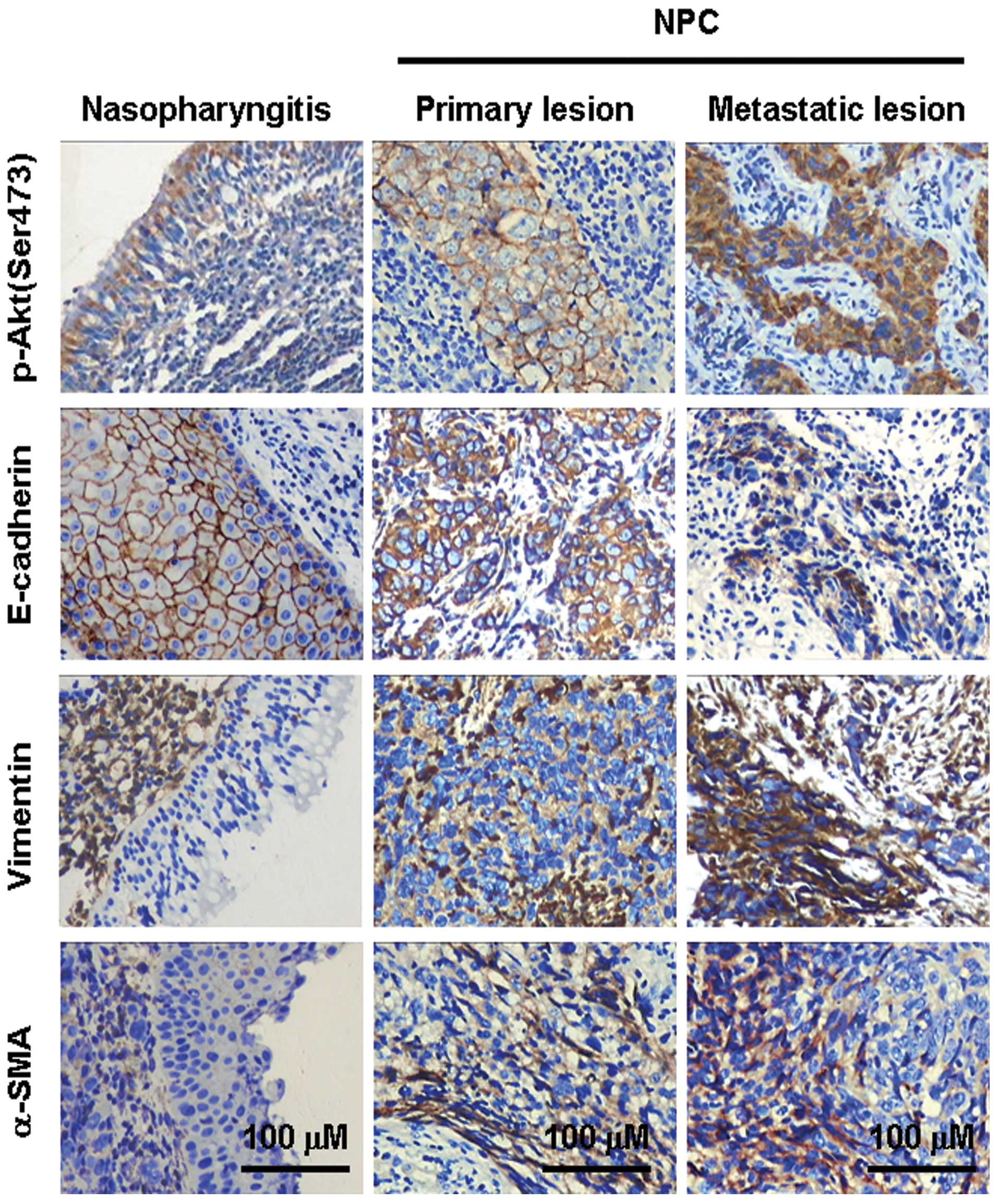
Positive expression of p-Akt (Ser473) in chronic
nasopharyngitis and NPC samples was observed in 25% (5/20) and
76.2% (99/130) of samples, respectively (P<0.001) (Table I, Fig.
1), indicating the activation of PI3K/Akt signaling in NPCs.
Expression levels of membrane E-cadherin, cytosol vimentin and
α-SMA proteins in chronic nasopharyngitis samples were 100%
(20/20), 0% and 0%, respectively. For NPC samples, expression
levels of these proteins were 75% (98/130), 41% (53/130) and 37%
(48/130) (P<0.05, 0.001 and 0.01, respectively). Spindle and
fibroblast-like tumor cells in NPC samples exhibited intense
vimentin and α-SMA staining. These results indicated that, when
compared with the benign chronic nasopharyngitis, the NPCs
presented a much stronger activation of PI3K/Akt signaling, obvious
downregulation in membrane E-cadherin and upregulation in cytosol
vimentin and α-SMA protein expressions.
 | Table IExpression of p-Akt (Ser473), membrane
E-cadherin, cytosol vimentin and α-SMA in clinical nasopharyngitis and NPC
samples. |
Table I
Expression of p-Akt (Ser473), membrane
E-cadherin, cytosol vimentin and α-SMA in clinical nasopharyngitis and NPC
samples.
| Conditions | n | p-Akt (Ser473) +
(%) | P-value | E-cadherin + (%) | P-value | Vimentin + (%) | P-value | α-SMA + (%) | P-value |
|---|
| Tissue type | | | <0.001 | | <0.05 | | <0.001 | | <0.01 |
| NPC | 130 | 99 (76.2) | | 98 (75.4) | | 53 (40.8) | | 48 (36.9) | |
| Nasopharyngitis | 20 | 5 (25.0) | | 20 (100.0) | | 0 | | 0 | |
| Metastasis | | | <0.01 | | <0.01 | | <0.001 | | <0.05 |
| With | 53 | 47 (88.7) | | 33 (62.3) | | 32 (60.4) | | 25 (47.2) | |
| Without | 77 | 52 (67.5) | | 65 (84.4) | | 21 (27.3) | | 23 (29.9) | |
| Matched lesion | | | <0.05 | | <0.05 | | <0.05 | | >0.05 |
| Primary | 23 | 10 (43.5) | | 18 (78.3) | | 15 (65.2) | | 7 (30.4) | |
| Secondary | 23 | 18 (78.3) | | 11 (47.8) | | 7 (30.4) | | 13 (56.5) | |
Activation of PI3K/Akt signaling and EMT
contribute to clinical NPC metastasis
Of the 130 NPC samples, 53 cases had signs of
cervical lymph node metastasis. High levels of p-Akt (Ser473)
expression were observed in cases with local lymph node metastasis
compared with those without metastasis [89% (47/53) vs. 68%
(52/77), P<0.01]. Expression levels of membrane E-cadherin,
cytosol vimentin and α-SMA in NPC samples with local lymph node
metastasis were 62% (33/53), 60% (32/53) and 47% (25/53), and 84%
(65/77), 27% (21/77) and 30% (23/77) in samples without local lymph
node metastasis (P<0.01, 0.001 and 0.05, respectively). To
further investigate the role of PI3K/Akt activation and EMT
occurrence in NPC metastasis, we analyzed the expression of p-Akt
(Ser473) and EMT genes in NPC samples matched with primary and
secondary lesions (n=23). Although expression of α-SMA was not
significantly different between primary and metastatic lesions,
higher p-Akt (Ser473) and vimentin expression levels, and lower
membrane E-cadherin expression levels, were observed in secondary
NPC metastatic lesions (P<0.05) (Table I).
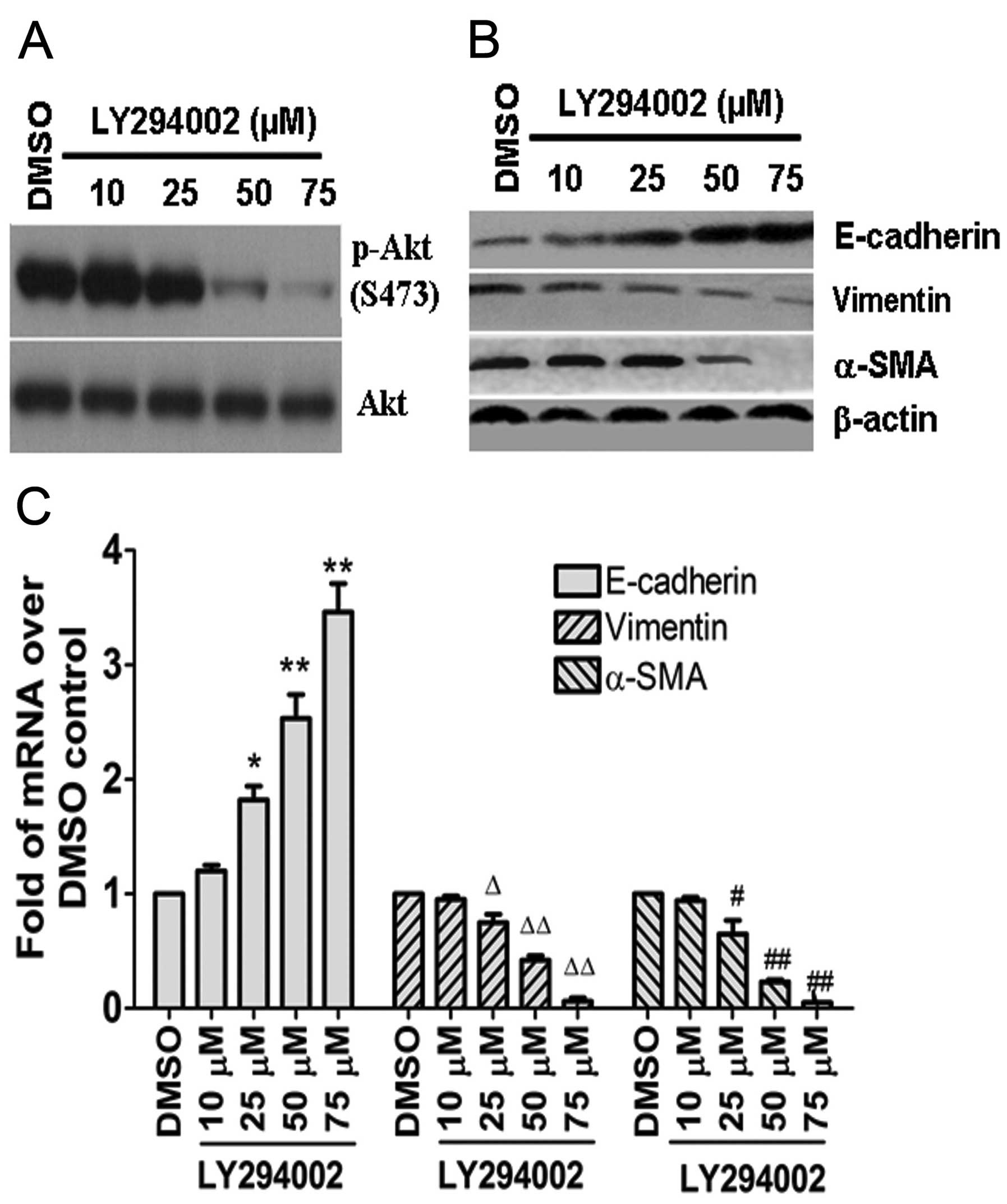
Inhibition of PI3K/Akt signaling alters
expression of p-Akt (Ser473) and EMT marker genes in CNE2Z cells in
vitro
Treatment of CNE2Z cells with LY294002 significantly
suppressed p-Akt (Ser473) expression in a concentration-dependent
manner; particularly at 75 μM, the expression of p-Akt (Ser473) was
almost completely attenuated (Fig.
2A). Treatment of CNE2Z cells with LY294002 led to upregulation
of E-cadherin and downregulation of vimentin and α-SMA at the
protein and mRNA levels (Fig. 2B and
C), and to EMrT in vitro.
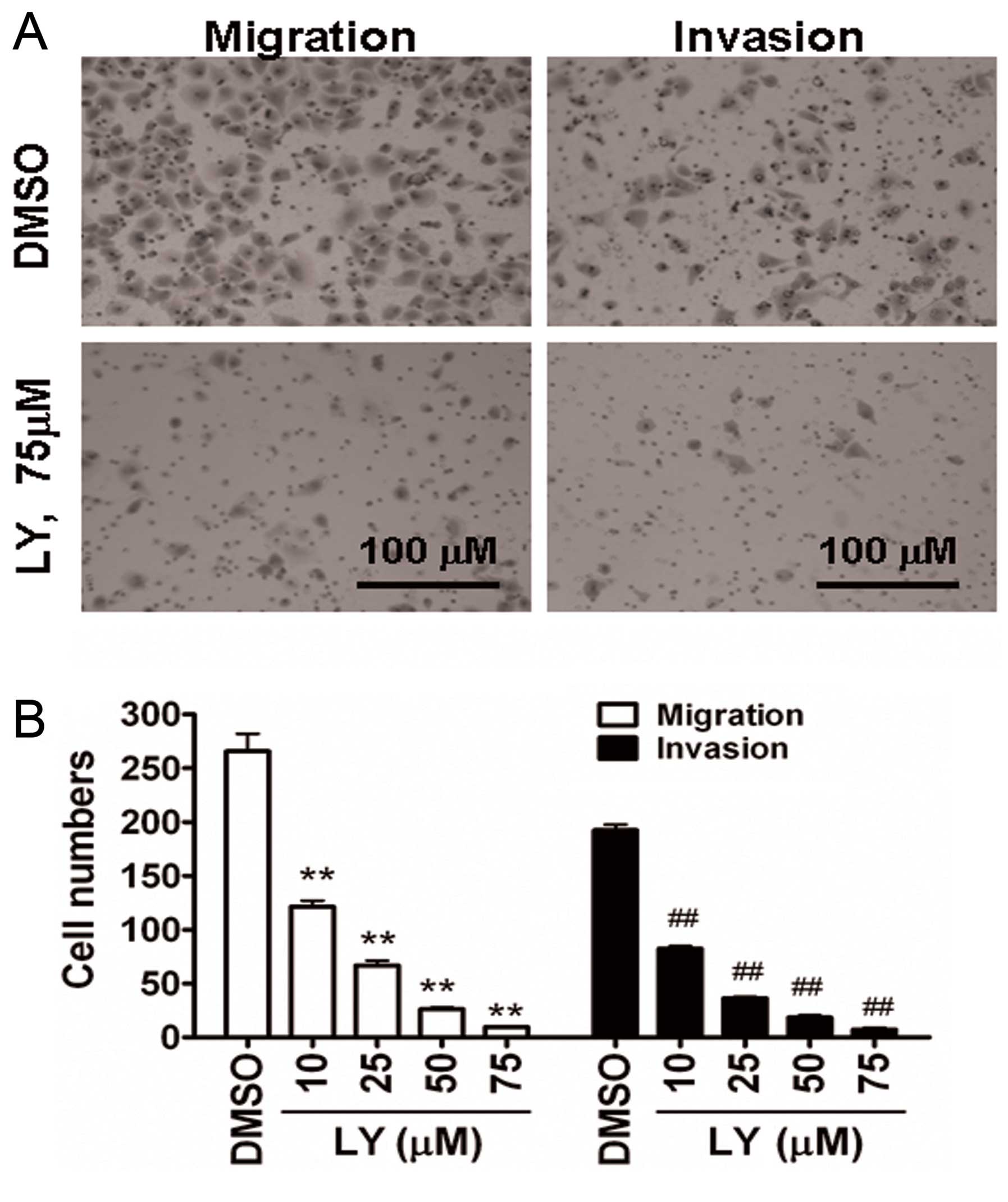
Inhibition of PI3K/Akt signaling
attenuates invasion and migration in CNE2Z cells in vitro
The ability of cellular migration and invasion was
used to assess functional changes in vitro following
inhibition of PI3K/Akt signaling. As we expected, LY294002
treatment of CNE2Z cells indeed significantly suppressed cell
migration and invasion in a concentration-dependent manner
(Fig. 3).
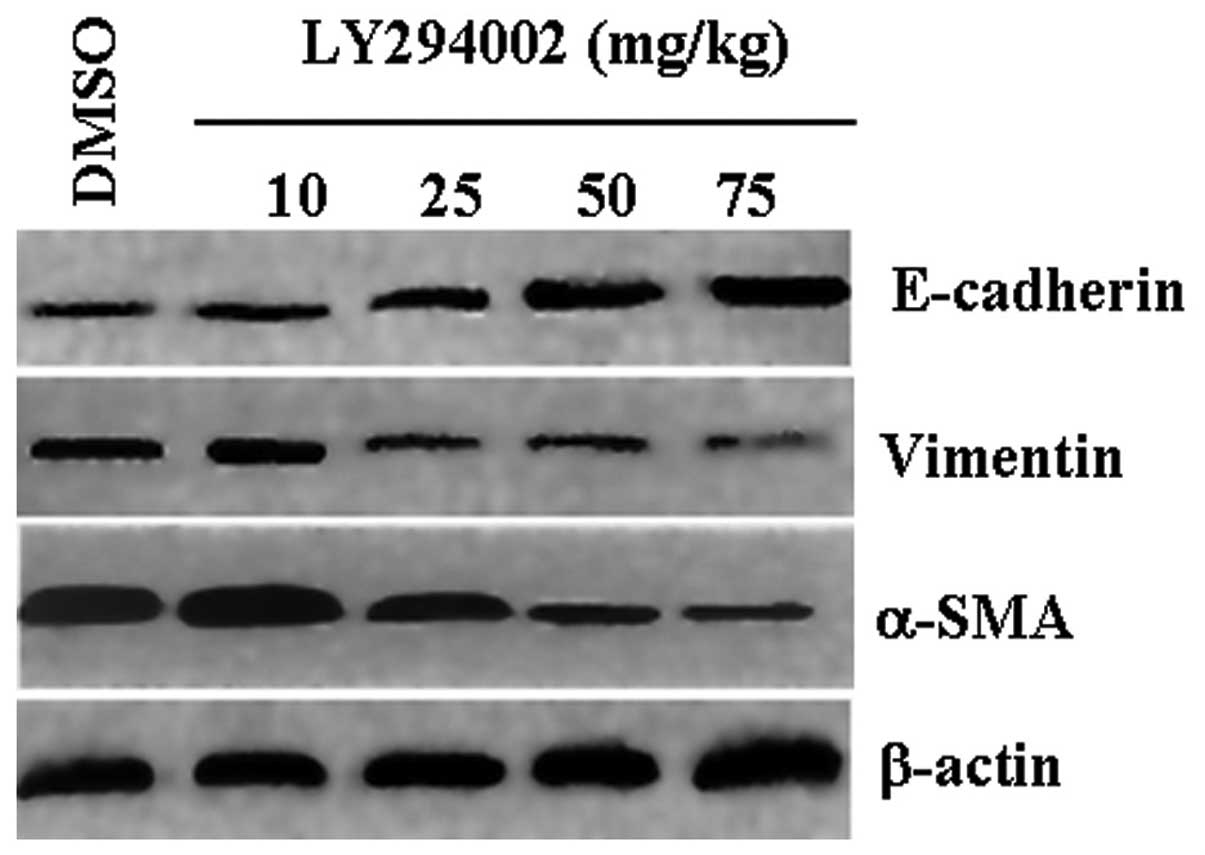
Effects of LY294002 on the expression of
EMT marker genes in vivo
Western blot results revealed that xenografts from
mice administered with DMSO displayed low levels of E-cadherin
expression and high levels of vimentin and α-SMA expression. Mice
treated with LY294002 had increased levels of E-cadherin, and
decreased levels of vimentin and α-SMA that were dependent on the
concentration of LY294002 (Fig. 4).
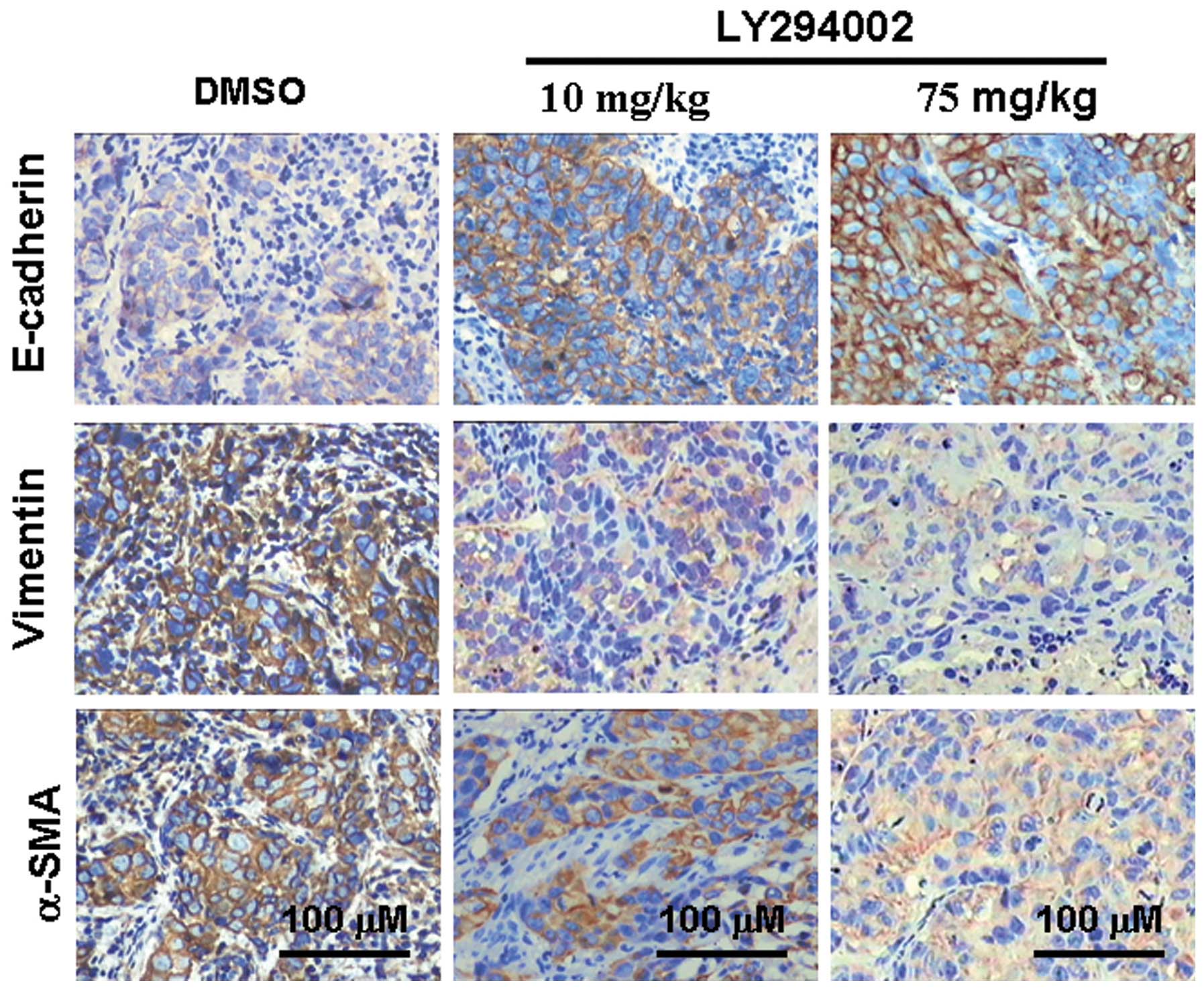
Consistent with the western blot results, the IHC results exhibited
the same tendencies (Fig. 5), and
these results were consistent with those found in vitro
(Fig. 2).
Effects of LY294002 on NPC cells in
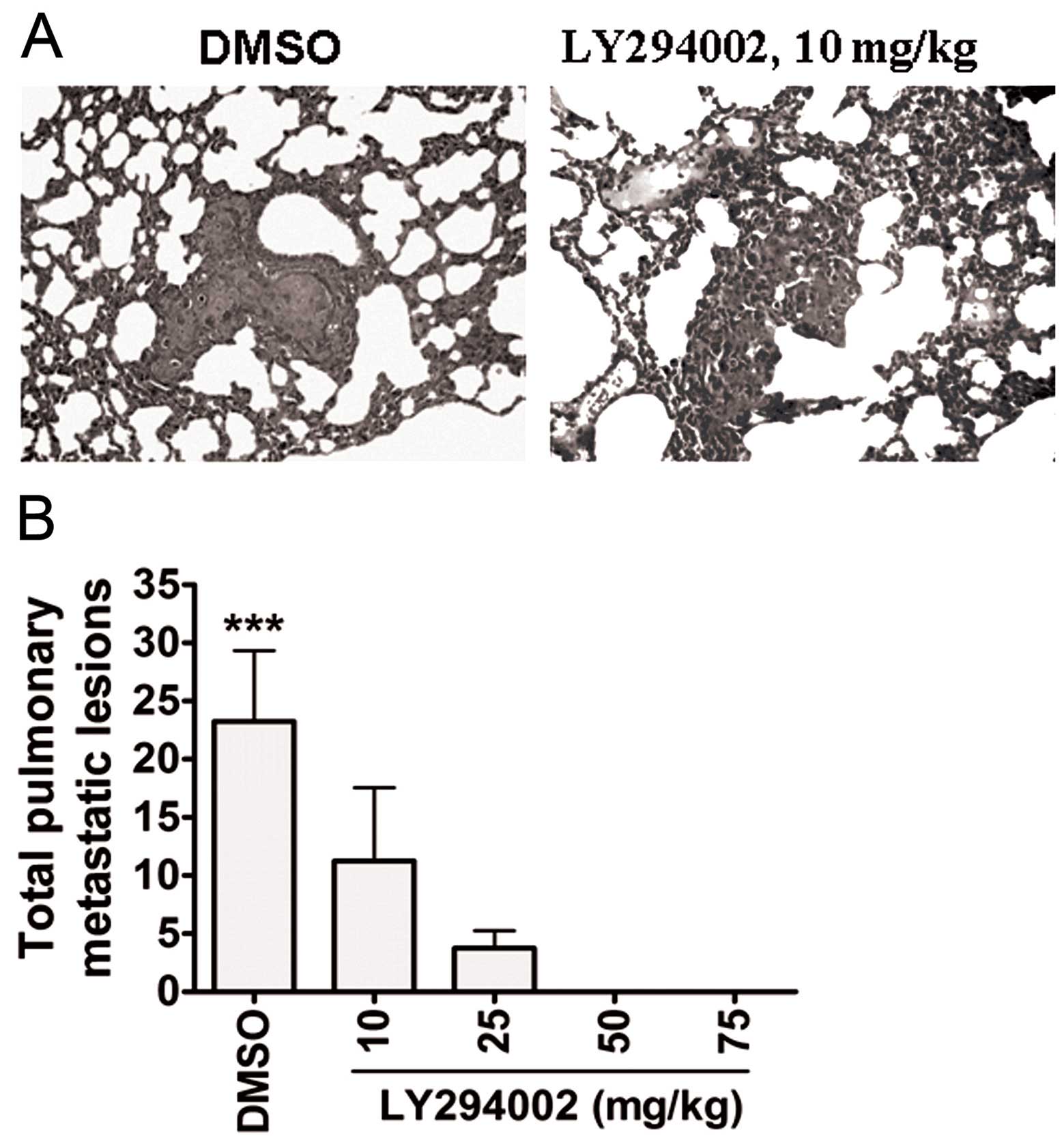
pulmonary metastasis in vivo
After all mice were euthanized, metastatic lesions
in organs such as the liver, spleen, lung and brain were examined.
At the gross level, metastatic lesions were not found in the liver,
spleen, lung and brain of any mice. At the microscopic level,
pulmonary metastatic lesions were observed in each group, with the
following mean total number of pulmonary metastatic lesions: 23.25,
11.25, 3.75, 0 and 0, respectively (P<0.001, Fig. 6).
Discussion
The PI3K/Akt cascade is known to be an important
signal transduction pathway involved in cell survival and
apoptosis. This pathway is activated by other factors that play a
critical role in regulating tumor cell growth, survival,
differentiation, motility and invasion (14). We have previously shown that
inhibition of PI3K/Akt signaling using LY294002 inhibits CNE2Z cell
proliferation, and induces apoptosis in vitro and in
vivo (20). As a hallmark of
malignant cancers, invasion and metastasis are major features that
contribute to cancerous progression (21). Previous findings have indicated that
metastasis of many epithelium-derived tumors have a close
correlation with EMT. During EMT, epithelial cell-derived tumor
cells acquire fibroblast-like properties. Loss of the E-cadherin
protein is a key molecular event during EMT (10,22).
In the present study, we examined the expression of
p-Akt (Ser473) and EMT markers in chronic nasopharyngitis and NPC
tissues. Our findings indicated that higher levels of p-Akt
(Ser473), vimentin and α-SMA expression, and lower membrane
E-cadherin expression levels were observed in clinical NPC samples,
indicating that activation of PI3K/Akt signaling and the occurrence
of EMT was associated with NPC pathogenesis. We then assessed the
relationship between changes in p-Akt (Ser473) and EMT marker
expression levels to NPC metastasis. Consistent with our
expectations, activation of PI3K/Akt signaling and occurrence of
EMT were associated with NPC metastasis. Results from the 23 cases
of NPC patients matched with primary and secondary lesions further
highlighted this relationship. We observed expression of E-cadherin
in the membranes of benign nasopharyngeal epithelia, while
decreased levels of membrane expression and increased cytoplasmic
E-cadherin expression were apparent in NPC tumor cells. Vimentin
and α-SMA expression was lacking in benign nasopharyngeal
epithelial cells, except for some stromal cells. Certain NPC cells,
particularly spindle and fibroblast-like tumor cells, displayed
increased levels of vimentin and α-SMA expression; however, these
were reduced in adjacent epithelial-like cancer cells. Our results
confirmed that EMT was detected in clinical NPC samples. The
occurrence of EMT has a close relationship with metastasis of NPC
cells, as detailed in other reports (23–25).
Treatment of the CEN2Z cell line with LY294002
significantly attenuated the levels of p-Akt(ser473)/Akt in a
concentration-dependent manner, indicating a suppression of
PI3K/Akt signaling in these cells. Increased levels of E-cadherin,
and lower levels of vimentin and α-SMA expression, were observed.
Consistent with our expectations, these molecular changes in
LY294002-treated CNE2Z cells resulted in reduced migration and
invasion of cells. Our findings in vitro showed that
inhibition of PI3K/Akt signaling led to EMrT in NPC cells. Our
results are supported by those from other research groups, which
showed that inhibition of Akt led to upregulation of E-cadherin in
lung (26) and gastric cancer
(27) cells.
The microenvironment of cancer cells is very complex
and flexible. Epithelial cell-derived cancer cells acquire features
of stromal cells and re-express E-cadherin to adapt to their new
environment for certain stimuli. This leads to EMrT and abrogated
cell motility (6,28). To assess the effects of LY294002 on
NPC cell metastasis in vivo, CNE2Z xenograft experiments
were conducted. After tumor-bearing Balb/c null mice were
administered LY294002 at various concentrations, primary tumor
masses were analyzed for activation of Akt and expression of EMT
markers. We observed attenuated Akt activation, which was
accompanied by upregulation of E-cadherin and downregulation of
vimentin and α-SMA. Thus, LY294002 administration in vivo
effectively leads to EMrT. Consistent with changes in EMT markers
in xenografts, significantly reduced pulmonary metastasis was also
observed. Therefore, we concluded that LY294002 administration
in vivo inhibits tumor cell metastasis via EMrT, which was
mediated by the suppression of PI3K/Akt signaling.
It has been reported that EMT of many
epithelium-derived tumors is regulated by multiple signal
transduction pathways that form complex networks (29). Our findings suggest that Akt kinase
was one of the major effectors of EMT signals downstream of PI3K in
NPC cells. Previous studies have demonstrated that EMT generates
cells with the properties of stem cells (30). Recent reports also indicate that EMT
occurs in NPC cells due to the existence of stem cell-like
cancerous cells (31–33). Future studies should focus on
investigating the contribution of Akt mediation to the biology of
cancer stem cell-like cells in NPC.
There were some limitations to the present study;
upstream PI3K/Akt signaling and inhibition of PI3K/Akt signaling
besides using LY294002 in NPC should be considered, and the number
of cases within animal experiments should be greater. Considering
that activation of PI3K/Akt signaling crosstalks with other
cascades (34), the present
investigation outlined the clinical significance of LY294002 in the
treatment of NPC.
In conclusion, we showed that EMT characteristics
were apparent in NPC tissues, and that administration of LY294002
in CNE2Z cells induced upregulation of E-cadherin and
downregulation of α-SMA and vimentin in vitro and in
vivo. This led to impaired cell metastasis via the
induction of EMrT. Our results highlight the possibility of
targeting the PI3K/Akt pathway as a therapeutic strategy for
controlling NPC cell invasion and metastasis. We consider that this
concept for NPC may then be applied to other carcinomas.
Acknowledgements
This study was supported by grants from the Research
Program of Guangdong Medical College (B2011018, B2010013 and
Z2013004), and the Guangdong Provincial Medical Research Foundation
(A2013421).
References
|
1
|
Cao SM, Simons MJ and Qian CN: The
prevalence and prevention of nasopharyngeal carcinoma in China.
Chin J Cancer. 30:114–119. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chang ET and Adami HO: The enigmatic
epidemiology of nasopharyngeal carcinoma. Cancer Epidemiol
Biomarkers Prev. 15:1765–1777. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang Y, Lin ZA, Pan JJ, et al: Concurrent
control study of different radiotherapy for primary nasopharyngeal
carcinoma: intensity-modulated radiotherapy versus conventional
radiotherapy. Ai Zheng. 28:1143–1148. 2009.(In Chinese).
|
|
4
|
Wu Y and Zhou BP: New insights of
epithelial-mesenchymal transition in cancer metastasis. Acta
Biochim Biophys Sin. 40:643–650. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lee JM, Dedhar S, Kalluri R and Thompson
EW: The epithelial-mesenchymal transition: new insights in
signaling, development, and disease. J Cell Biol. 172:973–981.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gao D, Vahdat LT, Wong S, Chang JC and
Mittal V: Microenvironmental regulation of epithelial-mesenchymal
transitions in cancer. Cancer Res. 72:4883–4889. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Talbot LJ, Bhattacharya SD and Kuo PC:
Epithelial-mesenchymal transition, the tumor microenvironment, and
metastatic behavior of epithelial malignancies. Int J Biochem Mol
Biol. 3:117–136. 2012.PubMed/NCBI
|
|
8
|
Thiery JP: Epithelial-mesenchymal
transitions in tumour progression. Nat Rev Cancer. 2:442–454. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huber MA, Kraut N and Beug H: Molecular
requirements for epithelial-mesenchymal transition during tumor
progression. Curr Opin Cell Biol. 17:548–558. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wells A, Yates C and Shepard CR:
E-cadherin as an indicator of mesenchymal to epithelial reverting
transitions during the metastatic seeding of disseminated
carcinomas. Clin Exp Metastasis. 25:621–628. 2008. View Article : Google Scholar
|
|
11
|
Chao YL, Shepard CR and Wells A: Breast
carcinoma cells re-express E-cadherin during mesenchymal to
epithelial reverting transition. Mol Cancer. 9:1792010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zheng H and Kang Y: Multilayer control of
the EMT master regulators. Oncogene. 33:1755–1763. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lee YJ and Han HJ: Troglitazone
ameliorates high glucose-induced EMT and dysfunction of SGLTs
through PI3K/Akt, GSK-3β, Snail1, and β-catenin in renal proximal
tubule cells. Am J Physiol Renal Physiol. 298:F1263–F1275.
2010.PubMed/NCBI
|
|
14
|
Nicholson KM and Anderson NG: The protein
kinase B/Akt signalling pathway in human malignancy. Cell Signal.
14:381–395. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sheng S, Qiao M and Pardee AB: Metastasis
and AKT activation. J Cell Physiol. 218:451–454. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Grille SJ, Bellacosa A, Upson J, et al:
The protein kinase Akt induces epithelial mesenchymal transition
and promotes enhanced motility and invasiveness of squamous cell
carcinoma lines. Cancer Res. 63:2172–2178. 2003.PubMed/NCBI
|
|
17
|
Shen Z, Zeng Y, Guo J, et al:
Over-expression of the special AT rich sequence binding protein 1
(SATB1) promotes the progression of nasopharyngeal carcinoma:
association with EBV LMP-1 expression. J Transl Med. 11:2172013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shen Z, Jiang X, Zeng C, et al: High
expression of ubiquitin-conjugating enzyme 2C (UBE2C) correlates
with nasopharyngeal carcinoma progression. BMC Cancer. 13:1922013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jie W, He QY, Luo BT, et al: Inhibition of
Pim-1 attenuates the proliferation and migration in nasopharyngeal
carcinoma cells. Asian Pac J Trop Med. 5:645–650. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jiang H, Fan D, Zhou G, Li X and Deng H:
Phosphatidylinositol 3-kinase inhibitor (LY294002) induces
apoptosis of human nasopharyngeal carcinoma in vitro and in vivo. J
Exp Clin Cancer Res. 29:342010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: the next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hong KO, Kim JH, Hong JS, et al:
Inhibition of Akt activity induces the mesenchymal-to-epithelial
reverting transition with restoring E-cadherin expression in KB and
KOSCC-25B oral squamous cell carcinoma cells. J Exp Clin Cancer
Res. 28:282009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Luo WR, Chen XY, Li SY, Wu AB and Yao KT:
Neoplastic spindle cells in nasopharyngeal carcinoma show features
of epithelial-mesenchymal transition. Histopathology. 61:113–122.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li XJ, Peng LX, Shao JY, et al: As an
independent unfavorable prognostic factor, IL-8 promotes metastasis
of nasopharyngeal carcinoma through induction of
epithelial-mesenchymal transition and activation of AKT signaling.
Carcinogenesis. 33:1302–1309. 2012. View Article : Google Scholar
|
|
25
|
Horikawa T, Yang J, Kondo S, et al: Twist
and epithelial-mesenchymal transition are induced by the EBV
oncoprotein latent membrane protein 1 and are associated with
metastatic nasopharyngeal carcinoma. Cancer Res. 67:1970–1978.
2007. View Article : Google Scholar
|
|
26
|
Chen XF, Zhang HJ, Wang HB, et al:
Transforming growth factor-β1 induces epithelial-to-mesenchymal
transition in human lung cancer cells via PI3K/Akt and MEK/Erk1/2
signaling pathways. Mol Biol Rep. 39:3549–3556. 2012.
|
|
27
|
Kang MH, Kim JS, Seo JE, Oh SC and Yoo YA:
BMP2 accelerates the motility and invasiveness of gastric cancer
cells via activation of the phosphatidylinositol 3-kinase
(PI3K)/Akt pathway. Exp Cell Res. 316:24–37. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ding S, Zhang W, Xu Z, et al: Induction of
an EMT-like transformation and MET in vitro. J Transl Med.
11:1642013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Thiery JP and Sleeman JP: Complex networks
orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell
Biol. 7:131–142. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Battula VL, Evans KW, Hollier BG, et al:
Epithelial-mesenchymal transition-derived cells exhibit
multilineage differentiation potential similar to mesenchymal stem
cells. Stem Cells. 28:1435–1445. 2010. View
Article : Google Scholar
|
|
31
|
Kong QL, Hu LJ, Cao JY, et al:
Epstein-Barr virus-encoded LMP2A induces an epithelial-mesenchymal
transition and increases the number of side population stem-like
cancer cells in nasopharyngeal carcinoma. PLoS Pathog.
6:e10009402010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Guo D, Xu BL, Zhang XH and Dong MM: Cancer
stem-like side population cells in the human nasopharyngeal
carcinoma cell line CNE-2 possess epithelial mesenchymal transition
properties in association with metastasis. Oncol Rep. 28:241–247.
2012.
|
|
33
|
Lin CH, Shen YA, Hung PH, Yu YB and Chen
YJ: Epigallocathechin gallate, polyphenol present in green tea,
inhibits stem-like characteristics and epithelial-mesenchymal
transition in nasopharyngeal cancer cell lines. BMC Complement
Altern Med. 12:2012012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jazirehi AR, Wenn PB and Damavand M:
Therapeutic implications of targeting the PI3Kinase/AKT/mTOR
signaling module in melanoma therapy. Am J Cancer Res. 2:178–191.
2012.PubMed/NCBI
|