Introduction
Neuroblastoma is the most common extracranial solid
tumor of children and accounts for 15% of pediatric cancer deaths
(1,2). Neuroblastoma may regress spontaneously
in children aged <18 months; however, neuroblastoma in children
>18 months is always associated with aggressive characteristics
and is considered as a high-risk disease. Even with intensive
combination treatment with surgery, chemotherapy and radiation, the
long term survival rate for patients with neuroblastoma remains
<40%, highlighting the necessity to explore novel therapies for
this devastating childhood disease (1–3).
Apoptosis is the process of programmed cell death
that plays a key role in maintenance of cell homeostasis and
differentiation. Resistance of neuroblastoma cells to apoptosis is
a major contributing factor to the aggressive nature of the
high-risk neuroblastoma (2–4). Among many mechanisms that cause
resistance of neuroblastoma cells to apoptosis, elevated expression
of antiapoptotic members of Bcl-2 family proteins such as Bcl-2,
Bcl-xL, and Mcl-1 has attracted particular attention, since these
proteins negatively regulate mitochondria-mediated apoptosis
(5). It was reported that >50%
of neuroblastoma tumor tissues and the majority of neuroblastoma
cell lines express a high level of Bcl-2 (4,6).
Moreover, overexpression of Bcl-2 protein was found to be closely
associated with poor response to apoptosis induction by
conventional therapies such as radiotherapy and chemotherapy
(7–9). Hence, restoration of apoptosis by
targeting Bcl-2 might be a promising strategy in managing
neuroblastoma.
It has been proposed that small-molecule Bcl-2
inhibitors ABT-737 and ABT-263 treat human cancer (10,11).
Preclinical studies showed that ABT-737 and ABT-263 are able to
potently induce apoptosis and exhibit strong anticancer activity in
certain types of cancer through inhibition of Bcl-2 and Bcl-xL
(10,11). In particular, clinical trials with
ABT-263 have shown promising anticancer activity in hematologic
tumors and lung cancer, demonstrating ABT-263 is a promising
anticancer drug (12,13). Nevertheless, a fundamental problem
of ABT-737 and ABT-263 is that the majority of solid tumors,
including neuroblastoma, are resistant to the apoptosis induction
by these two new drugs, posing a chief problem for the potential
utility in management of these cancers (14). Thus, major efforts have focused on
identifying approaches that could overcome
ABT-737/ABT-263-resistance through combinational strategies
(15–17).
Norcantharidin (NCTD) is a small-molecule anticancer
drug derived from a traditional Chinese medicine blister beetle
(18–20). We recently reported that NCTD could
overcome the resistance to ABT-737 in a panel of hepatocellular
cancer cell lines (21). In the
current study, we investigated the anticancer activity of ABT-263
in combination with NCTD in neuroblastoma SH-SY5Y and CHLA-119 cell
lines. We found that NCTD markedly increased the expression of
Noxa, an endogenous Mcl-1 inhibitor, which leads to significant
enhancement of ABT-263-mediated anticancer activity in these
neuroblastoma cells. Since NCTD is routinely used to treat patients
with cancer in clinic, our findings have translational significance
in managing neuroblastoma.
Materials and methods
Cell lines and compound preparations
Neuroblastoma cell lines SH-SY5Y and CHLA-119
purchased from the Cell Bank of the Chinese Academy of Sciences
Shanghai Institute of Cell Biology (Shanghai, China) were
maintained in a 1:1 mixture of F12 and Dulbecco’s modified Eagle’s
medium (DMEM) (HyClone/Thermo Fisher Scientific, Beijing, China)
supplemented with 10% heat-inactivated fetal FBS (Hangzhou Sijiqing
Biological Engineering Materials Co., Ltd, Hangzhou, China) in a
humidified atmosphere of 5% of CO2 in air at 37°C.
ABT-263 was purchased from Biochempartner (Shanghai, China) and was
dissolved in DMSO with a stock concentration of 100 mM and stored
at −20°C. NCTD was purchased from Shaanxi Huike Plants Exploitation
(Xi’an, China), and the purity was >98% as determined by
high-performance liquid, and was dissolved in DMEM with a stock
concentration of 1 mM and stored at −20°C.
MTT viability assay
Cell viability was measured by
3-[4,5-dimethylthiazol-2-thiazolyl]-2,5-diphenyl-tetrazolium
bromide (MTT) assay. The percentages of absorbance relative to
those of untreated control samples were plotted as a linear
function of drug concentration. Inhibition of cell viability was
measured by percentage of viable cells relative to the control: %
Inhibition = 100% × ODT/ODC, where ODT is the average OD value of
the treated samples and ODC is the average OD value of the control
samples.
Cell death and flow cytometry apoptosis
assays
Cell death was quantitated by microscopic
examination in trypan blue exclusion assays. Blue-stained and
considerably shrunken cells were counted as non-viable cells. SubG1
apoptosis in cells was examined by propidium iodide (PI; 50 μg/ml
in PBS) staining in the presence of RNase A (100 μg/ml) and flow
cytometry with a BD LSR II system (BD Biosciences, Shanghai,
China).
Apoptosis TUNEL assay
Apoptosis terminal deoxynucleotidyl transferase dUTP
nick end labeling (TUNEL) assay was performed with a commercial kit
of DeadEndTM Colorimetric TUNEL assay from Promega
(Shanghai, China).
Clonogenic assay
For clonogenic assay, 1,000 cells were seeded into
6-well dishes in 3 ml of medium. The next day, cells were treated
as indicated, and then maintained for 14 days at 37°C in a 5%
CO2 incubator. Cells were washed with drug-free medium,
stained with 0.05% methylene blue, and cell colonies (>50 cells)
were counted at 14 days. Assays were performed in duplicate with at
least three independent repetitions per treatment.
Western blot analysis and cell
fractionation
Western blot analyses and cell fractionation were
performed as previously described (21). Antibodies used were: anti-cytochrome
c rabbit polyclonal antibody (556432) from BD Biosciences
(Shanghai, China); anti-actin goat polyclonal antibody, and
HRP-conjugated secondary anti-mouse, anti-goat and anti-rabbit
antibodies from Santa Cruz Biotechnology (Shanghai, China);
anti-caspase-9 rabbit polyclonal antibody (9502), anti-caspase-3
rabbit polyclonal antibody (9662), anti-Bcl-2 rabbit polyclonal
antibody (2870), anti-Mcl-1 rabbit polyclonal antibody (4572) and
anti-COX IV from Cell Signaling Technology (Shanghai, China).
Protein expression levels were quantified by densitometry (ImageJ
program). The relative density was calculated as follows: Relative
density = (band density of the sample)/(band density of the
control).
RNA interference
The siRNA transfections were carried out using
Lipofectamine RNAiMax transfection reagent (Invitrogen) according
to the manufacturer’s instructions. Validated siRNA for Mcl-1 and
Noxa duplexes were purchased from Invitrogen (Shanghai, China). A
non-silencing control siRNA (siCTL) was used as the control.
Concentration for transfection was 10 nmol/l of each siRNA.
Statistical analysis
Statistical analyses were performed by one-way ANOVA
using SPSS (version 13.0; SPSS Inc, Chicago, IL, USA). P<0.05
was considered statistically significant, P<0.01 was considered
highly statistically significant.
Results
ABT-263 is ineffective in neuroblastoma
SH-SY5Y and CHLA-119 cell lines
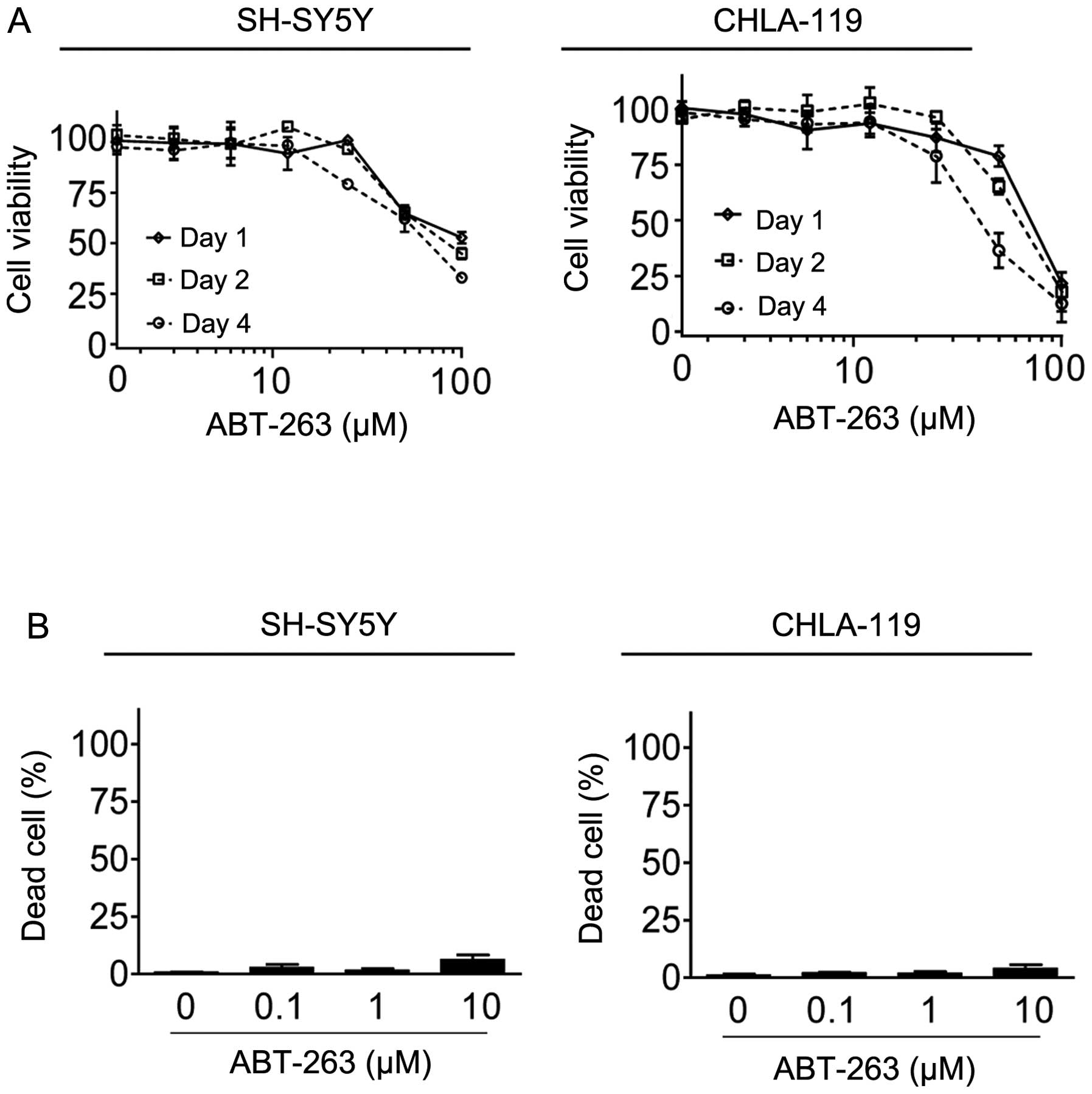
We initiated our study to evaluate the activity of
Bcl-2 inhibitor ABT-263 in SH-SY5Y and CHLA-119 cell lines, since
these two cell lines expressed a high level of Bcl-2 (4,6). MTT
cell proliferation assays showed that treatment with ABT-263 for 24
h achieved IC50 values of 70 and 98 μM, respectively, in
SH-SY5Y and CHLA-119 cell lines. ABT-263 at 10 μM inhibited cell
proliferation only by 7 and 4%, respectively, in the two cell lines
(Fig. 1A). Treatment with ABT-263
for a longer time (48 and 72 h) did not appreciably increase the
inhibitory effect on cell proliferation. Consistently, trypan blue
staining assays showed that treatment with ABT-263 at 10 μM for 48
h only induced approximately 6% and 7% cell death, respectively, in
the two cell lines (Fig. 1B). These
results showed that ABT-263 is ineffective in these neuroblastoma
cell lines.
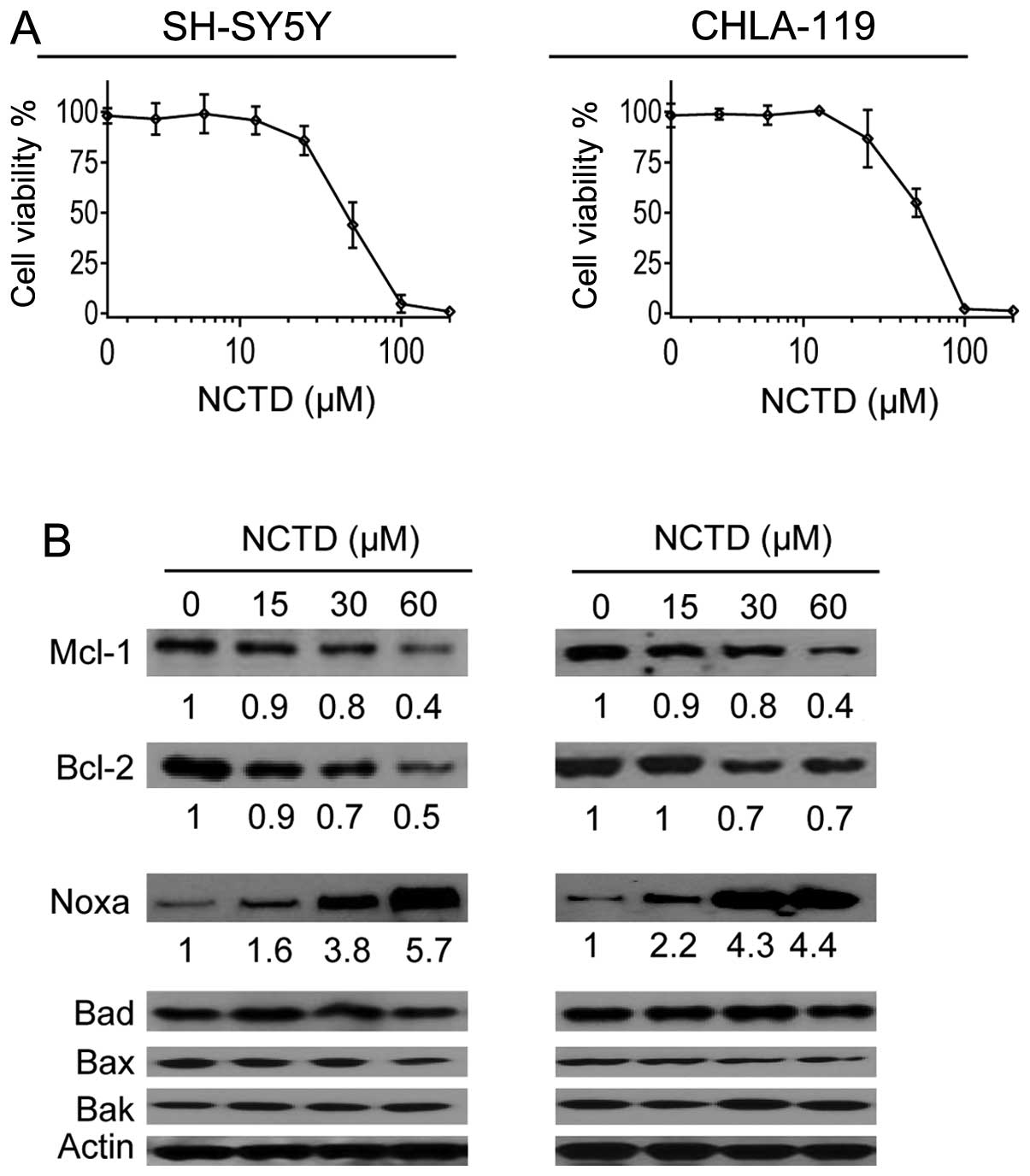
NCTD markedly increases Noxa expression
in neuroblastoma cells
A number of previous studies have reported that NCTD
could modulate the levels of Bcl-2 family proteins in oral cancer,
liver cancer and breast cancer cells (20–22).
We next investigated whether this drug had a similar effect on
neuroblastoma cells. MTT assays were used to determine a subtoxic
concentration range of NCTD in the cell lines. We found that NCTD
at 15–60 μM dose-dependently and partially inhibited cell viability
in both cell lines (Fig. 2A). Thus,
we treated SH-SY5Y and CHLA-119 cell lines with these sub-toxic
concentrations of NCTD for 48 h (Fig.
1B), and then examined the expression of several key members of
Bcl-2 family proteins in cells. Western blot analysis showed that
NCTD at these concentrations did not have an effect on the
expressions of Bad and Bax, Bak, and had a modest inhibitory effect
on the expressions of Mcl-1 and Bcl-2 in the two cell lines
(Fig. 1B). Contrarily, the same
treatments markedly increased the level of Noxa in the two cell
lines in a dose-dependent manner. Densitometry analysis of western
blotting results showed NCTD at 15, 30 and 60 μM increased the
level of Noxa 1.6-, 3.8- and 5.7-fold in the SH-SY5Y cell line
respectively, and increased the level of Noxa 2.2-, 4.3- and
4.4-fold, in the CHLA-119 cell line respectively (Fig. 1B).
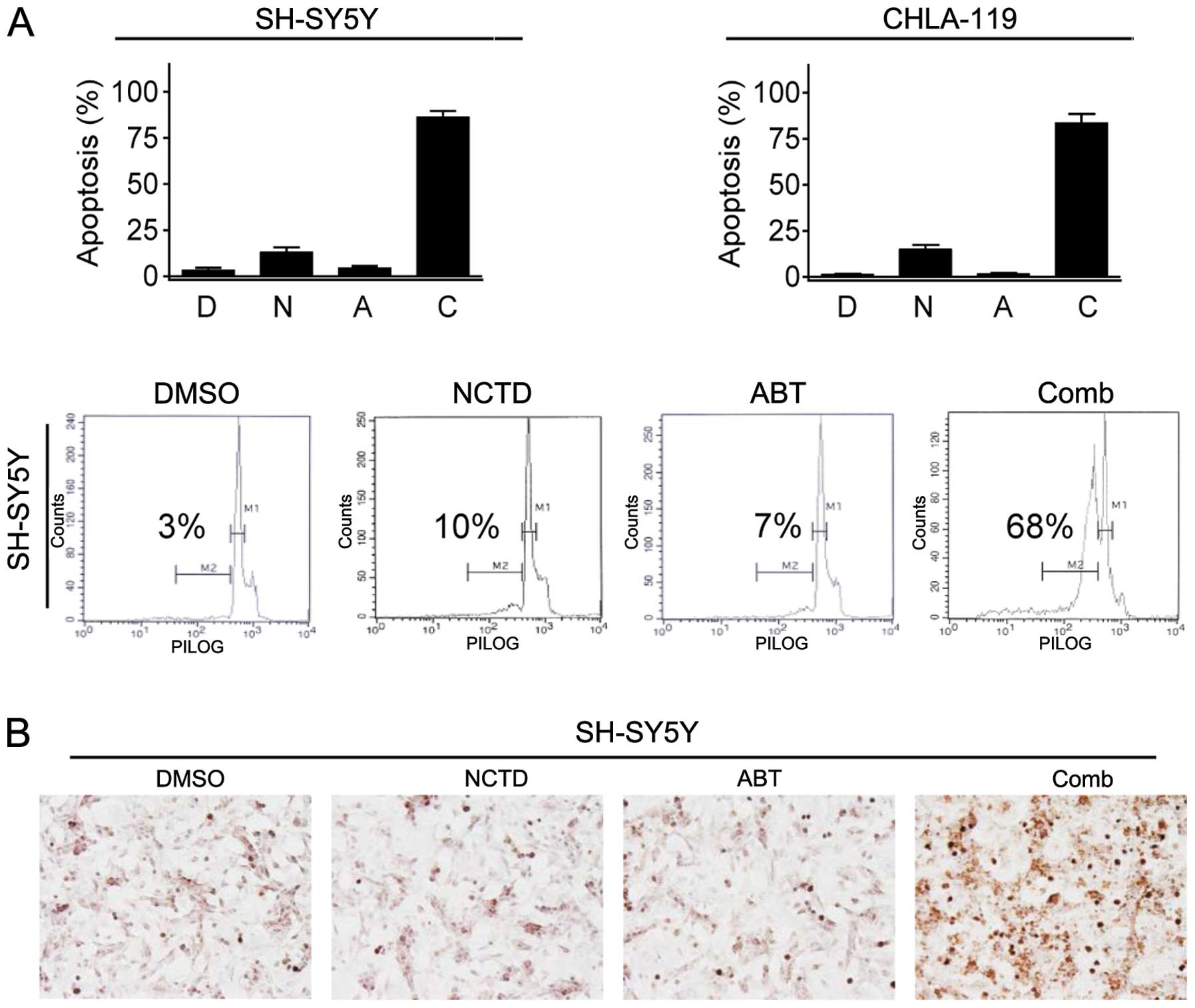
NCTD enhances ABT-263-mediated apoptosis
in neuroblastoma cells
We next investigated whether NCTD enhanced
ABT-263-mediated apoptosis induction in neuroblastoma cells. We
treated neuroblastoma cells with NCTD at 30 μM alone, ABT-263 at 2
μM alone or their combination for 24 h, and performed PI staining
and flow cytometry to quantify the percentage of apoptotic cells in
the “sub-G1” peak. We observed that single-agent treatments induced
apoptosis only in <10% of neuroblastoma cells (Fig. 3A). In contrast, their combination
induced 68% and 65% of cells undergoing apoptosis in SH-SY5Y and
CHLA-119 cell lines, respectively (Fig.
3A). We then performed TUNEL assay in SH-SY5Y cell line to
validate the results of flow cytometry assays. We found that
treatment with either agent alone induced only a very small
fraction of cells positively stained with TUNEL, whereas their
combination not only resulted in TUNEL positive in most cells, but
also led to cells shrinking into a dense, round mass, a distinct
morphological characteristic of apoptosis (Fig. 3B) (14). These results demonstrated that NCTD
potently enhanced ABT-263-triggered apoptosis in neuroblastoma
cells.
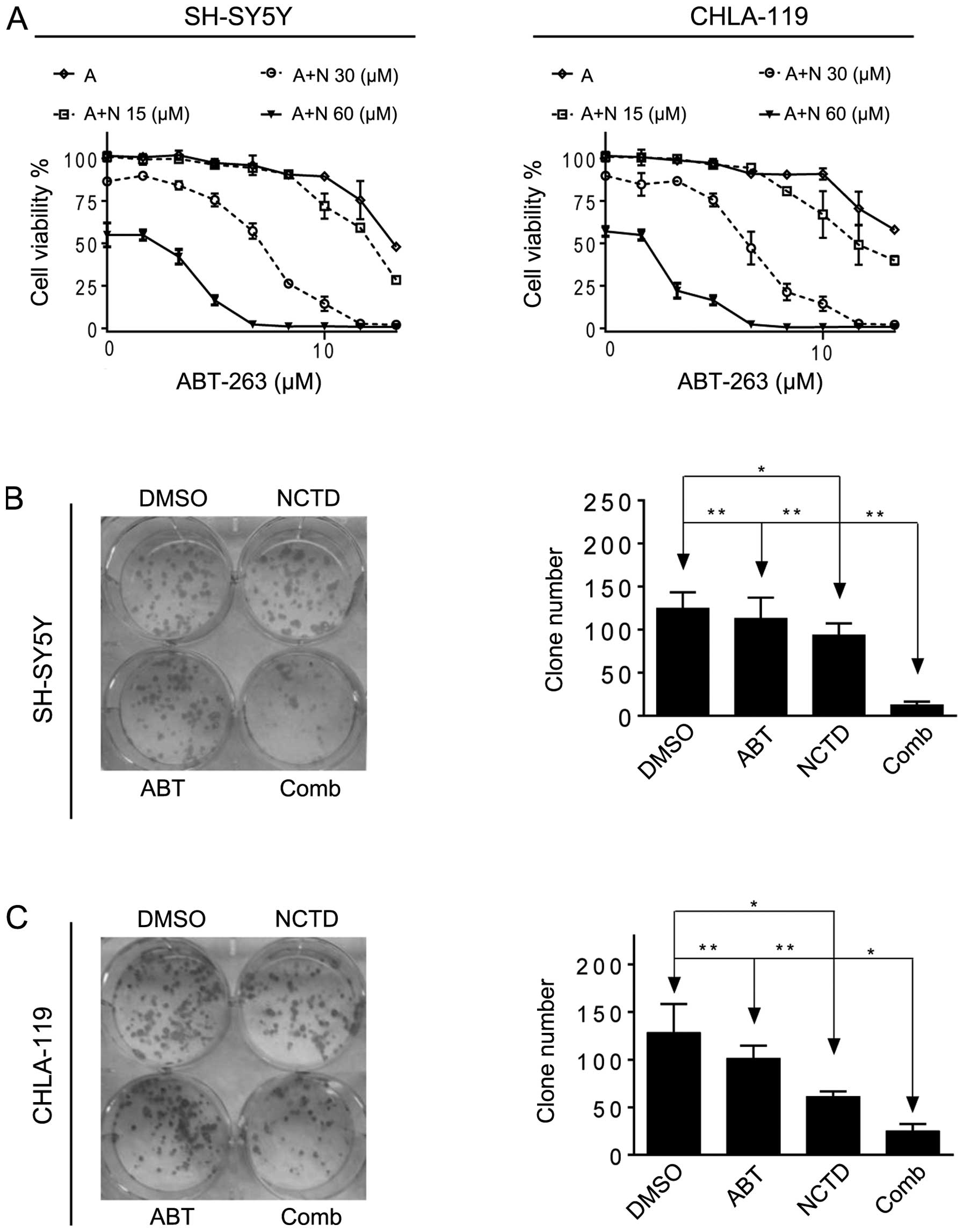
NCTD enhances ABT-263-mediated inhibition
of cell proliferation and clonal formation in neuroblastoma
cells
Subsequently, we investigated whether NCTD could
enhance ABT-263-mediated inhibition of cell proliferation in
neuroblastoma cells. We treated the cells with ABT-263 alone or in
combination with NCTD for 4 days, then examined cell proliferation
with MTT assays. We found that NCTD dose-dependently enhanced
ABT-263-mediated cell proliferation inhibition in the two cell
lines (Fig. 4A). Notably, the
enhancement of proliferation inhibition correlated well with
upregulation of Noxa by NCTD in the two cell lines (Figs. 2 and 4A). For instance, in the SH-SY5Y cell
line, NCTD at 15 μM, a concentration increasing the level of Noxa
1.6-fold only reduced the IC50 value by 45% (from 40 to
22 μM), while NCTD at 30 and 60 μM, concentrations increasing the
level of Noxa 3.8-fold and 5.7-fold, in combination with ABT-263
achieved 100% cell viability inhibition in the two neuroblastoma
cell lines.
Neuroblastoma cells were grown in the presence of
NCTD alone, ABT-263 alone or both for 2 weeks. Clonogenic assays
were performed to determine whether the combination of NCTD and
ABT-263 had long-term stronger anticancer activity in SH-SY5Y and
CHLA-1 cell lines. As shown in Fig.
4, ABT-263 alone showed no efficacy and NCTD at 30 μM showed
modest effect, whereas the combination achieved a significant
improvement in the inhibition of colony formation (combination vs.
ABT-263, P<0.001; combination vs. NCTD, P<0.01).
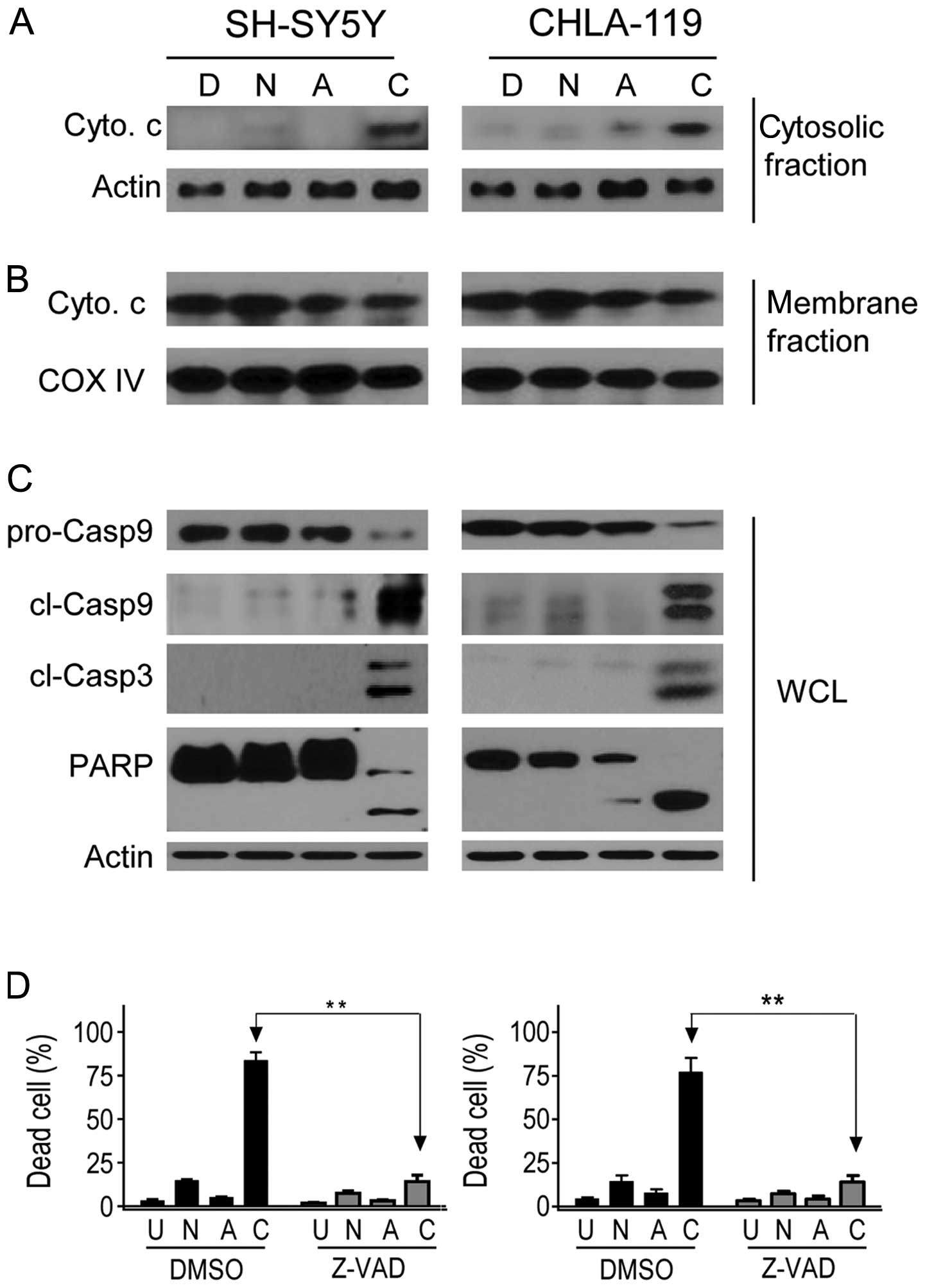
NCTD enhances ABT-263-triggered release
of cytochrome c and activation of caspases in neuroblastoma
cells
Cytochrome c released from the mitochondrial
intermembrane space into the cytosol is a crucial step in
ABT-263-triggered apoptosis (10,11,23).
We next investigated whether treatment with ABT-263 alone, NCTD
alone or both could induce cytosolic release of cytochrome c
in neuroblastoma cells. As shown in Fig. 5A, treatment with either NCTD or
ABT-263 alone induced a negligible release of cytochrome c.
By sharp contrast, co-treatment with the two drugs resulted in a
robust increase of cytochrome c release in the two cell
lines, which was accompanied by a modest decrease of cytochrome
c in mitochondria-enriched membrane fraction (Fig. 5B). Hence, these data indicated that
NCTD markedly enhanced ABT-263-triggered damage of the
mitochondrial outer membrane integrity and allowed the release of
the proapoptotic protein cytochrome c into the cytosol in
the neuroblastoma cells.
Activation of caspases is another key event
following cytochrome c release in apoptosis signaling
(24). We then treated the cells
with single agents or combination, and examined whether caspase-9,
-3 were activated in neuroblastoma cells with western blot
analysis. We found that as compared to the minimal effect by
single-agent treatments, combination treatment with ABT-263 and
NCTD for 24 h led to a robust accumulation of cleaved fragments of
initiator caspase-9 in the two cell lines. Similarly, substantial
accumulation of cleaved effector caspase-3 was observed in the
combination treatment but not in single-agent treatments. We also
observed that PARP, a substrate of caspase-3, was cleaved upon
treatment with the combination, but not by treatment with single
agents (Fig. 5C).
To determine whether caspases were required for the
combination effect of NCTD and ABT-263, SH-SY5Y and CHLA-119 cell
lines were pretreated with the 50 μM of pancaspase inhibitor
(zVAD.fmk) for 1 h before the addition of NCTD (30 μM) and ABT-263
(2 μM) (Fig. 5D). Inhibition of
caspase activity significantly attenuated the cell death induction
by the combination, evidently showing that a caspase-dependent
mechanism was involved in the anticancer activity of the two
drugs.
Collectively, these results demonstrate that NCTD
greatly enhances ABT-263-triggered apoptosis in neuroblastoma
cells.
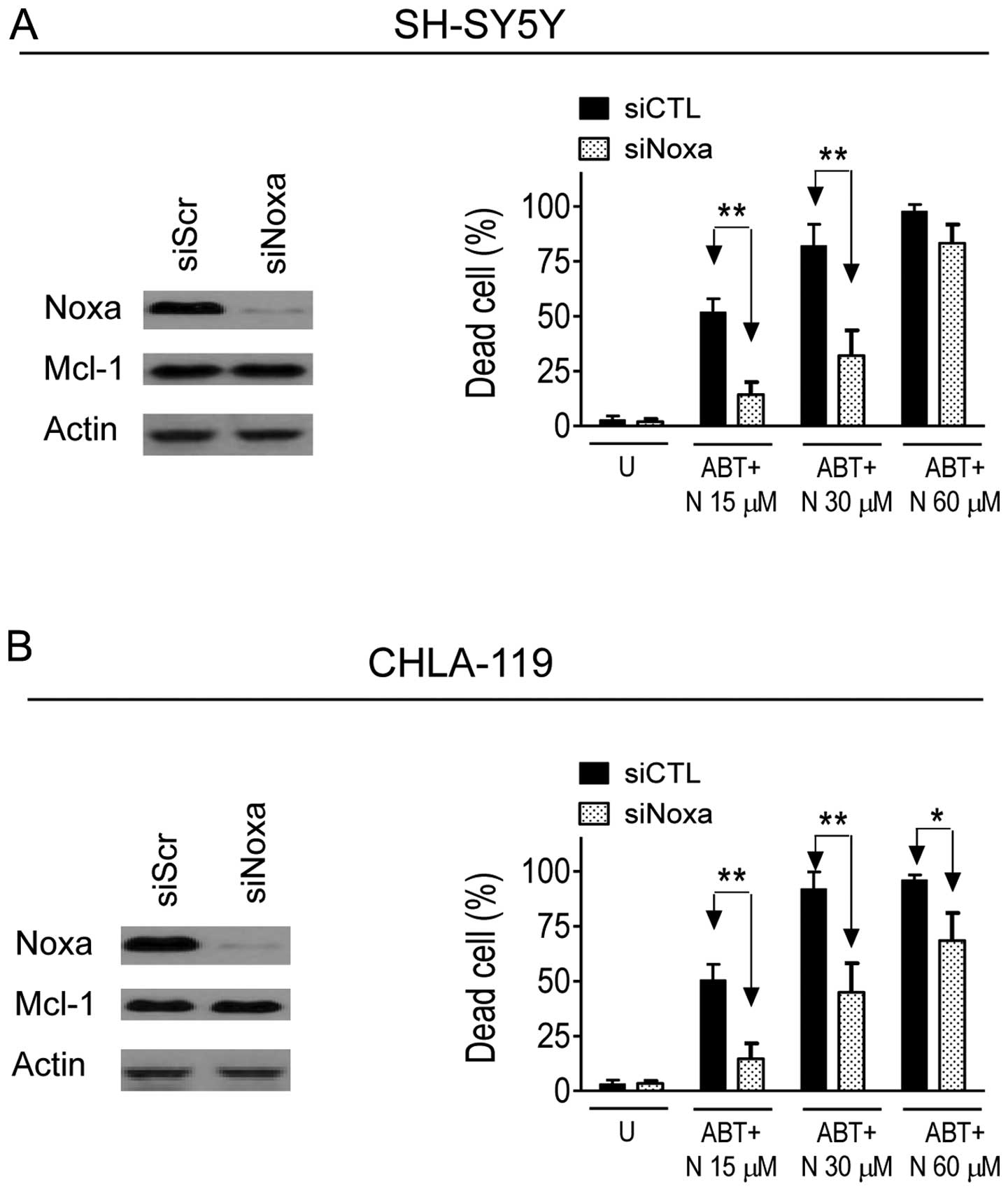
Noxa plays an essential role in the
enhancement of ABT-263-mediated cell death by NCTD in neuroblastoma
cells
Mcl-1 has been identified as the key mediator of
ABT-263-resistance in cancer cells (10,11,14,15).
Given the fact that Noxa is an endogenous inhibitor of Mcl-1
(25–27), we hypothesized that upregulation of
Noxa might play a role in the enhancement of ABT-263-mediated
anticancer activity by NCTD. To investigate this hypothesis, we
knocked down Noxa with specific siRNAs in the two neuroblastoma
cell lines. As shown in Fig. 6,
transfection for 48 h effectively suppressed Noxa expression in
SH-SY5Y and CHLA-119 cell lines, but had no effect on the
expression of Mcl-1 in the two cell lines. MTT assays showed that
inhibition of Noxa expression by siRNA significantly attenuated
cell viability inhibition mediated by ABT-263 in combination with
NCTD. For instance, ABT-263 in combination with NCTD at 30 μM
induced 82% cell death in siCTL transfected SH-SY5Y cells, while
the same treatments only caused 32% cell death in siNoxa
transfected cells (P<0.01) (Fig.
6). An inhibitory effect on the combination by siNoxa was also
observed in the CHLA-119 cell line. These results indicate that
upregulation of Noxa is required in the enhancement of
ABT-263-mediated anticancer activity by NCTD in neuroblastoma
cells.
Discussion
ABT-737 and ABT-263 are small molecule Bcl-2
inhibitors developed as novel anticancer drugs. Preclinical studies
demonstrated that these two drugs exhibit promising anticancer
efficacy in certain types of cancers. Therefore, ABT-737 and
ABT-263 represent promising anticancer therapies. Nonetheless, due
to low binding affinity to another member of antiapoptotic Bcl-2
protein, Mcl-1, ABT-737 and ABT-263 have very weak anticancer
activity in a large number of other cancer types, such as
neuroblastoma, which express an elevated level of Mcl-1. The
ineffectiveness of ABT-737 and ABT-263 in those cancers poses a key
problem for their potential application in a wide range of cancer
types (14–16). Accordingly, finding approaches to
overcome ABT-263/ABT-737-resistance through inhibition of Mcl-1,
directly or indirectly, is urgently needed.
The strategies that directly target Mcl-1 expression
have been investigated extensively. For instance, cyclin-dependent
kinase (CDK) inhibitors, such as flavopiridol and roscovitine, have
been found to enhance ABT-737 activity by suppressing Mcl-1
expression in several types of cancer (15). Meanwhile, an alternative strategy
that sensitizes cancer cells to ABT-263/ABT-737-mediated anticancer
activity is the use of agents capable of increasing Noxa
expression. Noxa belongs to the proapoptotic Bcl-2 BH3 subfamily,
and is known as an endogenous Mcl-1 inhibitor (25–27).
Increasing Noxa expression has been reported to enhance
ABT-263-activity in a number of cancer types. For instance, Simonin
et al found that platinum compounds enhance ABT-737-mediated
apoptosis in ovarian carcinoma cells by upregulation of Noxa
expression (27). Zinn et al
reported that chloroquine overcome ABT-737-resistance in small lung
cancer cells by increasing Noxa expression (28). In the current study, we observed
that NCTD considerably enhanced ABT-263-mediated anticancer
activity, which was accompanied by marked increase of Noxa
expression in neuroblastoma cells. By employing specific siRNA, we
confirmed that Noxa played an essential role in the enhancement of
ABT-263-mediated anticancer activity by NCTD. Since NCTD is a
conventional anticancer drug, and ABT-263 is being tested in
clinic, our study has more translational significance in ABT-based
cancer therapy.
NCTD is a demethylated analog of cantharidin, the
major bioactive constituent of Chinese blister beetle
Mylabris, which has been used in China to treat tumors,
inflammation and many other conditions for a long time. Previous
studies showed that NCTD could modulate the expression of Bcl-2 and
Mcl-1 in oral, breast and hepatocellular cancer cells (19–22,29).
Here, we explored whether this drug also inhibited antiapoptotic
Bcl-2 proteins in neuroblastoma cells. However, we observed that
NCTD at the applied concentrations only had a modest effect on
downregulation of Mcl-1 and Bcl-2. By contrast, NCTD could markedly
increase the expression of Noxa in the two neuroblastoma cell
lines. These discrepancies between this study and previous reports
may reflect the diverse response of different cancer types to NCTD,
and may also indicate the need for elucidation of the true
mechanism by which NCTD elicits anticancer activity in the
future.
Several pieces of evidence indicated that
combination treatment by two drugs triggered activation of the
mitochondrial apoptosis signaling pathway. Firstly, flow cytometery
and TUNEL staining assays showed that co-treatment with ABT-263 and
NCTD induced apoptosis in a large fraction of cells. Secondly,
western blot analysis showed that combination treatment induced
activation of several typical biomarkers of mitochondria-mediated
apoptosis, including cytosolic cytochrome c release,
activation of initiator caspase-9 and effector caspase-3 and PARP
cleavage. Thirdly, cell killing induced by the combination could be
largely rescued by a pancaspase inhibitor zVAD.fmk. Once again, our
findings support that the combinational effect of ABT-737/ABT-263
with other approaches is dependent on ABT-737/ABT-263-triggered
mitochondrial apoptosis signaling (15–17,21).
Acknowledgements
This research was supported by the National Natural
Science Foundation of China (81101685).
References
|
1
|
Meadows A, Baum E, Fossati-Bellani F, et
al: Second malignant neoplasms in children: an update from the late
effects study group. J Clin Oncol. 3:532–538. 1985.PubMed/NCBI
|
|
2
|
Welch C, Chen Y and Stallings R:
MicroRNA-34a functions as a potential tumor suppressor by inducing
apoptosis in neuroblastoma cells. Oncogene. 26:5017–5022. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
George R, Sanda T, Hanna M, et al:
Activating mutations in ALK provide a therapeutic target in
neuroblastoma. Nature. 455:975–978. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Goldsmith K, Lestini B, Gross M, et al:
BH3 response profiles from neuroblastoma mitochondria predict
activity of small molecule Bcl-2 family antagonists. Cell Death
Differ. 17:872–882. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kroemer G: The proto-oncogene Bcl-2 and
its role in regulating apoptosis. Nature Med. 3:614–620. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fang H, Harned T, Kalous O, Maldonado V,
DeClerck Y and Reynolds C: Synergistic activity of fenretinide and
the Bcl-2 family protein inhibitor ABT-737 against human
neuroblastoma. Clin Cancer Res. 17:7093–7104. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Castle V, Heidelberger K, Bromberg J, Ou
X, Dole M and Nuñez G: Expression of the apoptosis-suppressing
protein bcl-2, in neuroblastoma is associated with unfavorable
histology and N-myc amplification. Am J Pathol. 143:1543–1550.
1993.PubMed/NCBI
|
|
8
|
Dole M, Nuñez G, Merchant A, Maybaum J,
Rode C, Bloch C and Castle V: Bcl-2 inhibits chemotherapy-induced
apoptosis in neuroblastoma. Cancer Res. 54:3253–3259.
1994.PubMed/NCBI
|
|
9
|
Hanada M, Krajewski S, Tanaka S, et al:
Regulation of Bcl-2 oncoprotein levels with differentiation of
human neuroblastoma cells. Cancer Res. 53:4978–4986.
1993.PubMed/NCBI
|
|
10
|
Tse C, Shoemaker A, Adickes J, et al:
ABT-263: a potent and orally bioavailable Bcl-2 family inhibitor.
Cancer Res. 68:3421–3428. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Oltersdorf T, Elmore S, Shoemaker A, et
al: An inhibitor of Bcl-2 family proteins induces regression of
solid tumours. Nature. 435:677–681. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gandhi L, Camidge D, Ribeiro de Oliveira
M, et al: Phase I study of Navitoclax (ABT-263), a novel Bcl-2
family inhibitor, in patients with small-cell lung cancer and other
solid tumors. J Clin Oncol. 29:909–916. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rudin C, Hann C, Garon E, et al: Phase II
study of single-agent navitoclax (ABT-263) and biomarker correlates
in patients with relapsed small cell lung cancer. Clin Cancer Res.
18:3163–3169. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Konopleva M, Contractor R, Tsao T, et al:
Mechanisms of apoptosis sensitivity and resistance to the BH3
mimetic ABT-737 in acute myeloid leukemia. Cancer Cell. 10:375–388.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Billard C: BH3 mimetics: status of the
field and new developments. Mol Cancer Ther. 12:1691–1700. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Goldsmith K, Gross M, Peirce S, et al:
Mitochondrial Bcl-2 family dynamics define therapy response and
resistance in neuroblastoma. Cancer Res. 72:2565–2577. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Klymenko T, Brandenburg M, Morrow C, Dive
C and Makin G: The novel Bcl-2 inhibitor ABT-737 is more effective
in hypoxia and is able to reverse hypoxia-induced drug resistance
in neuroblastoma cells. Mol Cancer Ther. 10:2373–2383. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang G: Medical uses of mylabris in
ancient China and recent studies. J Ethnopharmacol. 26:147–162.
1989. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang E, Tang W, Zhang K, Cheng L and Mack
PO: Norcantharidin inhibits growth of human HepG2 cell-transplanted
tumor in nude mice and prolongs host survival. Cancer Lett.
117:93–98. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kok SH, Cheng SJ, Hong CY, et al:
Norcantharidin-induced apoptosis in oral cancer cells is associated
with an increase of proapoptotic to antiapoptotic protein ratio.
Cancer Lett. 217:43–52. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang S, Li G, Ma X, et al: Norcantharidin
enhances ABT-737-induced apoptosis in hepatocellular carcinoma
cells by transcriptional repression of Mcl-1. Cell Signal.
24:1803–1809. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Huang Y, Liu Q, Liu K, Yagasaki K and
Zhang G: Suppression of growth of highly-metastatic human breast
cancer cells by norcantharidin and its mechanisms of action.
Cytotechnology. 59:201–208. 2009. View Article : Google Scholar
|
|
23
|
Yang J, Liu X, Bhalla K, et al: Prevention
of apoptosis by Bcl-2: release of cytochrome c from mitochondria
blocked. Science. 275:1129–1132. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Green DR and Kroemer G: The
pathophysiology of mitochondrial cell death. Science. 305:626–629.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mazumder S, Choudhary G, Al-Harbi S and
Almasan A: Mcl-1 phosphorylation defines ABT-737 resistance that
can be overcome by increased NOXA expression in leukemic B cells.
Cancer Res. 72:3069–3079. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Okumura K, Huang S and Sinicrope F:
Induction of Noxa sensitizes human colorectal cancer cells
expressing Mcl-1 to the small-molecule Bcl-2/Bcl-xL inhibitor,
ABT-737. Clin Cancer Res. 14:8132–8142. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Simonin K, N’Diaye M, Lheureux S, et al:
Platinum compounds sensitize ovarian carcinoma cells to ABT-737 by
modulation of the Mcl-1/Noxa axis. Apoptosis. 18:492–508. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zinn R, Gardner E, Dobromilskaya I, et al:
Combination treatment with ABT-737 and chloroquine in preclinical
models of small cell lung cancer. Mol Cancer. 12:Mar 2–2013.
View Article : Google Scholar
|
|
29
|
Yang PY, Chen MF, Kao YH, et al:
Norcantharidin induces apoptosis of breast cancer cells:
involvement of activities of mitogen activated protein kinases and
signal transducers and activators of transcription. Toxicol In
Vitro. 25:699–707. 2011. View Article : Google Scholar : PubMed/NCBI
|