Introduction
Thyroid cancer, although a rare disease, is one of
the most common endocrine malignancies, and its incidence is
rapidly increasing worldwide (1).
Despite the considerable developments in surgical technique and
other therapeutic strategies, the survival rate of patients with
thyroid cancer remains low due to its rapid progression and
metastasis (2). It is critical to
find new methods to treat thyroid cancer (3).
Several reports have shown that signal transducers
and activators of transcription 3 (Stat3), a member of STAT family,
play a key role in carcinogenesis by promoting cell proliferation,
differentiation and cell cycle progression, as well as inhibition
of apoptosis (4,5). In addition, constitutive activation of
Stat3 and its overexpression have been detected in a wide variety
of human tumors, including prostate (6), breast (7), leukemia (8), lung (9), thyroid cancer (10) and squamous cell carcinoma of the
head and neck (11). Targets
inhibiting the expression of Stat3 by RNA interferences could
inhibit the proliferation and induce the apoptosis of cancer cells
in vitro or suppress tumor growth in a mouse model (12–14).
These data suggest that STAT3 may be a new target in the therapy of
thyroid cancer.
Gene associated with retinoid-IFN-induced mortality
19 (GRIM-19), an IFN/RA-inducible gene product, was recently
identified as a potential tumor suppressor that promotes
IFN/RA-induced cell death (15). It
has been shown that overexpression of GRIM-19 could inhibit cell
proliferation and induce apoptosis in human prostate, breast,
gastric cancer and renal carcinoma cells (16–19).
Moreover, GRIM-19 has been shown to be an inhibitor of signal
transducer and activator of transcription 3 (Stat3) by binding to
Stat3 and suppressing its transcriptional activation (20,21).
Taken together, these studies indicate that GRIM9 is a potential
tumor suppressor (22).
In the present study, the plasmid expressing GRIM-19
and Stat3-specific short hairpin RNA (p-Si-Stat3-GRIM-19) was
transfected into SW579 cells to examine the effect of co-expression
of GRIM-19 and Stat3-specific short hairpin RNA on cell
proliferation, cell apoptosis, cell migration and cell invasion in
a human thyroid carcinoma cell line (SW579 cells) in vitro
and on tumor growth in a thyroid cancer xenograft in
vivo.
Materials and methods
Plasmid and cell
Plasmid pSilencerTMneo3.1-H1-Stat3- siRNA
(pSi-Stat3), plasmid pGCsilencerTMneo3.1-H1-scramble
(pSi-Scramble), pcDNA3.1-GRIM-19 (pGRIM-19) and Co-expression
plasmid pcDNA3.1-GRIM-19-Si-Stat3 (pSi-Stat3-GRIM-19) were granted
for the Prostate Diseases Prevention and Treatment Research Center
and the Department of Pathophysiology, School of Basic Medicine,
Jilin University (Changchun, China). The human thyroid carcinoma
cell line, SW579, was obtained from the Cell Bank of the Chinese
Academy of Sciences (Shanghai, China).
Cell culture and transfection
The human thyroid carcinoma cell line SW579 was
grown in L-15 (HyClone, Logan, UT, USA) with 10% (v/v) fetal bovine
serum (FBS; HyClone). SW579 cells were transfected with various
plasmids using the Lipofectamine 2000 reagent (Invitrogen,
Carlsbad, CA, USA) according to the manufacturer’s instructions for
an additional 48–72 h before analysis of mRNA and protein levels,
cell apoptosis and cell proliferation.
Semi-quantitative reverse
transcription-PCR (RT-PCR)
The mRNA expression levels of Stat3, GRIM-19 and
related genes were examined using semi-quantitative RT-PCR. Cells
transfected with pSi-Stat3, pGRIM-19 or pSi-Stat3-GRIM-19 plasmids
were collected after 48 h. Total RNA was extracted using the TRIzol
reagent (Invitrogen). Reverse transcription was performed with 5 μg
of total RNA purified after DNAse I treatment using a commercially
available RT-PCR kit (Takara, Dalian, China), based on the
manufacturer’s instructions. GAPDH was used as control. Stat3
sense, 5′-GAGTCAGGCACTGTGGG-3′ and antisense,
5′-CGGTCGGTTTCTGCCTGTA-3′; GRIM-19 sense, 5′-TTGCCAGTTGTGGTGATC-3′
and antisense, 5′-AGACCCAGAAGGAGCCGC-3′; GAPDH sense,
5′-CCTTCATTGACCTCAACTA-3′ and antisense,
5′-GGAAGGCCATGCCAGTGAGC-3′.
Western blot analysis
Antibodies against Stat3, p-Stat3, survivin, Bcl-2,
GRIM-19, matrix metalloproteinase-2 (MMP-2), MMP-9 and β-actin were
obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Anti-rabbit or anti-mouse secondary horseradish
peroxidase-conjugated were bought from Amersham Biosciences
(Uppsala, Sweden). For western blot analyses, cells were harvested
at 48 h after transfection and lysed with lysis buffer (Takara,
Dalian, China). After centrifugation at 15,000 × g for 30 min, the
supernatants were analyzed for protein content using Bradford
reagent (Bio-Rad Laboratories, Hercules, CA, USA). After boiling at
100°C for 10 min, a total of 40 mg of protein mixed with SDS-PAGE
buffer were loaded onto 8–12% SDS-PAGE gel for electrophoresis. The
proteins were separated and then transferred onto polyvinylidene
difluoride membranes (Millipore Corp., Bedford, MA, USA). The
membranes were blocked in 5% non-fat milk at 37°C for 2 h and were
incubated with the primary antibodies at 4°C overnight. After
washing three times with TBST buffer, the membranes were incubated
with HRP-labeled anti-rabbit IgG or HRP-labeled anti-mouse IgG
secondary antibody at 37°C for 2 h. The membranes were subsequently
washed thoroughly with TBST buffer. The antibody-bound bands were
visualized using ECL reagents (ECL; Amersham, GE Healthcare,
Velizy-Villacoublay, France) to detect protein. The absorbances of
the positive bands in the analysis were measured by densitometry
using a GIS Analysis System (Tannon, Shanghai, China).
Measurement of SW579 cell viability
To measure the effect of plasmid pSi-Stat3, pGRIM-19
or co-expression plasmids pSi-Stat3-GRIM-19 on cell proliferation,
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay was used. The cell density of SW579 cells was adjusted to
5×104/ml, and added to a 96-well plate (100 μl/well). In
the blank controls, 100 μl of medium alone was added. At 24 h after
culture, cells were transfected with different plasmids. At 48 h
after culture, 20 μl of MTT (5 mg/ml) was added to each well
followed by incubation for 48 h at 37°C. Then, centrifugation was
performed at 2,000 × g for 10 min. The supernatant was removed, and
200 μl of dimethyl sulfoxide (DMSO) was added to each well followed
by shaking for 10 min. Absorbance was measured at 570 nm with a
microplate reader (Molecular Devices Corp., Sunnyvale, CA, USA) and
growth inhibition was calculated. The mean proliferation of cells
without any treatment was expressed as 100%.
Detection of apoptosis
SW579 cells were cultured in 6-well plates in L-15
containing 10% FBS medium and were treated with plasmid pSi-Stat3,
pGRIM-19 or co-expression plasmid pSi-Stat3-GRIM-19 for 48 h,
respectively. The coverslips were washed three times with
phosphate-buffered saline (PBS; pH 7.2) and single cell suspensions
were fixed in 1% PBS. Cells were stained with 100 μg/ml acridine
orange (AO) and 100 μg/ml ethidium bromide (EB) for 1 min. Then,
cells were observed under a fluorescence microscope. At least 200
cells were counted and the percentage of apoptotic cells was
calculated.
In addition, in the present study, we also detected
survivin and Bcl-2 protein expression by western blotting as an
additional indicator of apoptosis.
Wound healing assay
To assess the effect of pSi-Stat3, pGRIM-19 or
co-expression plasmid pSi-Stat3-GRIM-19 on cell migration, wound
healing assay was performed. SW579 cells (1×105) were
plated in 12-well plates in complete growth medium. After 24 h of
growth, a scratch was made through the confluent cell monolayer,
and then the cells were treated with the indicated plasmid in 3 ml
of complete medium. At 48 h post treatment, cells were stained with
hematoxylin and eosin (H&E). Cells invading the wound line were
observed under an inverted phase-contrast microscope (Leica DMR,
Wetzlar, Germany).
Cell invasion assay
Cell invasion was determined using Transwell
chambers. Transwell filters in 6-well plates were coated with
Matrigel, hydrated for ~2 h in the tissue culture incubator with
500 μl serum-free culture media in the bottom and 500 μl in the top
of the chamber. After hydration of the Matrigel, 5×105
SW579 cells were plated in 500 μl serum-free L-15 medium on the top
chamber, while 2 ml L-15 medium containing 10% FCS were placed in
the lower chambers. Indicated plasmids were added to the upper
chambers, respectively. Cells without any treatment were used as
control. After 48 h of incubation, the filters were removed, washed
three times with PBS and then fixed in 10% formalin for 15 min.
After fixing at room temperature, the chambers were rinsed in PBS
and stained with 0.2% crystal violet staining solution for 30 min.
After washing the chambers using PBS, the cells at the top of the
Matrigel membrane were carefully removed by a number of cotton
swabs. At this time, all cells that remain are the ones that have
invaded to the bottom side of the membrane. Cell invasion was
observed with an immunofluorescence microscope by counting the
cells that had invaded into the bottom of the Cell Culture Insert.
In addition, in the present study, we also detected VEGF, MMP-9 and
MMP-2 protein expression by enzyme-linked immunospecific assay
(ELISA) or western blotting as an additional indicator of
invasion.
ELISA
SW579 cells grown in 24-well plates were transfected
with the indicated plasmid for 72 h. Protein levels of vascular
endothelial growth factor (VEGF) in the cell supernatant were
determined by Human VEGF ELISA kit (Yanyu, Shanghai, China)
according to the manufacturer’s instructions. Samples were measured
in triplicate and were properly diluted to ensure that measured
values were within the concentration range of the standard
curve.
Tumor growth in vivo
Female BALB/c nude mice (Jilin Institute of
Experimental Animals) were inoculated with 2×106 cells
of SW579 cells s.c. into the right flank of the mice. Tumor size
was measured every 2–3 days, and tumor volume calculated as 0.5236
× width2 × length. When tumors grew to an average volume
of 75 mm3, mice were randomly divided into five groups
(n=10) and inoculated with 20 μg/50 μl per mouse via i.t. injection
of different plasmids. Immediately after injection, tumors were
pulsed with an electroporation generator (ECM 830, BTX). Pulses
were delivered at a frequency of 1/sec, 150 V/cm, with a length of
50 ms. Mice were sacrificed on day 36, tumor weight and volume was
determined, and the tumor blocks were evaluated by RT-PCR assay.
Mock mice received implanted tumor but did not receive any plasmid
construct therapy or electroporation.
On the 36th day, the animals were euthanized using
chloroform and their spleen tissues were collected and cultured for
a splenocyte surveillance study. Spleens from treated mice were
collected and single-cell spleen suspensions were pooled in
serum-free L-15 by filtering the suspension through a sieve mesh
with the aid of a glass homogenizer to exert gentle pressure on the
spleen fragments. The detailed assay of splenocyte proliferation
was based on one previously described (23).
Statistical analysis
Data from at least three independent experiments are
expressed as means ± SD. Statistical comparison of more than two
groups was performed using one-way ANOVA followed by a Tukey’s post
hoc test. Statistical analyses were undertaken using the
SPSS® statistical package, version 19.0 (SPSS, Inc.,
Chicago, IL, USA) and the GraphPad Prism version 5.01 (GraphPad
Software, San Diego, CA, USA) for Windows®. P<0.05
was considered to indicate a statistically significant
difference.
Results
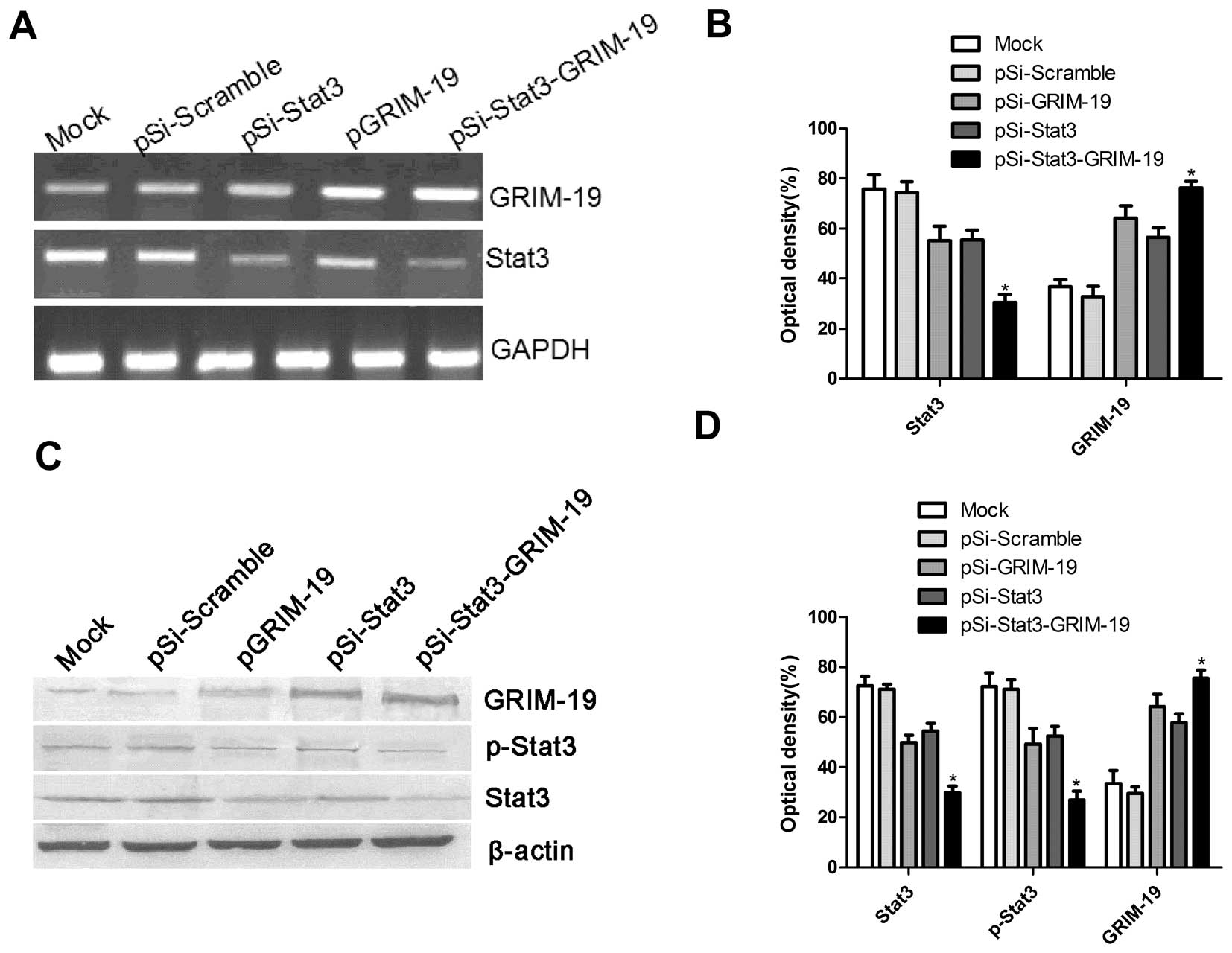
Effect of pSi-Stat3-GRIM-19 on mRNA and
protein expression of Stat3 and GRIM-19
We had three plasmids (pSi-Stat3, pGRIM-19 and
pSi-Stat3-GRIM-19) that are capable of expressing an shRNA that
targets the Stat3, tumor suppressor GRIM-19 either alone or in
combination. These plasmids were transfected into SW579 cells, a
thyroid carcinoma cell line, and their expression was determined
using western blot analysis and RT-PCR analyses. It was found that
Stat3 expression on mRNA level and protein level significantly
decreased after transfection of expression vectors pGRIM-19,
pSi-Stat3 and pGRIM-19-Si-Stat3, and that GRIM-19 expression on
mRNA level and protein level was significantly upregulated after
transfection with pGRIM-19, pGRIM-19-Si-Stat3 compared to those
that received Mock or pSi-Scramble via transfection (Fig. 1).
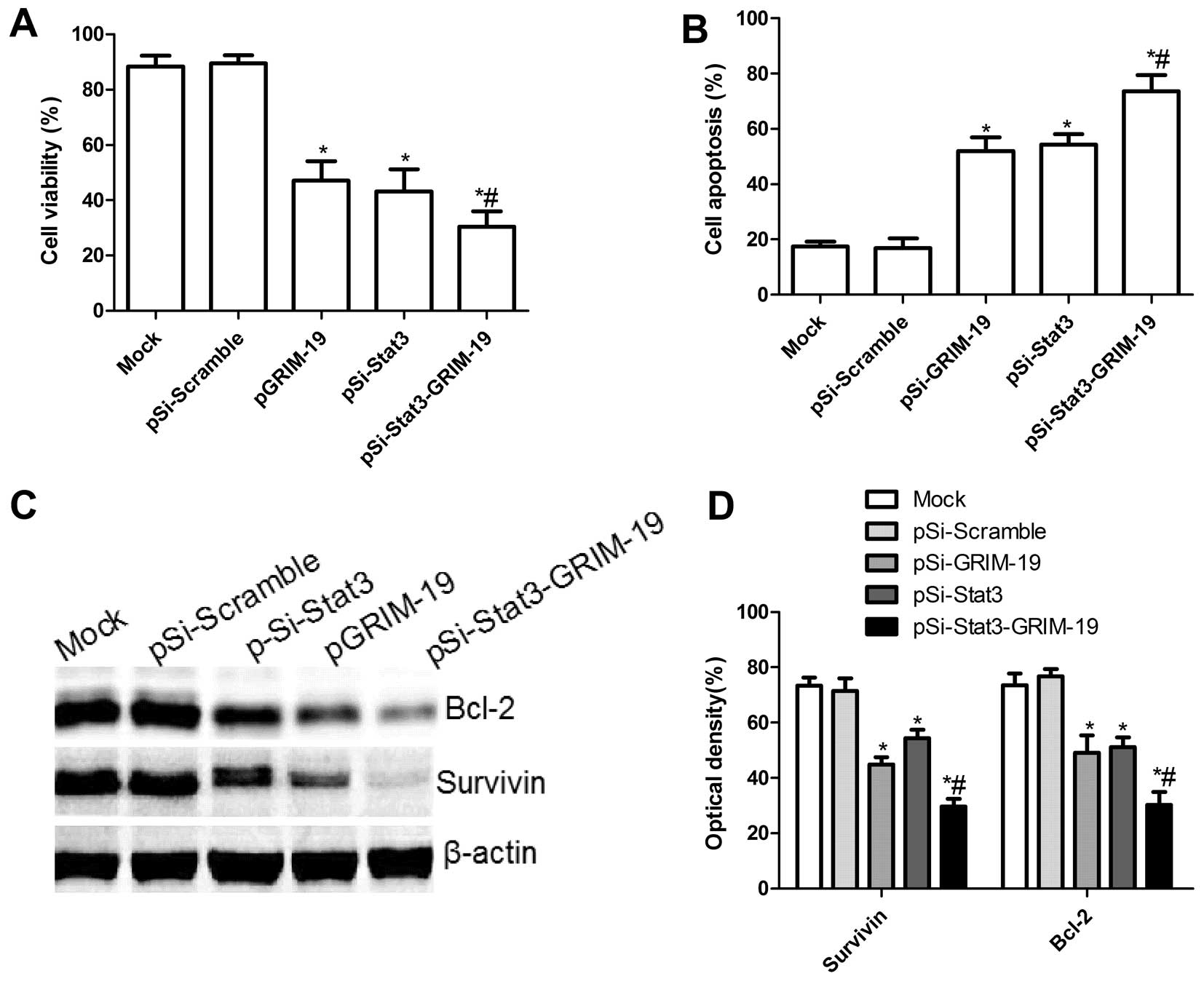
Effect of pSi-Stat3-GRIM-19 on cell
proliferation and apoptosis in SW579 cells
To investigate if plasmid pSi-Stat3, pGRIM-19 and
pSi-Stat3-GRIM-19 exert significantly different effects on cell
proliferation, MTT assay was performed for 72 h when SW579 were
transfected with individual expression vectors. Cell proliferation
in the pSi-Stat3, pGRIM-19 and pSi-Stat3-GRIM-19 groups was
significantly diminished compared to the Mock and pSi-Scramble
groups (P<0.05; Fig. 2A). Among
the SW579 cell groups treated with pSi-Stat3, pGRIM-19 and
pSi-Stat3-GRIM-19, the lowest incidence of cell proliferation was
observed in the pSi-Stat3-GRIM-19 treatment group. There was no
significant difference between the pSi-Stat3 and the pGRIM-19 group
(P>0.05).
To investigate whether plasmid pSi-Stat3, pGRIM-19
and pSi-Stat3-GRIM-19 could induce apoptosis, we analyzed the
apoptosis after treatment with pSi-Stat3, pGRIM-19 and
pSi-Stat3-GRIM-19. It was found that SW579 cells treated with the
pSi-Stat3 and the pGRIM-19 group significantly induced cell
apoptosis compared to the Mock and pSi-Scramble groups (P<0.05;
Fig. 2B). Treatment with
pSi-Stat3-GRIM-19 led to a marked increase in apoptotic cells
compared to the pSi-Stat3 and the pGRIM-19 group (P<0.05;
Fig. 2B).
To determine the potential mechanism of cell growth
inhibition in vitro, apoptosis inhibiting genes, survivin
and BCL-2, protein expression were detected using western blots.
Survivin and BCL-2 protein expression was significantly decreased
in pSi-Stat3, pGRIM-19 and pSi-Stat3-GRIM-19 treatment groups,
compared to the controls-Mock and pSi-Scramble groups (P<0.05;
Fig. 2C and D). Furthermore, the
group transfected with pSi-Stat3-GRIM-19 showed the most reduced
expression (Fig. 2C and D).
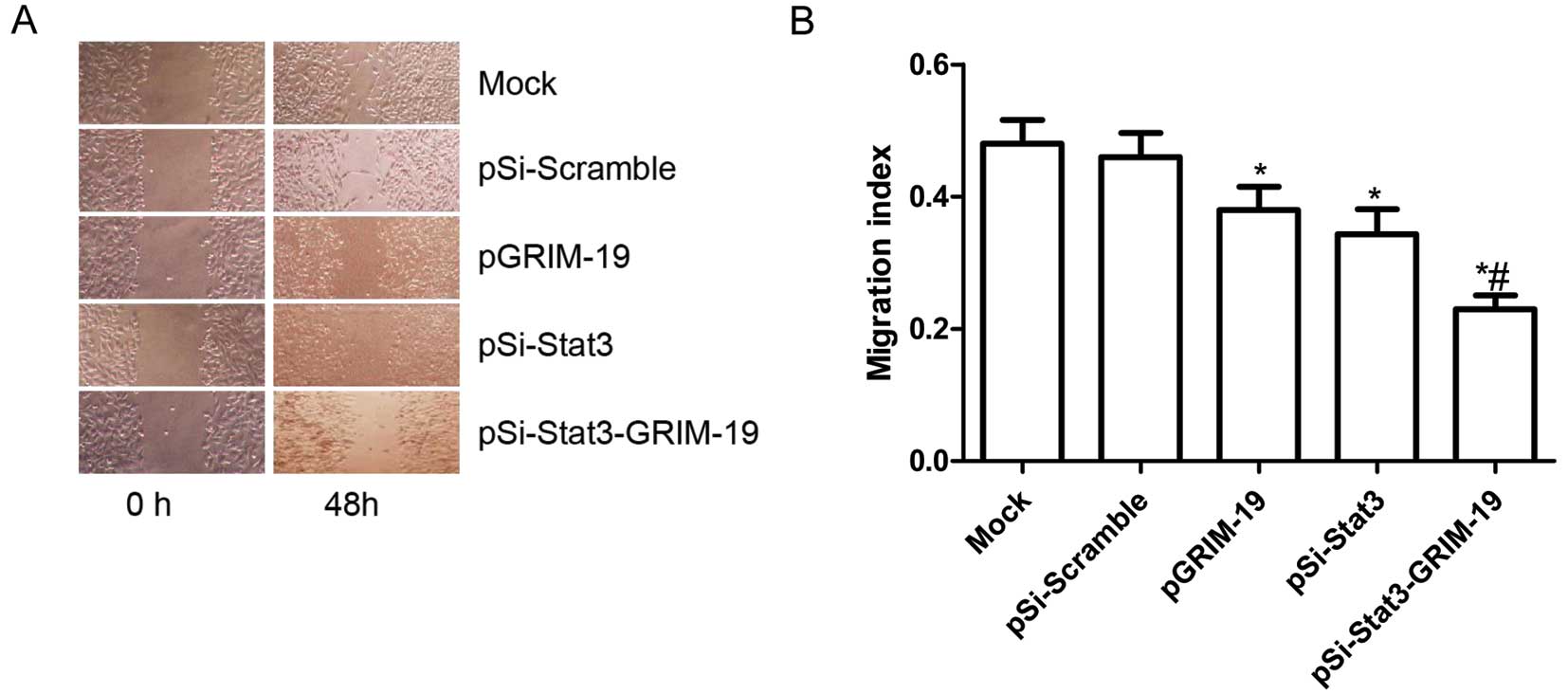
Effects of pSi-Stat3-GRIM-19 on cell
migration in SW579 cells
To ascertain the inhibitory effect of pSi-Stat3,
pGRIM-19 and pSi-Stat3-GRIM-19 on thyroid cancer on cell motility
in vitro, wound healing assay was performed to investigate
their effects on the migration potential of SW579 cells. A scratch
was introduced into confluent monolayers expressing different
treatment plasmids, and the time-dependent movement of cells into
the injured area was monitored microscopically. Cells in the Mock
and pSi-Scramble groups began migrating 8 h after scratching. There
was no significant change between 8 and 12 h. After 48 h, cells in
the pSi-Stat3 group, the pGRIM-19 group and the pSi-Stat3-GRIM-19
group migrated significantly less than those in the Mock and the
pSi-Scramble groups. Compared to the pSi-Stat3 or the pGRIM-19
groups, the cells in the pSi-Stat3-GRIM-19 group significantly
decreased in migration in SW579 cells (P<0.05; Fig. 3A and B).
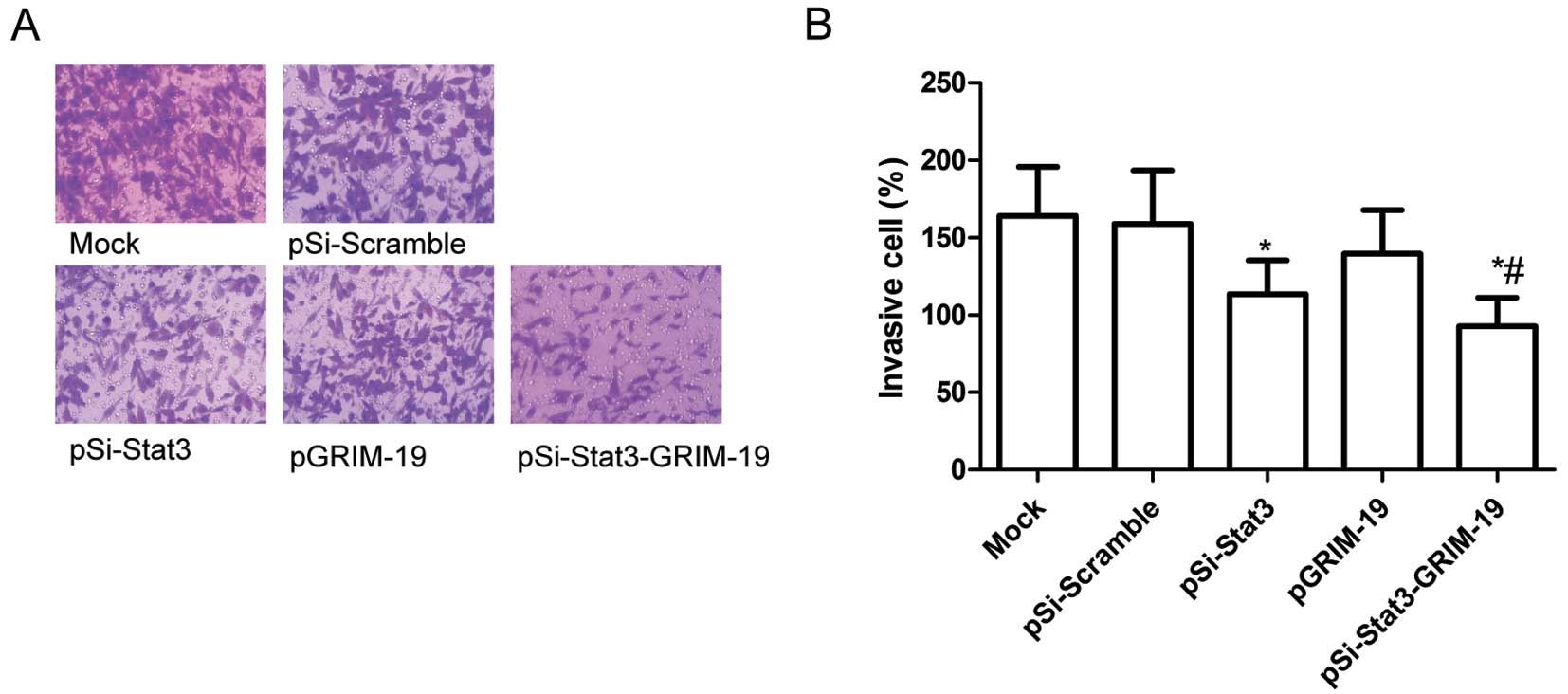
Effects of pSi-Stat3-GRIM-19 on cell
invasion in SW579 cells
The ability of pSi-Stat3, pGRIM-19 and
pSi-Stat3-GRIM-19 to reduce the invasiveness of thyroid cancer
cells was further investigated by the Transwell system assay. It
was found that invasion was also decreased significantly in the
pSi-Stat3 group, the pGRIM-19 group and the pSi-Stat3-GRIM-19
treatment group compared to the Mock and pSi-Scramble groups
(P<0.05; Fig. 4A and B).
Compared to the pSi-Stat3 or the pGRIM-19 groups, the
pSi-Stat3-GRIM-19 treatment group greatly inhibited SW579 cell
invasion (P<0.05; Fig. 4A and
B).
pSi-Stat3-GRIM-19 inhibits the
invasion-related protein in SW579 cells
To determine the potential mechanism of cell
migration inhibition and cell invasion inhibition in vitro,
invasion-associated protein expression was examined using ELISA and
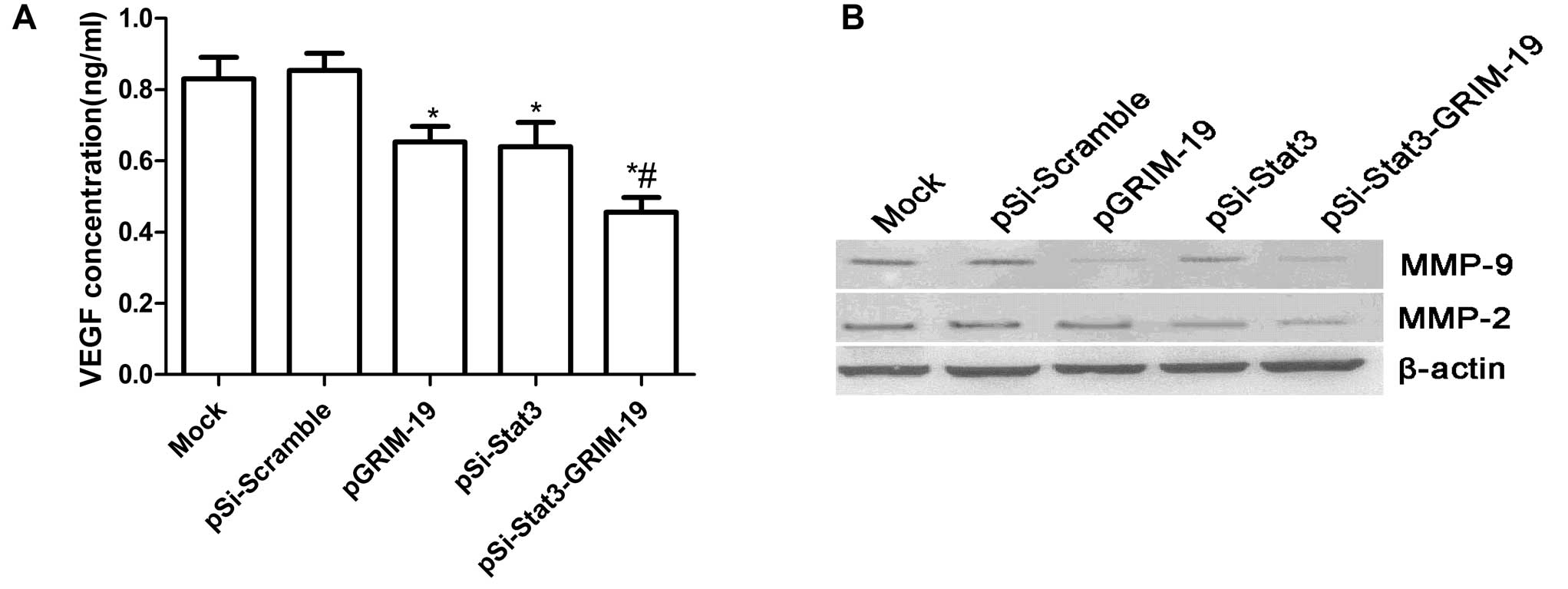
western blots. As shown in Fig. 5A,
ELISA analysis revealed that VEGF excretion in the supernatant from
pSi-Stat3 group, the pGRIM-19 group and the pSi-Stat3-GRIM-19 group
was significantly decreased (P<0.05), while no obvious changes
were observed in the Mock and pSi-Scramble groups. Western blot
analysis displayed a significant decrease in MMP-2, MMP-9 proteins
in the pSi-Stat3 group, the pGRIM-19 group and the
pSi-Stat3-GRIM-19 group infected SW579 cells compared to Mock and
pSi-Scramble groups (Fig. 5B). The
pSi-Stat3-GRIM-19 group showed maximally reduced expression
compared to either the pSi-Stat3 or pGRIM-19 groups (Fig. 5B). In conclusion, these results
suggest that the pSi-Stat3-GRIM-19 inhibitory effect on metastasis
of thyroid cancer was at least partially mediated by the
downregulation of MMP-9, MMP-2 and VEGF, which may contribute to
degradation of the extracellular matrix.
Co-expressed Stat3-shRNA and GRIM-19
synergistically suppress tumor growth in vivo
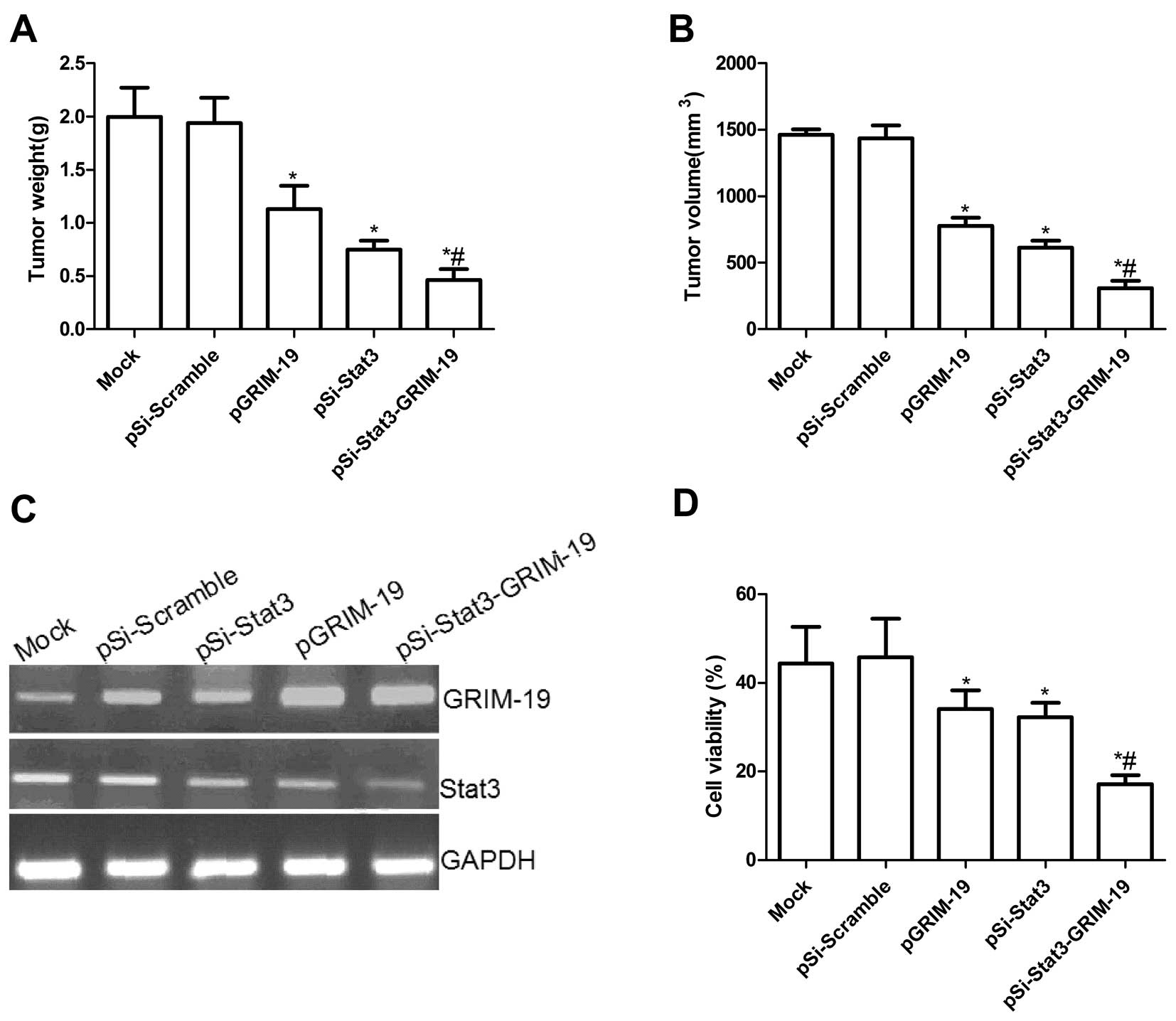
We next determined if the co-expressed GRIM-19 and
siRNA-Stat3 (pSi-Stat3-GRIM-19) could synergistically inhibit tumor
growth by a xenograft tumor model. Tumor growth was monitored for
36 days. On day 36, animals were sacrificed and final tumor weights
and tumor volume were determined. It was found that the tumor
weight was significantly lower in various treatment groups than in
the Mock and pSi-Scramble groups (P<0.05; Fig. 6A). Compared to either the pSi-Stat3
or the pGRIM-19 groups, the pSi-Stat3-GRIM-19 group showed
maximally reduced weight. Tumor volume in various treatment groups
was significantly (P<0.05) diminished when compared with the
scramble control. Compared with pGRIM-19 and pSi-Stat3, the
pSi-Stat3-GRIM-19 group had an obvious effect on tumor growth
(P<0.01; Fig. 6B). In addition,
in the present study, we also examined the expression of Stat3 and
GRIM-19 in grafted tumor tissues by RT-PCR analysis. It was found
that GRIM-19 expression levels were increased in the groups treated
with the pGRIM-19 and pSi-Stat3-GRIM-19 plasmid (Fig. 6C) and that Stat3 expression
decreased in the groups treated with pGRIM-19, pSi-Stat3 or
pSi-Stat3-GRIM-19 (P<0.05; Fig.
6C). We also assessed the efficacy of co-expressed GRIM-19 and
siRNA-Stat3 (pSi-Stat3-GRIM-19) in modulating splenocyte
proliferation using MTT assay. As shown in Fig. 6D, the inhibitory rates of pSi-Stat3,
pGRIM-19 and pSi-Stat3-GRIM-19 significantly increased compared to
the Mock group and pSi-Scramble group (P<0.01). The inhibitory
rates of the pSi-Stat3-GRIM-19 group were higher than those of the
pSi-Stat3 and the pGRIM-19 group. These results suggested that
co-expressed GRIM-19 and siRNA-Stat3 synergistically inhibited
tumor growth in vivo.
Discussion
It has been shown that abnormal apoptosis plays an
important role in the development and progression of cancer
(24) and therefore, controlling
the apoptosis of cancer cells has important biological and clinical
significance (25,26). Studies have shown Stat3 induced
activation via tyrosyl phosphorylation by larger number of
different oncogenic events. A large number of studies showed that
activated Stat3 could improve cell proliferation and inhibit cell
apoptosis (27–31). In addition, aberrantly active Stat3
promotes tumor cell growth and survival via an incessant induction
of pro-growth genes, such as cyclin D1, c-Myc, survivin, Bcl-xL,
Bcl-2, Mcl-1, VEGF and MMP-2 and MMP-9 (27–32),
whose products promote tumor cell proliferation, cell cycle
progression, metastasis, as well as inhibit apoptosis. Therefore,
downregulation of Stat3 expression could inhibit the proliferation
and induce apoptosis. In the present study, our results showed for
the first time that down-regulation of Stat3 by RNA interference in
thyroid cancer cells significantly suppressed the proliferation,
migration and invasion and induced cell apoptosis in vitro,
and inhibited tumor growth in vivo. These results were
agreement with previous results (12–14).
Although downregulation of Stat3 has been
successfully used for the suppression of tumor growth in
vitro or in vivo (12–14),
the therapeutic efficacy in vivo has not been fully assessed
(12). It is well known that RNA
interference does not completely block gene expression, especially
when the target mRNA is expressed at abnormally high levels
(33). Therefore, in the present
study, we chose the plasmid coexpressing GRIM-19 and Stat3-specific
short hairpin RNA (pSi-Stat3-GRIM-19) to inhibit thyroid tumor
growth in vitro and in vivo. GRIM-19, a
Stat3-inhibitory protein, has been shown to interact exclusively
with the transcriptional activating domain of Stat3 and to inhibit
autoregulatory Stat3 driven transcriptional activation (15,20,21).
Alchanati et al (19) showed
that reduction of GRIM-19 expression results in upregulation of
Stat3-regulated genes in these tumors, and restoration of GRIM-19
suppresses the growth-promoting activities of Stat3. GRIM-19 has
been identified as a potential tumor suppressor that promotes
IFN/RA-induced cell death (15),
and overexpression of GRIM-19 induces apoptosis, inhibit tumor
growth in various types of cancer (16–19).
Notably, Wen et al (34)
demonstrated that co-expressed survivin-shRNA and GRIM-19 can
induce the apoptosis of laryngeal cancer cells and inhibit their
proliferation, and co-expressed survivin-shRNA and GRIM-19 is more
effective than psi-survivin and p-GRIM-19. Zhang et al
(16) reported that the
co-expressed Stat3-specific shRNA and GRIM-19 synergistically and
more effectively suppressed prostate tumor growth and metastases
when compared with treatment with either single agent alone, which
was in agreement with our results. In the present study, our
results also showed that simultaneous expression of Stat3-specific
shRNA and GRIM-19 in thyroid cancer cells significantly suppressed
the proliferation, migration and invasion in vitro and tumor
growth in vivo, when compared to the controls either
Stat3-specific siRNA or GRIM19 alone. Our results as well as those
of others showed that co-expressed Stat3-specific shRNA and GRIM-19
might be a more effective gene therapy for the treatment of various
cancers.
Extracellular matrix degradation is a key step in
tumor invasion and metastasis, and it is mainly mediated by the
secretion of MMP-2 (32), MMP-9
(35,36) and VEGF (30,37).
It has been reported that downregulation of MMP-2, MMP-9 and VEGF
contributes to inhibition of cancer cell invasion and metastasis
(38,39). In the present study, we found that
co-expressed Stat3-specific shRNA and GRIM-19 could suppress
thyroid cancer cell migration and invasion and inhibit MMP-2, MMP-9
and VEGF expression.
In conclusion, our results demonstrated that
simultaneous expression of pSi-Stat3-GRIM-19 in SW579 tumor cells
significantly suppressed thyroid cell proliferation, migration and
invasion and induced cell apoptosis in vitro, and inhibited
thyroid tumor growth in vivo, when compared to the controls
either Stat3-specific siRNA or GRIM19 alone. These results suggest
that co-expressed Stat3-specific shRNA and GRIM-19 appears to have
therapeutic potential for the treatment of thyroid cancer.
Acknowledgements
The present study was supported by the Science and
Technology Research and Innovation Team funded by the Jilin
Province (JL2013018).
References
|
1
|
Jeong SY, Kim HW, Lee SW, Ahn BC and Lee
J: Salivary gland function 5 years after radioactive iodine
ablation in patients with differentiated thyroid cancer: direct
comparison of pre- and postablation scintigraphies and their
relation to xerostomia symptoms. Thyroid. 23:609–616. 2013.
|
|
2
|
Siironen P, Hagstrom J, Maenpaa HO, et al:
Anaplastic and poorly differentiated thyroid carcinoma: therapeutic
strategies and treatment outcome of 52 consecutive patients.
Oncology. 79:400–408. 2010. View Article : Google Scholar
|
|
3
|
Sakorafas GH, Lappas C, Mastoraki A,
Kotsilianou O, Makras P and Safioleas M: Extensive and concurrent
infiltration of an ectopic intrathoracic thyroid and thyroid gland
by papillary thyroid cancer. Am Surg. 76:E152–E153. 2010.PubMed/NCBI
|
|
4
|
Darnell JE Jr: STATs and gene regulation.
Science. 277:1630–1635. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lu Y, Fukuyama S, Yoshida R, et al: Loss
of SOCS3 gene expression converts STAT3 function from
anti-apoptotic to pro-apoptotic. J Biol Chem. 281:36683–36690.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mora LB, Buettner R, Seigne J, et al:
Constitutive activation of Stat3 in human prostate tumors and cell
lines: direct inhibition of Stat3 signaling induces apoptosis of
prostate cancer cells. Cancer Res. 62:6659–6666. 2002.PubMed/NCBI
|
|
7
|
Li L and Shaw PE: Autocrine-mediated
activation of STAT3 correlates with cell proliferation in breast
carcinoma lines. J Biol Chem. 277:17397–17405. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Epling-Burnette PK, Liu JH,
Catlett-Falcone R, et al: Inhibition of STAT3 signaling leads to
apoptosis of leukemic large granular lymphocytes and decreased
Mcl-1 expression. J Clin Invest. 107:351–362. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Song L, Turkson J, Karras JG, Jove R and
Haura EB: Activation of Stat3 by receptor tyrosine kinases and
cytokines regulates survival in human non-small cell carcinoma
cells. Oncogene. 22:4150–4165. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kim EJ, Park JI and Nelkin BD: IFI16 is an
essential mediator of growth inhibition, but not differentiation,
induced by the leukemia inhibitory factor/JAK/STAT pathway in
medullary thyroid carcinoma cells. J Biol Chem. 280:4913–4920.
2005. View Article : Google Scholar
|
|
11
|
Grandis JR, Drenning SD, Zeng Q, et al:
Constitutive activation of Stat3 signaling abrogates apoptosis in
squamous cell carcinogenesis in vivo. Proc Natl Acad Sci
USA. 97:4227–4232. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gao L, Zhang L, Hu J, et al:
Down-regulation of signal transducer and activator of transcription
3 expression using vector-based small interfering RNAs suppresses
growth of human prostate tumor in vivo. Clin Cancer Res.
11:6333–6341. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ling X and Arlinghaus RB: Knockdown of
STAT3 expression by RNA interference inhibits the induction of
breast tumors in immunocompetent mice. Cancer Res. 65:2532–2536.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee SO, Lou W, Qureshi KM, Mehraein-Ghomi
F, Trump DL and Gao AC: RNA interference targeting Stat3 inhibits
growth and induces apoptosis of human prostate cancer cells.
Prostate. 60:303–309. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Angell JE, Lindner DJ, Shapiro PS, Hofmann
ER and Kalvakolanu DV: Identification of GRIM-19, a novel cell
death-regulatory gene induced by the interferon-β and retinoic acid
combination, using a genetic approach. J Biol Chem.
275:33416–33426. 2000.PubMed/NCBI
|
|
16
|
Zhang L, Gao L, Li Y, et al: Effects of
plasmid-based Stat3-specific short hairpin RNA and GRIM-19 on PC-3M
tumor cell growth. Clin Cancer Res. 14:559–568. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang Y, Yang M, Yang H and Zeng Z:
Upregulation of the GRIM-19 gene suppresses invasion and metastasis
of human gastric cancer SGC-7901 cell line. Exp Cell Res.
316:2061–2070. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang G, Chen Y, Lu H and Cao X: Coupling
mitochondrial respiratory chain to cell death: an essential role of
mitochondrial complex I in the interferon-β and retinoic
acid-induced cancer cell death. Cell Death Differ. 14:327–337.
2007.PubMed/NCBI
|
|
19
|
Alchanati I, Nallar SC, Sun P, et al: A
proteomic analysis reveals the loss of expression of the cell death
regulatory gene GRIM-19 in human renal cell carcinomas. Oncogene.
25:7138–7147. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lufei C, Ma J, Huang G, et al: GRIM-19, a
death-regulatory gene product, suppresses Stat3 activity via
functional interaction. EMBO J. 22:1325–1335. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang J, Yang J, Roy SK, et al: The cell
death regulator GRIM-19 is an inhibitor of signal transducer and
activator of transcription 3. Proc Natl Acad Sci USA.
100:9342–9347. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nallar SC, Kalakonda S, Lindner DJ, et al:
Tumor-derived mutations in the gene associated with retinoid
interferon-induced mortality (GRIM-19) disrupt its anti-signal
transducer and activator of transcription 3 (STAT3) activity and
promote oncogenesis. J Biol Chem. 288:7930–7941. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kumar BN, Rajput S, Dey KK, et al:
Celecoxib alleviates tamoxifen-instigated angiogenic effects by
ROS-dependent VEGF/VEGFR2 autocrine signaling. BMC Cancer.
13:2732013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: the next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chau BN and Wang JY: Coordinated
regulation of life and death by RB. Nat Rev Cancer. 3:130–138.
2003. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Panguluri SK, Yeakel C and Kakar SS: PTTG:
an important target gene for ovarian cancer therapy. J Ovarian Res.
1:62008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Turkson J: STAT proteins as novel targets
for cancer drug discovery. Expert Opin Ther Targets. 8:409–422.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Masuda M, Suzui M, Yasumatu R, et al:
Constitutive activation of signal transducers and activators of
transcription 3 correlates with cyclin D1 overexpression and may
provide a novel prognostic marker in head and neck squamous cell
carcinoma. Cancer Res. 62:3351–3355. 2002.
|
|
29
|
Alas S and Bonavida B: Rituximab
inactivates signal transducer and activation of transcription 3
(STAT3) activity in B-non-Hodgkin’s lymphoma through inhibition of
the interleukin 10 autocrine/paracrine loop and results in
down-regulation of Bcl-2 and sensitization to cytotoxic drugs.
Cancer Res. 61:5137–5144. 2001.PubMed/NCBI
|
|
30
|
Aoki Y, Feldman GM and Tosato G:
Inhibition of STAT3 signaling induces apoptosis and decreases
survivin expression in primary effusion lymphoma. Blood.
101:1535–1542. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wei D, Le X, Zheng L, et al: Stat3
activation regulates the expression of vascular endothelial growth
factor and human pancreatic cancer angiogenesis and metastasis.
Oncogene. 22:319–329. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xie TX, Wei D, Liu M, et al: Stat3
activation regulates the expression of matrix metalloproteinase-2
and tumor invasion and metastasis. Oncogene. 23:3550–3560. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Elbashir SM, Harborth J, Weber K and
Tuschl T: Analysis of gene function in somatic mammalian cells
using small interfering RNAs. Methods. 26:199–213. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wen LJ, Gao LF, Jin CS, et al: Small
interfering RNA survivin and GRIM-19 co-expression salmonella
plasmid inhibited the growth of laryngeal cancer cells in vitro and
in vivo. Int J Clin Exp Pathol. 6:2071–2081. 2013.PubMed/NCBI
|
|
35
|
Farina AR, Tacconelli A, Vacca A, Maroder
M, Gulino A and Mackay AR: Transcriptional up-regulation of matrix
metalloproteinase-9 expression during spontaneous epithelial to
neuroblast phenotype conversion by SK-N-SH neuroblastoma cells,
involved in enhanced invasivity, depends upon GT-box and nuclear
factor kappaB elements. Cell Growth Differ. 10:353–367. 1999.
|
|
36
|
Bond M, Fabunmi RP, Baker AH and Newby AC:
Synergistic upregulation of metalloproteinase-9 by growth factors
and inflammatory cytokines: an absolute requirement for
transcription factor NF-κB. FEBS Lett. 435:29–34. 1998.PubMed/NCBI
|
|
37
|
Yang EV, Kim SJ, Donovan EL, et al:
Norepinephrine upregulates VEGF, IL-8, and IL-6 expression in human
melanoma tumor cell lines: implications for stress-related
enhancement of tumor progression. Brain Behav Immun. 23:267–275.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Braicu EI, Gasimli K, Richter R, et al:
Role of serum VEGFA, TIMP2, MMP2 and MMP9 in monitoring response to
adjuvant radiochemotherapy in patients with primary cervical cancer
- results of a companion protocol of the randomized NOGGO-AGO Phase
III Clinical Trial. Anticancer Res. 34:385–391. 2014.
|
|
39
|
Lai WW, Hsu SC, Chueh FS, et al: Quercetin
inhibits migration and invasion of SAS human oral cancer cells
through inhibition of NF-κB and matrix metalloproteinase-2/-9
signaling pathways. Anticancer Res. 33:1941–1950. 2013.PubMed/NCBI
|