Introduction
Hypoxia is a major determinant of the malignant
progression of tumor cells and their response to therapy (1). Clinical studies have shown a strong
correlation between tumor pO2, local control and overall
survival (2,3). Since the vast majority of human solid
tumors have median pO2 levels lower than that of the
surrounding normal tissues (4),
efforts have been made to exploit it as a target for cancer
treatments. At the molecular level, hypoxia is a powerful modulator
of gene expression. An important mediator of hypoxic response is
the interaction of a transcriptional complex termed hypoxia
inducible factor (HIF) with its cognate DNA recognition site, the
hypoxia-response element (HRE) (5,6). The
HRE sequences isolated from oxygen-responsive genes have been shown
to selectively induce gene expression in response to hypoxia when
placed upstream of a promoter, and hypoxia-targeted gene therapy
can be achieved (7,8).
Two well-established gene-directed enzyme prodrug
therapy strategies, in which Herpes simplex virus 1-thymidine
kinase (HSV1-TK) or cytosine deaminase (CD) was used in combination
with prodrug ganciclovir (GCV) or 5-fluorocytosine (5-FC)
respectively, have been proposed for hypoxia-driven gene therapy in
different tumor models (9,10). Double suicide gene combined with
HSV1-TK/GCV and CD/5-FC resulted in enhanced anticancer effects and
has been used in clinical trials (11,12).
However, hypoxia-targeted double suicide gene therapy has not been
reported. Studies from us and other groups have demonstrated that
co-transduction of uracil phosphoribosyltransferase (UPRT) with CD
gene to cancer cells improved antitumor efficacy (13,14).
Furthermore, our studies demonstrated that hypoxia-targeted
expression of bifunctional suicide gene CDUPRT enhanced the
cytotoxicity of 5-FC treatments and radiosensitization under
hypoxic conditions (15). We also
found the superior antitumor effects of triple suicide gene
approach-TKCDUPRT over double gene approach-TKCD in a rat prostate
tumor model (16).
In the present study, we generated a human
colorectal cancer model expressing triple suicide gene (HSV1-TK, CD
and UPRT) under the control of the hypoxia inducible promoter.
Significant radiosensitization effects were demonstrated both in
vitro and in vivo with GCV and 5-FC treatments. Our data
suggest that the hypoxia-inducible triple suicide gene therapy has
the ability to specifically target hypoxic cancer cells and
significantly improves the tumor control in combination with
radiotherapy. This novel model also appears to be a valuable tool
to study tumor hypoxia, radiation effects and hypoxia-targeted
radio-gene therapy.
Materials and methods
Cell culture
Human colorectal cancer HCT-8 cells obtained from
the American Type Culture Collection (Manassas, VA, USA) were grown
in RPMI-1640 medium (Mediatech, Herndon, VA, USA) supplemented with
10% fetal bovine serum (FBS; Gemini Bio Products, West Sacramento,
CA, USA), 100 U/ml penicillin and 100 μg/ml streptomycin (Gemini).
For normoxic culture, cells were incubated in a humidified
atmosphere containing 21% O2 and 5% CO2; for
hypoxic culture, cells were incubated in an Invivo2 400
Hypoxic workstation (Ruskinn Inc., Cincinnati, OH, USA) with a gas
mixture of 0.5% O2, 5% CO2 and 94.5%
N2.
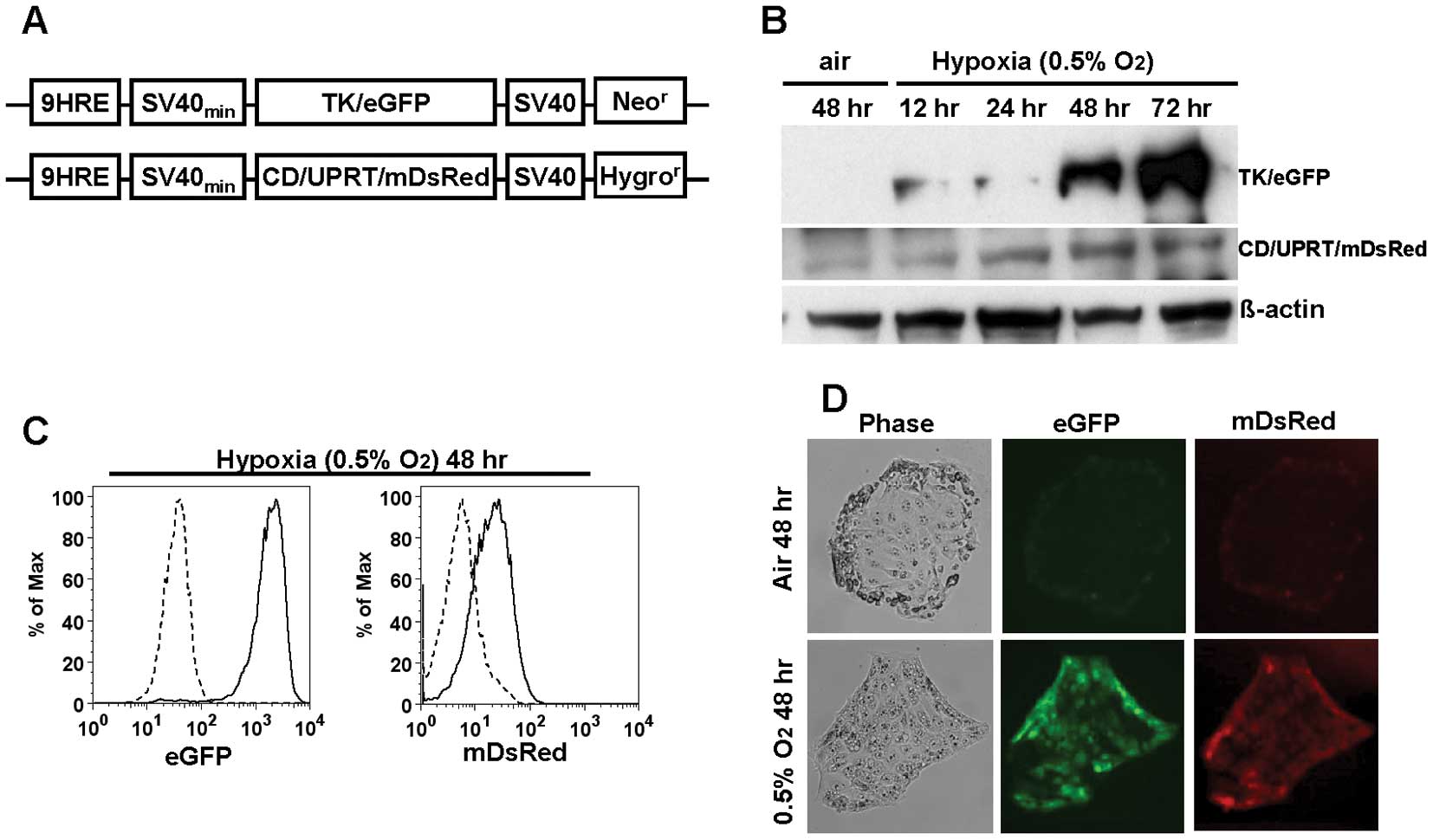
HCT-8 cells were co-transfected with two plasmids;
p9HRE-TK/eGFP, which contains the TK/eGFP fusion gene under the
control of a hypoxia-inducible promoter and a constitutively
expressed neomycin-resistance gene (Neor) (17), and p9HRE-CD/UPRT/mDsRed, which
contains the CD, UPRT and mDsRed fusion gene under the regulation
of the hypoxia-inducible promoter and a hygromycin B-resistance
gene (Hygr) (15)
(Fig. 1A). The hypoxia-inducible
promoter consists of nine tandem repeats of the HRE from human
erythropoietin gene linked to the SV40 minimal promoter (SV40min).
The stable cell line expressing HRE-regulated TK/eGFP and
CD/UPRT/mDsRed was established by fluorescence-activated cell
sorting using the cell sorter (MoFlo; Dako, Carpinteria, CA, USA)
(17) and designated as HCT8-HRE
cells. HCT-8 cells transfected with empty vector were used as the
control cells.
Western blot analysis
The cells were incubated under normoxic or 0.5%
O2 conditions for 12, 24, 48 or 72 h, and then whole
cell extracts were prepared. The protein was detected with the
sheep anti-yCD polyclonal antibody (Biotrend Chemikalien GmbH,
Cologne, Germany) and the anti-HSV1-TK monoclonal antibody (kindly
provided by Dr W.C. Summers, Yale University, New Haven, CT,
USA).
Flow cytometry
After incubation under normoxic or 0.5%
O2 conditions for 48 h, the cells were trypsinized,
centrifuged and re-suspended in PBS at 4°C. The fluorescence of
eGFP and mDsRed was measured using the cell sorter (MoFlo). The
data were analyzed with the FlowJo program (Tree Star Inc.,
Ashland, OR, USA).
Fluorescence microscopy
The cells were incubated under normoxic or 0.5%
O2 conditions for 48 h. The cells were then fixed with
freshly prepared 4% paraformaldehyde for 10 min and rinsed twice
with PBS. The fluorescent images were acquired with a fluorescence
microscope (Axiovert 200M, Zeiss).
Colony formation assay
The cells were treated with 5-FC (InvivoGen, San
Diego, CA, USA) or GCV (Sigma-Aldrich, St. Louis, MO, USA) at
various concentrations under normoxic or 0.5% O2
conditions for 48 h. The cells were then trypsinized, counted,
serially diluted and plated into 60-mm dishes. Clonogenic survival
was determined by counting crystal violet-stained colonies ~14 days
later. The survival fraction was normalized to the cell survival
without drug treatments and plotted as a function of the drug
concentration.
MTT assay
The cells were plated into 96-well plates
(104/well) and incubated with various concentrations of
GCV and/or 5-FC for 48 h under normoxic or 0.5% O2
conditions. The medium containing the drug was then replaced with
fresh medium, and cells were cultured for an additional 48 h under
the normoxic conditions. Tetrazolium dye (Sigma) was added and
allowed to react for 2 h at 37°C. After dissolving in DMSO, dye
conversion was read using an ELISA plate reader at 490 nm against
610 nm.
Bystander effect
The HCT-8 control cells were mixed with HCT8-HRE
cells at the 80:20 ratio and plated into 96-well plates. The cells
were treated with various concentrations of GCV and/or 5-FC for 48
h under 0.5% O2 conditions, and cell viability was
determined with the MTT assay described above.
Radiation survival assay
The cells were exposed to 5-FC (0.5 mg/ml) and GCV
(5 μg/ml) under normoxic or hypoxic conditions for 48 h, and then
irradiated for various doses using a Cs-137 unit (Mark 1 model 68;
J L Shepherd & Associates, San Fernando CA, USA) at ~2.0
Gy/min. Clonogenic survival was then determined by colony formation
assay. The radiation survival fractions were normalized for plating
efficiency using non-irradiated cells with or without drug
treatment and plotted as a function of radiation doses.
Xenograft model
Animal protocols were approved by the Institutional
Animal Care and Use Committee. Xenografts were formed by injecting
5×106 cells subcutaneously into the hind legs of
6–8-week old female nude mice (athymic nu/nu; NCI Frederick Cancer
Research Institute, Frederick, MD, USA). When the xenografts
reached ~10 mm in diameter, the mice were injected i.v. with
pimonidazole (80 mg/kg; HPI, MA). Two hours later, Hoechst 33342
(25 mg/kg; Sigma-Aldrich) was i.v. injected, and 1 min later, the
mice were sacrificed by CO2 breathing. Then, tumors were
excised and snap-frozen in OCT mounting medium (Sakura Finetek USA
Inc., Torrance, CA, USA). Frozen sections of 8-μm thickness were
prepared for the immunohistochemical analysis.
Immunohistochemical staining and
fluorescent microscopic imaging
Tumor sections were first imaged for the eGFP,
mDsRed and Hoechst 33342 fluorescence. They were then stained for
pimonidazole using FITC-conjugated murine anti-pimonidazole
monoclonal antibody (17). Finally,
the sections were stained with hematoxylin-eosin with the standard
protocol. Fluorescence images were acquired at ×50 magnification
using the fluorescence microscope (Axiovert 200 M) equipped with a
CCD digital camera, a computer-controlled motorized stage and
MetaMorph 7.0 Imaging software (Molecular Devices, Sunnyvale, CA,
USA).
Tumor growth delay
Each tumor was measured with digital caliper in
three orthogonal dimensions (a, b and c), and tumor volume was
calculated as πabc/6. When the tumor grew to ~100–150
mm3, the mice were divided into four groups (10
mice/group) and treated with: i) PBS, ii) 5-FC (i.p. 500 mg/kg) +
GCV (i.p. 30 mg/kg) daily for 14 days, iii) radiation (15 Gy) on
the 5th day, iv) 5-FC+GCV daily for 14 days followed by radiation
(15 Gy) on the 5th day, respectively. The tumor volume was measured
three times a week until the volume exceeded 1,000 mm3
and plotted as a function of time.
Statistical analysis
Averages are presented as the mean ± SE. Difference
in cytotoxicity and radiosensitivity was determined using the
Student’s t-test. A P-value of <0.01 was considered to indicate
a statistically significant difference.
Results
Hypoxia-induced expression of triple
suicide gene
HCT-8 stable cell lines expressing HRE driven
TK/eGFP and CD/UPRT/mDsRed were successfully established. Several
stable transfectants were established and their gene expressions
were similar as verified by western blot analysis, flow cytometry,
hence one of them was used for further experiments. Western blot
analysis showed that the expression level of TK/eGFP and
CD/UPRT/mDsRed protein was markedly increased after hypoxic
treatments in a time-dependent manner (Fig. 1B). Exposure of the HRE-TKCDUPRT
cells to hypoxia led to an increase of the hypoxia-regulated eGFP
and mDsRed fluorescence. In flow cytometric analysis, an 80-fold
increase in eGFP and 4-fold increase in mDsRed fluorescence was
observed in cells treated at 0.5% O2 for 48-h hypoxia
compared to normoxic control cells (Fig. 1C). The fluorescent microscopic
images of hypoxic and normoxic cell showed that strong eGFP and
mDsRed fluorescence signal was observed only in hypoxic cells,
confirming the results of flow cytometric analysis (Fig. 1D).
Hypoxia-induced TK/CD/UPRT expression
sensitizes HCT8-HRE cells to 5-FC and GCV
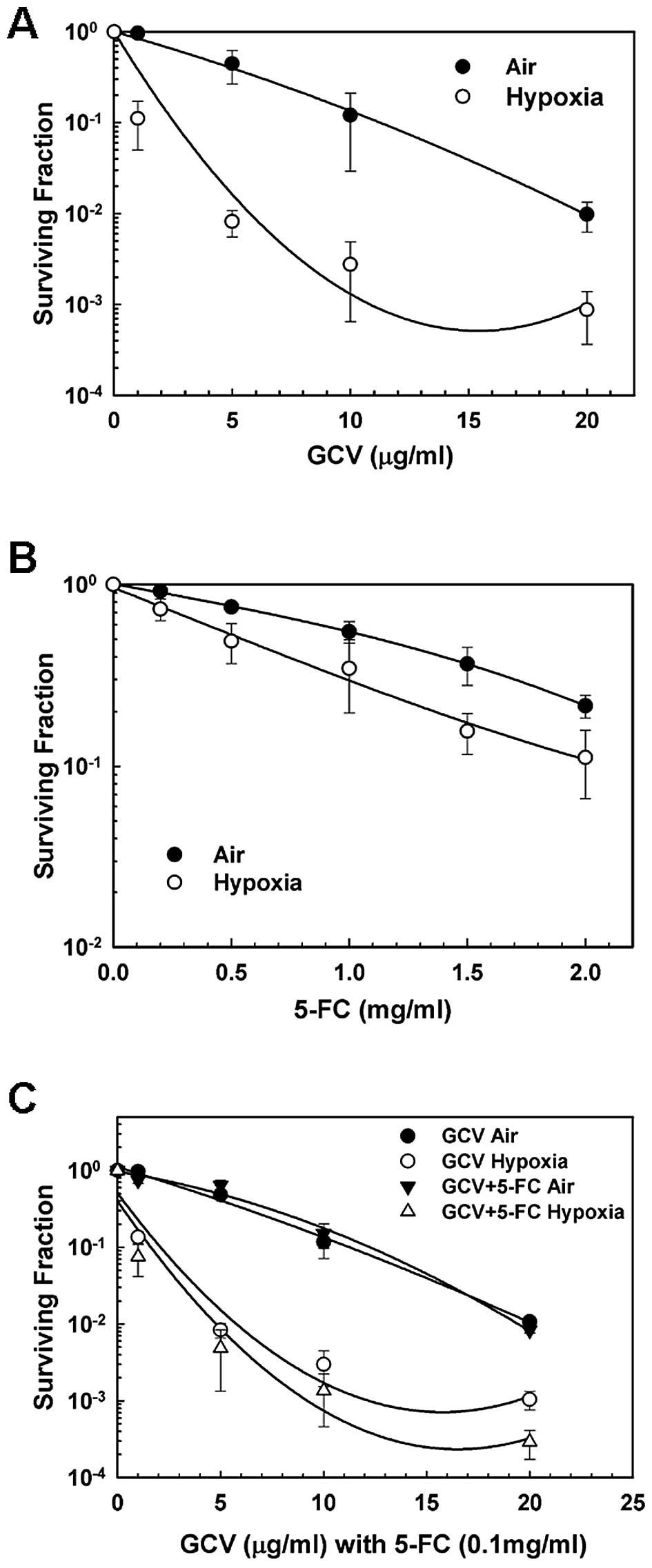
It was clearly shown that the cell clonogenic
survival was significantly inhibited by GCV (Fig. 2A) or 5-FC (Fig. 2B) under hypoxic conditions compared
to normoxic conditions. Whereas HCT8-HRE cells were sensitive to
GCV and 5-FC administered independently, concurrent prodrug
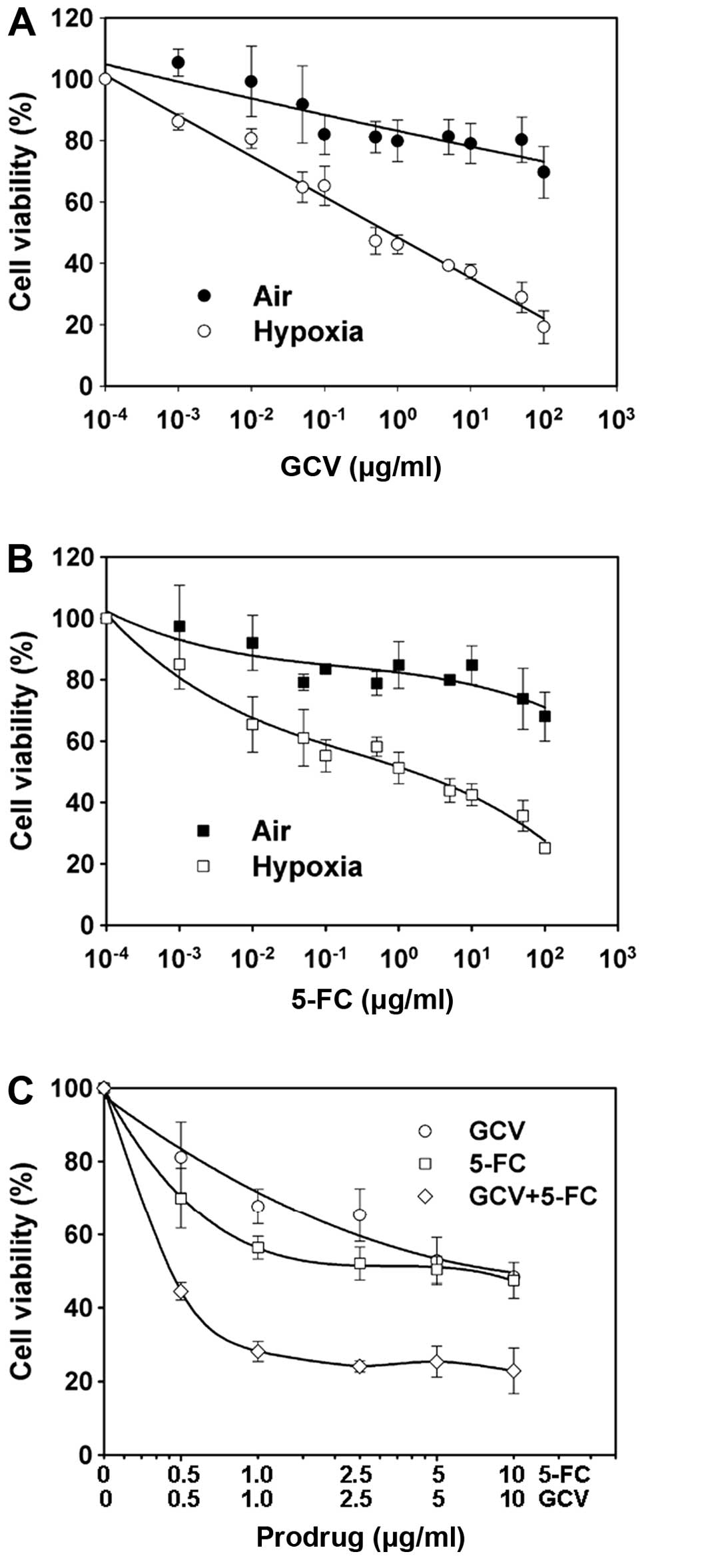
treatment resulted in much greater cytotoxicity (Fig. 2C). Another method, MTT assay, was
used to further assess hypoxia-induced cytotoxicity to GCV and
5-FC. It was found that cell viability under hypoxic conditions was
significantly inhibited by GCV or 5-FC, relative to that under
normoxic conditions (Fig. 3). It
was demonstrated again that concurrent prodrug treatment resulted
in much greater cytotoxicity.
Hypoxia-induced expression of TK/CD/UPRT
radiosensitizes HCT8-HRE tumor cells pre-treated with prodrug 5-FC
and GCV
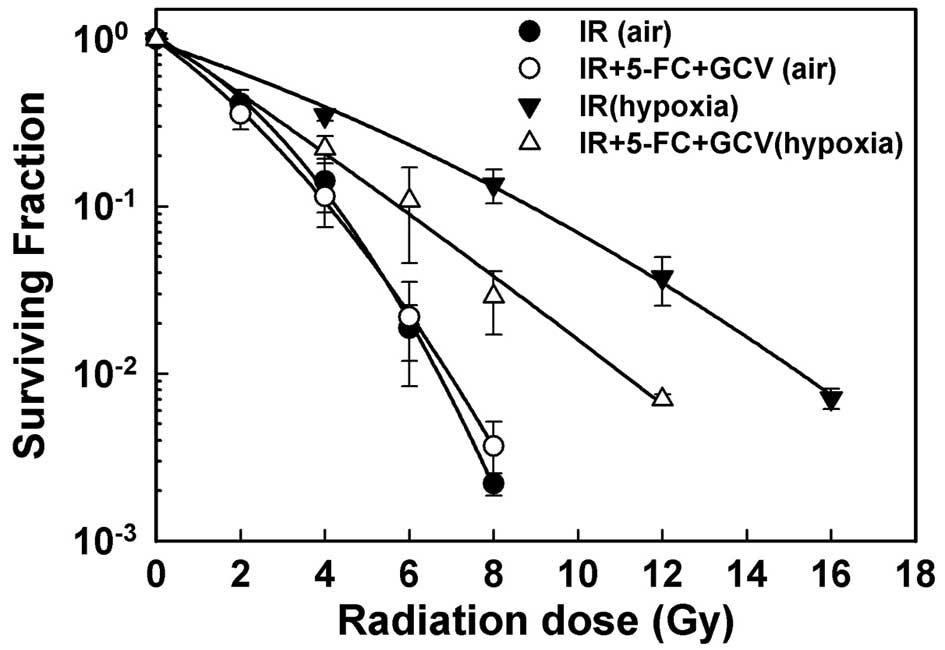
Data on the effect of 48 h GCV and 5-FC treatments
on the radiation response of normoxic and hypoxic HCT-HRE cells are
presented in Fig. 4. Treatments
with GCV plus 5-FC had little radiosensitizing effect in normoxic
HCT-HRE cells. However, drug treatments did enhance the
radiosensitivity of hypoxic HCT-HRE cells.
Hypoxia-induced triple suicide gene
expression in conjunction with GCV and 5-FC treatments exerts
bystander effects
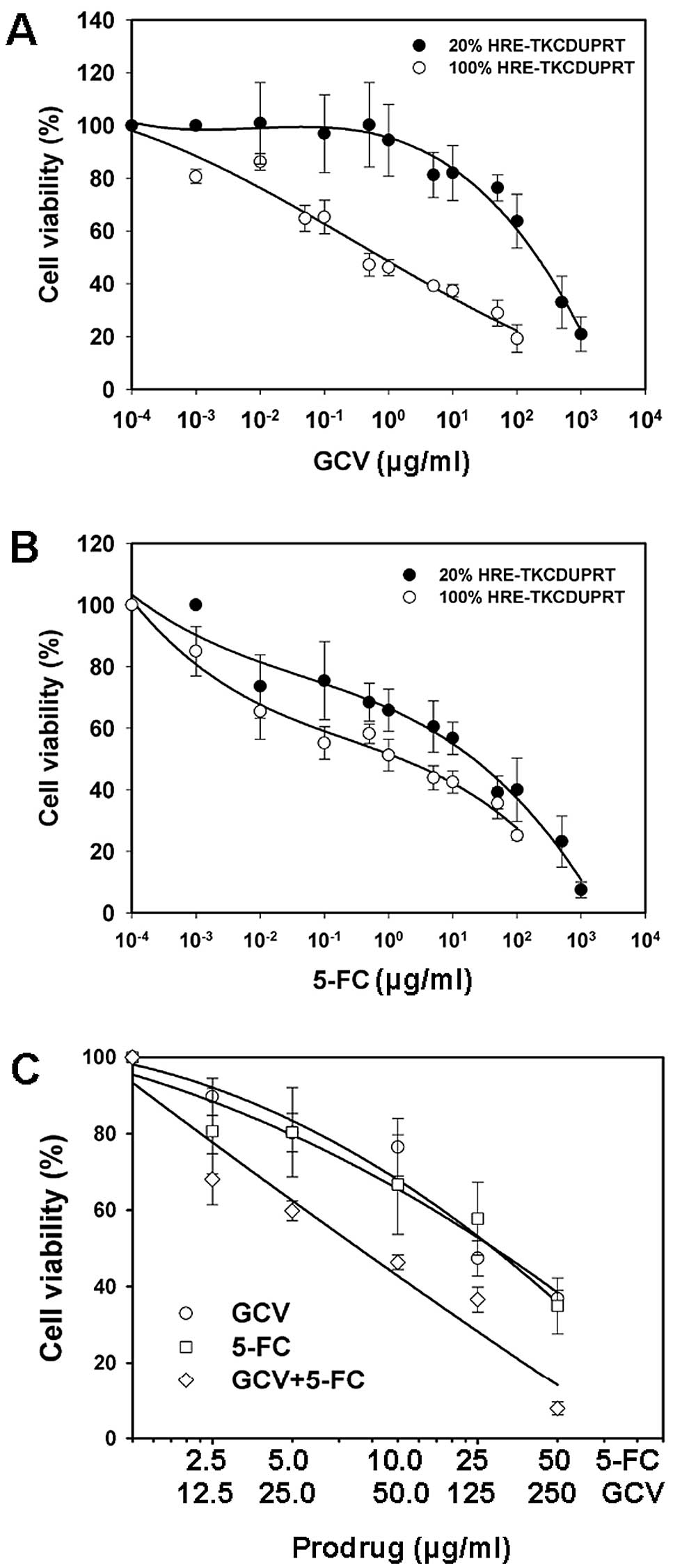
Bystander killing was examined in vitro by
culturing mixture with 20% HCT8-HRE cells and 80% control cells and
exposing the cells to increasing doses of GCV and/or 5-FC under the
hypoxic conditions. As shown in Fig.
5, the cell mixture of 20% HCT8-HRE cells was much less
sensitive to GCV than 100% HCT8-HRE cells under the hypoxic
conditions (Fig. 5A), whereas 20%
HCT8-HRE cell mixtures was slightly less sensitive to 5-FC than
100% HRE cells (Fig. 5B).
Furthermore, combined GCV and 5-FC treatments also led to greater
cytotoxicity in the mixed cell population (Fig. 5C).
Hypoxia-induced transgene expression in
vivo
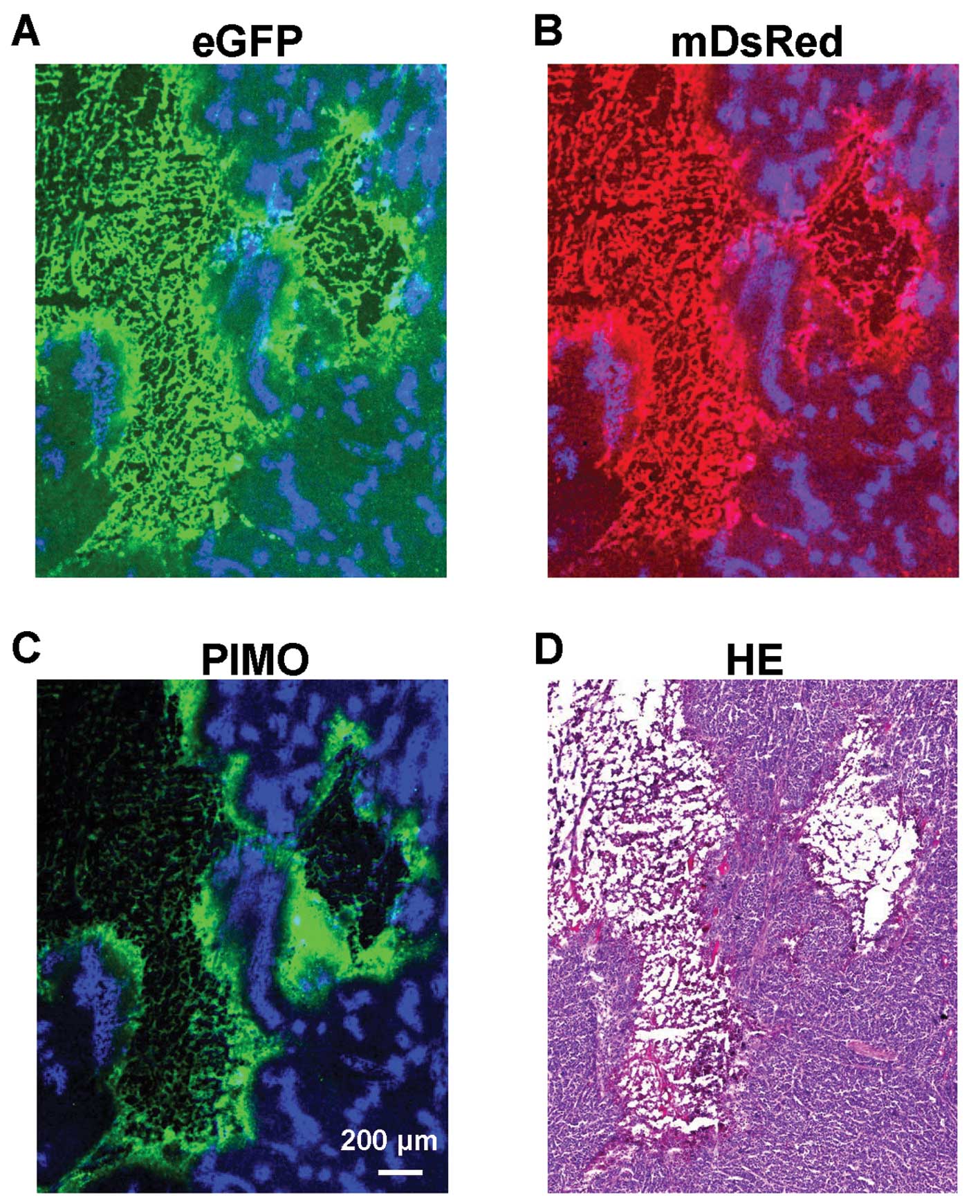
Hypoxia-driven transgene expression in vivo
was validated and compared with distributions of exogenous hypoxia
marker (pimonidazole) and blood perfusion marker (Hoechst 33342).
Fig. 6 provides a detailed
examination and comparison of the different biomarkers in magnified
views of a region from an HCT8-HRE tumor. The merged images of eGFP
and Hoechst (Fig. 6A) or mDsRed and
Hoechst (Fig. 6B) show that eGFP
(green) and mDsRed (red) are located between well perfused area
(blue) and the necrotic region, consistent with hypoxia-induced
expression of the eGFP and mDsRed reporter gene. Fig. 6C shows that the exogenous hypoxic
marker pimonidazole (green) is also accumulated in regions of low
perfusion.
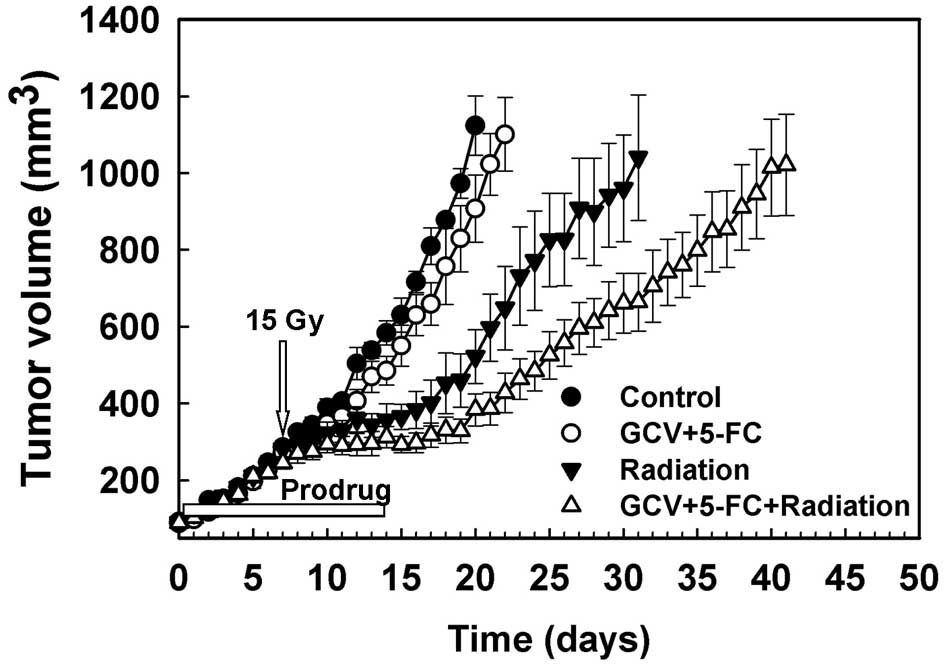
Radiation combined with 5-FC and GCV
treatments delays tumor growth
The response of the HCT8-HRE tumors to prodrug (GCV
and 5-FC) treatment with or without radiation was assessed using
the tumor growth delay assay. As shown in Fig. 7, 5-FC (500 mg/kg) and GCV (30 mg/kg)
treatments for 14 days alone or radiation (15 Gy) alone, slightly
delayed the tumor growth. However, treatments of GCV plus 5-FC
combined with irradiation showed significant tumor growth delay
(P<0.01).
Discussion
The hypoxic microenvironment, an important feature
of human solid tumors but absent in normal tissue, may provide an
opportunity for cancer-specific gene therapy. Studies have
demonstrated the potential of hypoxia/HRE-regulated gene therapy
for cancer with HSV1-TK (10,18),
bacterial CD (7,19,20)
and yeast CD (21,22) employed. To explore the possibility
of the hypoxia-targeted triple suicide gene therapy strategy, HCT-8
cell lines stably transfected with hypoxia-inducible vectors
(HRE-TK/eGFP and HRE-CD/UPRT/mDsRed) were established. Hypoxic
induction of the TK/eGFP and CD/UPRT/mDsRed protein (Fig. 1) led to increased sensitivity to GCV
and 5-FC (Figs. 2 and 3). At the same time, significant
radiosensitizing effects were detected in vitro after 5-FC
and GCV treatments in the hypoxia conditions (Fig. 4). More importantly, administration
of 5-FC and GCV in the tumor-bearing mice in combination with
localized irradiation resulted in significant tumor growth delay,
as compared with prodrug or radiation treatments alone (Fig. 7). These results clearly demonstrated
the potential of hypoxia-targeted radio-gene therapy with the
HRE-TKCDUPRT/GCV+5-FC approach.
One potential advantage of suicide gene therapy
relevant to clinical application is its radiosensitization effect.
Our data clearly showed that the co-expression of hypoxia-regulated
HSV1-TK, CD and UPRT triple genes not only significantly increased
the sensitivity of GCV and 5-FC, but also improved the
radiosensitizing effect of GCV and 5-FC in vitro (Fig. 4) and in vivo (Fig. 7). It has been shown that HSV-1
TK/GCV gene therapy may inhibit the repair of radiation-induced
sublethal DNA damage (23). Others
have suggested that the radiosensitization in CD/5-FC and
CD/UPRT/5-FC approaches is mediated through the inhibition of
thymidylate synthase by 5-FdUMP, resulting in the depletion of
deoxythymidine monophosphate pools and increased DNA strand break,
as well as redistribution of cells to the radiosensitive early S
phase (24). Previously, we
demonstrated that the CDUPRT/5-FC approach has greater
radiosensitization effect than the CD/5-FC system (13). The hypoxia-regulated CD/5-FC
approach has shown significant tumor controlling effects as
combined with irradation in different tumor models (19,22).
Therefore, it is not surprising that significant tumor control
effects were found as the triple suicide gene approach was combined
with radiation.
Another advantage of suicide gene therapy approaches
is the bystander effects. The HSV1-TK/GCV system requires direct
cell-to-cell contacts and active transport of toxic GCV metabolites
to neighboring cells through the gap junctions to exert their
bystander killing effect (25). On
the other hand, in the CD/5-FC system, the converted 5-FU is able
to diffuse across the cell membrane into adjacent cells without
going through the gap junction, resulting in a more powerful
bystander effect (26). Strong
bystander effect has also been shown with the CDUPRT/5-FC approach
(27). In the present study, the
cell mixture of 20% HRE-TKCDUPRT cells was more sensitive to 5-FC
than GCV under the hypoxic conditions (Fig. 5), whereas the sensitivity of the
100% HRE cells to GCV and 5-FC was similar as determined by MTT
assay (Fig. 3). This is partly due
to the low expression of gap junctions in the human colorectal
cells (25). Even so, combined GCV
and 5-FC treatments leads to greater cytotoxicity in mixed cell
populations (Fig. 5C), which
demonstrated the superior bystander killing in the hypoxia-driven
TKCDUPRT approach.
Several factors may impact on the efficacy of
hypoxia-targeted gene therapy. Firstly, the tumors with high
hypoxia fraction, for example large tumors, will be targets for
hypoxia-regulated suicide gene therapy. In the present study,
treatment of 5-FC plus GCV in tumor-bearing mice showed only mild
tumor growth delay (Fig. 7). It may
be due to low hypoxia fraction in the model used (Fig 6). Secondly, HIF activity in tumors
depends on availability of the HIF-1α/2α subunit, the levels of
which increase under hypoxic conditions and through activation of
oncogenes and/or inactivation of tumor suppressor genes (such as
p53). HCT-8 tumor cells bear wild-type p53 gene, which may limit
the hypoxia-induced gene expression in vivo. To investigate
the potential role of hypoxia-targeted triple suicide gene therapy,
tumor models with different hypoxia conditions and gene profile
were used.
The potential limitation of the present study is the
use of stably transfected cell lines and associated tumor models.
As two plasmids carrying hypoxia-regulated TK or CDUPRT were used,
the hypoxia-induced expression/function and cytotoxicity of TK and
CDUPRT were not comparable (Figs. 1
and 2). To translate this strategy
to clinical application, we are generating adenovirus vectors to
deliver hypoxia-regulated TK, CD and UPRT genes into in vivo
tumor models and to test whether triple gene transduction following
GCV and 5-FC treatment will improve the efficacy of radiotherapy.
To overcome the limitations in introducing gene therapy into
clinical use, advanced methods of hypoxia-targeted gene delivery
are also being developed.
In conclusion, our data suggest that the
hypoxia-inducible TK/GCV+CDUPRT/5-FC triple suicide gene therapy
may have the ability to specifically target hypoxic cancer cells
and improve the tumor control in combination with radiotherapy.
This novel model may also serve as a valuable tool to study the
correlations among tumor hypoxia, radiation and hypoxia-targeted
radio-gene therapy.
Acknowledgements
The present study was supported in part by grants
from the National Institutes of Health (P01 CA115675, R01 CA56909,
and R33 CA109772), the National Natural Science Foundation of China
(81001004 and 81272502) and the Shandong Natural Science Foundation
(ZR2010HL027).
References
|
1
|
Wilson WR and Hay MP: Targeting hypoxia in
cancer therapy. Nat Rev Cancer. 11:393–410. 2011. View Article : Google Scholar
|
|
2
|
Nordsmark M, Bentzen SM, Rudat V, et al:
Prognostic value of tumor oxygenation in 397 head and neck tumors
after primary radiation therapy. An international multi-center
study. Radiother Oncol. 77:18–24. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Brizel DM, Scully SP, Harrelson JM, et al:
Tumor oxygenation predicts for the likelihood of distant metastases
in human soft tissue sarcoma. Cancer Res. 56:941–943.
1996.PubMed/NCBI
|
|
4
|
Rampling R, Cruickshank G, Lewis AD,
Fitzsimmons SA and Workman P: Direct measurement of pO2
distribution and bioreductive enzymes in human malignant brain
tumors. Int J Radiat Oncol Biol Phys. 29:427–431. 1994.PubMed/NCBI
|
|
5
|
Keith B, Johnson RS and Simon MC: HIF1α
and HIF2α: sibling rivalry in hypoxic tumour growth and
progression. Nat Rev Cancer. 12:9–22. 2012.
|
|
6
|
Wang GL and Semenza GL: General
involvement of hypoxia-inducible factor 1 in transcriptional
response to hypoxia. Proc Natl Acad Sci USA. 90:4304–4308. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Marignol L, Lawler M, Coffey M and
Hollywood D: Achieving hypoxia-inducible gene expression in tumors.
Cancer Biol Ther. 4:359–364. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Greco O, Marples B, Joiner MC and Scott
SD: How to overcome (and exploit) tumor hypoxia for targeted gene
therapy. J Cell Physiol. 197:312–325. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Marignol L, Foley R, Southgate TD, Coffey
M, Hollywood D and Lawler M: Hypoxia response element-driven
cytosine deaminase/5-fluorocytosine gene therapy system: a highly
effective approach to overcome the dynamics of tumour hypoxia and
enhance the radiosensitivity of prostate cancer cells in
vitro. J Gene Med. 11:169–179. 2009. View Article : Google Scholar
|
|
10
|
Koshikawa N, Takenaga K, Tagawa M and
Sakiyama S: Therapeutic efficacy of the suicide gene driven by the
promoter of vascular endothelial growth factor gene against hypoxic
tumor cells. Cancer Res. 60:2936–2941. 2000.PubMed/NCBI
|
|
11
|
Freytag SO, Stricker H, Pegg J, et al:
Phase I study of replication-competent adenovirus-mediated
double-suicide gene therapy in combination with conventional-dose
three-dimensional conformal radiation therapy for the treatment of
newly diagnosed, intermediate- to high-risk prostate cancer. Cancer
Res. 63:7497–7506. 2003.
|
|
12
|
Rogulski KR, Zhang K, Kolozsvary A, Kim JH
and Freytag SO: Pronounced antitumor effects and tumor
radiosensitization of double suicide gene therapy. Clin Cancer Res.
3:2081–2088. 1997.PubMed/NCBI
|
|
13
|
Xing L, Sun X, Deng X, et al: Expression
of the bifunctional suicide gene CDUPRT increases
radiosensitization and bystander effect of 5-FC in prostate cancer
cells. Radiother Oncol. 92:345–352. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Porosnicu M, Mian A and Barber GN: The
oncolytic effect of recombinant vesicular stomatitis virus is
enhanced by expression of the fusion cytosine deaminase/uracil
phosphoribosyltransferase suicide gene. Cancer Res. 63:8366–8376.
2003.
|
|
15
|
Sun X, Xing L, Deng X, et al: Hypoxia
targeted bifunctional suicide gene expression enhances radiotherapy
in vitro and in vivo. Radiother Oncol. 105:57–63.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xing L, Sun X, Deng X, et al: A triple
suicide gene strategy that improves therapeutic effects and
incorporates multimodality molecular imaging for monitoring gene
functions. Cancer Gene Therapy. 20:358–365. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
He F, Deng X, Wen B, et al: Noninvasive
molecular imaging of hypoxia in human xenografts: comparing
hypoxia-induced gene expression with endogenous and exogenous
hypoxia markers. Cancer Res. 68:8597–8606. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Binley K, Askham Z, Martin L, et al:
Hypoxia-mediated tumour targeting. Gene Ther. 10:540–549. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu J, Harada H, Ogura M, Shibata T and
Hiraoka M: Adenovirus-mediated hypoxia-targeting cytosine deaminase
gene therapy enhances radiotherapy in tumour xenografts. Br J
Cancer. 96:1871–1878. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee CH, Wu CL and Shiau AL:
Hypoxia-induced cytosine deaminase gene expression for cancer
therapy. Hum Gene Ther. 18:27–38. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang D, Ruan H, Hu L, et al: Development
of a hypoxia-inducible cytosine deaminase expression vector for
gene-directed prodrug cancer therapy. Cancer Gene Ther. 12:276–283.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen JK, Hu LJ, Wang D, Lamborn KR and
Deen DF: Cytosine deaminase/5-fluorocytosine exposure induces
bystander and radiosensitization effects in hypoxic glioblastoma
cells in vitro. Int J Radiat Oncol Biol Phys. 67:1538–1547.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Freytag SO, Kim JH, Brown SL, Barton K, Lu
M and Chung M: Gene therapy strategies to improve the effectiveness
of cancer radiotherapy. Expert Opin Biol Ther. 4:1757–1770. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hwang HS, Davis TW, Houghton JA and
Kinsella TJ: Radiosensitivity of thymidylate synthase-deficient
human tumor cells is affected by progression through the G1
restriction point into S-phase: implications for fluoropyrimidine
radiosensitization. Cancer Res. 60:92–100. 2000.
|
|
25
|
McMasters RA, Saylors RL, Jones KE,
Hendrix ME, Moyer MP and Drake RR: Lack of bystander killing in
herpes simplex virus thymidine kinase-transduced colon cell lines
due to deficient connexin43 gap junction formation. Hum Gen Ther.
9:2253–2261. 1998. View Article : Google Scholar
|
|
26
|
Shirakawa T, Gardner TA, Ko SC, et al:
Cytotoxicity of adenoviral-mediated cytosine deaminase plus
5-fluorocytosine gene therapy is superior to thymidine kinase plus
acyclovir in a human renal cell carcinoma model. J Urol.
162:949–954. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Khatri A, Zhang B, Doherty E, et al:
Combination of cytosine deaminase with uracil phosphoribosyl
transferase leads to local and distant bystander effects against
RM1 prostate cancer in mice. J Gene Med. 8:1086–1096. 2006.
View Article : Google Scholar : PubMed/NCBI
|