Introduction
Breast cancer is the most commonly diagnosed cancer
and the second leading cause of cancer-related mortality among
women in the US (1). Breast cancer
is a type of hormone-dependent malignancy, and sustained exposure
to estrogen is known to contribute to the development and
progression of this disease (2).
The biological effects of estrogen are mediated by the estrogen
receptor (ER) through classic genomic and non-genomic actions
(3). In the US, ~75% of breast
cancers express ER (4), and are
classified as ER-positive breast cancer, thus corroborating the use
of selective ER modulators (SERMs) as estrogen antagonists
(5). Tamoxifen, the most commonly
used SERM, has been accepted as the gold standard of endocrine
therapy and has been utilized successfully both in neoadjuvant and
adjuvant settings (6). However, a
substantial proportion of breast cancer patients with localized
disease and nearly all with advanced disease, despite an initial
positive drug response, develop acquired tamoxifen resistance
(7). To date, application of
commercially available drugs to circumvent tamoxifen resistance
remains sparse, and therefore, searching for novel pharmaceutical
agents for re-sensitization to endocrine therapy is urgent.
A771726, the main active metabolite of leflunomide,
also named teriflunomide, has been approved by the FDA for the
treatment of multiple sclerosis (MS) (8). A771726 exerts anti-inflammatory and
immunomodulatory actions by inhibiting the activity of
proliferating B- and T-cells through dihydroorotate dehydrogenase
(DHODH) inhibition-dependent and -independent patterns (9). Although the precise underlying
mechanisms remain elusive, A771726 has high plasma concentrations
(10), a long elimination half-time
(11), high treatment compliance
(12) and multiple molecular
targets (13), implying more
possibilities for broader clinical applications, even beyond the
treatment of MS.
Actually, A771726 has been reported to exert a
potent anticancer effect in treating multiple human neoplasms
(14–17). Notably, A771726 has been used as an
agent with direct cytotoxicity to chemoresistant chronic
lymphocytic leukemia (CLL) (18),
and has also been utilized to enhance chemosensitization (19,20)
and assist targeting therapy (21,22) in
the treatment of other types of tumors. Further investigation on
the application of A771726 in endocrine therapy for breast cancer
has not yet been documented. Primary mechanisms of acquired
resistance to tamoxifen in ER-positive breast cancer include
deregulation of cell cycle and apoptotic machinery accompanied by
altered modulation of various regulators, such as Bcl-2, p21,
activation of receptor tyrosine kinase signaling leading to the
increased activity of Src and MAPK pathways, and alteration of ER
and ER co-regulators (23). In the
present study, using an acquired endocrine-resistant cell model,
MCF-7/LCC9, we investigated, for the first time, to our knowledge,
the anticancer activity and tamoxifen-resistance reversal of
A771726. Furthermore, we performed a comprehensive profiling
analysis of A771726-regulated molecules in MCF-7/LCC9 cells and
validated that A771726 may modulate extensive cellular processes
and multiple signaling pathways that are critical for endocrine
resistance. Here, our study provides new clues for the reversal of
resistance to endocrine therapy and the potential clinical
applications of A771726 in the management of tamoxifen resistance
in breast cancer patients.
Materials and methods
Cell culture and drug treatment
MCF-7 and tamoxifen-resistant MCF-7/LCC9 cells were
kindly provided by Dr Robert Clarke (Georgetown University Medical
Centre, Washington, DC, USA). Cells were routinely cultured in
complete medium with 5% fetal bovine serum (FBS) (Invitrogen,
Carlsbad, CA, USA) at 37°C in 5% CO2. 4-Hydroxytamoxifen
(4-OHT) was purchased from Sigma-Aldrich (St. Louis, MO, USA), and
A771726 was kindly provided by Cinkate Corporation (Oak Park, IL,
USA). For the tamoxifen sensitivity analysis, 2,000 cells/well were
plated in phenol red-free IMEM plus 5% charcoal dextran-treated FBS
(Tissue Culture Biologicals, Los Alamitos, CA, USA) for 24 h and
were then treated with a combination of A771726 and OHT for 7
days.
Sulforhodamine B (SRB) assay
Cells were seeded in 96-well plates (Corning, Acton,
MA, USA) and fixed by trichloroacetic acid (Sinopharm Reagent,
Shanghai, China). Dried cells were stained by SRB (Sigma-Aldrich),
which then was solubilized by Tris-base (Sinopharm Reagent). The
absorbance was read on an automated microplate reader (VERSAmax;
Molecular Devices, Sunnyvale, CA, USA).
Clonogenic assay
Cells were plated at 500 cells/well in 6-well plates
(Corning) and incubated with various concentrations of A771726 for
2 weeks. Then, the colonies were stained with 0.5% crystal violet
and counted.
Flow cytometric assay
For cell apoptosis analysis, after treatment of
A771726 for 48 h, cells were detached and washed with cooled PBS.
Cells were then re-suspended and stained by Annexin V and propidium
iodide (PI) (BD Pharmingen, San Diego, CA, USA). Analysis was
performed on a FACSCalibur analyzer (Becton-Dickinson, San Jose,
CA, USA). With respect to cell cycle analysis, cells were harvested
and fixed by 70% cooled ethanol, and then stained with PI
(Sigma-Aldrich). Analysis was also carried out on a FACSCalibur
analyzer.
Western blot assay
All cells were treated with A771726 for 72 h. The
protein extracts were separated by SDS-PAGE and transferred to NC
membranes (Millipore, Bedford, MA, USA). Src and p-Src proteins
were visualized by a chemiluminescence system (GE Healthcare,
Piscataway, NJ, USA).
Microarray analysis
Briefly, samples were utilized to synthesize
double-stranded complementary DNAs (cDNAs), which were labelled and
hybridized to dual-channel mRNA microarray version 1.0 (CapitalBio
Corp., Beijing, China) following the manufacturer’s instructions.
Data were selected by using a threshold of >2- and <−2-fold
change under false discovery rate (FDR) protection (P<0.05).
Statistical analysis
Statistical significance was evaluated by the
Student’s t-test with GraphPad Prism software 5.0. P-values
<0.05 were considered to indicate statistically significant
results.
Results
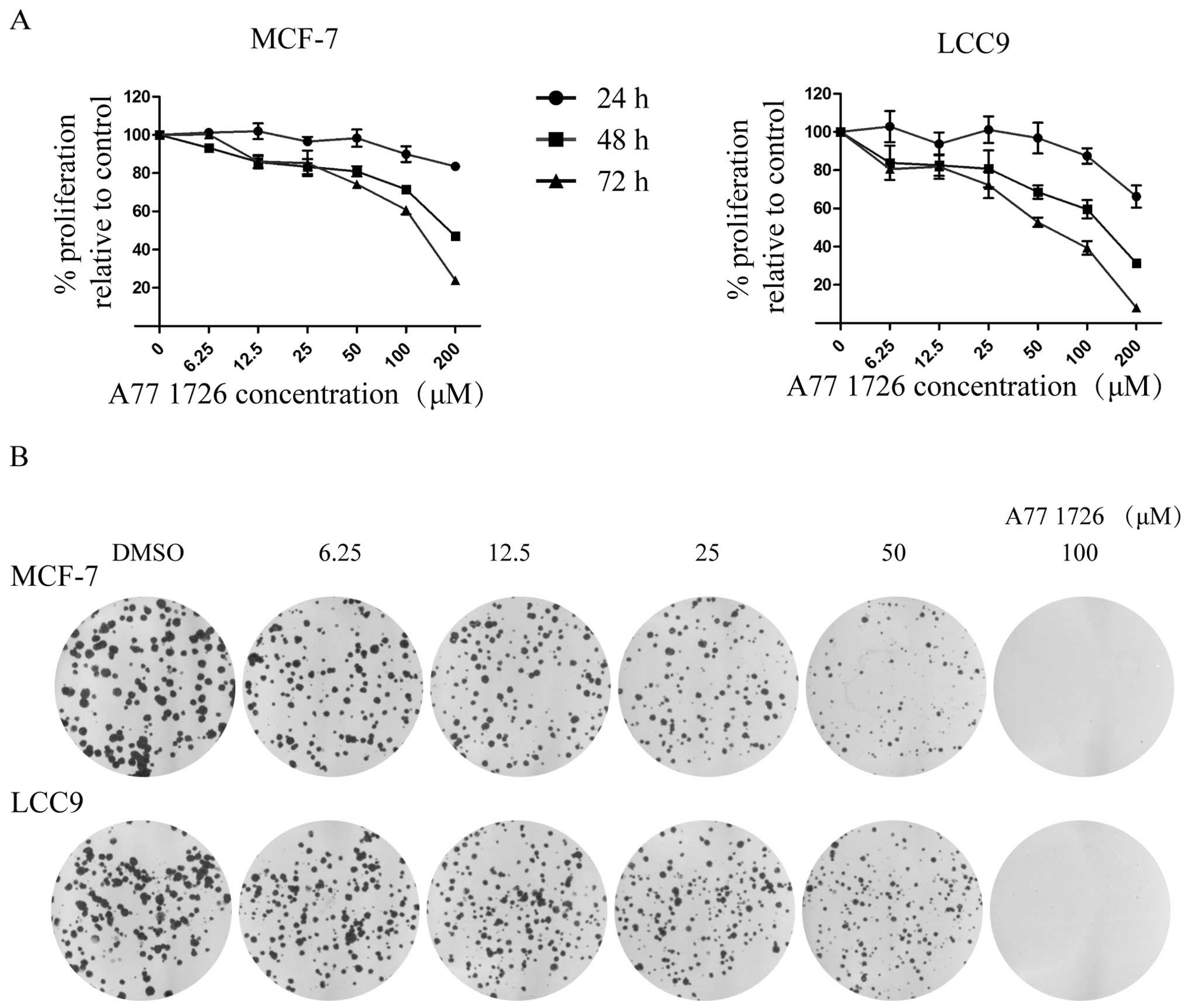
A771726 inhibits the cell proliferation
of endocrine-resistant breast cancer cells
To examine the biological effects on cell
proliferation of tamoxifen-resistant cells and their sensitive
counterparts, we treated the cells with different doses of A771726
and performed an SRB assay (Fig.
1A). Viability of the cells decreased in a dose- and
time-dependent manner. Moreover, tamoxifen-resistant MCF-7/LCC9
cells were more sensitive to inhibition by this agent than the
tamoxifen-sensitive MCF-7 cells; marked early antiproliferative
effects were observed within a 24-h period in the MCF-7/LCC9 cells
but not until 48 h in the MCF-7 cells, respectively. Lower
IC50 values were observed for A771726 in the MCF-7/LCC9
cells (72 h post-treatment: IC50=64.72 μM in MCF-7/LCC9
and 122.79 μM in MCF-7 cells).
A771726 suppresses the colony-forming
ability of endocrine-resistant breast cancer cells
To further demonstrate the antiproliferative effects
of A771726 on the growth of MCF-7/LCC9 cells, a colony formation
assay was carried out. As shown in Fig.
1B, the colony numbers of MCF-7 and MCF-7/LCC9 cells were
significantly decreased in dose-dependent manner, and long-term
treatment of A771726 at the concentration of 100 μM almost
completely diminished the colony formation capability. The results
were consistent with that of the SRB assay further indicating that
A771726 inhibits the in vitro proliferation of MCF-7/LCC9
cells.
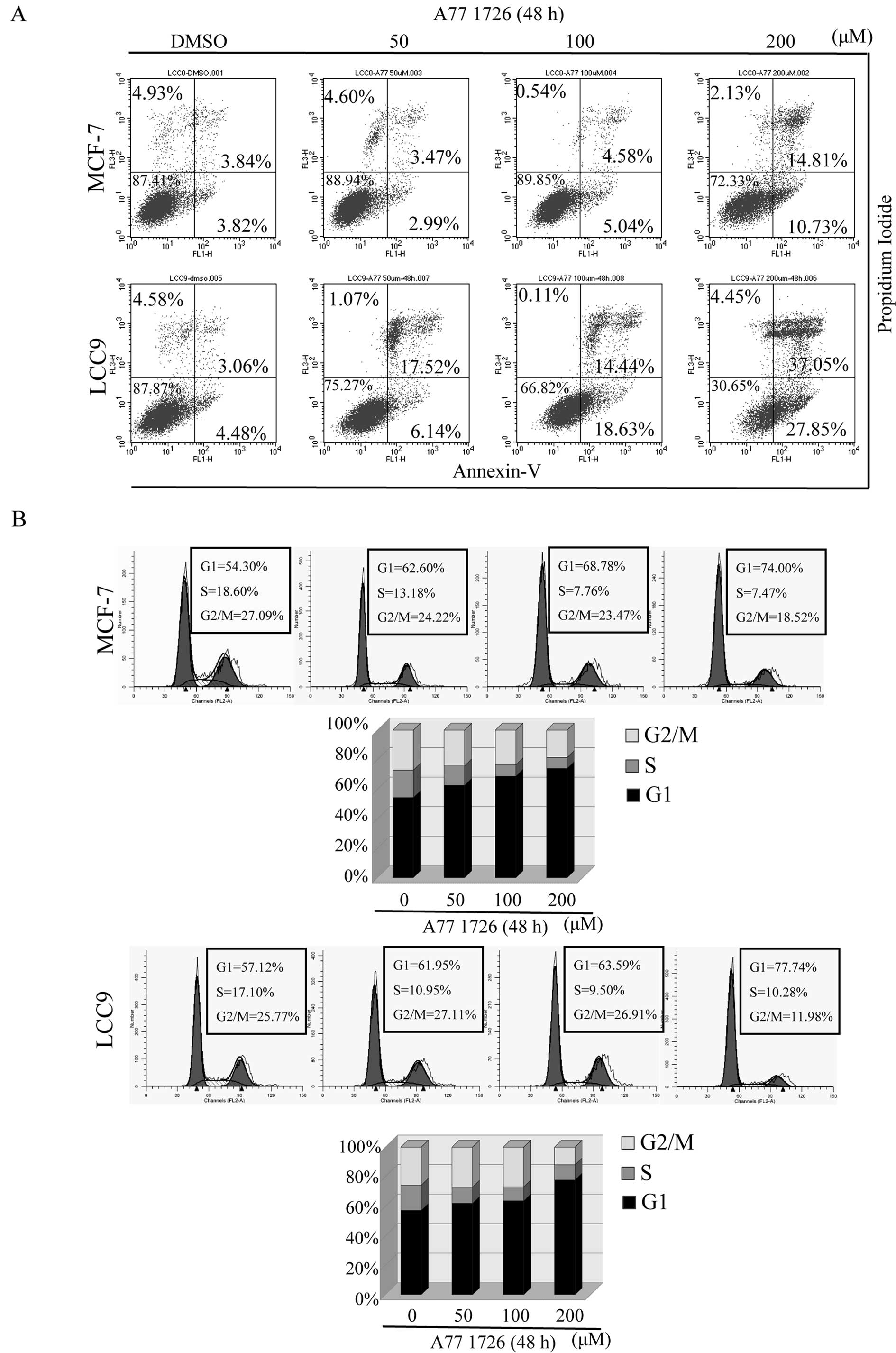
A771726 induces apoptosis in
endocrine-resistant breast cancer cells
We aimed to ascertain whether the cell growth
inhibition induced by A771726 was due to the induction of apoptosis
in tamoxifen-resistant cells. Therefore, we incubated MCF-7/LCC9
and MCF-7 cells with A771726 for 48 h. Our experiments showed that
A771726 induced apoptosis in a dose-dependent manner in the
endocrine-resistant and wild-type cells (Fig. 2A). When exposed to 50 and 100 μM
A771726, few MCF-7 cells were PI+ or Annexin
V+, indicating that low doses of A771726 did not induce
significant apoptosis while the highest concentration of 200 μM
did. However, MCF-7/LCC9 cells were more sensitive to A771726; the
lowest concentration of 50 μM significantly increased the
percentage of Annexin V+/PI+ cells. Following
treatment with higher doses of A771726 at 100 and 200 μM, the
percentage of Annexin V+/PI− cells was
increased from 4.48 to 18.63 and 27.85%, as well as the percentage
of Annexin V+/PI+ cells from 3.06 to 37.05%
at 200 μM, indicating A771726 at various concentrations induced
apoptosis and necrosis in endocrine-resistant cell.
A771726 induces G1 phase arrest in
endocrine-resistant breast cancer cells
In order to determine whether A771726 has a cell
cycle arrest effect, MCF-7/LCC9 and MCF-7 cells treated with
increasing doses of A771726 for 48 h were subjected to flow
cytometric analysis. As shown in Fig.
2B, A771726 caused a dose-dependent accumulation of cells in
the G1 phase fraction, while reducing cell accumulation in the S
and G2/M phases. The G1 phase fraction increased from 57.12 to
61.95%, 63.59 and 77.74% at 50, 100 and 200 μM A771726 in resistant
cells, and from 54.30 to 62.60%, 68.78 and 74.00% in wild-type
cells at the same concentrations of A771726, accompanied by maximal
decreases of 6.82% in the S phase and 13.79% in the G2/M phase in
MCF-7/LCC9 cells and 10.53% in the S phase and 8.59% in the G2/M
phase in MCF-7 cells.
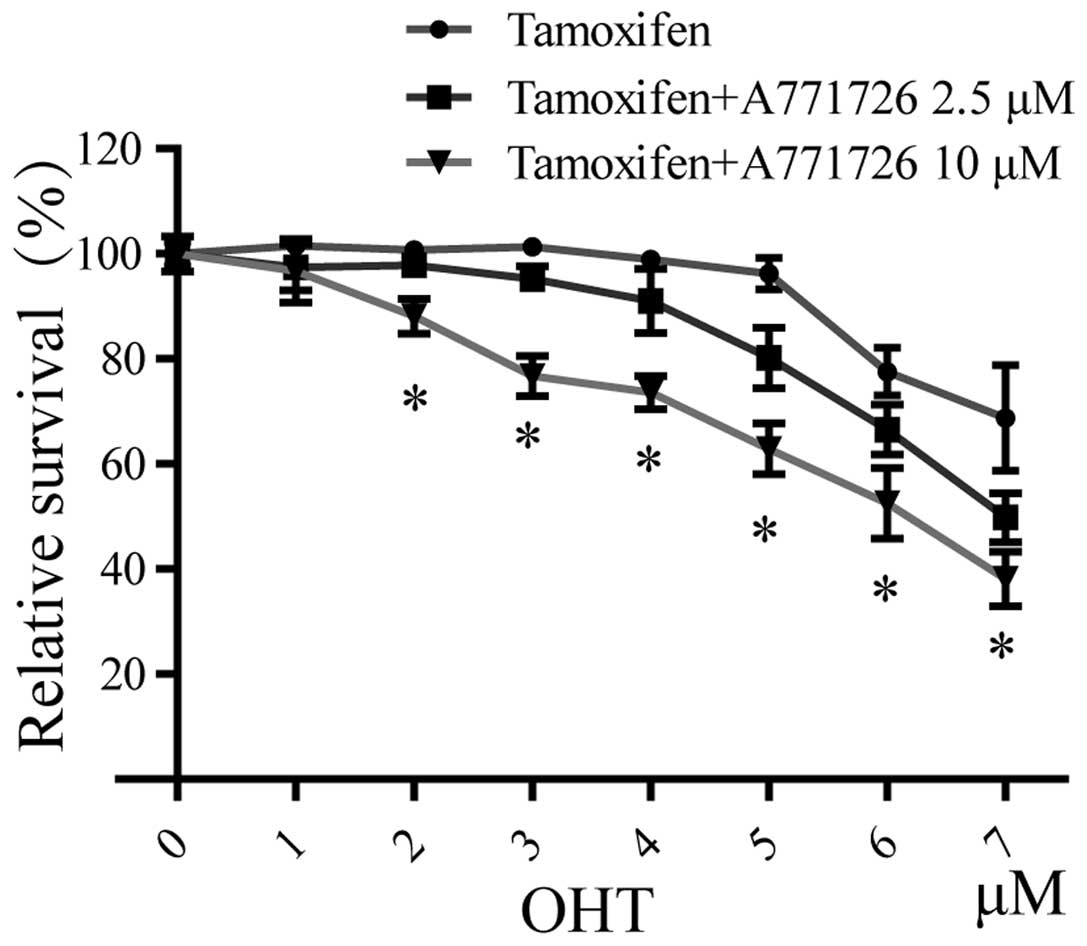
A771726 reverses tamoxifen resistance in
endocrine-resistant breast cancer cells
To further examine the effects of A771726 on
tamoxifen sensitivity, hormone-resistant MCF-7/LCC9 cells were
treated with different doses of A771726 and tamoxifen (alone or in
combination) for 7 days in estrogen-depleted media containing 5%
charcoal-stripped serum (CS). As shown in Fig. 3, cell viability assay recorded over
a range of 4-OHT concentrations from 1 to 7 μM confirmed that these
cells indeed were resistant to OHT, compared with their sensitive
counterparts (data not shown). After receiving A771726 plus OHT,
dose-dependent increases in the sensitivity of MCF-7/LCC9 cells to
OHT were observed. The IC50 value of OHT in the
A771726-treated MCF-7/LCC9 cells was decreased to 7.0 and 5.9 μM at
2.5 and 10 μM, indicating significant recovery of drug
sensitivity.
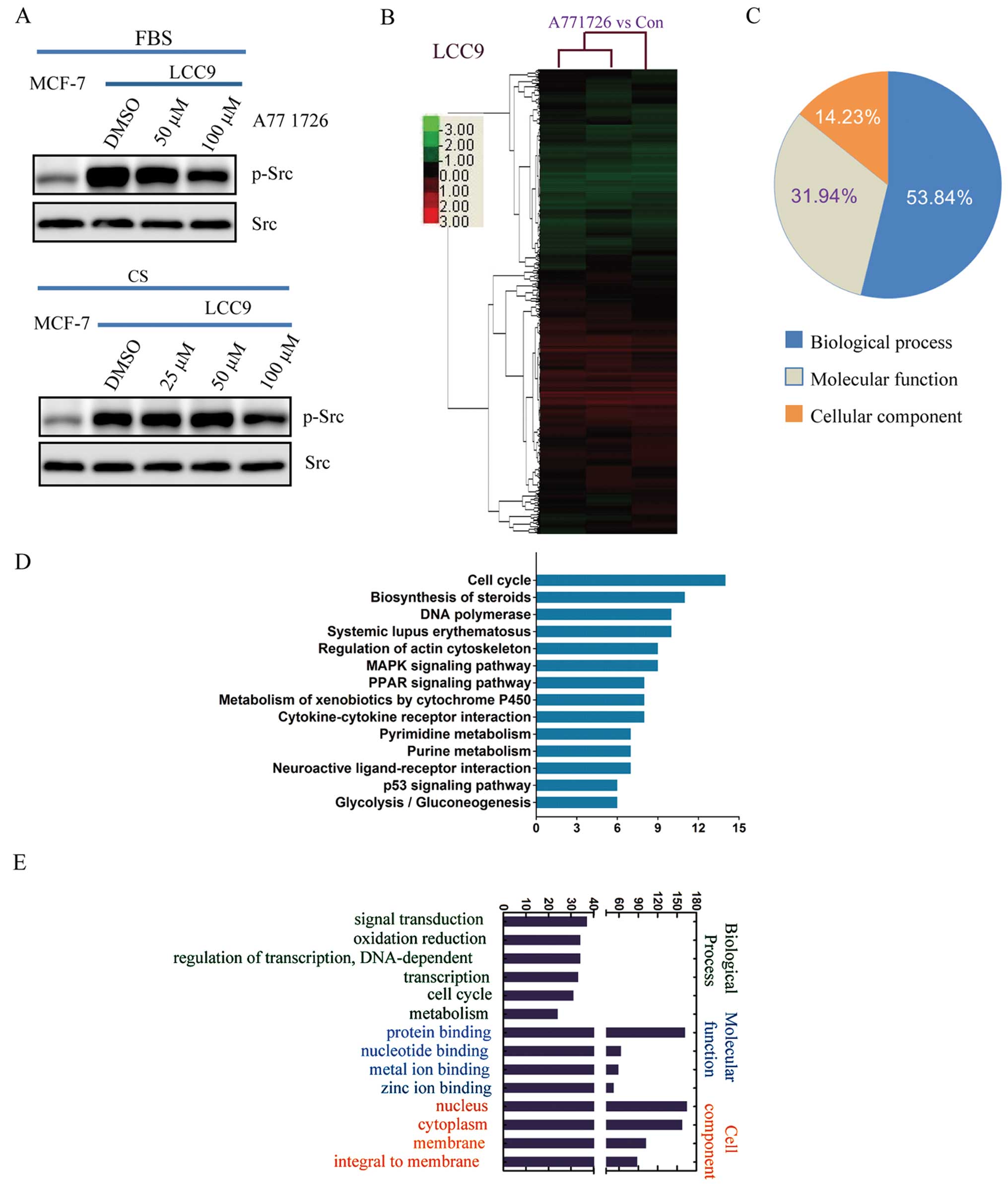
A771726 inhibits Src activation in
endocrine-resistant breast cancer cells
To investigate the mechanisms by which A771726
reverses the drug resistance of MCF-7/LCC9 cells, we analyzed the
state of Src phosphorylation, one key driver of tamoxifen
resistance (Fig. 4A). Although no
change in total Src expression was detected between MCF-7 and
MCF-7/LCC9 cells, our results revealed that the phosphorylation of
Src (Tyr418) was increased in the resistant cells, both in complete
medium with 5% FBS and in phenol red-free medium with 5% CS, and
interestingly at a higher level in FBS. Following A771726
treatment, the Src phosphorylation was significantly suppressed at
100 μM A771726 at 72 h.
A771726 regulates global gene expression
and multiple signaling pathways in endocrine-resistant breast
cancer cells
To systematically gain further insight into the
functional effects of A771726 in tamoxifen resistance reversal and
other biological functions, we performed large-scale mRNA
expression profiling following treatment of 100 μM A771726 for 72 h
in MCF-7/LCC9 cells. As shown in Fig.
4B, clustering results of all significantly regulated probes
were demonstrated. Respectively, we identified 410 upregulated
probes and 285 downregulated probes (Table I). To investigate the functional
roles of the A771726-regulated genes, pathway analysis was
performed, and genes modulated by A771726 in the MCF-7/LCC9 cells
were significantly enriched in multiple KEGG pathways, represented
by cell cycle, apoptosis, MAPK, metabolism and P53 signaling
pathways (Fig. 4D). Gene ontology
(GO) analysis showed that 53.84% of all differentially expressed
genes were associated with biological process, 31.94% with
molecular function and 14.23% with cellular component (Fig. 4C). The gene classes most
significantly affected by A771726 were signal transduction and
oxidation reduction in biological process, protein and nucleotide
binding in molecular function and nucleus and cytoplasm in cell
component (Fig. 4E).
 | Table IDifferentially regulated genes in
cell cycle, apoptosis and MAPK signaling pathways following A771726
treatment in MCF-7/LCC9 cells. |
Table I
Differentially regulated genes in
cell cycle, apoptosis and MAPK signaling pathways following A771726
treatment in MCF-7/LCC9 cells.
| Signaling pathway
(oligo ID) | Gene symbol | Gene name | Fold-change
(LCC9-A771726 vs. LCC9-DMSO) |
|---|
| Cell cycle |
| H200008365 | CDKN1A | Cyclin-dependent
kinase inhibitor 1 (p21) | 3.62 |
| H300012891 | CHEK1 |
Serine/threonine-protein kinase Chk1 | 0.19 |
| H300020974 | CCNE2 | G1/S-specific
cyclin-E2 | 0.41 |
| H200015433 | E2F2 | Transcription
factor E2F2 | 0.42 |
| H200006619 | MAD2L1 | Mitotic spindle
assembly checkpoint protein MAD2A | 0.44 |
| H200007139 | CCNA2 | Cyclin-A2 | 0.46 |
| Apoptosis |
|
opHsV0400006063 | CAPN2 | Calpain-2 catalytic
subunit precursor | 4.66 |
| H300022026 | BAX | BAX protein | 2.00 |
| H300008499 | IRF1 | Interferon
regulatory factor 1 | 1.63 |
| H300005750 | HSPA5 | 78 kDa
glucose-regulated protein precursor (GRP 78) | 0.43 |
| H300019718 | BIRC5 | Baculoviral IAP
repeat-containing protein 5 (survivin) | 0.49 |
| H200006652 | BCL2 | Apoptosis regulator
Bcl-2 | 0.6011 |
| MAPK |
| H300008649 | CACNG1 | Voltage-dependent
calcium channel γ-1 subunit | 4.85 |
| H200000600 | DUSP4 | Dual specificity
protein phosphatase 4 | 3.31 |
| H200013671 | PLA2G3 | Group 3 secretory
phospholipase A2 precursor | 2.88 |
| H300000118 | DUSP3 | Dual specificity
protein phosphatase 3 | 2.14 |
| H300019880 | ATF2 | Cyclic
AMP-dependent transcription factor ATF-2 | 0.44 |
| H300016729 | MAP2K6 | Dual specificity
mitogen-activated protein kinase kinase 6 | 0.44 |
Discussion
In the present study, we provide evidence that
A771726 may have direct chemopreventive potential and resistance
reversal when concurrently used in endocrine-resistant cells. We
found that this anti-inflammatory agent is capable of suppressing
the growth of MCF-7/LCC9 cells in vitro by inducing cell
apoptosis and G1 phase arrest. Moreover, by analyzing
A771726-regulated gene expression profiles in
anti-estrogen-resistant cells, we demonstrated the multiple
biological functions and signaling pathways that were involved.
Although recently approved by the FDA for use in the
relapsing forms of MS in the US, A771726 has also been investigated
as a potential anticancer agent in a handful of human tumors
(14–17,21).
However, the lethal effect of A771726 in breast cancer has not been
thoroughly investigated. In the present study, we report the potent
cytotoxicity and attenuation of colony-forming activity of A771726
in sensitive ER-positive breast cancer cells (Fig. 1A and B). Notably, consistent with
the direct anticancer effect in drug-resistance CLL (18), we also showed that A771726 led to
cell death in endocrine-resistant cells (Fig. 1A and B). Moreover, we discovered
that the tamoxifen-resistant MCF-7/LCC9 cells were more sensitive
to A771726-mediated killing than the parental MCF-7 cells (Fig. 1A), indicating that
resistance-related signals may be responsible for the differential
response to A771726. Transcriptional repression or somatic deletion
of CDKN1A (encoding p21), resulting in the alleviation of the
inhibitory effect on CDK activity, contributes to decreased
anti-estrogen sensitivity in breast cancer (24,25).
Although no change in Myc, a suppressor of p21 (24), was observed (data not shown),
significant upregulation of p21 mRNA (Table I) was determined, leading to the
inability of growth maintenance and retrieval of tamoxifen
sensitivity.
Tamoxifen has the ability to elicit cell apoptosis
via an ER-dependent and -independent manner, disturbing the balance
between pro- and anti-survival functions (26). However, when endocrine resistance
develops, cancer cells also lose the response to tamoxifen-induced
apoptosis (27). Accumulating
evidence suggests that the increased expression of anti-apoptotic
Bcl-2 family members, such as Bcl-2 and Bcl-xL, and functional
defect of pro-apoptotic members, such as BAK and BIK, contribute to
the emergence of attenuated responses to tamoxifen (23). In the present study, A771726
treatment in resistant cells induced downregulation of
anti-apoptotic Bcl-2 by 40% and upregulation of the pro-apoptotic
Bax (Table I), leading to an
increase in the BAX:Bcl-2 ratio, shifting the balance towards cell
death (28) and restoring the
tamoxifen-induced apoptosis in MCF-7/LCC9 cells (29). In addition, compared with the
sensitive controls, MCF-7/LCC9 cells expressed lower IRF1 (30), whose enhanced expression by A771726
(by ~60%) may mediate cell death by inversing the balance of Bcl-2
family members and downregulating the inhibitor of apoptosis,
survivin (Table I) (31), thus, determining the cell fate
decision to again undergo drug apoptosis. Interestingly, we also
revealed that autophagy may be involved in A771726-induced
sensitivity restoration by inhibiting expression of GRP78 (Table I), which could activate
mTOR-regulated dependent pro-survival autophagy with increased
LC3-II and decreased p62 in induction of anti-estrogen resistance
(32). Overall, our results showed
that A771726 could recover the deregulated apoptosis and autophagy
in tamoxifen-resistant cells via modulating multiple signals
involved in the determination of cell fate.
In addition to the disruption of cell death
pathways, cell cycle perturbation by A771726, consistent with data
in the treatment of multiple myeloma cells (19), could also be observed in
endocrine-resistant cells. Cyclin E2 is induced in the late G1
phase and activates cyclin-dependent kinase CDK2, leading to
inactivation of pRB and activation of E2F transcription factors to
promote entry into S phase (33).
However, in the development of impaired tamoxifen responsiveness,
aberrant regulation of several cell cycle regulators, such as
cyclin E2 and E2F2, also anti-estrogen targets, was found to be
associated with the inability of induction of G1 phase-specific
cell cycle arrest and the consequent reduction in cancer growth
(34,35). Interestingly, our genome-wide
analysis in MCF-7/LCC9 cells showed that A771726 significantly
decreased the expression of cyclin E2 and E2F2 (Table I) and CDK2 by 25%, accompanied by
the upregulation of the antagonist of CDK inhibitor, p21 (33), contributing to the delay of cell
cycle progression from the G1 to the S phase (Fig. 2B) and enhancement of
tamoxifen-induced growth inhibition (Fig. 3). Moreover, downregulation of GRP78
(Table I), also a key driver of
tamoxifen resistance (32), may be
involved in G1 arrest (36).
Prior experiments have demonstrated that the
resistant cell line MCF-7/LCC9 has an elevated level of Src
phosphorylation (37), which was
also observed in the present study, regardless of whether the
medium contained estrogen or not (Fig.
4A). Pharmacological inhibition of Src, like that in A771726
treatment in resistant cells, may be effective in preventing the
emergence of tamoxifen resistance (38). For the first time, we demonstrated
the global pattern of cellular functions and signaling pathways of
A771726 targets using GO and pathway analysis. As for pathway
mapping, we revealed that multiple oncogenic signaling pathways,
cell cycle, apoptosis, MAPK, metabolism and p53 pathway, were
regulated by A771726 (Fig. 4D),
which have been reported to contribute to acquisition of tamoxifen
resistance (23,39). Although precise underlying
regulation patterns of the biological processes have not yet been
uncovered, A771726 modulated signal transduction, oxidation
reduction, transcription, cell cycle and metabolism (Fig. 4E), consistent with that in the
pathway analysis. Moreover, the A771726-related gene products were
mainly located in the nuclei, indicating its involvement in the
regulation of transcription.
In summary, to the best of our knowledge for the
first time, we demonstrated the mechanistic evidence for the potent
anticancer effect and tamoxifen resistance reversal by A771726 in
endocrine-resistant MCF-7/LCC9 cells. Our findings demonstrated
that A771726 mediated these effects through modulation of extensive
related signaling pathways and cellular processes. The data provide
support for the potential clinical applications of A771726 for
ER-positive breast cancer patients with poor response to endocrine
therapy.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (nos. 81202549, 81202088,
81302147 and 81302145), the National Science and Technology Major
Project of the Ministry of Science and Technology of China (no.
2012ZX09301001-007 to M.G.), the Leading Academic Discipline
Project of Shanghai Municipal Education Commission (J50208), the
Cancer Foundation of China (0901), the Science and Technology
Commission of Shanghai Municipality (09411961400), and the Shanghai
Charity Cancer Research Centre.
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar
|
|
2
|
Chlebowski RT, Manson JE, Anderson GL, et
al: Estrogen plus progestin and breast cancer incidence and
mortality in the Women’s Health Initiative Observational Study. J
Natl Cancer Inst. 105:526–535. 2013.
|
|
3
|
McDonnell DP and Norris JD: Connections
and regulation of the human estrogen receptor. Science.
296:1642–1644. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Brouckaert O, Paridaens R, Floris G, Rakha
E, Osborne K and Neven P: A critical review why assessment of
steroid hormone receptors in breast cancer should be quantitative.
Ann Oncol. 24:47–53. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pinkerton JV and Thomas S: Use of SERMs
for treatment in postmenopausal women. J Steroid Biochem Mol Biol.
142C:142–154. 2014. View Article : Google Scholar
|
|
6
|
Goncalves R, Ma C, Luo J, Suman V and
Ellis MJ: Use of neoadjuvant data to design adjuvant endocrine
therapy trials for breast cancer. Nat Rev Clin Oncol. 9:223–229.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Early Breast Cancer Trialists’
Collaborative Group (EBCTCG). Effects of chemotherapy and hormonal
therapy for early breast cancer on recurrence and 15-year survival:
an overview of the randomised trials. Lancet. 365:1687–1717.
2005.PubMed/NCBI
|
|
8
|
Ali R, Nicholas RS and Muraro PA: Drugs in
development for relapsing multiple sclerosis. Drugs. 73:625–650.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Oh J and O’Connor PW: Teriflunomide.
Neurol Clin Pract. 3:254–260. 2013. View Article : Google Scholar
|
|
10
|
Tallantyre E, Evangelou N and
Constantinescu CS: Spotlight on teriflunomide. Int MS J. 15:62–68.
2008.
|
|
11
|
Wiese MD, Rowland A, Polasek TM, Sorich MJ
and O’Doherty C: Pharmacokinetic evaluation of teriflunomide for
the treatment of multiple sclerosis. Expert Opin Drug Metab
Toxicol. 9:1025–1035. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Oh J and O’Connor PW: Safety,
tolerability, and efficacy of oral therapies for
relapsing-remitting multiple sclerosis. CNS Drugs. 27:591–609.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Oh J and O’Connor PW: An update of
teriflunomide for treatment of multiple sclerosis. Ther Clin Risk
Manag. 9:177–190. 2013.PubMed/NCBI
|
|
14
|
Zhu S, Yan X, Xiang Z, Ding HF and Cui H:
Leflunomide reduces proliferation and induces apoptosis in
neuroblastoma cells in vitro and in vivo. PLoS One. 8:e715552013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hail N Jr, Chen P, Kepa JJ and Bushman LR:
Evidence supporting a role for dihydroorotate dehydrogenase,
bioenergetics, and p53 in selective teriflunomide-induced apoptosis
in transformed versus normal human keratinocytes. Apoptosis.
17:258–268. 2012. View Article : Google Scholar
|
|
16
|
Ringshausen I, Oelsner M, Bogner C,
Peschel C and Decker T: The immunomodulatory drug Leflunomide
inhibits cell cycle progression of B-CLL cells. Leukemia.
22:635–638. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hail N Jr, Chen P and Bushman LR:
Teriflunomide (leflunomide) promotes cytostatic, antioxidant, and
apoptotic effects in transformed prostate epithelial cells:
evidence supporting a role for teriflunomide in prostate cancer
chemoprevention. Neoplasia. 12:464–475. 2010.
|
|
18
|
Dietrich S, Krämer OH, Hahn E, et al:
Leflunomide induces apoptosis in fludarabine-resistant and
clinically refractory CLL cells. Clin Cancer Res. 18:417–431. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Baumann P, Mandl-Weber S, Völkl A, et al:
Dihydroorotate dehydrogenase inhibitor A771726 (leflunomide)
induces apoptosis and diminishes proliferation of multiple myeloma
cells. Mol Cancer Ther. 8:366–375. 2009. View Article : Google Scholar
|
|
20
|
Uckun FM: Rationally designed anti-mitotic
agents with pro-apoptotic activity. Curr Pharm Des. 7:1627–1639.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
White RM, Cech J, Ratanasirintrawoot S, et
al: DHODH modulates transcriptional elongation in the neural crest
and melanoma. Nature. 471:518–522. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Somnay Y, Chen H and Kunnimalaiyaan M:
Synergistic effect of pasireotide and teriflunomide in carcinoids
in vitro. Neuroendocrinology. 97:183–192. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Musgrove EA and Sutherland RL: Biological
determinants of endocrine resistance in breast cancer. Nat Rev
Cancer. 9:631–643. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mukherjee S and Conrad SE: c-Myc
suppresses p21WAF1/CIP1 expression during estrogen
signaling and antiestrogen resistance in human breast cancer cells.
J Biol Chem. 280:17617–17625. 2005.PubMed/NCBI
|
|
25
|
Abukhdeir AM, Vitolo MI, Argani P, et al:
Tamoxifen-stimulated growth of breast cancer due to p21 loss. Proc
Natl Acad Sci USA. 105:288–293. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Riggins RB, Bouton AH, Liu MC and Clarke
R: Antiestrogens, aromatase inhibitors, and apoptosis in breast
cancer. Vitam Horm. 71:201–237. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Razandi M, Pedram A, Jordan VC, Fuqua S
and Levin ER: Tamoxifen regulates cell fate through mitochondrial
estrogen receptor beta in breast cancer. Oncogene. 32:3274–3285.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Martin LA and Dowsett M: BCL-2: a new
therapeutic target in estrogen receptor-positive breast cancer?
Cancer Cell. 24:7–9. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nehra R, Riggins RB, Shajahan AN, Zwart A,
Crawford AC and Clarke R: BCL2 and CASP8 regulation by NF-κB
differentially affect mitochondrial function and cell fate in
antiestrogen-sensitive and -resistant breast cancer cells. FASEB J.
24:2040–2055. 2010.
|
|
30
|
Ning Y, Riggins RB, Mulla JE, Chung H,
Zwart A and Clarke R: IFNγ restores breast cancer sensitivity to
fulvestrant by regulating STAT1, IFN regulatory factor 1, NF-κB,
BCL2 family members, and signaling to caspase-dependent apoptosis.
Mol Cancer Ther. 9:1274–1285. 2010.
|
|
31
|
Schwartz JL, Shajahan AN and Clarke R: The
role of interferon regulatory factor-1 (IRF1) in overcoming
antiestrogen resistance in the treatment of breast cancer. Int J
Breast Cancer. 2011:9121022011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cook KL, Shajahan AN, Wärri A, Jin L,
Hilakivi-Clarke LA and Clarke R: Glucose-regulated protein 78
controls cross-talk between apoptosis and autophagy to determine
antiestrogen responsiveness. Cancer Res. 72:3337–3349. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Geng Y and Sicinski P: Differences in
regulation and function of E-cyclins in human cancer cells. Cell
Cycle. 12:11652013. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Caldon CE, Sergio CM, Kang J, et al:
Cyclin E2 overexpression is associated with endocrine resistance
but not insensitivity to CDK2 inhibition in human breast cancer
cells. Mol Cancer Ther. 11:1488–1499. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Miller TW, Balko JM, Fox EM, et al:
ERα-dependent E2F transcription can mediate resistance to estrogen
deprivation in human breast cancer. Cancer Discov. 1:338–351.
2011.
|
|
36
|
Lin JA, Fang SU, Su CL, et al: Silencing
glucose-regulated protein 78 induced renal cell carcinoma cell line
G1 cell-cycle arrest and resistance to conventional chemotherapy.
Urol Oncol. 32:29.e1–29.e11. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jiang M, Huang O, Zhang X, et al: Curcumin
induces cell death and restores tamoxifen sensitivity in the
antiestrogen-resistant breast cancer cell lines MCF-7/LCC2 and
MCF-7/LCC9. Molecules. 18:701–720. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hiscox S, Jordan NJ, Smith C, et al: Dual
targeting of Src and ER prevents acquired antihormone resistance in
breast cancer cells. Breast Cancer Res Treat. 115:57–67. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Martinez-Outschoorn UE, Goldberg A, Lin Z,
et al: Anti-estrogen resistance in breast cancer is induced by the
tumor microenvironment and can be overcome by inhibiting
mitochondrial function in epithelial cancer cells. Cancer Biol
Ther. 12:924–938. 2011. View Article : Google Scholar
|