Introduction
Osteosarcoma (OS) is a primary malignant bone tumor
mostly occurring in adolescents and young adults with a second peak
at middle age (1). OS patients are
generally treated with high doses of neoadjuvant chemotherapy to
prevent the outgrowth of micrometastases (2). However, 45% of OS patients develop
distant metastases, leading to the death of OS patients (3). OS is also a genetic disease, and
single or multiple mutations in genes related to growth and
metastasis form the molecular genetic basis of malignant
transformation and tumor progression (4). Thus, identification of molecular
targets that can be exploited in the clinic to treat metastatic
disease is desperately needed.
Tumor necrosis factor-α-inducible protein-1
(TNFAIP1) is a protein which can be induced by tumor necrosis
factor α (TNFα) and interleukin-6 (IL-6), and may play roles in DNA
synthesis, cell apoptosis and human diseases (5). TNFAIP1 has been identified to be
highly expressed in Alzheimer’s disease brains (6) and hepatitis B virus (7), and is associated with diabetic
nephropathy (8). Moreover,
overexpression of TNFAIP1 was found to correlate with histological
grade and a poor prognosis in breast cancer (9), and its allelic partner TNFAIP2 was
found to be aberrantly expressed in malignant cells of classical
Hodgkin lymphoma and primary mediastinal large B-cell lymphoma
(10), and contributes to the rapid
progression of cervical cancer (11), indicating TNFAIP may serve as a
useful new marker of cancer.
TNFAIP1 expression was found to be downregulated in
lipopolysaccharide (LPS)-induced liver injury (12). Yet, few reports have demonstrated
the expression and function of TNFAIP1 in OS tissues. More
importantly, TNFRSF11A gene encoding receptor activator of nuclear
factor-κB (NF-κB) has been confirmed to be responsible for the
progression of OS (13), and
activation of NF-κB participates in the anti-apoptotic effect in
TNFα-treated Ewing sarcoma cells (14). In the present study, the expression
of TNFAIP1 was examined, and its function and molecular mechanisms
were evaluated in human OS. We found that knockdown of TNFAIP1
inhibited the growth and invasion, and induced apoptosis in OS
cells through inhibition of the NF-κB pathway.
Materials and methods
Materials
The OS cell lines (MG-63 and U-2 OS) used for
experiments were obtained from the Institute of Biochemistry and
Cell Biology (Shanghai, China). Lentiviral-mediated TNFAIP1 siRNA
(siTNFAIP1), the negative control (NC) vector, and virion-packaging
elements were purchased from GeneChem (Shanghai, China). Human OS
tissues and the corresponding adjacent non-cancerous tissues
(ANCTs) were collected from the Department of Orthopedic Surgery,
Changzheng Hospital. The OS tissue microarray was constructed by
Shanghai Outdo Biotech Co. Ltd. (Shanghai, China). All the
antibodies were purchased from Cell Signaling Technology (Boston,
MA, USA). TNFAIP1 primer was synthesized by ABI (Framingham, MA,
USA).
Drugs and reagents
Dulbecco’s modified Eagle’s medium (DMEM) and fetal
bovine serum (FBS) were purchased from Thermo Fisher Scientific
Inc. (Waltham, MA, USA); TRIzol reagent and Lipofectamine 2000 were
obtained from Invitrogen (Carlsbad, CA, USA); M-MLV reverse
transcriptase was purchased from Promega (Madison, WI, USA);
SYBR-Green Master Mix was obtained from Takara (Otsu, Japan); and
the ECL Plus kit was obtained from GE Healthcare (Piscataway, NJ,
USA). The cell apoptosis kit was from KeyGen Biology (China).
Clinical samples and data
The tissue microarray was prepared for
immunohistochemical (IHC) analysis. Human OS tissues and the
corresponding ANCTs were obtained from biopsies in a total of 45
consecutive OS cases admitted to our hospital from January 2005 to
December 2011. The baseline characteristics of the patients before
neoadjuvant chemotherapy are summarized in Table I. The present study was approved by
the Medical Ethics Committee of the Second Military Medical
University, and written informed consent was obtained from the
patients or their parents before sample collection. Two
pathologists respectively reviewed all of the cases.
 | Table ICorrelation of TNFAIP1 expression and
clinicopathological factors of the OS patients. |
Table I
Correlation of TNFAIP1 expression and
clinicopathological factors of the OS patients.
| | TNFAIP1 expression
(n) | |
|---|
| |
| |
|---|
| Variables | Cases (n) | − | + | P-value |
|---|
| Total | 45 | 12 | 33 | |
| Age (years) | | | | 0.714 |
| <20 | 28 | 8 | 20 | |
| ≥20 | 17 | 4 | 13 | |
| Gender | | | | 0.964 |
| Male | 26 | 7 | 19 | |
| Female | 19 | 5 | 14 | |
| Histology | | | | 0.641 |
| Osteoblastic | 21 | 7 | 14 | |
| Chondroblastic | 15 | 3 | 12 | |
| Fibroblastic | 9 | 2 | 7 | |
| Ennecking
staging | | | | 0.492 |
| I | 13 | 3 | 10 | |
| II | 24 | 8 | 16 | |
| III | 8 | 1 | 7 | |
| Distant
metastasis | | | | 0.029 |
| No | 18 | 8 | 10 | |
| Yes | 27 | 4 | 23 | |
Tissue microarray
The Advanced tissue arrayer (ATA-100; Chemicon
International, Tamecula, CA, USA) was used to create holes in a
recipient paraffin block and to acquire cylindrical core tissue
biopsies with a diameter of 1 mm from the specific areas of the
‘donor’ block. The tissue core biopsies were transferred to the
recipient paraffin block at defined array positions. The tissue
microarrays contained tissue samples from 45 formalin-fixed
paraffin-embedded cancer specimens with known diagnosis, and the
corresponding ANCTs from these patients. The block was incubated in
an oven at 45°C for 20 min to allow complete embedding of the
grafted tissue cylinders in the paraffin of the recipient block,
and then stored at 4°C until microtome sectioning.
Immunohistochemical staining
Tissue microarray sections were processed for IHC
analysis of TNFAIP1 protein as follows. IHC examinations were
carried out on 3-mm thick sections. For anti-TNFAIP1 IHC, unmasking
was performed with 10 mM sodium citrate buffer, pH 6.0, at 90°C for
30 min. For anti-TNFAIP1 IHC, antigen unmasking was not necessary.
Sections were incubated in 0.03% hydrogen peroxide for 10 min at
room temperature to remove endogenous peroxidase activity, and then
in blocking serum [0.04% bovine serum albumin (A2153;
Sigma-Aldrich, Shanghai, China) and 0.5% normal goat serum (X0907;
Dako Corporation, Carpinteria, CA, USA) in phosphate-buffered
saline (PBS)] for 30 min at room temperature. The anti-TNFAIP1
antibody was used at a dilution of 1:200. The antibody was
incubated overnight at 4°C. Sections were then washed three times
for 5 min in PBS. Non-specific staining was blocked with 0.5%
casein and 5% normal serum for 30 min at room temperature. Finally,
staining was developed using diaminobenzidine substrate, and
sections were counterstained with hematoxylin. Normal serum or PBS
was used to replace the anti-TNFAIP1 antibody in the negative
controls.
Quantification of protein expression
The expression of TNFAIP1 was semi-quantitatively
estimated as total immunostaining scores, which were calculated as
the product of a proportion score and an intensity score. The
proportion and intensity of the staining were evaluated
independently by two observers. The proportion score reflected the
fraction of positive staining cells (0, none; 1, <10%; 2, 10 to
≥25%; 3, >25 to 50%; 4, >50%), and the intensity score
represented the staining intensity (0, no staining; 1, weak; 2,
intermediate; 3, strong). Finally, a total expression score was
given ranging from 0 to 12. Based on the analysis in advance,
TNFAIP1 expression was categorized into two groups: low-level
TNFAIP1 expression (score 0–3) and high-level TNFAIP1 expression
(score 4–12). The scoring was independently assessed by two
pathologists.
Cell culture and transfection
OS cell lines were cultured in DMEM supplemented
with 10% heat-inactivated FBS, 100 U/ml of penicillin and 100 μg/ml
of streptomycin. Cells in this medium were placed in a humidified
atmosphere containing 5% CO2 at 37°C. Cells were
subcultured at a 1:5 dilution in medium containing 300 μg/ml G418
(an aminoglycoside antibody, commonly used as a stable transfection
reagent in molecular genetic testing). On the day of transduction,
OS cells were replated at 5×104 cells/well in 24-well
plates containing serum-free growth medium with Polybrene (5
mg/ml). When reaching 50% confluency, the cells were transfected
with the recombinant experimental or control virus at the optimal
multiplicity of infection (MOI) of 50, and cultured at 37°C in 5%
CO2 for 4 h. Then the supernatant was discarded and
serum containing growth medium was added. At 4 days
post-transduction, transduction efficiency was measured by the
frequency of green fluorescent protein (GFP)-positive cells.
Positive and stable transfectants were selected and expanded for
further study. The TNFAIP1 siRNA (siTNFAIP1) vector-infected clone,
the negative control vector-infected cells and the untreated cells
were designated as the Lv-siTNFAIP1 group, NC group and CON group,
respectively.
Quantitative real-time PCR
To quantitatively determine the mRNA expression
level of TNFAIP1 in OS cells, real-time PCR was performed. Total
RNA was extracted from each clone using TRIzol according to the
manufacturer’s protocol. Reverse transcription was carried out
using M-MLV, and cDNA amplification was performed using the
SYBR-Green Master Mix kit according to the manufacturer’s
guidelines. The TNFAIP1 gene was amplified using a specific
oligonucleotide primer, and the β-actin gene was used as an
endogenous control. Data were analyzed using the comparative Ct
method (2−ΔΔCt). Three separate experiments were
performed for each clone.
Western blot assay
OS cell lines were harvested and extracted using
lysis buffer (Tris-HCl, SDS, mercaptoethanol and glycerol). Cell
extracts were boiled for 5 min in loading buffer, and then an equal
amount of cell extracts was separated on 15% SDS-PAGE gels.
Separated protein bands were transferred onto polyvinylidene
fluoride (PVDF) membranes, which were subsequently blocked in 5%
skim milk powder. Primary antibodies against TNFAIP1, p65NF-κB,
PCNA, matrix metalloproteinase-2 (MMP-2) and caspase-3 were diluted
according to the manufacturer’s instructions and incubated
overnight at 4°C. Subsequently, horseradish peroxidase-linked
secondary antibodies were added at a dilution of 1:1,000 and
incubated at room temperature for 2 h. The membranes were washed 3
times with PBS, and the immunoreactive bands were visualized using
the ECL Plus kit according to the manufacturer’s instructions. The
relative protein levels in the different cell lines were normalized
to the concentration of β-actin. Three separate experiments were
performed for each clone.
Cell proliferation assay
Cell proliferation was analyzed using the MTT assay.
Briefly, cells infected with the siTNFAIP1 virus were incubated in
96-well-plates at a density of 1×105 cells/well with
DMEM supplemented with 10% FBS. Cells were treated with 20 μl of
MTT dye at 0, 24, 48 and 72 h, and subsequently incubated with 150
μl of DMSO for 5 min. The color reaction was measured at 570 nm
using an Enzyme Immunoassay Analyzer (Bio-Rad, Hercules, CA, USA).
The proliferation activity was calculated for each clone.
Transwell invasion assay
Transwell filters were coated with Matrigel (3.9
μg/μl; 60–80 μl) on the upper surface of a polycarbonate membrane
(diameter, 6.5 mm; pore size, 8-μm). After incubation at 37°C for
30 min, the Matrigel solidified and served as the extracellular
matrix for analysis of tumor cell invasion. Harvested cells
(1×105) in 100 μl of serum-free DMEM were added into the
upper compartment of the chamber. A total of 200 μl of conditioned
medium derived from NIH3T3 cells was used as a source of
chemoattractant, which was placed in the bottom compartment of the
chamber. After 24 h of incubation at 37°C with 5% CO2,
the medium was removed from the upper chamber. The non-invaded
cells on the upper side of the chamber were scraped off with a
cotton swab. Cells that had migrated from the Matrigel into the
pores of the inserted filter were fixed with 100% methanol, stained
with hematoxylin, then mounted and dried at 80°C for 30 min. The
number of cells invading through the Matrigel was counted in 3
randomly selected visual fields from the central and peripheral
portion of the filter by using an inverted microscope (x200
magnification). Each assay was repeated 3 times.
Analysis of cell apoptosis
To detect cell apoptosis, OS cells treated with
siTNFAIP1 were trypsinized, washed with cold PBS and resuspended in
binding buffer according to the instructions provided in the
apoptosis kit. FITC-Annexin V and PI were added to the fixed cells
for 20 min in darkness at room temperature. Then, Annexin V binding
buffer was added to the mixture before the fluorescence was
measured on the FACSort flow cytometer. Cell apoptosis was analyzed
using CellQuest software (Becton-Dickinson, USA). Three separate
experiments were performed for each clone.
Statistical analysis
SPSS 20.0 was used for statistical analysis.
Kruskal-Wallis H and Chi-square tests were used to analyze the
expression rate in all groups. One-way analysis of variance (ANOVA)
was used to analyze the differences between groups. The LSD method
of multiple comparisons was used when the probability for ANOVA was
statistically significant. Statistical significance was set at
P<0.05.
Results
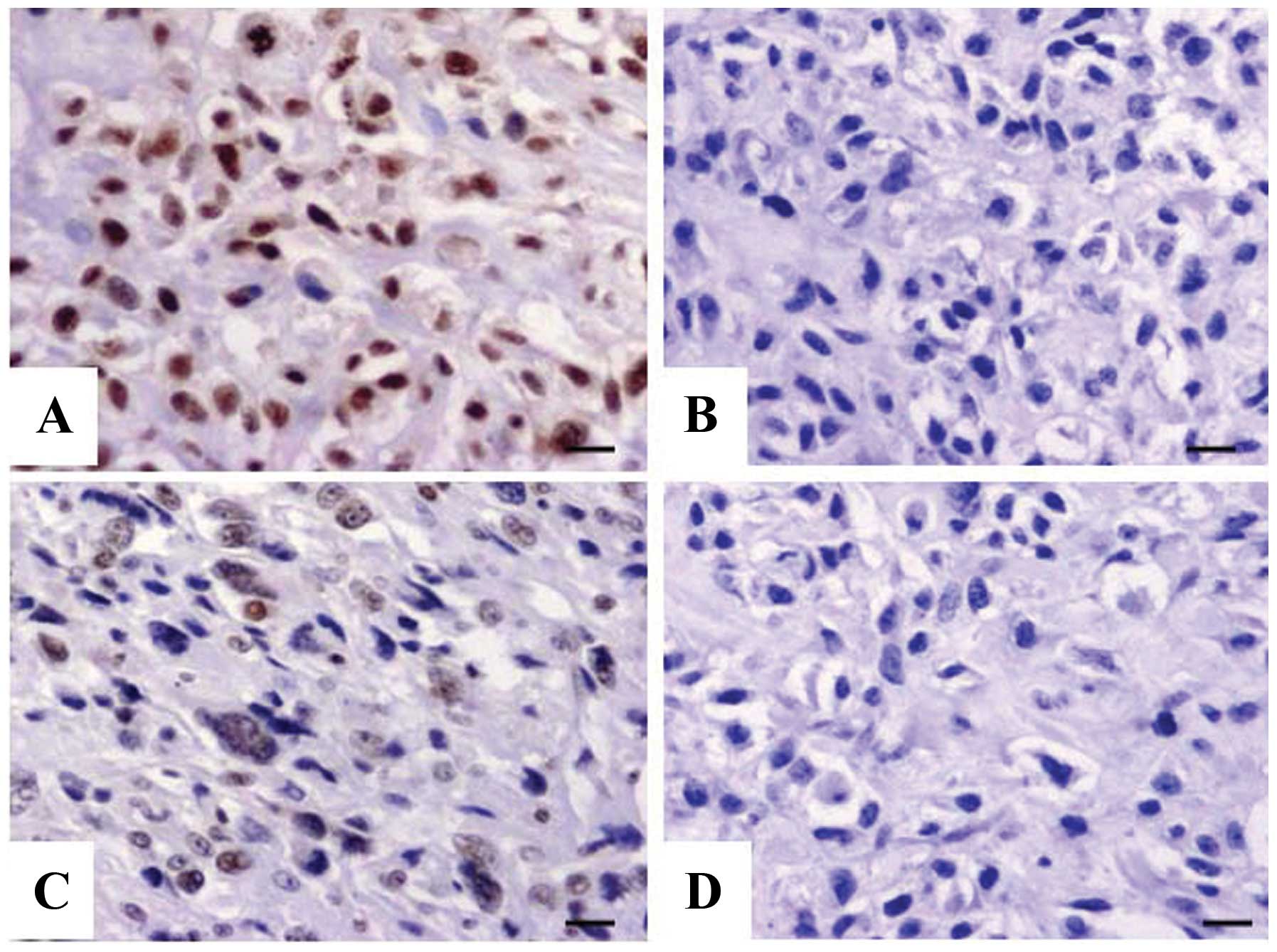
Expression of TNFAIP1 in OS tissues
The expression of TNFAIP1 protein was evaluated
using IHC staining in OS tissues. Different levels of positive
expression of TNFAIP1 protein were detected in the OS tissues
(Fig. 1A and B) and the ANCTs
(Fig. 1C and D). Positive TNFAIP1
immunostaining was mainly localized in the nucleus of the OS tissue
cells. According to the TNFAIP1 immunoreactive intensity, the
positive expression of TNFAIP1 in the OS tissues was significantly
increased compared with that in the ANCTs (P=0.018) (Table II).
| Table IIExpression of TNFAIP1 protein in OS
tissues. |
Table II
Expression of TNFAIP1 protein in OS
tissues.
| Target | Sample | TNFAIP1 expression
(n) | Total | Positive rate
(%) | χ2 | P-value |
|---|
|
|---|
| − | + | ++ | +++ |
|---|
| TNFAIP1 | OS | 12 | 17 | 10 | 6 | 45 | 73.3 | | |
| ANCTs | 23 | 13 | 6 | 3 | 45 | 48.9 | 5.607 | 0.018 |
Correlation of TNFAIP1 expression with
clinicopathological characteristics
The correlation of TNFAIP1 expression with various
clinicopathologic factors was analyzed. As shown in Table I, increased expression of TNFAIP1
was closely correlated with the distant metastasis of OS patients
(P=0.029). However, no significant association was found between
TNFAIP1 expression and other factors including age, gender of the
patients, and histology and Ennecking stage of the tumor
(P>0.05, respectively).
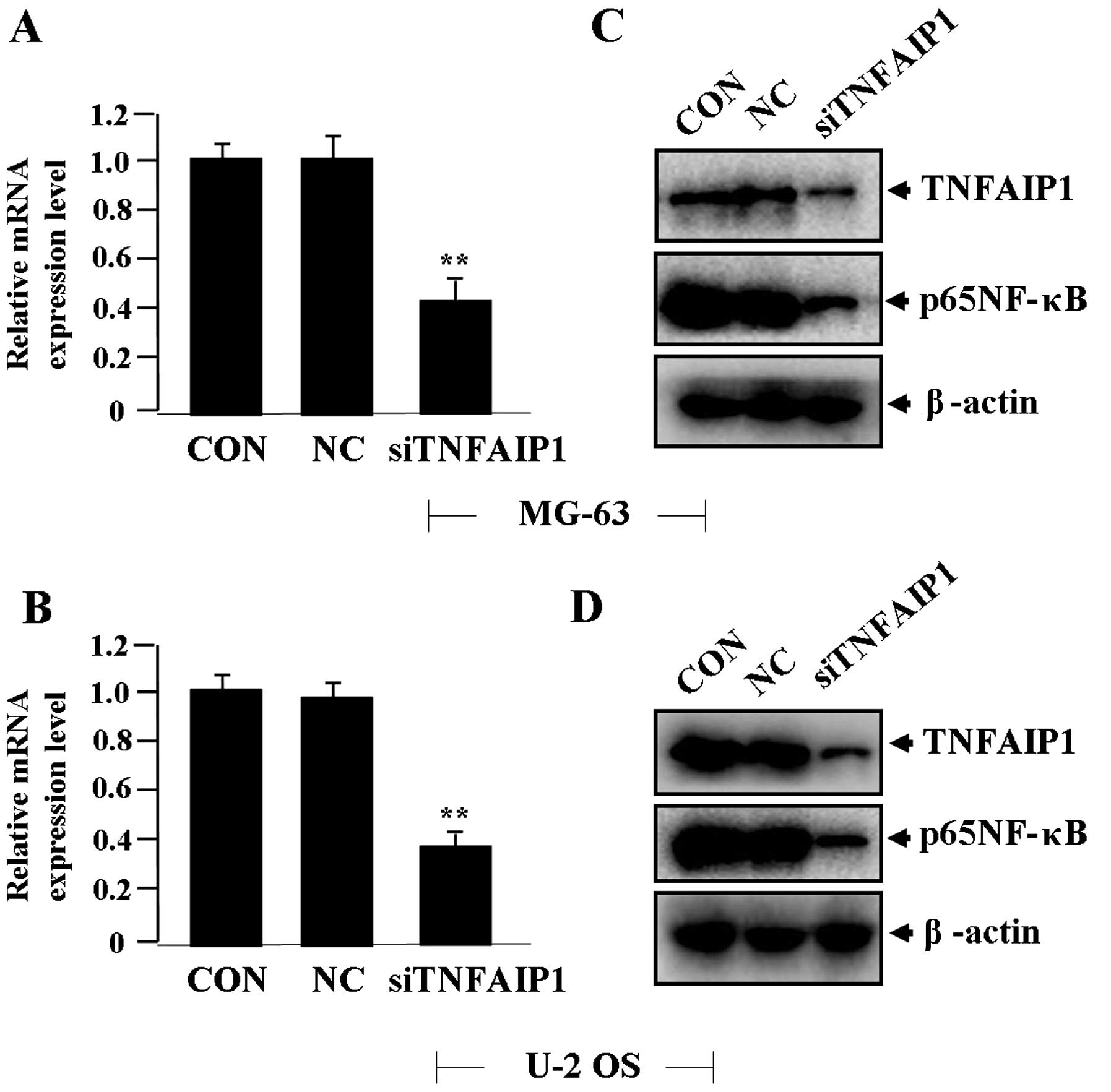
Effect of TNFAIP1 knockdown on the
expression of NF-κB
Lentiviruses of different multiplicity of infection
(MOI) were transfect into OS cells (MG-63 and U-2 OS), and the
transfection efficiency of siTNFAIP1 (MOI=50) reached >70%.
After siTNFAIP1 was transfected into the OS cells for 24 h, the
expression levels of TNFAIP1 mRNA (Fig.
2A and B) and protein levels of TNFAIP1 and p65NF-κB (Fig. 2C and D) were detected by real-time
PCR and western blotting, indicating decreased expression of
TNFAIP1 and p65NF-κB in the siTNFAIP1 group compared with the NC
and CON groups.
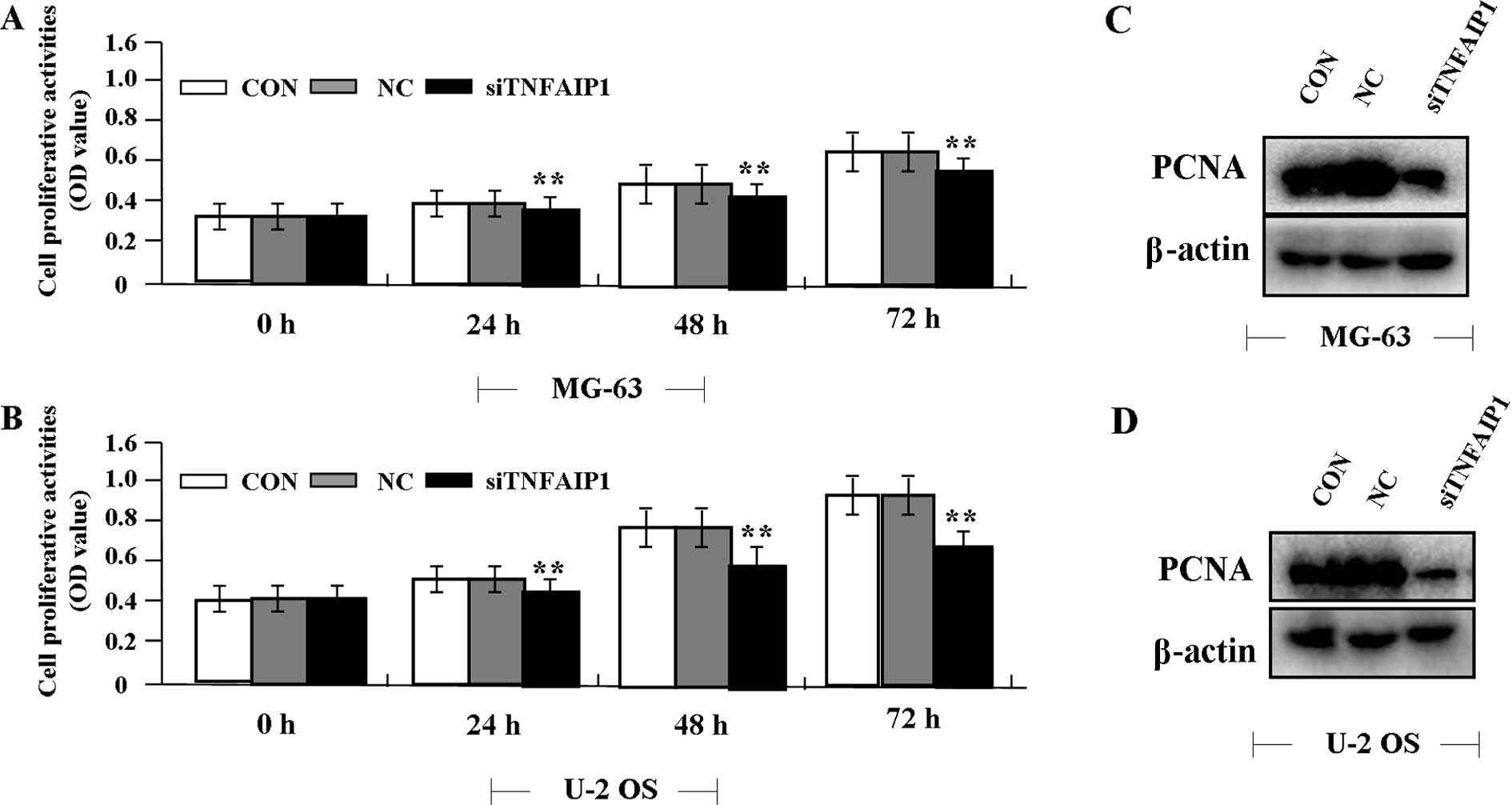
Effect of TNFAIP1 knockdown on cell
proliferation
Deregulated cell proliferation is a hallmark of
cancer. To confirm the effect of TNFAIP1 on OS cell growth, we
examined cell proliferative activities by MTT assay. The results
showed that knockdown of TNFAIP1 diminished the proliferative
activities of the OS cells in a time-dependent manner compared to
the NC group (Fig. 3A and B). In
addition, the expression level of PCNA protein, examined by western
blotting (Fig. 3C and D), was found
to be significantly downregulated in the siTNFAIP1 group when
compared with the level in the NC and NC groups.
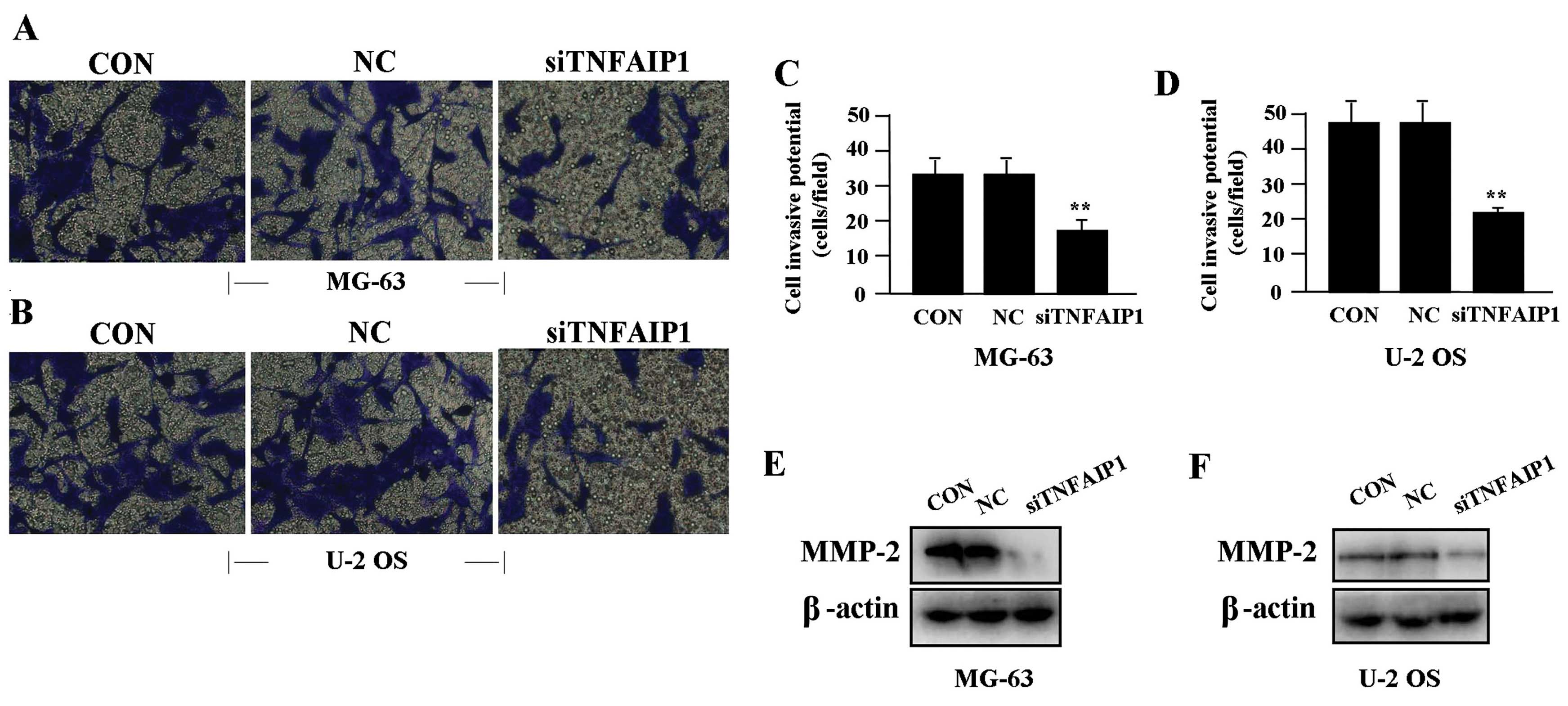
Effect of TNFAIP1 knockdown on cell
invasion
To determine the effect of TNFAIP1 on cell invasion,
a Transwell assay was performed. The invasive potential of tumor
cells in the Transwell assay was determined by the ability of cells
to invade a matrix barrier containing laminin and type IV collagen,
the major components of the basement membrane. Representative
micrographs of Transwell filters are shown in Fig. 4A and B. We found that the invasive
potential of OS cells was markedly lowered in the siTNFAIP1 group
when compared to the invasive potential in the NC and CON groups
(P<0.01) (Fig. 4C and D). In
addition, the expression level of MMP-2 protein, examined by
western blotting (Fig. 4E and F),
was found to be significantly downregulated in the siTNFAIP1 group
when compared to the level in the NC and CON groups.
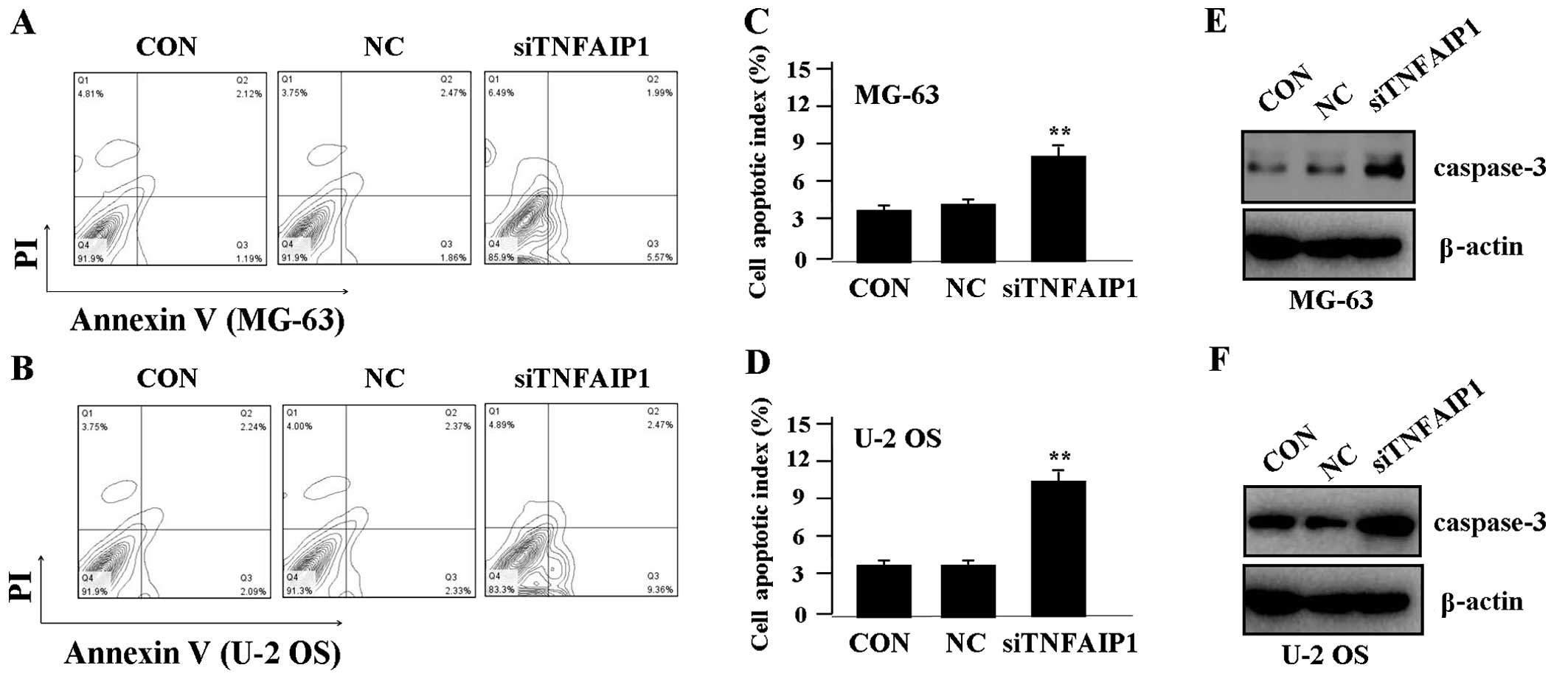
Effect of TNFAIP1 knockdown on cell
apoptosis
To determine whether TNFAIP1 affects OS cell
apoptosis, flow cytometric analysis with PI/FITC-Annexin V staining
was performed. We found that the apoptosis indices of the OS cells
were significantly higher in the siTNFAIP1 group when compared to
the indices in the NC and CON groups (Fig. 5A–D). Additionally, to determine
whether TNFAIP1 regulates the expression of caspase-3 protein,
western blotting was carried out. The protein level of caspase-3
was found to be upregulated in the siTNFAIP1 group in comparison
with the level in the NC and CON groups (Fig. 5E and F).
Discussion
TNF-α is an inflammatory cytokine that is present in
the microenvironment of many types of tumors, and is known to
promote tumor progression. Overexpression of TNFAIP1 or TNFAIP2
correlates with tumor invasion and metastasis, and serves as an
independent prognostic indicator for breast cancer (9) and nasopharyngeal carcinoma (15). The TNFAIP2 miRNA binding site (s8126
T>C SNP) is also correlated with a significantly elevated risk
of gastric cancer, suggesting a marker for susceptibility to
gastric cancer (16). To verify the
correlation of TNFAIP1 expression with OS, we examined the
expression of TNFAIP1 in human OS tissues. The present study showed
that TNFAIP1 was markedly upregulated in the nuclei of OS tissue
cells when compared to the ANCTs, and was positively correlated
with the distant metastasis of OS patients. These studies
demonstrated that TNFAIP1 may serve as a potential biomarker
involved in the development of OS.
Moreover, the function of TNFAIP requires further
research in cancer. First, TNFAIP1 confers acquired resistance to
paclitaxel, while knockdown of TNFAIP1 was found to increase the
tumor response to paclitaxel (17),
suggesting that TNFAIP may represent a valuable therapeutic target
for the treatment of cancer. Furthermore, TNFAIP1 was found to be
downregulated by miR-372/-373 and participates in cell apoptosis
and proliferation via the NF-κB pathway (18,19).
In addition, TNFAIP2 was found to promote tumor progression in
nasopharyngeal carcinoma (NPC) (20), while knockdown of TNFAIP2 was found
to markedly reduce the migration and invasion of NPC cells
(15). The present study also
showed that knockdown of TNFAIP1 repressed cell proliferation and
invasion, and induced cell apoptosis in OS cells, which is
supported by data that TNFAIP2 can mediate the inhibitory effects
of retinoic acid on endometrial cancer cell growth, invasion and
apoptosis escape (21), suggesting
TNFAIP1/2 as a potential target for retinoic acid in acute
promyelocytic leukemia (22). Yet,
research found that simultaneous expression of RhoB and TNFAIP1
resulted in a marked increase in apoptosis in HeLa cells (23), which is opposite from our findings
concerning the decrease in cell apoptosis induced by TNFAIP1 in OS
cells, suggesting that the underlying molecular mechanisms of
TNFAIP1 in cancer require further exploration.
Activation of NF-κB promotes the development of OS
(14), while inhibition of NF-κB
activity suppresses the tumorigenicity of Ewing sarcoma EW7 cells
(24). It has been reported that
NF-κB-mediated transcriptional upregulation of TNFAIP2 by the
Epstein-Barr virus oncoprotein, LMP1, promotes cell motility in NPC
(20). However, few studies have
shown that TNFAIP1 inhibits the transcriptional activities of NF-κB
(25). In the present study, we
found that knockdown of TNFAIP1 downregulated the expression of
NF-κB at the translational level in OS cells, suggesting that
TNFAIP1 may promote the progression of OS through activation of the
NF-κB pathway.
PCNA is a nuclear protein that is expressed in
proliferating cells and may be required for maintaining cell
proliferation, and is used as a marker for cell proliferation of
colon cancer (26). MMP-2 is
believed to be a key enzyme involved in the degradation of type IV
collagen, and a high level of MMP-9 in tissues is associated with
tumor invasion and metastasis (27). NF-κB was found to promote the growth
and invasion and block the apoptosis of tumor cells via regulation
of the expression of PCNA (28),
MMP-2 (29) and apoptosis (30). Moreover, it was found that TNFAIP1
promotes cell growth and gene amplification in a PCNA-dependent way
(5,31), yet no report has demonstrated the
regulation of the expression of MMP-2 and caspase-3 by TNFAIP1. In
the present study, we found that knockdown of TNFAIP1 downregulated
the expression of PCNA and MMP-9 and upregulated the expression of
caspase-3 in OS cells, suggesting that TNFAIP1 may promote the
progression of OS through NF-κB pathway-mediated regulation of
PCNA, MMP-2 and caspase-3 expression.
In conclusion, our findings indicate that high
expression of TNFAIP1 is associated with the distant metastasis of
OS, and knockdown of TNFAIP1 inhibits the growth and invasion, and
induces apoptosis in OS cells through inhibition of the NF-κB
pathway, suggesting that TNFAIP1 may act as a potential therapeutic
target for the treatment of cancer.
References
|
1
|
Kuijjer ML, van den Akker BE, Hilhorst R,
et al: Kinome and mRNA expression profiling of high-grade
osteosarcoma cell lines implies Akt signaling as possible target
for therapy. BMC Med Gen. 7:42014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Buddingh EP, Anninga JK, Versteegh MI, et
al: Prognostic factors in pulmonary metastasized high-grade
osteosarcoma. Pediatr Blood Cancer. 54:216–221. 2010.PubMed/NCBI
|
|
3
|
Bacci G, Longhi A, Versari M, et al:
Prognostic factors for osteosarcoma of the extremity treated with
neoadjuvant chemotherapy: 15-year experience in 789 patients
treated at a single institution. Cancer. 106:1154–1161.
2006.PubMed/NCBI
|
|
4
|
Savage SA, Mirabello L, Wang Z, et al:
Genome-wide association study identifies two susceptibility loci
for osteosarcoma. Nat Genet. 45:799–803. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yang L, Liu N, Hu X, et al: CK2
phosphorylates TNFAIP1 to affect its subcellular localization and
interaction with PCNA. Mol Biol Rep. 37:2967–2973. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Link CD, Taft A, Kapulkin V, et al: Gene
expression analysis in a transgenic Caenorhabditis elegans
Alzheimer’s disease model. Neurobiol Aging. 24:397–413. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lin MC, Lee NP, Zheng N, et al: Tumor
necrosis factor-α-induced protein 1 and immunity to hepatitis B
virus. World J Gastroenterol. 11:7564–7568. 2005.
|
|
8
|
Gupta J, Gaikwad AB and Tikoo K: Hepatic
expression profiling shows involvement of PKC epsilon, DGK eta,
Tnfalpha, and Rho kinase in type 2 diabetic nephropathy rats. J
Cell Biochem. 111:944–954. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Grinchuk OV, Motakis E and Kuznetsov VA:
Complex sense-antisense architecture of TNFAIP1/POLDIP2 on 17q11.2
represents a novel transcriptional structural-functional gene
module involved in breast cancer progression. BMC Genomics.
11(Suppl 1): S92010. View Article : Google Scholar
|
|
10
|
Kondratiev S, Duraisamy S, Unitt CL, et
al: Aberrant expression of the dendritic cell marker TNFAIP2 by the
malignant cells of Hodgkin lymphoma and primary mediastinal large
B-cell lymphoma distinguishes these tumor types from
morphologically and phenotypically similar lymphomas. Am J Surg
Pathol. 35:1531–1539. 2011. View Article : Google Scholar
|
|
11
|
Dumon K, Rossbach C, Harms B, et al: Tumor
necrosis factor-alpha (TNF-alpha) gene polymorphism in surgical
intensive care patients with SIRS. Langenbecks Arch Chir Suppl
Kongressbd. 115(Suppl I): 387–390. 1998.(In German).
|
|
12
|
Liu XW, Lu FG, Zhang GS, et al: Proteomics
to display tissue repair opposing injury response to LPS-induced
liver injury. World J Gastroenterol. 10:2701–2705. 2004.PubMed/NCBI
|
|
13
|
Sparks AB, Peterson SN, Bell C, et al:
Mutation screening of the TNFRSF11A gene encoding receptor
activator of NFκB (RANK) in familial and sporadic Paget’s disease
of bone and osteosarcoma. Calcif Tissue Int. 68:151–155. 2001.
|
|
14
|
Javelaud D and Besançon F: NF-κB
activation results in rapid inactivation of JNK in TNFα-treated
Ewing sarcoma cells: a mechanism for the anti-apoptotic effect of
NF-κB. Oncogene. 20:4365–4372. 2001.
|
|
15
|
Chen LC, Chen CC, Liang Y, et al: A novel
role for TNFAIP2: its correlation with invasion and metastasis in
nasopharyngeal carcinoma. Mod Pathol. 24:175–184. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xu Y, Ma H, Yu H, et al: The miR-184
binding-site rs8126 T>C polymorphism in TNFAIP2 is
associated with risk of gastric cancer. PLoS One.
8:e649732013.PubMed/NCBI
|
|
17
|
Zhu Y, Yao Z, Wu Z, et al: Role of tumor
necrosis factor alpha-induced protein 1 in paclitaxel resistance.
Oncogene. Aug 5–2013.(Epub ahead of print). View Article : Google Scholar
|
|
18
|
Zhou C, Li X, Zhang X, et al: microRNA-372
maintains oncogene characteristics by targeting TNFAIP1 and affects
NFκB signaling in human gastric carcinoma cells. Int J Oncol.
42:635–642. 2013.PubMed/NCBI
|
|
19
|
Zhang X, Li X, Tan Z, et al: MicroRNA-373
is upregulated and targets TNFAIP1 in human gastric cancer,
contributing to tumorigenesis. Oncol Lett. 6:1427–1434.
2013.PubMed/NCBI
|
|
20
|
Chen CC, Liu HP, Chao M, et al:
NF-κB-mediated transcriptional upregulation of TNFAIP2 by the
Epstein-Barr virus oncoprotein, LMP1, promotes cell motility in
nasopharyngeal carcinoma. Oncogene. Aug 26–2013.(Epub ahead of
print). View Article : Google Scholar
|
|
21
|
Cheng YH, Utsunomiya H, Pavone ME, et al:
Retinoic acid inhibits endometrial cancer cell growth via multiple
genomic mechanisms. J Mol Endocrinol. 46:139–153. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rusiniak ME, Yu M, Ross DT, et al:
Identification of B94 (TNFAIP2) as a potential retinoic acid target
gene in acute promyelocytic leukemia. Cancer Res. 60:1824–1829.
2000.PubMed/NCBI
|
|
23
|
Kim DM, Chung KS, Choi SJ, et al: RhoB
induces apoptosis via direct interaction with TNFAIP1 in
HeLa cells. Int J Cancer. 125:2520–2527. 2009.PubMed/NCBI
|
|
24
|
Javelaud D, Poupon MF, Wietzerbin and
Besançon F: Inhibition of constitutive NF-κB activity suppresses
tumorigenicity of Ewing sarcoma EW7 cells. Int J Cancer.
98:193–198. 2002.
|
|
25
|
Hu X, Yan F, Wang F, et al: TNFAIP1
interacts with KCTD10 to promote the degradation of KCTD10 proteins
and inhibit the transcriptional activities of NF-κB and AP-1. Mol
Biol Rep. 39:9911–9919. 2012.PubMed/NCBI
|
|
26
|
Risio M: Cell proliferation in colorectal
tumor progression: an immunohistochemical approach to intermediate
biomarkers. J Cell Biochem (Suppl). 16G:79–87. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hornebeck W, Lambert E, Petitfrère E and
Bernard P: Beneficial and detrimental influences of tissue
inhibitor of metalloproteinase-1 (TIMP-1) in tumor progression.
Biochimie. 87:377–383. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shah SJ and Sylvester PW: γ-Tocotrienol
inhibits neoplastic mammary epithelial cell proliferation by
decreasing Akt and nuclear factor κB activity. Exp Biol Med.
230:235–241. 2005.
|
|
29
|
Philip S, Bulbule A and Kundu GC:
Osteopontin stimulates tumor growth and activation of promatrix
metalloproteinase-2 through nuclear factor-κB-mediated induction of
membrane type 1 matrix metalloproteinase in murine melanoma cells.
J Biol Chem. 276:44926–44935. 2001.
|
|
30
|
Moalic-Juge S, Liagre B, Duval R, et al:
The anti-apoptotic property of NS-398 at high dose can be mediated
in part through NF-κB activation, hsp70 induction and a decrease in
caspase-3 activity in human osteosarcoma cells. Int J Oncol.
20:1255–1262. 2002.PubMed/NCBI
|
|
31
|
Zhou J, Hu X, Xiong X, et al: Cloning of
two rat PDIP1 related genes and their interactions with
proliferating cell nuclear antigen. J Exp Zool A Comp Exp Biol.
303:227–240. 2005. View Article : Google Scholar : PubMed/NCBI
|