Introduction
microRNAs (miRNAs), small non-coding RNAs that are
20–22 nucleotides in length, play important roles in cancer
including proliferation, aggressiveness and development of
metastasis (1–3). miRNAs have the ability to modulate key
cellular processes that define the cell phenotype, making them
highly promising therapeutic targets. Glioblastoma multiforme (GBM)
is the most common and aggressive primary brain tumor in adults.
Elucidation of the molecular pathogenesis of GBM is crucial to
improve the overall survival of patients (4). The current standard therapy for GBM
includes surgical resection or biopsy, followed by a combination of
radiation (RT) and chemotherapy with temozolomide (5). Temozolomide (TMZ), an alkylating
agent, is a cytotoxic antitumor prodrug against malignant brain
tumors. At physiological pH, TMZ undergoes decomposition to the
active compound MTIC (6,7). Although TMZ may restrain GBM growth,
TMZ resistance is common and accounts for GBM treatment failure
(8). Recent analyses have revealed
that several miRNAs are clinically implicated in GBM, with some
reports indicating the association of the regulation of miRNA
expression and the chemosensitivity to TMZ (8–11). For
example, expression levels of miR-181b and miR-181c may serve as
predictive markers of response to RT/TMZ therapy in glioblastoma
patients (12). Chronic TMZ
exposure was found to result in acquired TMZ resistance and
elevated miR-21 expression. Concomitant treatment with miR-21
inhibitor and TMZ resulted in a significantly higher apoptosis
rather than TMZ treatment alone (8). miR-21 inhibitor combined with TMZ
significantly enhanced human glioblastoma stem cell apoptosis
(8). miR-125b-2 is overexpressed in
glioblastoma multiforme tissues and the corresponding stem cells
(GBMSCs); downregulation of miR-125b-2 expression in GBMSCs could
allow TMZ to induce GBMSC apoptosis (13). Our previous research found that
miR-128, miR-149 and miR-181a/b were significantly downregulated
and may be tumor suppressors in astrocytomas (14). Further research validated that
miR-181a/b/c/d were significantly downregulated in astrocytomas and
glioblastoma of different grades, and their overexpression
inhibited the growth and invasion of glioblastoma cells and
enhanced the chemosensitivity of TMZ by Rap1B-mediated cytoskeletal
remodeling. Rap1B is a member of the Rap1 family, and is a Ras-like
GTPase that regulates several basic cellular functions, such as
growth, adhesion and migration (15). Rap1B was also found to be a possible
negative regulator of miR-128 and miR-149 by a docking study of
miRNA/mRNA omics in human grade I-III astrocytomas (14). Recent studies suggest that miR-128
and miR-149 play an important role in various diseases, including
the progression of malignant tumors. miR-128 is a brain-enriched
miRNA, which has tissue-specific and developmental-specific
expression patterns, mainly in neurons rather than in astrocytes
(16). Downregulation of miR-128
has been reported in several cancer types, for example,
neuroblastoma (17) glioblastoma
(14,18,19)
prostate (20), lung cancer
(21) and MLL-AF4 acute lymphocytic
leukemia (22). miR-128 exerted a
pro-apoptotic effect in a p53 transcription-dependent and
-independent manner via the PUMA-Bak axis in HEK293T cells (human
embryonic kidney), HCT116 cells (human colon carcinoma cells) and
MCF-7 cells (human breast adenocarcinoma), and miR-128 regulated
the chemotherapeutic sensitivity of breast tumor-initiating cells
via Bmi-1 and ABCC5 (23). The
overexpression of miR-128 was found to suppress p70S6K1 and
expression of its downstream signaling molecules HIF-1 and VEGF,
and to attenuate cell proliferation, tumor growth and angiogenesis
(24).
Recent studies have revealed the essential role of
miR-149 in various types of diseases, including the progression of
various types of malignant tumors. However, the results are
controversial, and miR-149 has been shown to function as both a
tumor suppressor (25) and as an
oncogene (26) in the development
of multiple types of solid tumors. p53 directly upregulates miR-149
which in turn targets glycogen synthase kinase-3α, resulting in
increased expression of Mcl-1 and resistance to apoptosis in
melanoma cells (26). miR-149 was
found to be significantly downregulated in GC cell lines and
clinical samples in comparison to normal gastric epithelial cells
and adjacent non-tumor tissues, respectively (27). miR-149 suppresses tumor cell
motility, and the pre-miR-149 polymorphism may affect the
processing of miR-149, resulting in a change in the abundance of
the mature form of miRNA, which, in turn, modulates tumor
progression and patient survival (28). miR-149 was found to be downregulated
and to function as a tumor suppressor by inhibiting oncogene
expression, and it inhibited the proliferation and invasion of
glioma cells by blocking AKT1 signaling in glioblastoma (29).
In the present study, we further confirmed changes
in the expression of miR-128 and miR-149 in astrocytomas of
different grades (WHO I, II and III) and glioblastoma (WHO IV), and
we investigated the effect of miR-128 and miR-149 on cellular
proliferation, invasion and its functional mechanism by targeting
Rap1B. We also demonstrated that miR-128 or miR-149 enhanced the
chemosensitivity of temozolomide by Rap1B-mediated cytoskeletal
remodeling in glioblastoma.
Materials and methods
Samples
Human glioma-derived cell lines U251 and U87 were
obtained from the Cell Center of the Peking Union Medical College
in China. All human primary brain tumor samples were obtained from
randomly selected cancer patients at the Second Affiliated Hospital
of Xiangya, China, and all of the diagnoses were pathologically
confirmed. Written informed consent was obtained from each patient
who participated in the study before surgery, and all of the
protocols were reviewed by the Joint Ethics Committee of the
Central South University Health Authority and performed in
accordance with national guidelines.
Cell culture
The U251 and U87 cells were grown in Dulbecco’s
modified Eagle’s medium (DMEM) with 10% fetal calf serum (FCS) and
standard antibiotics. All cells were maintained at 37°C under an
atmosphere of 5% CO2 and 95% air.
RNA extraction and real-time PCR
Real-time PCR was carried out as previously
described (30). Total RNA was
extracted using TRIzol® reagent (Invitrogen, Carlsbad,
CA, USA) from samples. Real-time PCR reactions were performed using
a qSYBR-Green-containing PCR kit (Invitrogen) and human β-actin or
U6 snRNA was used as an endogenous control for mRNA or miRNA
detection, respectively. Expression of each gene was quantified by
measuring Ct values and normalized using the 2−ΔΔct
method relative to U6 snRNA or β-actin.
Luciferase assay
The 3′-untranslated regions (UTRs) of the Rap1B gene
were synthesized, annealed, and then inserted into the pMIR-REPORT™
luciferase vector (Ambion) using the HindIII (aagctt) and
SpeI (actagt) sites located downstream from the stop codon
of luciferase. There was one region of the Rap1B gene 3′-UTRs for
miR-128 and miR-149 in TargetScan 5.2 software. The Luc-wt or
Luc-mut plasmid was co-transfected with miR-128 or miR-149 mimics
into U251 cells, and the pMIR-REPORT β-galactosidase control vector
was transfected as a control. Firefly luciferase activity was
measured consecutively using the luciferase assay system (Promega)
48 h after transfection. All the experiments were performed at
least three times.
Protein extraction and western
blotting
Protein extraction and western blot analysis were
performed as previously described (30). Rabbit polyclonal primary antibodies
against Rap1B, Cdc42, RhoA and N-cadherin were purchased from Cell
Signaling Technology (Beverly, MA, USA), and mouse monoclonal
antibodies against GAPDH and α-tubulin were purchased from Santa
Cruz Biotechnology, Inc. (Santa Cruz, CA, USA).
Colony formation assay
Synthesized RNA duplexes of scrambled miRNAs,
miR-128 and miR-149 mimics were obtained from GeneChem (Shanghai,
China). Temozolomide (T-2706; RD, USA) was obtained from Sigma.
Cells were seeded at 1×103 cells/well in 6-well plates
and transfected with miR-128 or miR-149 mimics followed by
treatment with 100 μM TMZ after transfection. Two weeks later,
cells were fixed in methanol for 30 min and stained with 1% crystal
violet dye, and the number of colonies was counted. Experiments
were repeated three times, and values are expressed as mean and
standard deviation.
Cell viability assay
Cell viability was determined by the MTT assay as
previously described (30).
Briefly, 1,500 cells/well were seeded into 96-well plates and were
treated by miR-128 or miR-149 mimic transient transfection and/or
TMZ administration, and MTT (Sigma-Aldrich, St. Louis, MO, USA)
assays were performed daily at different times. In this assay, the
medium was replaced with fresh medium containing 0.5 mg/ml MTT for
4 h and then carefully removed. Subsequently, 150 μl of dimethyl
sulfoxide (DMSO) was added to each well and mixed for 10 min, and
the optical density of each well was determined with a scanning
multi-well spectrophotometer at a wavelength of 490 nm. The
experiment was repeated three times; six parallel samples were
measured each time.
Wound closure assay
The cells were transfected with the synthetic
miR-128 or miR-149 mimics and grown to 90% confluency in a 6-well
dish. A wound was created using a sterile 10-μl pipette tip
followed by a wash with 1× phosphate-buffered saline (PBS) to
remove detached cells. Then, the cells were cultured in medium with
2% serum, and migration at the corresponding wound site was
documented using a micro-scope (Nikon) at different time-points (0,
12, 24 and 48 h).
Transwell migration assay
The Transwell migration assay was performed as
previously described (18).
Immunofluorescence
U251 cells were transfected with miR-128 or miR-149
mimics, Rap1B siRNA and NC or treatment with TMZ, respectively. The
cells were fixed with 4% paraformaldehyde in PBS for 20 min,
permeabilized in 0.25% Triton X-100 and blocked with 5% BSA in PBS.
Then, cells with or without treatment were stained with
FITC-labeled phalloidin (P5282; Sigma, Seelze, Germany) and DAPI
(C1002; Beyotime Institute of Biotechnology, Shanghai, China).
Images were acquired by laser scanning confocal microscopy
(TCS-SP5; Leica Microsystems, Wetzlar, Germany).
Statistical analysis
Differences between the variables of the groups were
tested using the Student’s t-test or one-way ANOVA, using the SPSS
15.0 program. A P-value of <0.05 was considered to indicate a
statistically significant result.
Results
miR-128 and miR-149 expression in the
astrocytomas of different grades
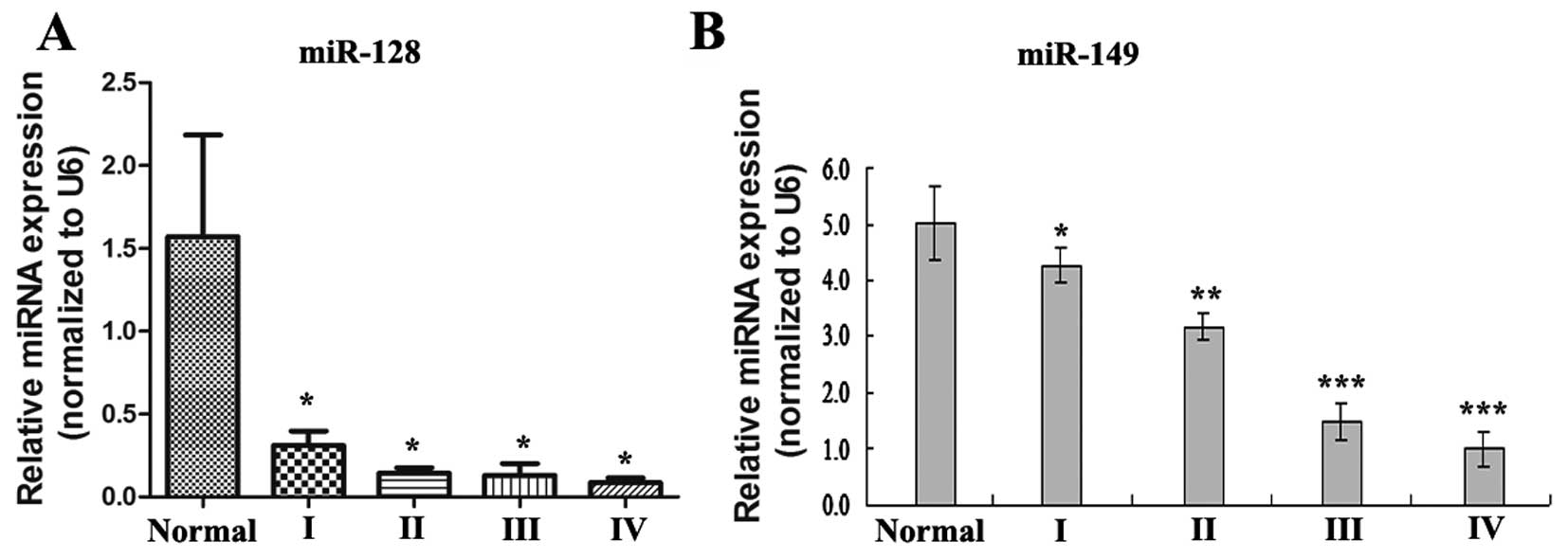
As shown in Fig. 1,
compared to expression in normal brain tissues (n=8), the
expression of miR-128 and miR-149 was decreased in most of the
primary astrocytomas (n=20), in which the expression of miR-149
gradually decreased with pathological progression in the different
WHO grade astrocytomas. However, the expression of miR-128 did not
gradually decrease with pathological progression in the different
WHO grade astrocytomas. The expression of miR-128 was not
statistically significantly different between grades II, III and IV
(P>0.05), but there was statistically significant differences
between grade I and grades II, III and IV astrocytomas
(P<0.05).
Expression of Rap1B is negatively
regulated by miR-128 and miR-149
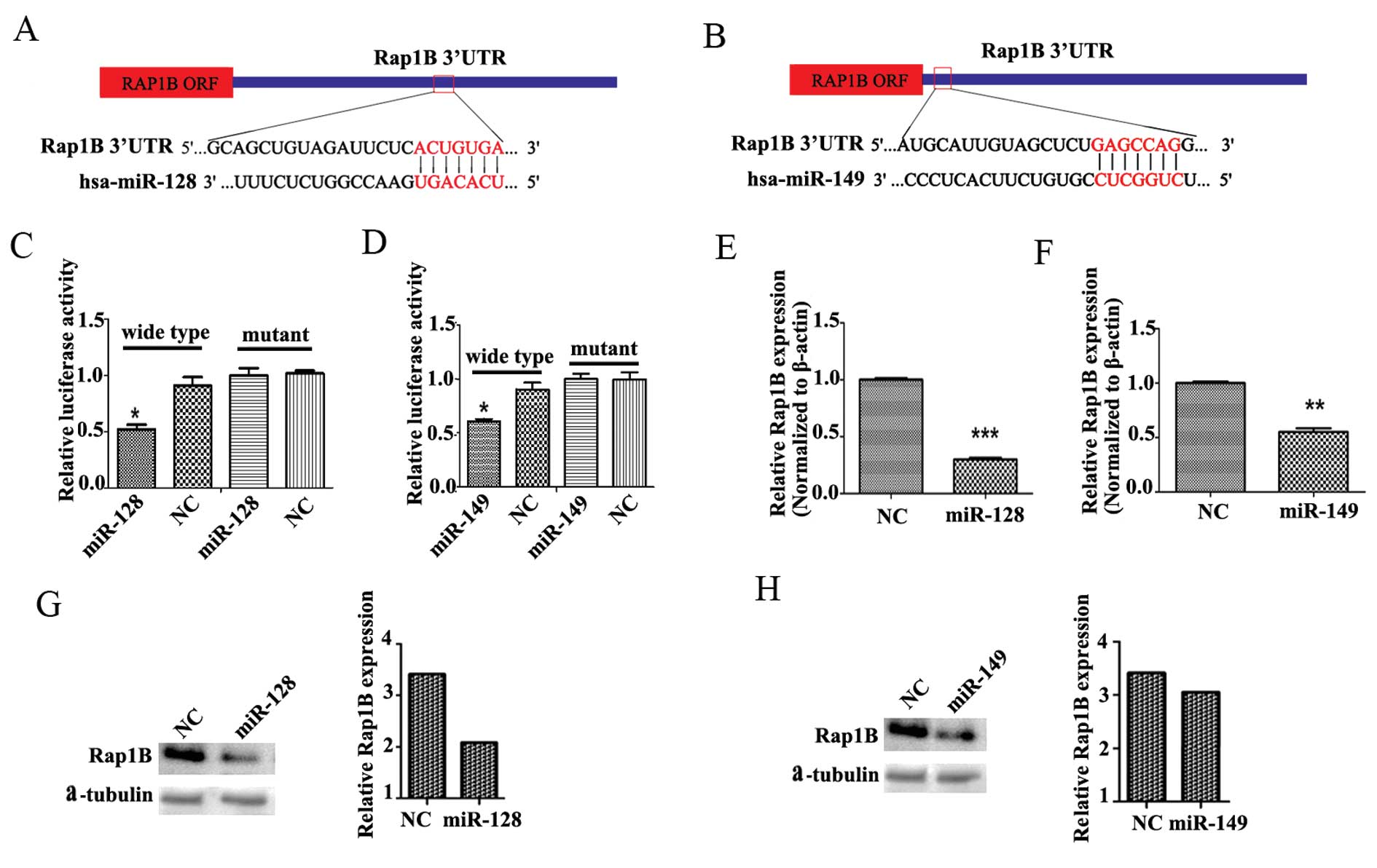
As shown in Fig. 2A and
B, the miRNA target prediction program TargetScan5.2 was used
to identify miR-128 and miR-149 interaction sites in the 3′-UTR of
Rap1B. miR-128 and miR-149 mimics had their own binding sites for
the 3′-UTR of Rap1B. The luciferase reporter assays showed that
miR-128 and miR-149 combined with the sites of the 3′-UTR of Rap1B,
respectively. Rap1B is a common bona fide target of miR-128 and
miR-149 (Fig. 2C and D).
Transfection of the miR-128 and miR-149 mimics resulted in a marked
reduction in Rap1B at both the mRNA (Fig. 2E and F) and protein levels (Fig. 2G and H) in U251 cells, as compared
with negative control (NC) transfection. We also previously
evaluated the expression of Rap1B in the astrocytomas tissues and
normal brain tissues, and found that the expression of Rap1B was
gradually increased with pathological progression in the WHO grade
astrocytomas (18). In the present
study, we found that the expression of miR-128 or miR-149 and Rap1B
was in an inverse trend in the astrocytoma and normal brain
tissues. The above data indicate that miR-128 and miR-149
negatively regulate the expression of Rap1B.
TMZ inhibits Rap1B expression by
upregulating miR-128 and miR-149
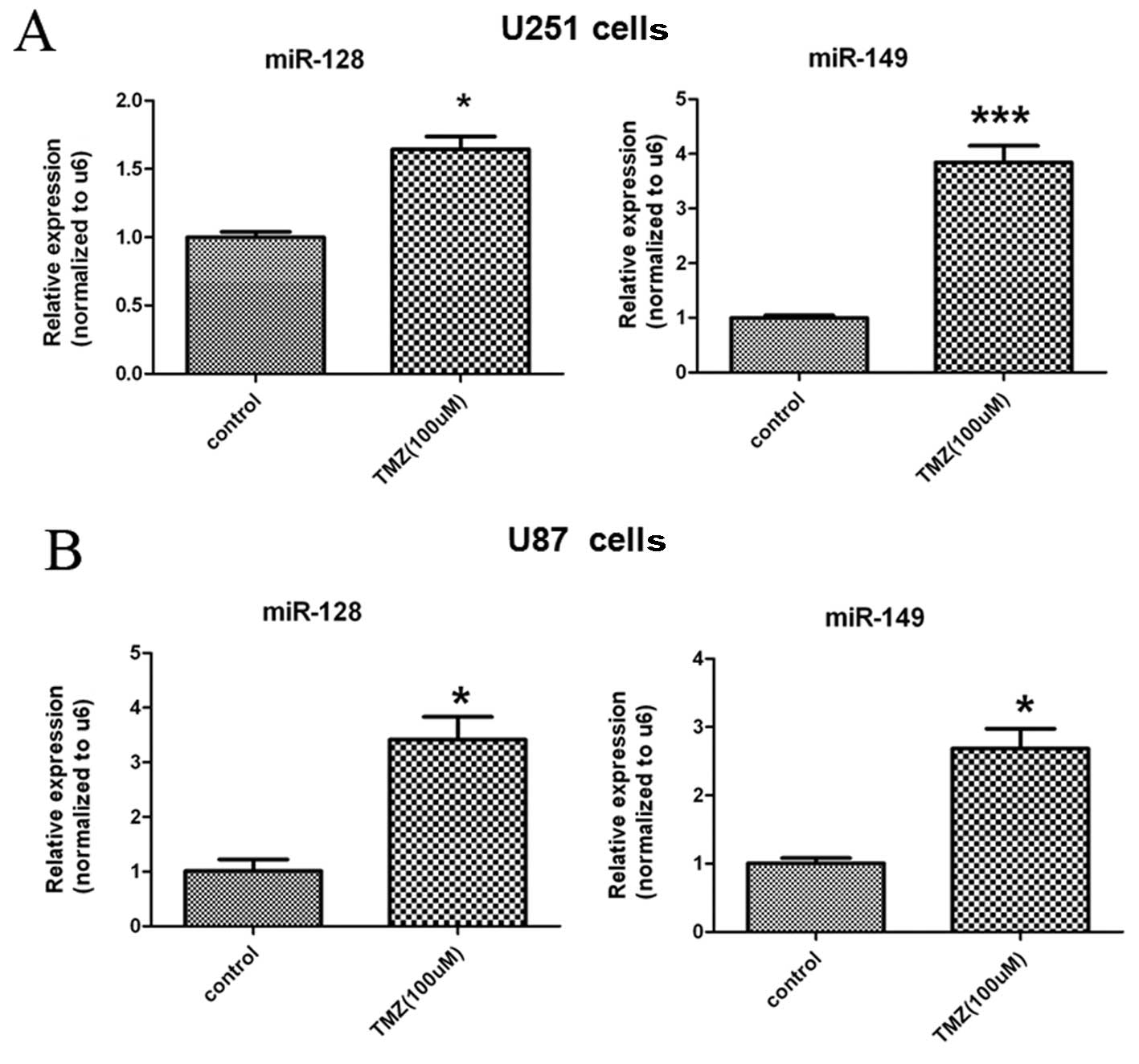
As shown in Fig. 3,
we observed that TMZ increased the expression of miR-128 and
miR-149 by real-time PCR when glioblastoma U251 and U87 cells were
treated with 100 μM TMZ. Our previous research demonstrated that
TMZ also decreased the expression of Rap1B at the mRNA and protein
levels by real-time PCR and western blotting (18). Given Rap1B is a common target of
miR-128 and miR-149 (Fig. 2), we
proposed that TMZ may inhibit Rap1B expression by upregulating
miR-128 and miR-149.
miR-128 and miR-149 suppress cell
proliferation and invasion, and alter cytoskeletal remodeling by
affecting Rap1B-associated small GTPase
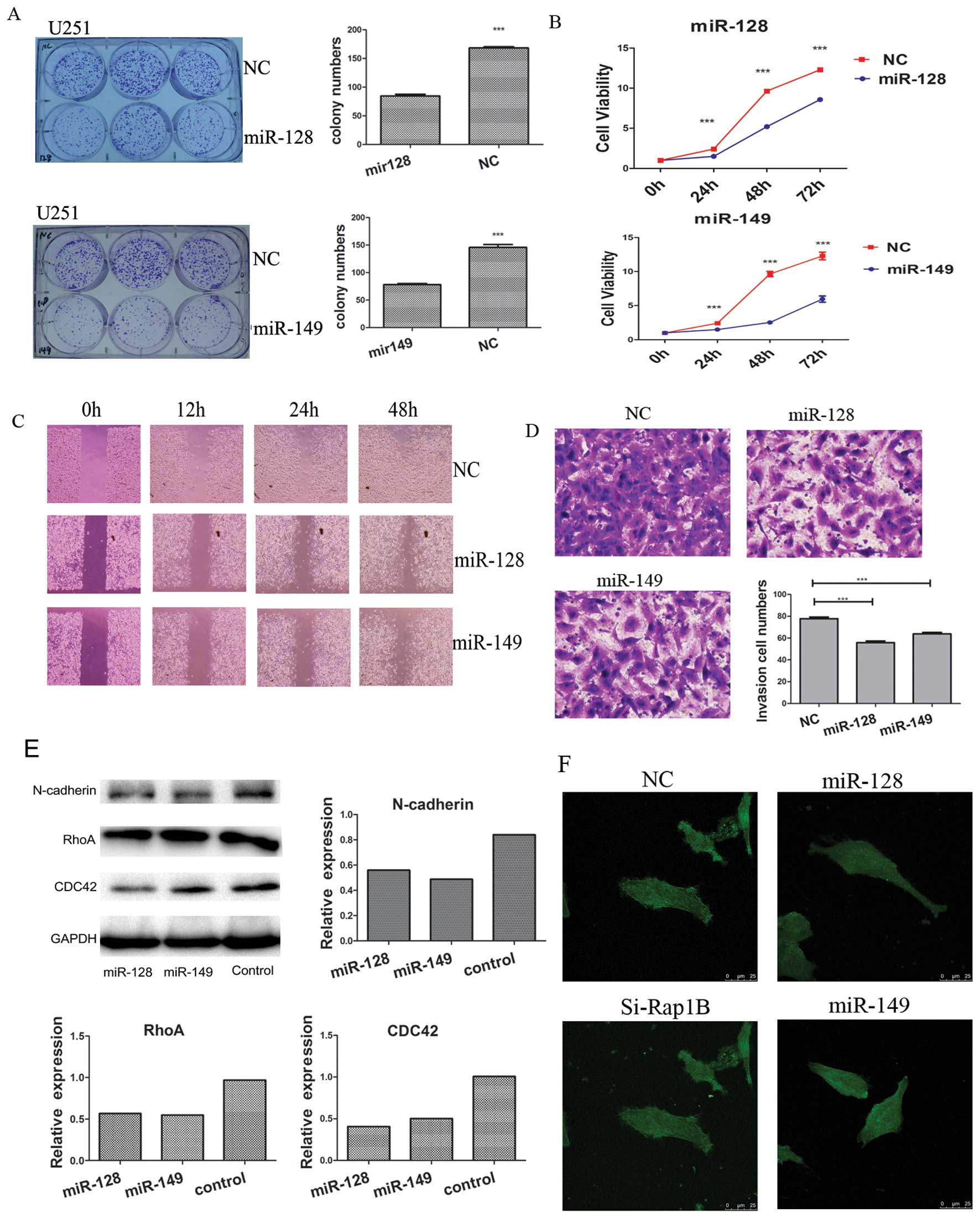
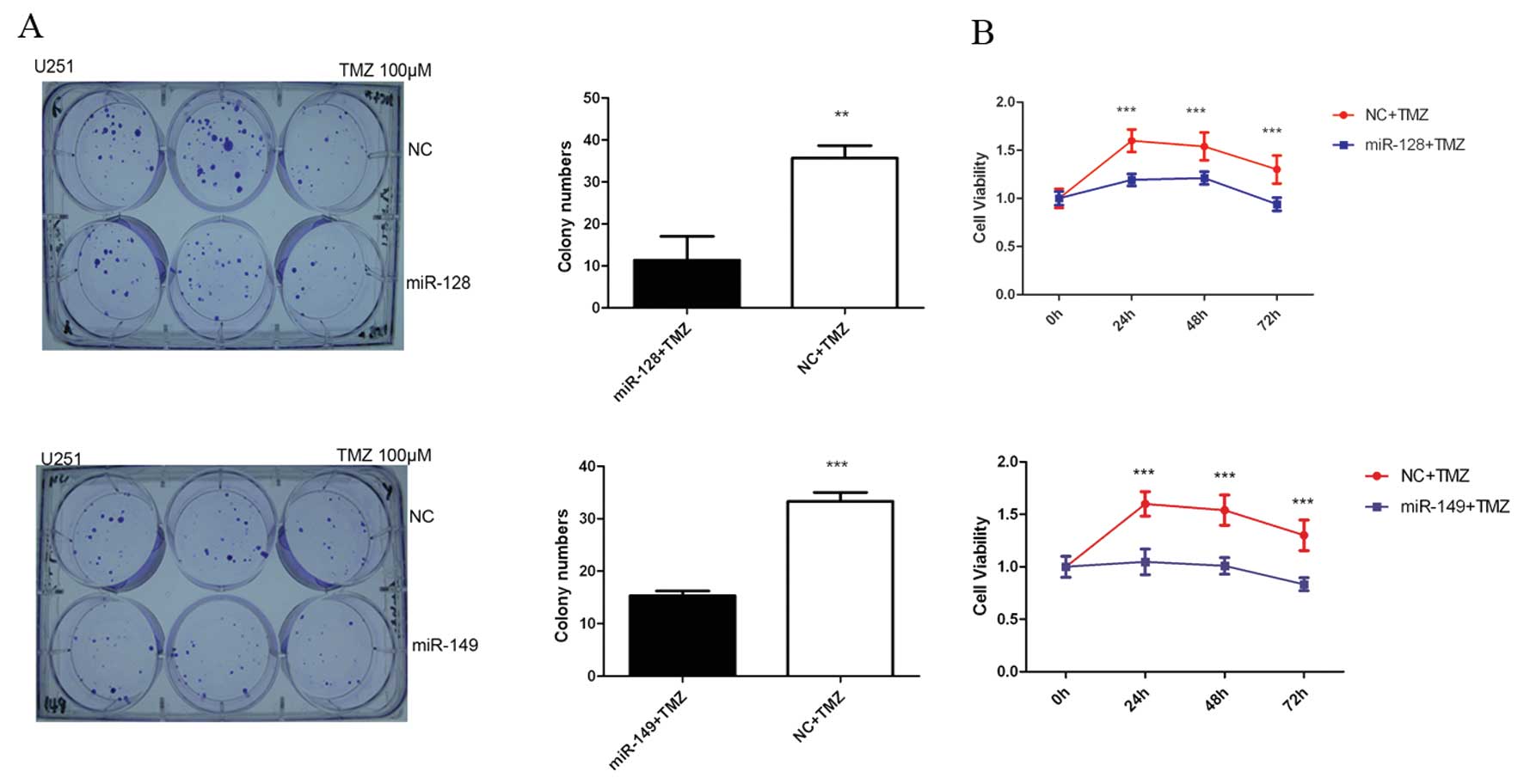
As shown in Fig. 4A,
compared with that of the negative control, miR-128 and miR-149
mimics inhibited cell colony formation in U251 cells (P<0.001).
MTT results indicated that miR-128 and miR-149 mimics also
inhibited the proliferation of U251 cells at 72 and 96 h
(P<0.001) (Fig. 4B). The
wound-healing assay showed that miR-128 and miR-149 inhibited the
migration and motility of U251 cells. Overexpression of miR-128 and
miR-149 using mimics clearly inhibited wound gap closure in a
time-dependent manner (Fig. 4C).
miR-128 and miR-149 mimics also inhibited the invasion of U251
cells at 48 h (P<0.001) (Fig.
4D). It has been demonstrated that TMZ can inhibit the
expression of Cdc42, RhoA and N-cadherin, and attenuate the
expression of F-actin and its polarized distribution in U251 cells
(18). In the present study, our
results also indicated that miR-128 and miR-149 mimics inhibited
the expression of Cdc42, RhoA and N-cadherin (Fig. 4E), suggesting that miR-128 and
miR-149 may be involved in cytoskeletal remodeling. FITC-labeled
phalloidin staining showed that the expression of F-actin-rich
cytoskeleton elements was strong and polarized at the edge of the
U251 cells. However, the expression of F-actin was attenuated, and
its polarized distribution died away when U251 cells were treated
with miR-128 or miR-149 mimics and Si-Rap1B (Fig. 4F).
miR-128 and miR-149 increase the
chemosensitivity of TMZ in glioblastoma cells
miR-128 and miR-149 inhibit the cellular
proliferation, invasion and alter cytoskeletal remodeling by
targeting inhibition of Rap1B. Therefore, we aimed to ascertain
whether miR-128 and miR-149 enhance the chemosensitivity of
temozolomide. The colony formation and MTT assays indicated that
miR-128 and miR-149 each enhanced the killing effect of TMZ on U251
cells when cells were administered miR-128 and miR-149 for 6 h,
followed by subsequent treatment with 100 μM TMZ (Fig. 5).
Discussion
GBM is characterized by high genetic heterogeneity
and infiltrative brain invasion patterns, and aberrant miRNA
expression has been associated with the hallmark malignant
properties of GBM (31). microRNAs
(miRNAs) are modulators of gene expression and play critical roles
in various cellular processes. Several miRNAs have been
characterized as tumor suppressors or oncogenes in cancer, and
recent studies suggest that certain miRNAs are implicated in
cytoskeletal remodeling and cell motility (32,33).
The cytoskeleton controls cell motility, invasion and adhesion.
miR-23b is able to directly inhibit a number of genes implicated in
cytoskeletal remodeling in breast cancer cells. Inhibition of
miR-23b, using a specific sponge construct, was found to lead to an
increase in cell migration and metastatic spread in vivo,
identifying it as a metastatic suppressor microRNA. Clinically, low
miR-23b expression correlates with the development of metastases in
breast cancer patients (32).
miR-30c was found to regulate invasion by targeting the
cytoskeleton network genes encoding Twinfilin 1 (TWF1) and vimentin
(VIM) in breast cancer cells (34).
In the present study, we assessed the endogenous expression of
miR-128 and miR-149 in 20 primary astrocytomas (grade I, 4 cases;
grade II, 6 cases; grade III, 5 cases; and glioblastoma, 5 cases)
and 8 normal brain tissues by quantitative real-time PCR. Our
results indicated that the expression of miR-128 and miR-149 was
downregulated in glioblastoma, and the expression of miR-149
gradually decreased with pathological progression in different
grade astrocytomas. However, the expression of miR-128 was not
different in WHO I, II, III grade astrocytomas and grade IV
glioblastoma although there was a significant different in normal
brain tissues and astrocytoma. These data revealed that aberrant
expression of miR-149 was involved in the initiation and the
malignant progression of astrocytomas, whereas, aberrant expression
of miR-128 was associated with the initiation of the
astrocytomas.
Our previous research found that Rap1B may also be a
negative regulator of miR-128 and miR-149 by a docking study of
miRNA/mRNA omics in human grade I–III astrocytomas (14). In the present study, we further
confirmed that miR-128 and miR-149 have their own binding sites
with Rap1B, and authenticated that Rap1B is a common target gene of
miR-128 and miR-149, which negatively regulate the expression of
Rap1B. RAP1 is a Ras-like GTPase that regulates several basic
cellular functions, such as migration, adhesion and growth. RAP1A
and RAP1B are two closely related isoforms of RAP1 (15). Rap1B, a small GTPase, is associated
with the cytoskeleton during cell activation (35). Its downregulated expression is
involved in the lysophosphatidic acid-induce inhibitory migration
in glioma cells (36). Rho family
GTPases, including Rac1, RhoA and cell division cycle 42 (Cdc42),
participate in the regulation of adhesion, actin cytoskeletal
reorganization and cell migration (37), and have been described as key
regulators of cell migration (38).
Several of the Rho GTPases shown to promote glioblastoma (GB) cell
invasion have also been correlated with poor patient prognosis
(39,40). We further elucidated the biological
function of miR-128 and miR-149. The results indicate that miR-128
and miR-149 mimics inhibited the proliferation and the invasion of
U251 cells and also mediated inhibition of expression of Cdc42,
RhoA and N-cadherin except for Rap1B. The overexpression of miR-128
and miR-149 or Rap1B siRNA could alter the F-actin cytoskeletal
reorganization and its expression level. These functional studies
suggest that miR-128 and miR-149 function as tumor-suppressor genes
by Rap1B-mediated changes in the actin cytoskeleton and inhibition
of cell adhesion and invasion and cell proliferation.
TMZ is the standard first-line agent for GBM. TMZ is
compromised by its dependence for activity on DNA mismatch repair
and the repair of the chemosensitive DNA lesion,
O6-methylguanine, by
O6-methylguanine-DNA-methyltransferase. The
present study found that miR-128 and miR-149 inhibit the cellular
proliferation, invasion and alter cytoskeletal remodeling by
targeted inhibition of Rap1B. We then presumed that miR-128 and
miR-149 enhance the chemosensitivity of temozolomide. Our data
confirmed that miR-128 and miR-149 did increase the cellular
inhibitory proliferation effects of TMZ in glioblastoma U251 cells
as determined by colony formation and MTT assays, suggesting that
the overexpression of miR-128 and miR-149 may enhance the
chemosensitivity of TMZ.
In summary, the present study demonstrated that the
expression of miR-128 and miR-149 was downregulated in
glioblastoma; the lower expression of miR-128 and miR-149
contributed to astrocytoma tumorigenesis. Overexpression of miR-128
and miR-149 inhibited the invasion and proliferation of
glioblastoma by targeting Rap1B-mediated cytoskeletal alteration
and related molecules, such as Cdc42, RhoA and N-cadherin. miR-128
and miR-149 enhanced the chemosensitivity of human glioblastoma
cells to TMZ. Therefore, restoration of miR-128 and miR-149
expression might provide a therapeutic strategy for GBM treatment.
The combination of miR-128 and miR-149 mimics and TMZ could be an
effective therapeutic strategy for suppressing the growth of
GBM.
Acknowledgements
The present study was supported by grants from the
National Science Foundation of China (81272297) and the Hunan
Province Natural Sciences Foundations of China (11JJ1013).
References
|
1
|
Pichiorri F, Palmieri D, De Luca L, et al:
In vivo NCL targeting affects breast cancer aggressiveness through
miRNA regulation. J Exp Med. 210:951–968. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pizzini S, Bisognin A, Mandruzzato S, et
al: Impact of microRNAs on regulatory networks and pathways in
human colorectal carcinogenesis and development of metastasis. BMC
Genomics. 14:5892013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yang J, Zhao H, Xin Y and Fan L:
MicroRNA-198 inhibits proliferation and induces apoptosis of lung
cancer cells via targeting FGFR1. J Cell Biochem. 115:987–995.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen L, Zhang Y, Yang J, Hagan JP and Li
M: Vertebrate animal models of glioma: understanding the mechanisms
and developing new therapies. Biochim Biophys Acta. 1836:158–165.
2013.PubMed/NCBI
|
|
5
|
Chou KN, Lin YC, Liu MY and Chang PY:
Temozolomide-related acute lymphoblastic leukemia with
translocation (4;11)(q21;q23) in a glioblastoma patient. J Clin
Neurosci. 21:701–704. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Danson SJ and Middleton MR: Temozolomide:
a novel oral alkylating agent. Expert Rev Anticancer Ther. 1:13–19.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Stupp R, Gander M, Leyvraz S and Newlands
E: Current and future developments in the use of temozolomide for
the treatment of brain tumours. Lancet Oncol. 2:552–560. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wong STS, Zhang XQ, Zhuang JTF, Chan HL,
Li CH and Leung GKK: MicroRNA-21 inhibition enhances in
vitro chemosensitivity of temozolomide-resistant glioblastoma
cells. Anticancer Res. 32:2835–2841. 2012.PubMed/NCBI
|
|
9
|
Shi L, Chen JA, Yang JA, Pan TH, Zhang SG
and Wang ZM: MiR-21 protected human glioblastoma U87MG cells from
chemotherapeutic drug temozolomide induced apoptosis by decreasing
Bax/Bcl-2 ratio and caspase-3 activity. Brain Res. 1352:255–264.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Qian XM, Ren Y, Shi ZD, et al:
Sequence-dependent synergistic inhibition of human glioma cell
lines by combined temozolomide and miR-21 inhibitor gene therapy.
Mol Pharm. 9:2636–2645. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ujifuku K, Mitsutake N, Takakura S, et al:
miR-195, miR-455–3p and miR-10a* are implicated in acquired
temozolomide resistance in glioblastoma multiforme cells. Cancer
Lett. 296:241–248. 2010.
|
|
12
|
Slaby O, Lakomy R, Fadrus P, et al:
MicroRNA-181 family predicts response to concomitant
chemoradiotherapy with temozolomide in glioblastoma patients.
Neoplasma. 57:264–269. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shi L, Zhang SG, Feng K, et al:
MicroRNA-125b-2 confers human glioblastoma stem cells resistance to
temozolomide through the mitochondrial pathway of apoptosis. Int J
Oncol. 40:119–129. 2012.PubMed/NCBI
|
|
14
|
Li D, Chen P, Li XY, et al: Grade-specific
expression profiles of miRNAs/mRNAs and docking study in human
grade I–III astrocytomas. OMICS. 15:673–682. 2011.PubMed/NCBI
|
|
15
|
Wittchen ES, Aghajanian A and Burridge K:
Isoform-specific differences between Rap1A and Rap1B GTPases in the
formation of endothelial cell junctions. Small GTPases. 2:65–76.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nuovo GJ: In situ detection of precursor
and mature microRNAs in paraffin embedded, formalin fixed tissues
and cell preparations. Methods. 44:39–46. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Evangelisti C, Florian MC, Massimi I, et
al: MiR-128 up-regulation inhibits Reelin and DCX expression and
reduces neuroblastoma cell motility and invasiveness. FASEB J.
23:4276–4287. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang Y, Chao T, Li R, et al: MicroRNA-128
inhibits glioma cells proliferation by targeting transcription
factor E2F3a. J Mol Med. 87:43–51. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Costa PM, Cardoso AL, Nobrega C, et al:
MicroRNA-21 silencing enhances the cytotoxic effect of the
antiangiogenic drug sunitinib in glioblastoma. Hum Mol Genet.
22:904–918. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Khan AP, Poisson LM, Bhat VB, et al:
Quantitative proteomic profiling of prostate cancer reveals a role
for miR-128 in prostate cancer. Mol Cell Proteomics. 9:298–312.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Weiss GJ, Bemis LT, Nakajima E, et al:
EGFR regulation by microRNA in lung cancer: correlation with
clinical response and survival to gefitinib and EGFR expression in
cell lines. Ann Oncol. 19:1053–1059. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kotani A, Ha D, Schotte D, den Boer ML,
Armstrong SA and Lodish HF: A novel mutation in the miR-128b gene
reduces miRNA processing and leads to glucocorticoid resistance of
MLL-AF4 acute lymphocytic leukemia cells. Cell Cycle. 9:1037–1042.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhu Y, Yu F, Jiao Y, et al: Reduced
miR-128 in breast tumor-initiating cells induces chemotherapeutic
resistance via Bmi-1 and ABCC5. Clin Cancer Res. 17:7105–7115.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shi ZM, Wang J, Yan Z, et al: miR-128
inhibits tumor growth and angiogenesis by targeting p70S6K1. PLoS
One. 7:e327092012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lin RJ, Lin YC and Yu AL:
miR-149* induces apoptosis by inhibiting Akt1 and E2F1
in human cancer cells. Mol Carcinog. 49:719–727. 2010.PubMed/NCBI
|
|
26
|
Jin L, Hu WL, Jiang CC, et al:
MicroRNA-149*, a p53-responsive microRNA, functions as
an oncogenic regulator in human melanoma. Proc Natl Acad Sci USA.
108:15840–15845. 2011.PubMed/NCBI
|
|
27
|
Wang Y, Zheng XS, Zhang ZY, et al:
MicroRNA-149 inhibits proliferation and cell cycle progression
through the targeting of ZBTB2 in human gastric cancer. PLoS One.
7:e416932012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tu HF, Liu CJ, Chang CL, et al: The
association between genetic polymorphism and the processing
efficiency of miR-149 affects the prognosis of patients with head
and neck squamous cell carcinoma. PLoS One. 7:e516062012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pan SJ, Zhan SK, Pei BG, Sun QF, Bian LG
and Sun BM: MicroRNA-149 inhibits proliferation and invasion of
glioma cells via blockade of AKT1 signaling. Int J Immunopathol
Pharmacol. 25:871–881. 2012.PubMed/NCBI
|
|
30
|
Tang HL, Wang ZY, Liu XP, et al: LRRC4
inhibits glioma cell growth and invasion through a
miR-185-dependent pathway. Curr Cancer Drug Targets. 12:1032–1042.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Guo Y, Yan KP, Fang JS, Qu Q, Zhou M and
Chen FH: Let-7b expression determines response to chemotherapy
through the regulation of cyclin D1 in glioblastoma. J Exp Clin
Cancer Res. 32:412013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pellegrino L, Stebbing J, Braga VM, et al:
miR-23b regulates cytoskeletal remodeling, motility and metastasis
by directly targeting multiple transcripts. Nucleic Acids Res.
41:5400–5412. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Avellino R, Carrella S, Pirozzi M, et al:
miR-204 targeting of Ankrd13A controls both mesenchymal neural
crest and lens cell migration. PLoS One. 8:e610992013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bockhorn J, Yee K, Chang YF, et al:
MicroRNA-30c targets cytoskeleton genes involved in breast cancer
cell invasion. Breast Cancer Res Treat. 137:373–382. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bertoni A, Tadokoro S, Eto K, et al:
Relationships between Rap1b, affinity modulation of integrin
αIIbβ3, and the actin cytoskeleton. J Biol Chem. 277:25715–25721.
2002.PubMed/NCBI
|
|
36
|
Malchinkhuu E, Sato K, Maehama T, et al:
Role of Rap1B and tumor suppressor PTEN in the negative regulation
of lysophosphatidic acid-induced migration by isoproterenol in
glioma cells. Mol Biol Cell. 20:5156–5165. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Vega FM and Ridley AJ: Rho GTPases in
cancer cell biology. FEBS Lett. 582:2093–2101. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
de Toledo M, Anguille C, Roger L, Roux P
and Gadea G: Cooperative anti-invasive effect of Cdc42/Rac1
activation and ROCK inhibition in SW620 colorectal cancer cells
with elevated blebbing activity. PLoS One. 7:e483442012.PubMed/NCBI
|
|
39
|
Feng H, Hu B, Liu KW, et al: Activation of
Rac1 by Src-dependent phosphorylation of Dock180Y1811
mediates PDGFRα-stimulated glioma tumorigenesis in mice and humans.
J Clin Invest. 121:4670–4684. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fortin Ensign SP, Mathews IT, Eschbacher
JM, Loftus JC, Symons MH and Tran NL: The Src homology 3
domain-containing guanine nucleotide exchange factor is
overexpressed in high-grade gliomas and promotes tumor necrosis
factor-like weak inducer of apoptosis-fibroblast growth
factor-inducible 14-induced cell migration and invasion via tumor
necrosis factor receptor-associated factor 2. J Biol Chem.
288:21887–21897. 2013.
|