Introduction
Prostate cancer is the most commonly diagnosed
cancer and the second leading cause of death in males (1,2).
Although substantial progress in treatment has been achieved,
therapies for prostate cancer are far from satisfactory, the reason
predominantly being the propensity of these tumors to metastasize.
Efforts to identify new genes and signaling pathways that promote
tumor progression have revealed that the oxidored-nitro domain
containing protein 1 (NOR1) gene (also known as OSCP1) plays an
important role in prostate cancer as well other cancer types
(3–5). Evidence suggests that the expression
of NOR1 is significantly decreased in prostate cancer cells
(3); however, the specific function
of NOR1 in the progression of prostate cancer remains to be
elucidated.
NOR1, a novel tumor-suppressor gene, was first
isolated from a nasopharyngeal carcinoma (6). The human NOR1 gene (chromosome 1 open
reading frame 102) is located on 1p34.3 (7) and has nine introns and 10 exons
according to the human genome working sequence (8). Moreover, data from the PROSITE
database suggest that the NOR1 gene has four N-myristoylation
sites, two tyrosine phosphorylation sites and cAMP and
cGMP-dependent protein kinase phosphorylation sites (8). NOR1 is expressed in many tissues and
cells, and more recently, it was shown to restrict expression in
the testis and neurons. Thus, this gene has been identified as a
potential cancer/testis/brain antigen (3). NOR1 is usually highly expressed in
many tissues, such as brain, testis, the upper respiratory tract
(such as the nasopharynx), but more weakly expressed in many types
of cancer tissues, such as nasopharyngeal carcinoma, lung, ovarian,
vulvar, kidney, leukemia, uterus, testis, thyroid gland, cervical
and prostate (5,7,9,10).
Cancer cell proliferation and apoptosis are
important in tumor progression. Reagents or molecules that can
inhibit cancer cell proliferation and induce apoptosis may suppress
tumor progression. Apoptosis, a form of programmed cell death, is
regulated by the Bcl-2 family and caspase family of proteins
(11). Evidence suggests that
genes, such as p53 (12,13), caspase-3 (14), caspase-9 (13), Bcl-2 (15), Bcl-xl (15) and Bax (16) regulate cell apoptosis. Ouyang and
colleagues (5) found that the
overexpression of NOR1 inhibits human nasopharyngeal carcinoma and
cervical cancer cell proliferation and induces apoptosis. Authors
of that group also showed that NOR1 overexpression can activate
caspase-9, increase pro-apoptotic Bax and p53 expression and
decrease anti-apoptotic Bcl-2 expression (5). Although NOR1 has many positive effects
in many types of cancer, the function and signaling mechanism of
NOR1 overexpression in prostate cancer cells has not been defined.
Therefore, it was hypothesized that NOR1 overexpression may inhibit
the growth of prostate cancer cells and induce cell apoptosis. In
the present study, we demonstrated that NOR1 overexpression
activates caspase-3 and −9 and increases the expression of Bax and
Bak, while decreasing the level of Bcl-2 and Bcl-xl in DU145 and
PC3 prostate cancer cells. To the best of our knowledge, the study
provides the first direct evidence in prostate cancer cells that
NOR1 overexpression can lead to cell growth inhibition and
apoptosis induction by altering the expression of apoptosis-related
genes through the MAPK signaling pathway.
Materials and methods
Animals
Procedures were carried out according to the
protocols approved by the Ethics Committee for Animal
Experimentation of the People’s Hospital of Zhengzhou (Zhengzhou,
China). All efforts were made to minimize the suffering of the
animals. MF-1 nude mice (6–8 weeks old) were obtained from the
Animal Center of Zhengzhou University, Zhengzhou, China.
Antibodies
Rabbit Anti-NOR1 polyclonal antibody, and mouse
anti-caspase-3, mouse anti-Bcl-2, mouse anti-Bcl-xl, mouse
anti-Bax, mouse anti-Bak, and mouse anti-β-actin monoclonal
antibodies, as well as HRP-conjugated rabbit anti-mouse IgG and
HRP-conjugated goat anti-rabbit IgG were purchased from Abcam
(Cambridge, MA, USA).
Cell culture
The DU145 (HTB-4) and PC3 human prostate cancer cell
lines were obtained from the American Type Cell Collection (ATCC;
Manassas, VA, USA). The cells were cultured in RPMI-1640 medium
(Gibco-BRL, Gaithersburg, MD, USA) supplemented with 10% fetal
bovine serum (FBS; Gibco-BRL), 1% penicillin-streptomycin and 1%
glutamine at 37°C under a humidified atmosphere of 5%
CO2.
Clone and expression vectors construction
of NOR1
pcDNA3.1-myc-his-NOR1 was constructed as previously
described (5). Briefly, total RNA
was extracted from the DU145 and PC3 prostate cancer cell lines
using TRIzol (Invitrogen, Carlsbad, CA, USA) according to the
manufacturer’s instructions. RNA (2 μg) was reverse transcribed
into cDNA using the PrimeScript RT reagent kit (Takara, Shiga,
Japan). The cDNAs were used for the amplification of NOR1 using two
primers (17): sense
5′-GTGCTGCTGGTCACTTTCAA-3′ and antisense
5′-GGATACTCGGATGTGCAGGT-3′. pcDNA3.1-myc-his-NOR1 was generated
using the ViraPower Adenoviral Expression System (Invitrogen)
according to the operating protocols. The transfection experiment
was performed with pcDNA3.1-myc-his-NOR1 (NOR1) or the control
plasmid (control) using Lipofectamine 2000 according to the
manufacturer’s instructions.
Cell viability assay
The viability of the DU145 and PC3 human prostate
cancer cell lines was determined using the trypan blue exclusion
assay, as previously described (18). Briefly, transfected and control
cells were seeded in 96-well plates at 1×106 cells/well
and cultured in RPMI-1640 for different periods of time (0, 24, 48,
72 and 96 h). Trypan blue (40 μl; Sigma, St. Louis, MO, USA) was
added to cell suspensions (10 μl) in PBS. The cells were then
washed three times with DMSO, followed by counting under a light
microscope.
MTT proliferation assay
DU145 and PC3 cell proliferation was detected by the
previously described
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay (19). Briefly, the cells
were transfected as described above, and then plated in 96-well
plates. The cells were cultured for 72 h, then 20 μl MTT (5 mg/ml)
was added to the cancer cells for 4 h at 37°C. DMSO (200 μl) was
added to solubilize the crystals at room temperature. The optical
density (OD) was measured at 490 nm by a spectrophotometer
(Multiskan MK3; Thermo Fisher Scientific, Waltham, MA, USA). Data
were calculated using averaged results.
Detection of apoptotic cells by flow
cytometry
To analyze apoptotic PC3 cells by Annexin V-FITC and
propidium iodide (PI) staining by flow cytometry (Beckman-Coulter,
Brea, CA, USA), 1×104 cells/well were plated in 96-well
plates. The cells were trypsinized and harvested by centrifugation.
Furthermore, the cells were incubated with Annexin V and PI for 20
min at room temperature. Apoptotic cells were then analyzed by flow
cytometry (Beckton-Dickinson, Franklin Lakes, NJ, USA).
Nuclear morphological observation of PC3
cancer cells
PC3 cancer cells were plated in 96-well plates at
1×105 cells/well. The cells were fixed using 4%
formaldehyde, and washed twice with PBS. Subsequently, the cells
were stained with Hoechst 33258 staining solution using the Hoechst
Staining kit according to the manufacturer’s instructions (20,21).
Stained nuclei of PC3 cells were observed under a reflected
fluorescence microscope (Nikon MF30 LED; Nikon, Tokyo, Japan).
Total RNA extraction and quantitative
reverse transcription-PCR
NOR1 mRNA levels were measured in the PC3 prostate
cancer cell line according to a previously described method, with
minor modifications (22). Total
RNA was isolated from cells using TRIzol, according to the
manufacturer’s instructions. RNA was reverse transcribed using a
high capacity RNA-to-cDNA kit (Applied Biosystems). RT-qPCR was
performed using an ABI PRISM 7000 Sequence Detection system
(Applied Biosystems) with SYBR-Green ReadyMix (Applied Biosystems).
The primer sequences used for RT-qPCR were: NOR1 forward
5′-ACCTGCACATCCGAGTATCC-3′ and reverse 5′-CTGGCCAAGAAATTCAGCTC3′;
β-actin forward 5′-TCCCTGGAGAAGAGCTACGA-3′ and reverse
5′-AGGAAGGAAGGCTGGAAGAG-3′. The primers were purchased from Sangon
Biotech Co., Ltd. (Shanghai, China). Expression levels of the
relative genes were calculated using the 2−ΔΔCT method
(23). β-actin mRNA was used as an
internal control.
Western blot assay
The protein expression in PC3 cells was determined
according to a previously described method, with minor
modifications (24). To obtain
whole-cell lysates, the cells were resuspended in ice-cold lysis
buffer [1% Nonidet P-40, 0.1% SDS, 0.1% sodium deoxycholate, 150 mM
NaCl, 1 mM EDTA, 10 mM Tris-HCl (pH 7.4)]. The protein
concentration in the lysates was determined using a BCA protein
assay kit (Beyotime Institute of Biotechnology, Nantong, China).
The proteins (40 μg/lane) were subjected to electrophoresis on 10%
SDS-polyacrylamide gels and electrophoretically transferred to
Immobilon-P membranes (Millipore, Bedford, MA, USA). After blocking
with 5% skim milk, the membranes were probed with mouse monoclonal
and rabbit polyclonal antibodies followed by HRP-conjugated rabbit
anti-mouse IgG or HRP-conjugated goat anti-rabbit IgG. Bound
proteins were detected using the ECL reagent (Boehringer Mannheim,
Mannheim, Germany).
Small interfering RNA transfection
PC3 prostate cancer cells with the NOR1 protein were
transfected with NOR1 siRNA (siNOR1) or the control siRNA (siMock)
using Lipofectamine 2000 (Invitrogen). The NOR1 siRNA
oligonucleotides used were: 5′-GGCCAGCATGGATAAGCTCTA-3′; and siRNA
scrambled: 5′-GGATAAAGCGTGCCCGTACTA-3′. Stable transfectants of PC3
cells were selected with culture medium containing puromycin
(Invitrogen). Viable colonies were picked up and transferred to new
dishes 2–3 weeks after transfection. siNOR1- and siMock-transfected
cells were used for subsequent experiments.
Prostate cancer model
Male MF-1 nude mice (6–8 weeks old) were housed
under pathogen-free conditions. PC-3 cancer cells
(1×106) infected with a control or NOR1-overproducing
vector were implanted subcutaneously in 6- to 8-week-old male nude
mice, respectively. Tumor growth was monitored regularly using
calipers. The tumor volume was calculated as follows: (larger
diameter) × (smaller diameter) × (width) (25).
Statistical analysis
Data are expressed as means ± SEM. The differences
between the control and experimental groups were compared by the
Dunnett’s test. P<0.05 was considered statistically significant.
All experiments were repeated at least three times.
Results
Increased NOR1 levels in DU145 and PC3
prostate cancer cell lines transduced with
pcDNA3.1-myc-his-NOR1
To overexpress NOR1 in the DU145 and PC3 prostate
cancer cell lines, we used a eukaryotic expression vector,
pcDNA3.1-myc-his, which encodes NOR1 under normal conditions. The
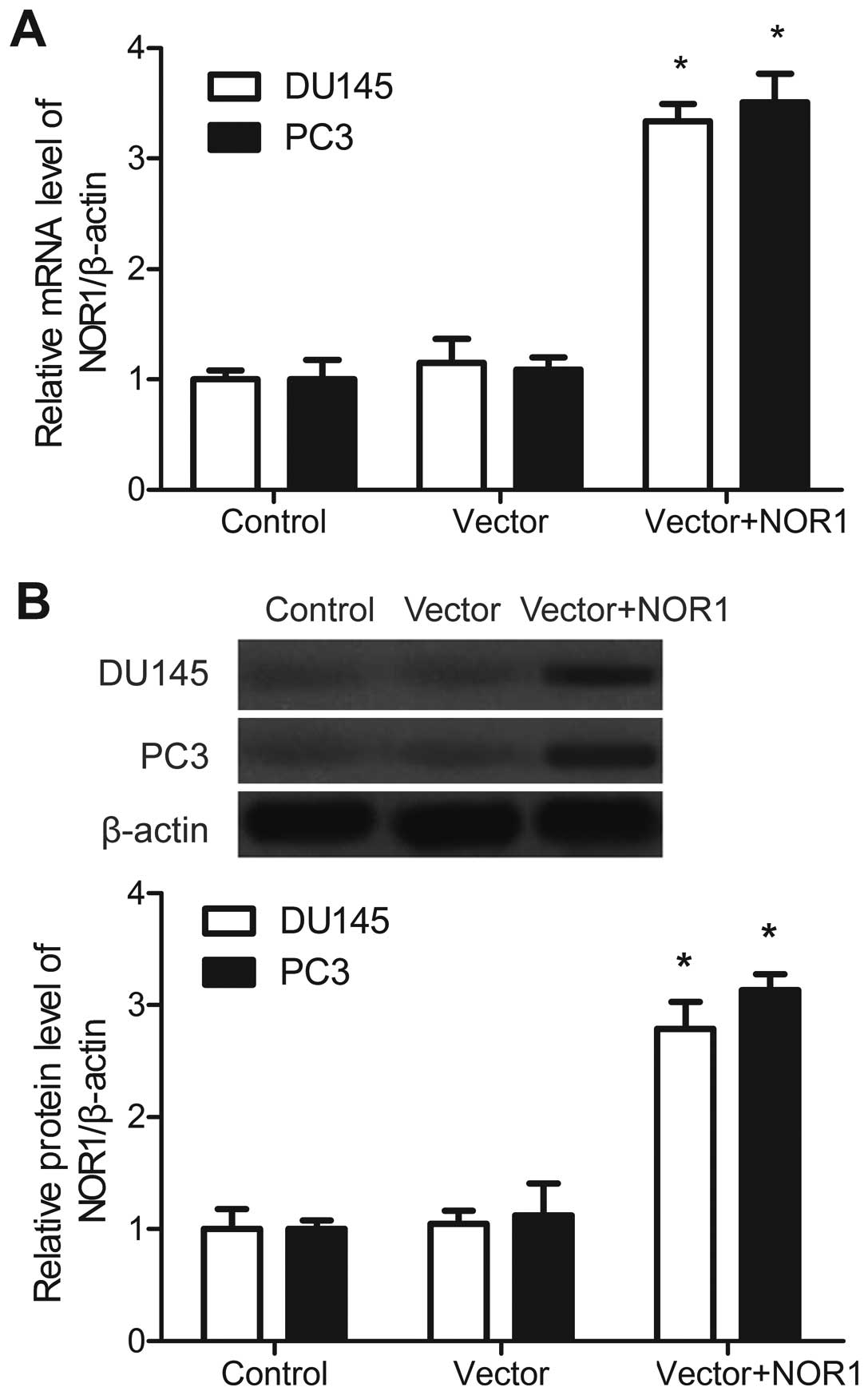
expression of NOR1 in prostate cancer cells was determined by
RT-qPCR and western blot analysis. The RT-qPCR results (Fig. 1A) showed that NOR1 mRNA levels in
DU145 and PC3 cells transduced with pcDNA3.1-myc-his-NOR1 for 72 h
were much higher than in the control group. The protein results
indicated that NOR1 protein levels were higher in DU145 and PC3
cells transduced with pcDNA3.1-myc-his-NOR1 for 72 h (Fig. 1B). The mRNA and protein expression
was also upregulated in DU145 and PC3 transduced with
pcDNA3.1-myc-his-NOR1 for 24, 48 and 96 h (data not shown).
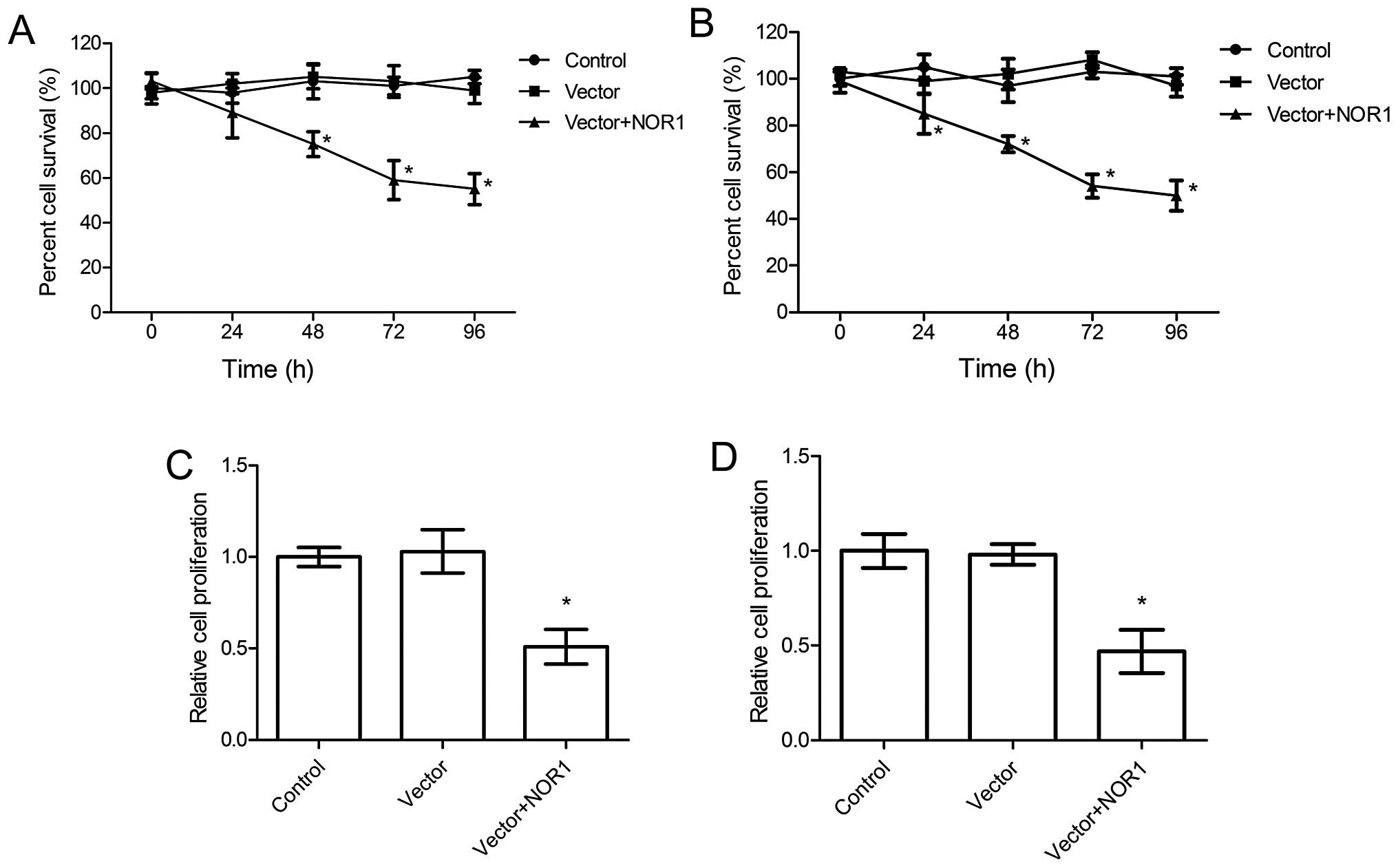
Effect of NOR1 overexpression on DU145
and PC3 cell viability and proliferation
The cytotoxic effect of NOR1 overexpression on DU145
and PC3 cells was determined by the trypan blue exclusion assay.
The results showed that the viability of DU145 and PC3 cells was
significantly reduced after NOR1 overexpression in a time-dependent
manner (Fig. 2A). We also measured
the effect of NOR1 overexpression on DU145 and PC3 cell
proliferation using an MTT assay. As shown in Fig. 2B, there was a marked decrease in the
proliferation of DU145 and PC3 cells with NOR1 overexpression. We
found that NOR1 overexpression led to an almost 49 and 53% decrease
in DU145 and PC3 cell numbers, respectively. This finding indicated
a positive effect of NOR1 on the inhibition of proliferation and
survival of DU145 and PC3 cell. Since PC3 cells were more sensitive
to NOR1 overexpression, this cell line was selected for subsequent
experiments.
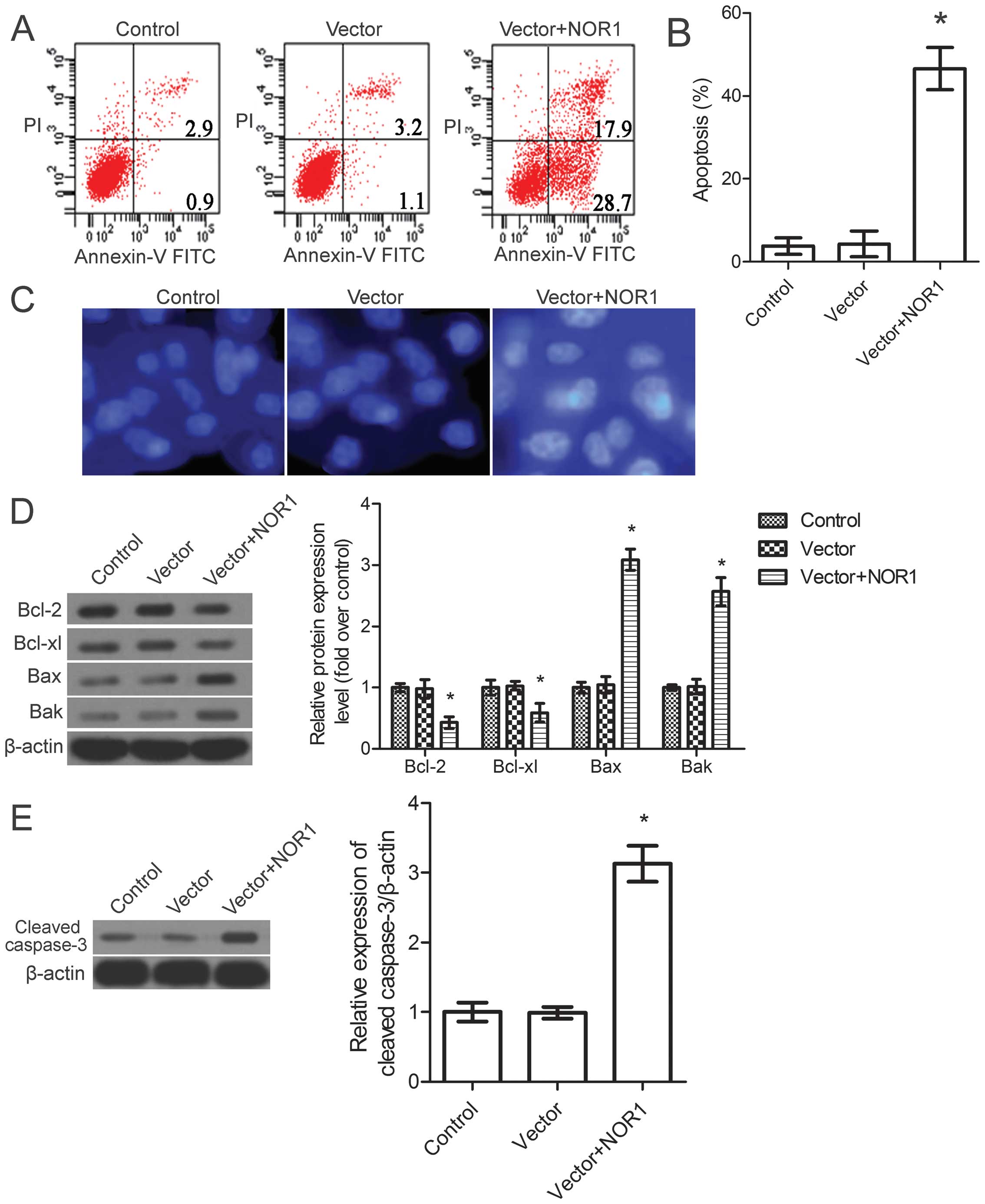
Effect of NOR1 overexpression on PC3 cell
apoptosis
Evidence suggests that NOR1 overexpression can
induce human nasopharyngeal carcinoma and cervical cancer cell
apoptosis (5). Thus, we tested
whether the change in PC3 cell numbers in our experiments was also
mediated by NOR1-induced apoptosis. Apoptosis was first measured by
the flow cytometric analysis of PC3 cells double-stained with
Annexin V and PI. The results suggested that NOR1 overexpression
induced cell apoptosis, and the results showed a significant
increase in the percentage of early (Annexin V-positive and
PI-negative) and late (Annexin V-negative and PI-positive)
apoptosis. The total apoptotic rate increased by 46% when cells
were transduced with the NOR1 vector for 72 h (Fig. 3A and B).
To confirm that NOR1 overexpression induces
apoptosis in PC3 cells, an additional apoptotic marker, Hoechst
33258 staining, was evaluated (26). Membrane-permeable Hoechst 33258 is a
blue fluorescent dye that stains the cell nucleus (27). Thus, we measured the changes in cell
nuclear morphology by Hoechst staining using fluorescence
microscopy. As shown in Fig. 3C,
the control and vector groups of PC3 cells showed uniformly light
blue nuclei under the fluorescent microscope. The NOR1 cells showed
smaller nuclei and condensed staining because of karyopyknosis and
chromatin condensation.
Given the profound roles of Bcl-2 and the caspase
family in triggering apoptosis, the anti-apoptotic proteins Bcl-2
and Bcl-xl and the pro-apoptotic proteins Bax and Bak, as well as
the activity of caspase-3 in NOR1 overexpression-mediated apoptosis
in PC3 cells were evaluated. The western blot results revealed that
the expression of Bcl-2 and Bcl-xl was decreased, whereas the
expression of Bax and Bak was simultaneously upregulated (Fig. 3D; P<0.05). Moreover, NOR1
overexpression showed a marked increase in the activity of
caspase-3 (Fig. 3E; P<0.05).
Thus, it appears that, in the PC3 prostate cancer cell line,
apoptosis was induced by NOR1 overexpression through a
caspase-dependent pathway.
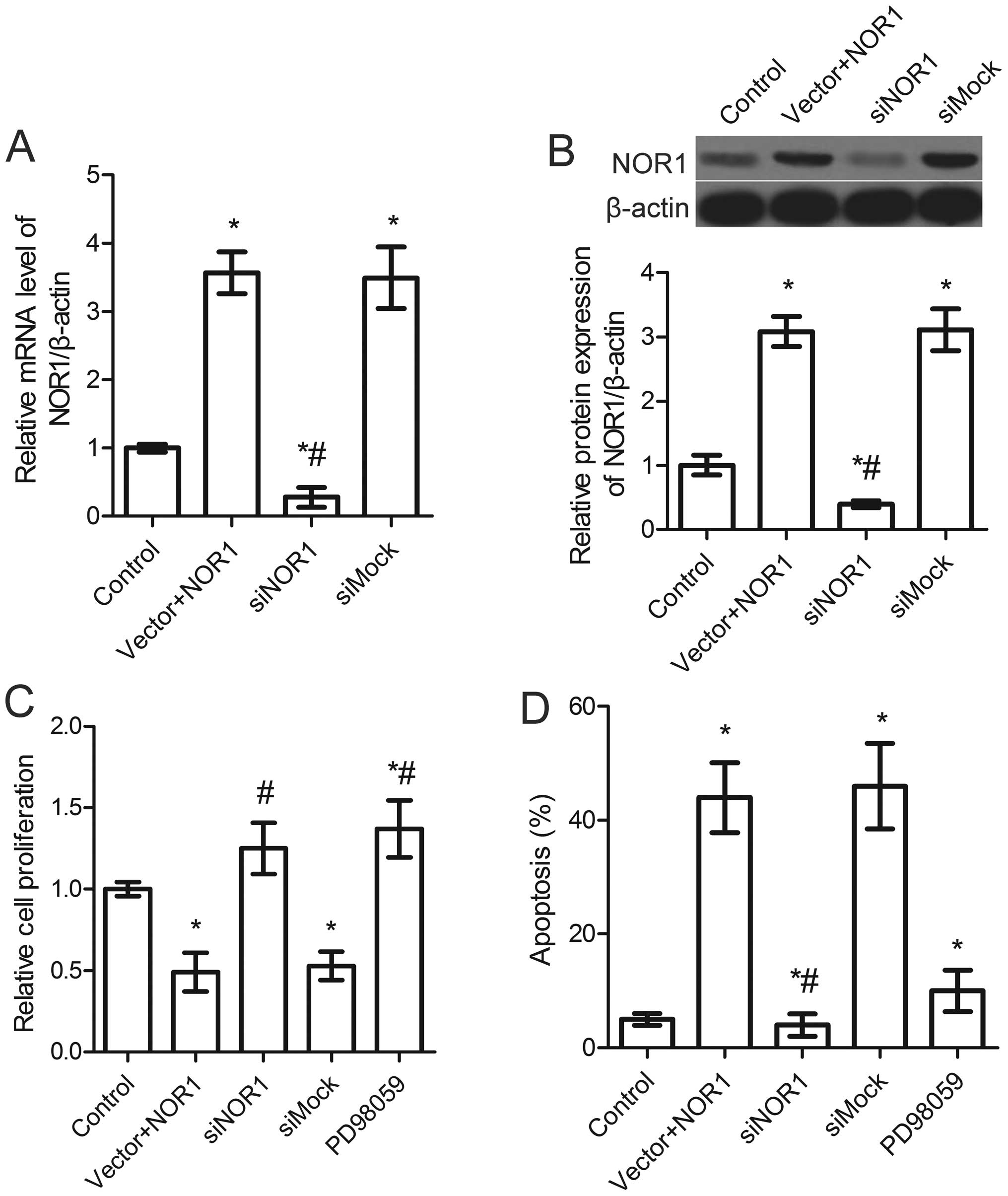
Specific inhibition of NOR1 mRNA and
protein expression by NOR1-specific siRNA
The above results were designed to reveal the main
effects of overexpressing NOR1 in prostate cancer cells. To detect
the effects of inhibiting the expression of NOR1 in
NOR1-overexpressing cells, a complementary approach was employed
using the RNA interference method. PC3 cells were transfected with
NOR1 siRNA (siNOR1) or control siRNA (siMock) and analyzed after 48
h of growth. The RT-qRCR and western blot results showed that the
level of NOR1 in cells transfected with siNOR1 was significantly
decreased (Fig. 4A and B). The
effect of NOR1 silencing on cell proliferation and apoptosis was
determined. The results shown in Fig.
4C indicate that there was a significant increase in PC3 cell
growth in the siNOR1-transfected group compared with the
siMock-transfected group. Moreover, cell apoptosis was also
markedly decreased in the siNOR1-transfected group (Fig. 4D and E). These results showed that
growth inhibition and apoptosis in PC3 cells is associated with the
overexpression of NOR1.
Overexpression of NOR1 is directly
related to the activation of MAPK
An important role for the p38 MAPK pathway in
apoptosis had been described in many cell types, including prostate
cancer cells (28–30). Moreover, Gui et al (8) found that the NOR1 gene can activate
the MAPK signaling pathway. Therefore, to determine a potential
mechanism that might be involved in the pro-apoptotic and growth
inhibition effect of NOR1 in prostate cancer cells, the effect of
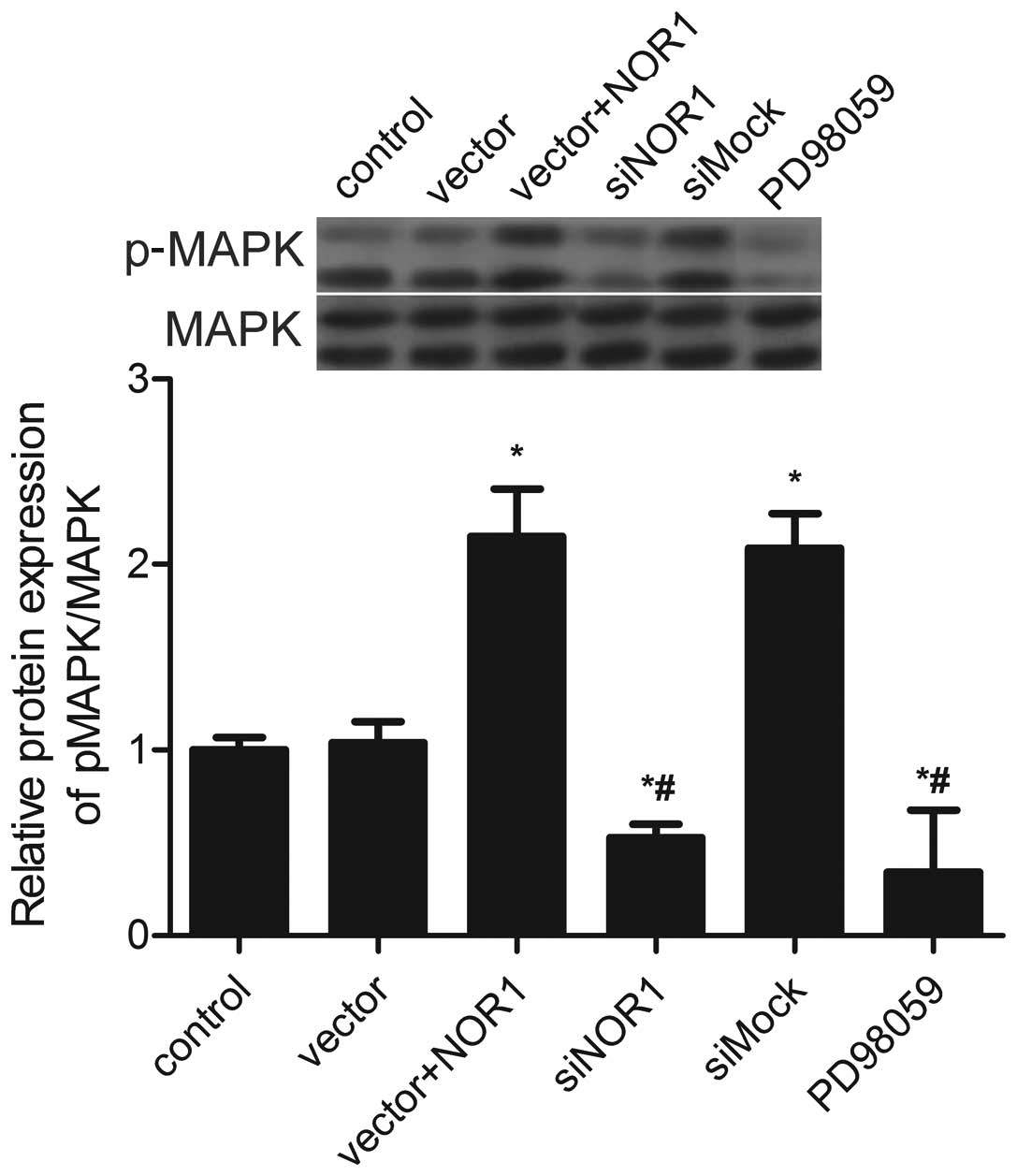
NOR1 overexpression on the level of MAPK and phosphorylated MAPK
(pMAPK) was assessed. The results showed that pMAPK was
significantly increased in NOR1-overexpressing cells and decreased
in siNOR1-transfected cells (Fig.
5). This result showed that the MAPK pathway is inhibited in
siNOR1-transfected cells.
To confirm whether MAPK mediates NOR1
overexpression-induced cell apoptosis, cells were treated with the
MAPK inhibitor PD98059. As shown in Fig. 5, PD98059 inhibited NOR1
overexpression-induced MAPK activation. Apoptosis and proliferation
in PC3 cells were determined and the results showed a marked
increase in PC3 cell growth and a significant decrease in apoptosis
in cells treated with PD98059 compared with the NOR1-overexpressing
group (Fig. 4D).
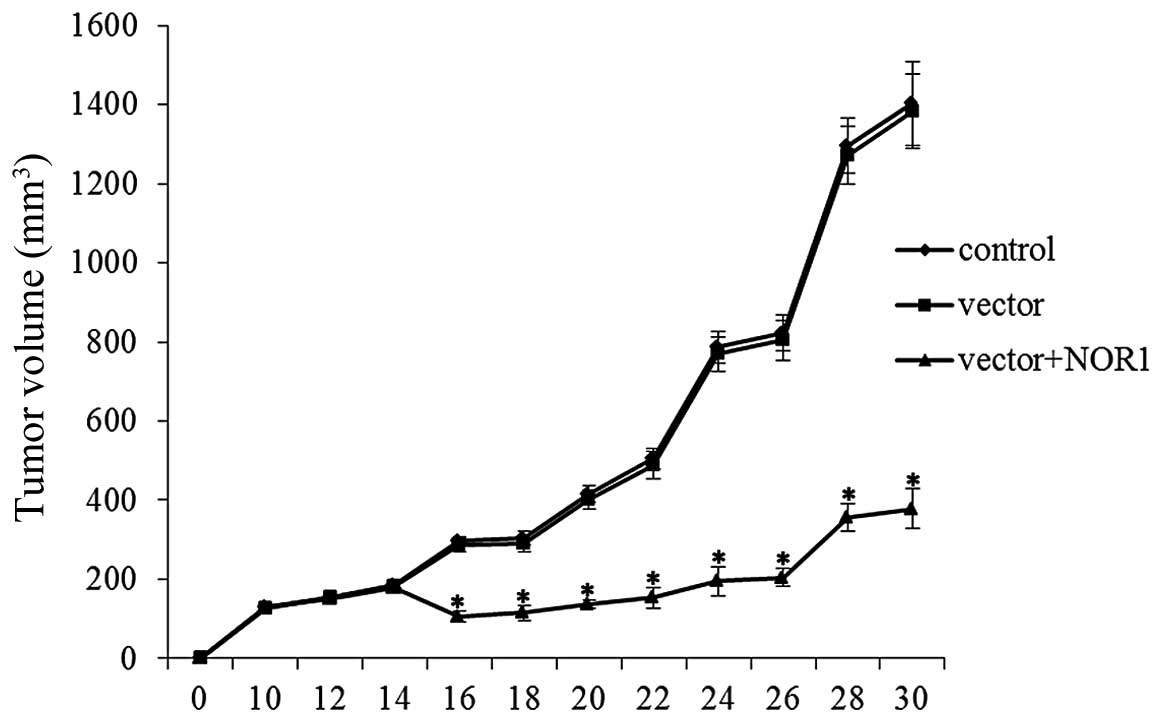
Effect of NOR1 overexpression on prostate
cancer growth
To examine whether NOR1 overexpression would induce
efficient antitumor effects, normal PC3 cells, control
transfectants (vector) and NOR1-overproducing cells (vector+NOR1)
were injected into the flanks of nude mice, and the tumor size was
measured at frequent intervals (Fig.
6). The results showed that NOR1 overexpression significantly
inhibited tumor growth in vivo. This result showed that NOR1
overexpression induces a marked antitumor effect.
Discussion
The main findings of the present study demonstrate
that: i) NOR1 overexpression can induce growth inhibition and
apoptosis in prostate cancer cells; ii) the anti-apoptotic effect
of NOR1 is apparently involved in regulating the expression of
apoptosis-related genes, i.e., caspase-3, Bcl-2, Bcl-xl, Bax and
Bak; iii) NOR1 silencing has no apparent effect on PC3 cell growth
and apoptosis; iv) NOR1 overexpression can activate the MAPK
pathway; and v) NOR1 overexpression induces stronger antitumor
effects. To the best of our knowledge, these data demonstrate for
the first time that NOR1 can inhibit the progression of prostate
cancer in vitro and in vivo, which can be explained
by growth inhibition and pro-apoptotic effects of NOR1 in prostate
cancer cells.
In the present study, we analyzed the effects of
NOR1 overexpression on prostate cancer cell proliferation and
apoptosis. The human NOR1 gene, located on 1p34.3 (7), is a novel tumor-suppressor gene.
Evidence suggests that the NOR1 promoter region is always
methylated in nasopharyngeal carcinoma and leukemia, leading to
lower NOR1 expression in these cancer cells (4,7).
Moreover, some studies suggest that NOR1 is downregulated in
prostate cancer cells compared with normal prostate tissue
(5). To determine the antitumor
effects of NOR1, we constructed a recombinant expression vector
(pcDNA3.1-myc-his-NOR1) and transfected the vector into DU145 and
PC3 cells. The RT-qPCR and western blot results indicate that NOR1
expression is significantly upregulated in these two types of cells
(Fig. 1). In the present study, the
overexpression of NOR1 in prostate cancer cells induced cell growth
inhibition and apoptosis. We found that a high level of NOR1 was
associated with a decrease in cell proliferation and an increase in
cell apoptosis (Figs. 2 and
3). Simultaneously, we found that
NOR1 siRNA increased cell survival and decreased apoptosis
(Fig. 4). These data suggest that
the upregulation of NOR1 expression was able to inhibit the
progression of prostate cancer.
Apoptosis, which is also known as programmed cell
death, is an evolutionarily conserved cell suicide process executed
by caspases and regulated by the Bcl-2 protein family (31,32).
In the present study, we found that NOR1 overexpression can
downregulate the anti-apoptotic genes Bcl-2 and Bcl-xl and
upregulate the pro-apoptotic genes Bax and Bak (Fig. 3). Moreover, we found that NOR1
overexpression induced a marked increase in the activity of
caspase-3 (Fig. 3). Therefore,
these results show that NOR1 overexpression-induced PC3 prostate
cancer cell line apoptosis is dependent on the caspase pathway.
Further investigation suggested that the mechanism of NOR1
overexpression-induced growth inhibition and apoptosis occurred
through the MAPK signaling pathway. NOR1 overexpression upregulated
the level of pMAPK, whereas suppression of the expression of NOR1
inhibited the phosphorylation of MAPK (Fig. 5). These results show that the
expression of NOR1 activated the MAPK signaling pathway. The
present study demonstrates that the MAPK inhibitor PD98059
attenuated the action of NOR1 overexpression-induced cell growth
inhibition and apoptosis (Fig. 4).
We also found that NOR1 overexpression induced strong antitumor
effects in a prostate cancer model.
In summary, our findings have shown that the
overexpression of NOR1 contributes to the inhibition of prostate
cancer. Overexpression of NOR1 inhibits the proliferation of
prostate cancer cells and promotes cell apoptosis. Furthermore, our
study suggests that the activity of caspase-3, and the expression
of Bcl-2, Bcl-xl, Bax and Bak are also regulated by NOR1, thus,
modulating apoptosis in PC3 cells. The present study also suggests
that NOR1 modulates the proliferation and apoptosis of PC3 cells
through the MAPK signaling pathway. Additionally, NOR1
overexpression induced strong antitumor effects in a prostate
cancer model. These findings support the hypothesis that increasing
NOR1 expression may be an ideal therapeutic approach for the
treatment of prostate cancer.
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar
|
|
2
|
Rasheed SAK, Teo CR, Beillard EJ,
Voorhoeve PM and Casey PJ: MicroRNA-182 and microRNA-200a control
G-protein subunit α-13 (GNA13) expression and cell invasion
synergistically in prostate cancer cells. J Biol Chem.
288:7986–7995. 2013.PubMed/NCBI
|
|
3
|
Xiang B, Wang W, Li W, Li X, Li X and Li
G: Differential expression of oxidored nitro domain containing
protein 1 (NOR1), in mouse tissues and in normal and cancerous
human tissues. Gene. 493:18–26. 2012. View Article : Google Scholar
|
|
4
|
Li W, Li X, Wang W, et al: NOR1 is an
HSF1-and NRF1-regulated putative tumor suppressor inactivated by
promoter hypermethylation in nasopharyngeal carcinoma.
Carcinogenesis. 32:1305–1314. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ouyang J, Wu M, Huang C, Cao L and Li G:
Overexpression of oxidored-nitro domain containing protein 1
inhibits human nasopharyngeal carcinoma and cervical cancer cell
proliferation and induces apoptosis: Involvement of mitochondrial
apoptotic pathways. Oncol Rep. 29:79–86. 2013.
|
|
6
|
Nie X, Zhang B, Li X, et al: Cloning,
expression, and mutation analysis of NOR1, a novel human gene
down-regulated in HNE1 nasopharyngeal carcinoma cell line. J Cancer
Res Clin Oncol. 129:410–414. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kroeger H, Jelinek J, Estécio MR, et al:
Aberrant CpG island methylation in acute myeloid leukemia is
accentuated at relapse. Blood. 112:1366–1373. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gui R, Liu J, Li D, Tang H, Feng Z and Nie
X: NOR1 enhanced CB1954 induced cell cytotoxicity in HepG2 is
dependent on Grb2 mediated MAPK pathway. In: Bioinformatics and
Biomedical Engineering (iCBBE), 2010 4th International Conference
on IEEE; pp. 1–7. 2010
|
|
9
|
Shen L, Kantarjian H, Guo Y, et al: DNA
methylation predicts survival and response to therapy in patients
with myelodysplastic syndromes. J Clin Oncol. 28:605–613. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jelinek J, Gharibyan V, Estecio MR, et al:
Aberrant DNA methylation is associated with disease progression,
resistance to imatinib and shortened survival in chronic
myelogenous leukemia. PloS One. 6:e221102011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Brentnall M, Rodriguez-Menacol L, De
Guevara RL, Cepero E and Boise LH: Caspase-9, caspase-3 and
caspase-7 have distinct roles during intrinsic apoptosis. BMC Cell
Biol. 14:322013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kuribayashi K, Finnberg NK, Jeffers JR,
Zambetti GP and El-Deiry WS: The relative contribution of
pro-apoptotic p53-target genes in the triggering of apoptosis
following DNA damage in vitro and in vivo. Cell Cycle.
10:2380–2389. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
D’Sa-Eipper C, Leonard JR, Putcha G, et
al: DNA damage-induced neural precursor cell apoptosis requires p53
and caspase 9 but neither Bax nor caspase 3. Development.
128:137–146. 2001.PubMed/NCBI
|
|
14
|
Porter AG and Jänicke RU: Emerging roles
of caspase-3 in apoptosis. Cell Death Differ. 6:99–104. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Boucher MJ, Morisset J, Vachon PH, Reed
JC, Lainé J and Rivard N: MEK/ERK signaling pathway regulates the
expression of Bcl-2, Bcl-XL, and Mcl-1 and promotes survival of
human pancreatic cancer cells. J Cell Biochem. 79:355–369. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wei MC, Zong WX, Cheng EHY, et al:
Proapoptotic BAX and BAK: a requisite gateway to mitochondrial
dysfunction and death. Science. 292:727–730. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kobayashi Y, Shibusawa A, Saito H, et al:
Isolation and functional characterization of a novel organic solute
carrier protein, hOSCP1. J Biol Chem. 280:32332–32339. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lin JG, Chen GW, Li TM, Chouh ST, Tan TW
and Chung JG: Aloe-emodin induces apoptosis in T24 human bladder
cancer cells through the p53 dependent apoptotic pathway. J Urol.
175:343–347. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Trotta AP, Need EF, Selth LA, et al:
Knockdown of the cochaperone SGTA results in the suppression of
androgen and PI3K/Akt signaling and inhibition of prostate cancer
cell proliferation. Int J Cancer. 133:2812–2823. 2013.PubMed/NCBI
|
|
20
|
Zhao D, Lin F, Wu X, et al: Pseudolaric
acid B induces apoptosis via proteasome-mediated Bcl-2 degradation
in hormone-refractory prostate cancer DU145 cells. Toxicol In
Vitro. 26:595–602. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu Y, Zhang SP and Cai YQ: Cytoprotective
effects of selenium on cadmium-induced LLC-PK1 cells apoptosis by
activating JNK pathway. Toxicol In Vitro. 21:677–684. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tucci P, Agostini M, Grespi F, et al: Loss
of p63 and its microRNA-205 target results in enhanced cell
migration and metastasis in prostate cancer. Proc Natl Acad Sci
USA. 109:15312–15317. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak K and Schmittgen T: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2 (-Delta Delta C (T) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kato T, Fujita Y, Nakane K, et al: ETS1
promotes chemoresistance and invasion of paclitaxel-resistant,
hormone-refractory PC3 prostate cancer cells by up-regulating MDR1
and MMP9 expression. Biochem Biophys Res Commun. 417:966–971. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tanaka H, Kono E, Tran CP, et al:
Monoclonal antibody targeting of N-cadherin inhibits prostate
cancer growth, metastasis and castration resistance. Nat Med.
16:1414–1420. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Aghaei M, Karami-Tehrani F, Panjehpour M,
Salami S and Fallahian F: Adenosine induces cell-cycle arrest and
apoptosis in androgen-dependent and -independent prostate cancer
cell lines, LNcap-FGC-10, DU-145, and PC3. Prostate. 72:361–375.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu X, Gao X, Jin L, et al:
Antiproliferation and cell apoptosis inducing bioactivities of
constituents from Dysosma versipellis in PC3 and Bcap-37
cell lines. Cell Div. 6:142011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
De Zutter GS and Davis RJ: Pro-apoptotic
gene expression mediated by the p38 mitogen-activated protein
kinase signal transduction pathway. Proc Natl Acad Sci USA.
98:6168–6173. 2001.
|
|
29
|
Murillo H, Schmidt LJ and Tindall DJ:
Tyrphostin AG825 triggers p38 mitogen-activated protein
kinase-dependent apoptosis in androgen-independent prostate cancer
cells C4 and C4-2. Cancer Res. 61:7408–7412. 2001.PubMed/NCBI
|
|
30
|
Tanaka Y, Gavrielides MV, Mitsuuchi Y,
Fujii T and Kazanietz MG: Protein kinase C promotes apoptosis in
LNCaP prostate cancer cells through activation of p38 MAPK and
inhibition of the Akt survival pathway. J Biol Chem.
278:33753–33762. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Marsden VS, O’Connor L, O’Reilly LA, et
al: Apoptosis initiated by Bcl-2-regulated caspase activation
independently of the cytochrome c/Apaf-1/caspase-9 apoptosome.
Nature. 419:634–637. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cory S and Adams JM: The Bcl2 family:
regulators of the cellular life-or-death switch. Nat Rev Cancer.
2:647–656. 2002. View
Article : Google Scholar : PubMed/NCBI
|