Introduction
The RNA-dependent protein kinase (PKR) is a
serine/threonine kinase that was originally identified as a
component of the interferon (IFN)-induced anti-viral response
(1,2). In response to viral infection, IFN
induces the expression of a number of genes, including the human
PKR gene. The anti-viral action of PKR is due to its activation by
viral double-stranded RNA (dsRNA) which results in the
phoshorylation of the α-subunit of eukaryotic translational
initiation factor 2 (eIF 2α) with the subsequent shut-down of
protein synthesis and inhibition of virus replication. It was
demonstrated that PKR is ubiquitously expressed and can also be
activated by cellular RNAs (3,4) and
proteins (5,6), suggesting that the function of PKR is
beyond the anti-viral defense.
Several studies have shown that PKR plays a role in
the regulation of important cell processes such as apoptosis,
signal transduction, cell proliferation and differentiation
(1,4,7,8). It
has been reported that PKR acts as a tumor suppressor or stimulator
protein, depending on the tumor type (1,7,8). The
role of PKR in cancer has been extensively studied, however, the
results are controversial. In a previous study, we showed that a
point mutation in the RNA-binding domain I results in the loss of
PKR activity in acute lymphoblastic leukemia (9). By contrast, PKR silencing by RNA
interference (RNAi) significantly reduced the number of pulmonary
metastatic nodules in the B16-F10 melanoma. In that experimental
model of metastasis, our findings suggested that the effect of PKR
knockdown is mediated by the transcription factor NF-κB through the
reduction of the phosphorylation of its inhibitor IκBβ by PKR
(10).
RNAi has become a powerful tool for investigating
gene function in mammalian cells (10,11).
The potential therapeutic of RNAi technology has also been explored
in cancer since RNAi is able to selectively knockdown critical
genes involved in cell proliferation (12). The RNAi technology holds promise for
cancer treatment, however, a number of obstacles remain to be
overcome before RNAi can be used in the clinic. Notably, the main
challenge is to develop methods to specifically deliver RNAi to
tumor cells. In the last few years, several strategies have been
used to surmount this barrier (13–15)
and the intratumoral injection of RNAi emerges as a promising
approach in the case of solid tumors (16).
In the present study, we investigated the effect of
the intratumoral PKR short hairpin (shRNA)-expressing plasmid on
the growth of B16-F10 melanoma in C57BL/6 mice. Since the effect of
PKR on B16-F10 melanoma cells is mediated by NF-κB, we also
examined the level of its inhibitor IκBβ in the B16-F10 melanoma
cells following the in vivo knockdown of PKR expression.
Materials and methods
Culture of tumor cells
B16-F10 melanoma cells were plated in tissue culture
flasks and cultured in RPMI-1640 medium (Life Technologies,
Carlsbad, CA, USA) supplemented with 10% inactivated fetal calf
serum (Life Technologies), 2 mM L-glutamine (Life Technologies) and
1% penicillin/streptomycin (100 U/ml; Life Technologies) in a
humidified incubator at 37°C and 5% CO2.
Target sequence selection of PKR mRNA and
plasmid vector construction
Three shRNAs targeting murine PKR and one scrambled
shRNA (used as a control) with sense and antisense sequences were
used. The selection of shRNA sequences was based on the shRNA
Target Finder and Design Tool available at Dharmacon website. Each
shRNA contains a sense strand of 19 nucleotides followed by a short
spacer (AAGTTCTCT), and an antisense strand and stop signal (TTTTT)
for RNA polymerase III.
siRNA sequences were selected a Blast search. The
selected siRNA sequences were expressed against sequence tag
libraries to ensure that only a single gene was targeted. A
predicted secondary structure of the intended mRNA was assessed to
avoid a steric hindrance of its binding. Each freeze-dried shRNA
was reconstituted with RNase-free water to prepare a stock
solution. Screening for the two inserts was performed by digestion
with PstI (Promega, Madison, WI, USA). The targeting
sequence and location of each shRNA in PKR cDNA is shown in
Table I.
 | Table ISequences of shRNA
oligonucleotides. |
Table I
Sequences of shRNA
oligonucleotides.
| shRNA | Target
positiona | Sequence |
|---|
| PKR-1 | 24 |
5′-ACGGAGGGCGAATAGATTT-3′ |
| PKR-2 | 320 |
5′-AAACGCTGCAGCCAAATTA-3′ |
| PKR-3 | 998 |
5′-ACTCAATCACGTCAACATT-3′ |
| Scrambled | - |
5′-ACGGAGGGCGAATAGATTT-3′ |
Transfection of B16-F10 melanoma
cells
B16-F10 melanoma cells were plated in tissue culture
flasks at a density of 7×105 cells. After an overnight
incubation and a confluence of ~70–80%, the cells were transfected
with 30 μg of each PKR shRNA (PKR-1, PKR-2 and PKR-3 shRNA) or
scrambled shRNA using 30 μl of Lipofectamine 2000 (Life
Technologies). The plasmid and Lipofectamine 2000 were diluted in
serum-free medium, left at room temperature for 5 min, mixed
immediately, and incubated for 20 min at room temperature at a v/w
ratio of liposomes to shRNA of 1:1. The culture medium was removed
and the shRNA-lipid complex (1.5 ml total volume) was added. The
transfection efficiency (~75–80%) was evaluated using green
fluorescent protein (GFP).
We examined whether the three PKR shRNAs were
effective in reducing PKR expression in cultured B16-F10 cells at
48 h after transfection when the cells were lysed and the amount of
PKR mRNA as well as the amount of PKR protein were measured.
RNA isolation
Total cellular RNA was extracted using TRIzol LS
reagent (Life Technologies). Cellular RNA in the aqueous phase was
transferred to sterile RNAse-free 1.5-ml microcentrifuge tubes and
precipitated by adding an equal volume of isopropanol.
Centrifugation at 10,000 rpm for 10 min was then performed. The
pellets were washed with 70% ethanol, air-dried and ressuspended in
DEPC water. RNA concentrations were determined by
spectrophotometric absorbance at 260 nm. The quality of extracted
RNA was determined by electrophoresis. RNA (1 μg) was boiled in
formamide loading buffer and snap cooled on ice. Samples were
subjected to electrophoresis with 1% agarose gels in 1X TBE buffer
(89 mM Tris-HCl, 2.5 mM EDTA, and 89 mM boric acid). Nucleic acids
were visualised by ethidium bromide staining and observed under
ultraviolet light. The presence of intact 28S and 18S rRNA bands
showed good quality cellular RNA. The RNA preparations were used
for semi-quantitative RT-PCR.
Analysis of PKR expression by reverse
transcription-PCR
Primer pairs designed to amplify PKR and β-actin
were synthesized by IDT (Integrated DNA Technologies, Coralville,
IA, USA). The primer sequences used included: PKR, sense,
5′-GTGGACATCTTTGCTTTGGGCCTT-3′ and antisense,
5′-TGTTCCTCCATTCAGCCAAGGTCT-3′ (GenBank accession no. M93567);
β-actin, sense, 5′-TGGAATCCTGTGGCA TCCATGAAAC-3′ and antisense,
5′-TAACGCAGCAGT AACAGTCCG-3′ (GenBank accession no. BC014861).
Total cellular RNA was extracted using TRIzol reagent (Life
Technologies) and reverse-transcribed with 0.5 μg of the oligodT
primer, 1 unit of reverse transcriptase, 1 unit of RNase inhibitor,
5 μl of 5X buffer and 4 μl MgCl2 (Life Technologies). We
used 2 μl of each reaction for PCR in 2 μl of sense and antisense
primers, respectively, 0.75 μl of MgCl2 (25 mmol/l), 2.5
μl of deoxynucleotide triphosphates (1.25 mM), 2.5 μl of 10X PCR
buffer, and 1 unit of Taq DNA polymerase in a 25-μl reaction
volume. β-actin was used as a control to assess the sample
integrity and to normalize posterior results. PCR conditions for
β-actin were 4-min denaturation at 94°C, 35 cycles of 1 min at
94°C, 1 min at 52°C and 2 min at 72°C, and 10-min elongation at
72°C in a thermocycler (Abgene, Epsom, UK). PCR conditions for PKR
were 4-min denaturation at 94°C, 37 cycles of 1 min at 94°C, 1 min
at 67°C and 1 min at 72°C, and 10-min elongation at 72°C in a
thermocycler (Abgene). PCR products of β-actin (364 bp) and PKR
(290 bp) were analysed by electrophoresis in a 1.5% agarose gel and
visualized using UV fluorescence after staining with ethidium
bromide. Quantification of PKR bands was performed by using
ImageQuant software, version 3.3 (Molecular Dynamics, Sunnyvale,
CA, USA). The gray scale ratio of PKR/β-actin was calculated and
the results were expressed as a percentage. The clone with the
highest inhibition of PKR expression was selected for in
vivo studies.
Western blot analysis
B16-F10 melanoma cells were washed with ice-cold
Tris-buffered saline (TBS) and lysed with 3 volumes of lysis buffer
consisting of 20 mM Tris-HCl (pH 7.6), 50 mM KCl, 400 mM NaCl, 1 mM
EDTA, 0.2 mM phenylmethylsulfonyl fluoride, aprotinin (2 μg/ml),
leupeptin (2 μg/ml), 1 mM dithiotreitol, 1% Triton X-100 and 20%
glycerol. The lysates were centrifuged at 10,000 rpm for 20 min and
the supernatant was stored at −70°C. The soluble proteins were then
measured according to the Lowry method. β-actin was used as a
loading control. Total cellular protein (30 μg) was separated by
electrophoresis through a 10% SDS-PAGE resolving gel with an
SDS-PAGE stacking gel. After electrophoresis, the proteins were
transferred onto a Hybond-C supported nitrocellulose membrane (GE
Healthcare, Little Chalfont, UK) by electroblotting for 4 h at 45
V, 25°C, in transfer buffer (3.94 g Tris-HCl, 18.80 g glycine, 240
ml methanol, and 10% SDS). The membranes were blocked with 10%
dried milk in TBS (20 mM Tris, and 500 mM NaCl) at room temperature
overnight, washed twice and then incubated at room temperature with
mouse anti-PKR monoclonal antibody, rabbit anti-IκBβ polyclonal
antibody (Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA) or
mouse anti-β-actin monoclonal antibody (Santa Cruz Biotechnology)
in TBS buffer for 80 min. The membrane was washed in TBS 1X
Tween-20 for 20 min, and secondary anti-mouse antibodies or
anti-rabbit antibodies labeled with horseradish peroxidase (GE
Healthcare) were added and the membrane was incubated at room
temperature for 60 min. The membrane was washed twice in TBS-T for
20 min and in TBS for 5 min. Antibody-labeled protein bands were
visualized with ECL detection reagents (GE Healthcare) according to
the manufacturer’s instructions. Quantification of bands was
performed by using the ImageQuant software, version 3.3 (Molecular
Dynamics) and the results were expressed as a percentage.
Transfection of B16-F10 melanoma cells
for in vivo assays
For in vivo experiments, B16-F10 melanoma
cells were transfected with PKR-2 or scrambled shRNA for 5 h.
Following transfection, the cells were detached with EDTA, washed
twice in PBS and then resuspended in RPMI at 2.0×106
cells/ml. Cell viability was assessed by trypan blue staining and
was >95%.
Animals
All the protocols involving animals were reviewed
and approved by our Institutional Animal Care Committee. C57BL/6
mice, weighing 20–25 g, that were raised at the Central Animal
Laboratory of the School of Medicine of Ribeirão Preto, SP, Brazil
were used.
Tumorigenic assay
Based on the in vitro findings, PKR-2 shRNA
was selected for the in vivo experiments. Tumor cells
transfected with PKR-2 or scrambled shRNA for 5 h were harvested
with EDTA, washed with RMPI, and resuspended in serum-free DMEM.
The cells were then inoculated subcutaneously (4.0×105
cells) into the right flank of mice (n=10 per group). The mice were
sacrificed 14 days post-inoculation, tumors were excised and
weighed on a microbalance Sartorius Supermicro (model S4;
Sartorius, Goettingen, Germany).
Intratumoral injection of PKR-2
shRNA
A total of 4.0×105 cells/ml melanoma
cells were subcutaneously inoculated in 0.2 ml of RPMI-1640 medium
into the right flank of C57BL/6 mice (n=10 per group). After 7 days
of tumor inoculation mice received a single intratumoral injection
of 2 μg of shRNA PKR-2 complexed with 2 μl of Lipofectamine 2000
dissolved in 50 μl of RPMI. Some mice received an intratumoral
injection of 2 μg scrambled shRNA-expressing plasmid complexed with
2 μl of Lipofectamine 2000 as a negative control. The mice were
sacrificed 7 days aftter intratumoral injection and the tumors was
weighed.
Statistical analysis
Data are expressed as means ± SD. Student’s t-test
was used to compare treated with control groups. P<0.05 was
considered significant.
Results
Transfection of B16-F10 melanoma cells
with PKR-2 shRNA-expressing plasmid results in the degradation of
PKR mRNA
To reduce the expression of PKR mRNA in B16-F10
melanoma cells, three PKR shRNAs were designed. Each PKR shRNA was
annealed and ligated into the psiSTRIKE vector controlled by Pol
III U6 promoter. The tumor cells were transfected at 5, 24 and 48 h
with the plasmid-based PKR-1, PKR-2, PKR-3 shRNA or scrambled
shRNA-expressing plasmids with Lipofectamine 2000. Following
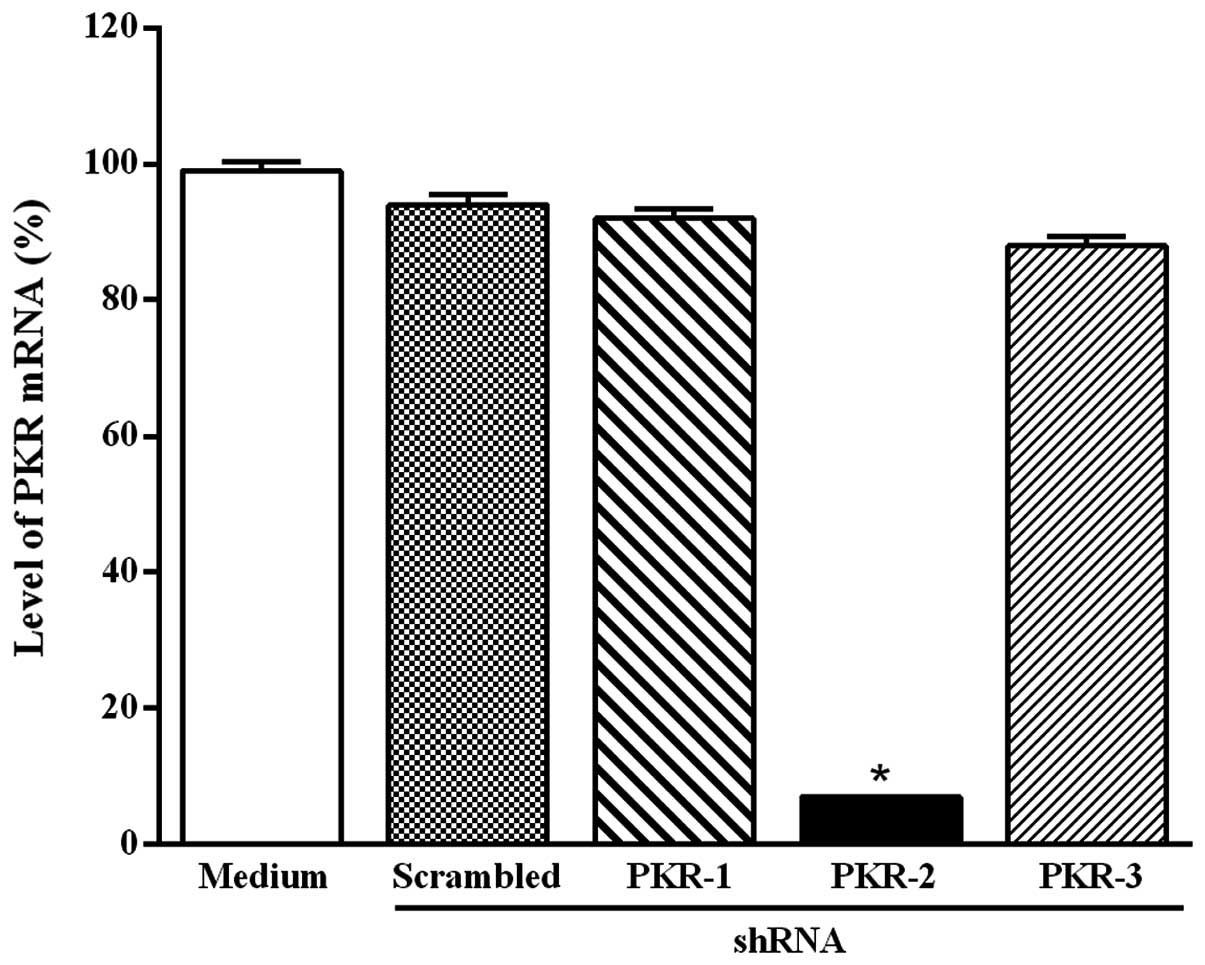
transfection, PKR degradation was monitored by RT-PCR. Fig. 1 shows that only the plasmid-based
PKR-2 shRNA significantly reduced the level of PKR mRNA after 48 h
of transfection and this effect continued for up to 4 days (data
not shown).
Transfection of B16-F10 melanoma cells
with PKR-2 shRNA-expressing plasmid reduces the level of PKR
protein
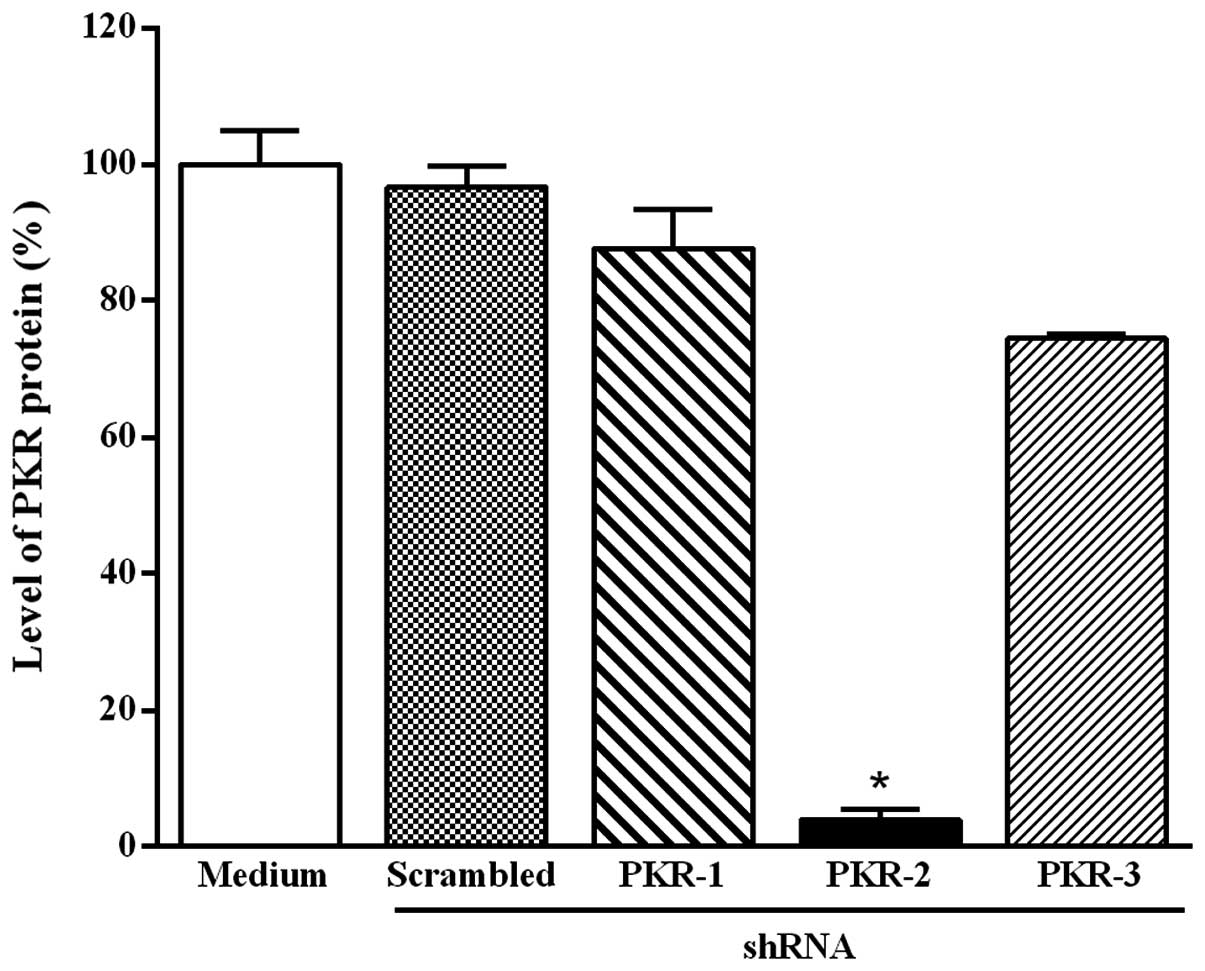
The effect of the transfection of B16-F10 melanoma
cells with the three PKR shRNA-expressing plasmids was also
monitored by western blot analysis. The downregulation of PKR
protein expression was also significantly detected only with the
plasmid-based PKR-2 shRNA at 48 h after transfection (Fig. 2). This effect was observed for up to
4 days and the reduction of PKR protein level at 5 and 24 h after
transfection was smaller than that at 48 h (data not shown).
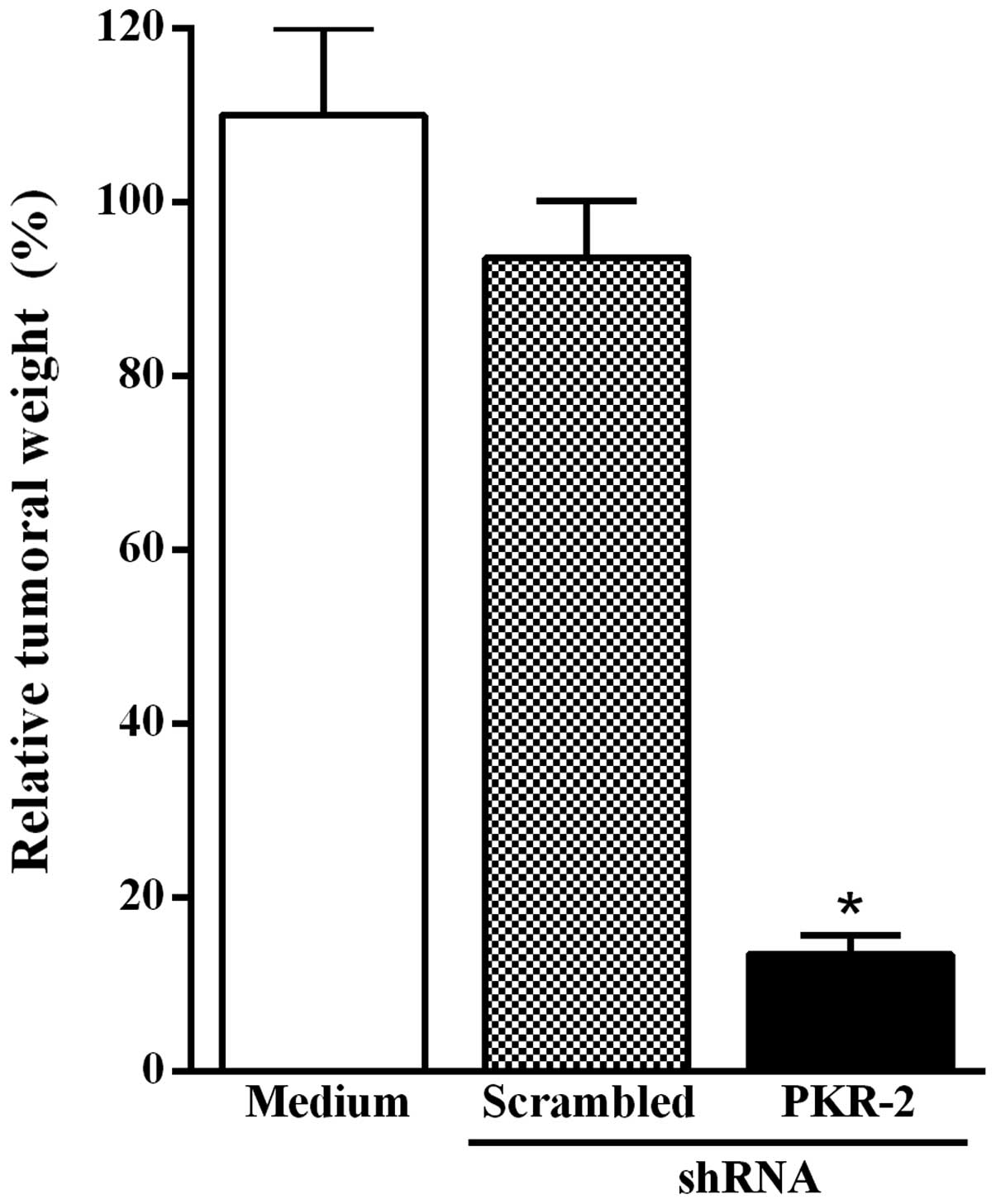
Transfection of B16–10 melanoma cells
with the PKR-2 shRNA-expressing plasmid inhibits tumor growth
Based on results shown in Figs. 1 and 2, the effect of the transfection of
B16-F10 melanoma cells with the PKR-2 shRNA-expressing plasmid on
tumor growth was subsequently investigated. The transfected tumor
cells were injected subcutaneously into C57BL/6 mice. The animals
were sacrificed 14 days after the inoculation of B16-F10 melanoma
cells and tumors were excised and weighed. Fig. 3 shows that tumor growth was
significantly inhibited when B16-F10 melanoma cells were
transfected with the PKR-2 shRNA-expressing plasmid as compared to
tumor cells transfected with the plasmid-based scrambled shRNA.
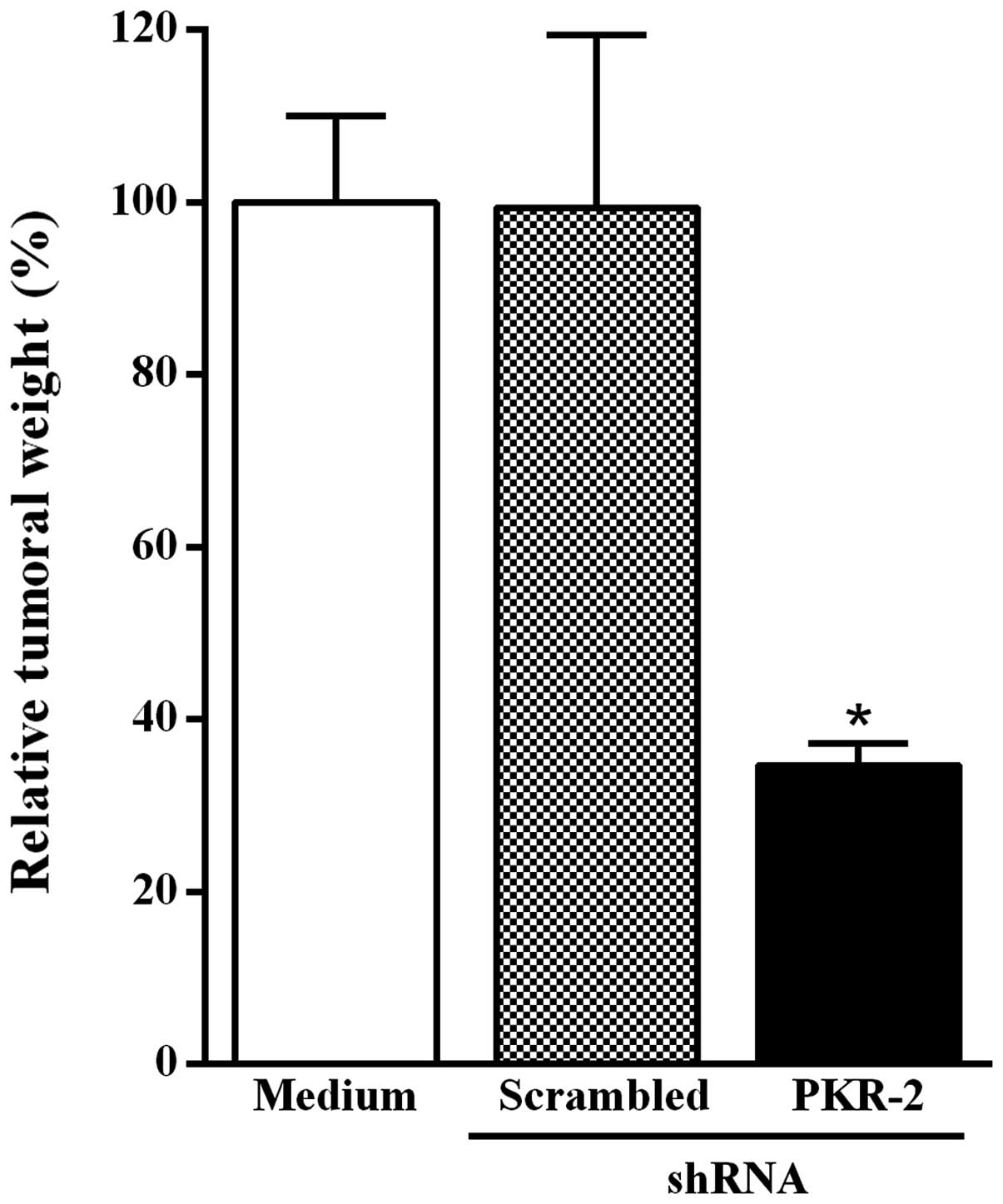
Intratumoral injection of the PKR-2
shRNA-expressing plasmid inhibits B16-F10 melanoma growth
To investigate the effect of the intratumoral
injection of the PKR-2 shRNA-expressing plasmid on tumor growth,
mice were inoculated subcutaneously with B16-F10 melanoma cells.
Fig. 4 shows that the reduction of
tumor weight was significantly increased in mice that had received
a single intratumoral injection of the PKR-2 shRNA-expressing
plasmid as compared to animals injected with the plasmid-based
scrambled shRNA.
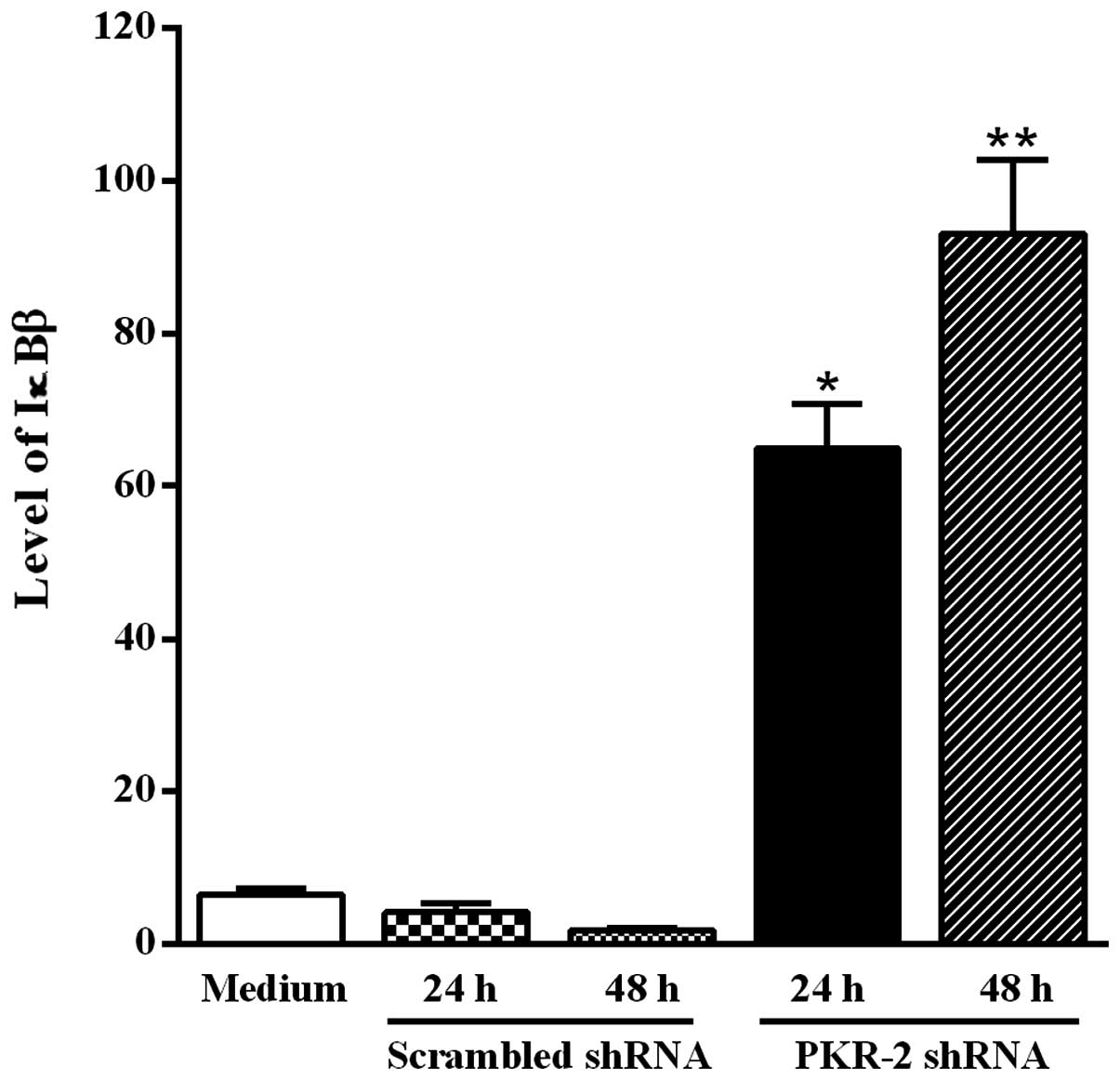
Transfection of B16-F10 melanoma cells
with PKR-2 shRNA- expressing plasmid increases the level of
IκBβ
PKR is able to activate IKK, which phosphorylates
the inhibitor IκBβ of the transcription factor NF-κB. Moreover, the
phosphorylation of IκBβ results in its degradation. To determine
whether downregulation of the activated PKR by the PKR-2 shRNA was
accompanied by an increase of the IκBβ level, tumor cells were
transfected with the PKR-2 shRNA-expressing plasmid and western
blot analysis was performed using the anti-IκBβ antibody. Fig. 5 shows that the IκBβ level was
significantly increased in B16-F10 melanoma cells transfected with
PKR-2 shRNA-expressing plasmid when compared to tumor cells
transfected with the plasmid-based scrambled shRNA.
Discussion
RNAi-based therapy is known to be effective and
elicit a gene silencing response, and that double-stranded RNA
molecules must be delivered to the target cell. However, delivery
of RNAi has been challenging and despite many efforts to overcome
this obstacle, current approaches are limited to few clinical
indications (12,17). Over the last few years, attempts
have been made to increase the efficacy of siRNA delivery including
chemical modifications and nanoparticle-based encapsulation of
siRNA (18,19).
In the present study, RNAi technology was applied to
elucidate the role played by PKR in B16-F10 melanoma since the
function of this protein kinase in tumorigenesis remains
controversial. A plasmid vector was employed for the expression of
anti-PKR shRNA and the delivery was performed by intratumoral
injection which is promising for solid tumors. Results of the
present study show that transfection of the B16-F10 melanoma cells
with the PKR-2-expressing plasmid was effective in inducing a
statistically significant reduction of the levels of PKR mRNA and
protein expression, whereas PKR-1 and PKR-3 shRNA had no effect.
Therefore, the PKR-2 shRNA-expressing plasmid was selected for
subsequent experiments. We then investigated whether transfection
with the PKR-2 shRNA-expressing plasmid reduces the potential of
B16-F10 melanoma cells to develop tumor following subcutaneous
injection in mice. The results show that PKR silencing inhibits
tumor growth, suggesting that PKR acts as a tumor supressor, a
finding that is consistent with previous results using an
experimental model of metastasis (10).
It is known that in cancer research, the results
obtained in vitro may not be reproducible in vivo due
to the complexity of biological systems. Thus, we investigated the
effect of the intratumoral injection of the PKR-2 shRNA-expressing
plasmid in the pre-established subcutaneous B16-F10 melanoma. We
found that PKR silencing inhibits tumor growth when compared to
animals injected with the plasmid-based scrambled shRNA. It is of
note that this effect was obtained with only a single injection of
the PKR-2 shRNA-expressing plasmid and this approach for RNAi
delivery was effective for at least 7 days. Previous studies have
demonstrated that shRNA expression vectors are able to sustain gene
silencing in human cell lines (16,20).
PKR is known to activate NF-κB by directly
phosphorylating its inhibitor IκBα (21) or indirectly by activating the IκB
kinase (IKK) complex (22). The
phosphorylated IκB undergoes polyubiquitination which targets it
for degradation by the ubiquitin-proteasome system, thereby
releasing NF-κB to translocate to the nucleus. Notably, NF-κB
regulates the transcription of genes involved in
immuno-inflammatory responses, cell cycle progression, inhibition
of apoptosis and cell adhesion, thus promoting carcinogenesis and
cancer progression.
A notable finding of the present study was the
observation that IκBβ level is significantly increased in B16-F10
melanoma cells transfected with PKR-2 shRNA-expressing plasmid when
compared to tumor cells transfected with the plasmid-based
scrambled shRNA. This result suggests that the inhibition of
B16-F10 melanoma growth due to PKR silencing may be mediated by the
transcription factor NF-κB. It is known that the inhibition of the
degradation of the IκBβ inhibitor is an efficient mechanism to
avoid the persistent activation of NF-κB (23,24).
Recently, it was demonstrated that the okadaic acid-induced
phosphorylation of I-κBα was mediated by PKR (25).
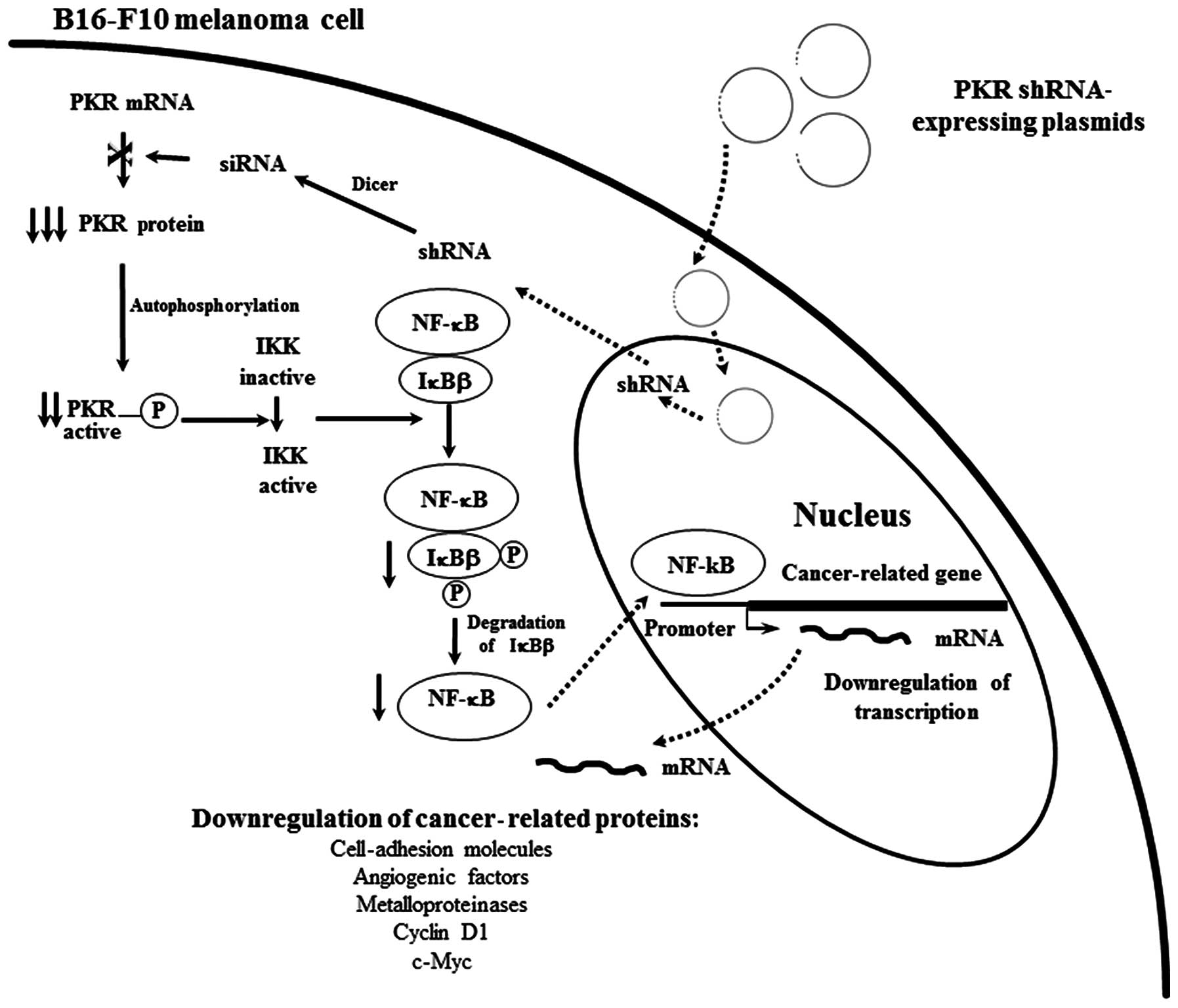
Based on results of the present study and our
previous findings (4,26,27),
we suggest a molecular mechanism that may explain the finding that
silencing PKR expression in B16-F10 melanoma cells by the
intratumoral injection of the PKR shRNA-expressing plasmids results
in the reduction of tumor growth. Uptake of the injected plasmids
by tumor cells occurs (Fig. 6) and
thus PKR shRNA are transcribed in the nucleus, export for the
cytoplasm and are processed by Dicer to generate siRNA, which
induces the specific degradation of PKR mRNA. Therefore, the level
of PKR protein is decreased with the subsequent reduction of
phosphorylated IκBβ, resulting in inhibition of the nuclear
translocation of NF-κB. Thus, the expression of cancer-related
proteins with promoters containing NF-κB-binding sites such as
c-myc, cyclin D1, cell adhesion molecules, angiogenic factors and
metalloproteinases, were downregulated, which may result in the
growth inhibition of pre-existing B16-F10 melanoma.
In conclusion, the present study validates the
hypothesis that the direct administration of RNAi-based
therapeutics into the target tumor is a promising approach for
overcoming obstacles of systemic delivery. Our results also suggest
that the intratumoral injection of PKR shRNA-expressing vector is a
novel therapeutic approach for human solid tumors such as cutaneous
melanoma and breast cancer since PKR is overexpressed in these
tumors.
Acknowledgements
The present study was supported by FAPESP
(06/57963-1). We would like to thank Cacilda D. Pereira and Zuleica
A.S. Moraes for the technical assistance.
References
|
1
|
Clemens MJ and Elia A: The double-stranded
RNA-dependent protein kinase PKR: Structure and function. J
Interferon Cytokine Res. 17:503–524. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
García MA, Gil J, Ventoso I, Guerra S,
Domingo E, Rivas C and Esteban M: Impact of protein kinase PKR in
cell biology: from antiviral to antiproliferative action. Microbiol
Mol Biol. 70:1032–1060. 2006.PubMed/NCBI
|
|
3
|
Murad JM, de Souza LR and De Lucca FL: PKR
activation by a non-coding RNA expressed in lymphocytes of mice
bearing B16 melanoma. Blood Cells Mol Dis. 37:128–133. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Watanabe MAE, Souza LR, Murad JM and De
Lucca FL: Activation of the RNA dependent protein kinase (PKR) of
lymphocytes by regulatory RNAs: implications for immunomodulation
in HIV infection. Curr HIV Res. 3:329–337. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Patel RC and Sen GC: PACT, a protein
activator of the interferon-induced protein kinase, PKR. EMBO J.
17:4379–4390. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Singh M and Patel RC: Increased
interaction between PACT molecules in response to stress signals is
required for PKR activation. J Cell Biochem. 113:2754–2764. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Williams BR: PKR: a sentinel kinase for
cellular stress. Oncogene. 18:6112–6120. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Marchal JA, Lopez GJ, Peran M, Comino A,
Delgado JR, García-García JA, Conde V, Aranda FM, Rivas C, Esteban
M and Garcia MA: The impact of PKR activation: from
neurodeggeneration to cancer. FASEB J. 28:1965–1974. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Murad JM, Tone LG, Souza LR and De Lucca
FL: A point mutation in the RNA-binding domain I results in
decrease of PKR activation in acute lymphoblastic leukemia. Blood
Cells Mol Dis. 34:1–5. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Delgado-André N and De Lucca FL: Knockdown
of PKR expression by RNAi reduces pulmonary metastatic potential of
B16-F10 melanoma cells in mice: possible role of NF-kappaB. Cancer
Lett. 258:118–125. 2007.PubMed/NCBI
|
|
11
|
Gao L, Zhang L, Hu J, Li F, Shao Y, Zhao
D, Kalvakolanu DV, Kopecko DJ, Zhao X and Xu DQ: Downregulation of
signal transducer and activator of transcription 3 expression using
vector-based small interfering RNAs suppresses growth of human
prostate tumor in vivo. Clin Cancer Res. 11:6333–6341. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Deng Y, Wang CC, Choy KW, Du Q, Chen J,
Wang Q, Li L, Chung TK and Tang T: Therapeutic potentials of gene
silencing by RNA interference: principles, challenges, and new
strategies. Gene. 538:217–227. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Díaz MR and Vivas-Mejia PE: Nanoparticles
as drug delivery systems in cancer medicine: emphasis on
RNAi-containing nanoliposomes. Pharmaceuticals. 6:1361–1380.
2013.PubMed/NCBI
|
|
14
|
Li CH, Parker A, Menocal E, Xiang S,
Borodyansky L and Fruehauf JH: Delivery of RNA interference. Cell
Cycle. 5:2103–2109. 2006. View Article : Google Scholar
|
|
15
|
Sakurai Y, Hatakeyama H, Sato Y, Hyodo M,
Akita H and Harashima H: Gene silencing via RNAi and siRNA
quantification in tumor tissue using MEND, a liposomal siRNA
delivery system. Mol Ther. 21:1195–1203. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Deharvengt SJ, Gunn JR, Pickett SB and
Korc M: Intratumoral delivery of shRNA targeting cyclin D1
attenuates pancreatic cancer growth. Cancer Gene Ther. 17:325–333.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fujita Y, Takeshita F, Kuwano K and Ochiya
T: RNAi therapeutic platforms for lung diseases. Pharmaceuticals.
6:223–250. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nikitenko NA and Prassolov VS: Non-viral
delivery and therapeutic application of small interfering RNAs.
Acta Naturae. 5:35–53. 2013.PubMed/NCBI
|
|
19
|
Das J, Das S, Paul A, Samadder A,
Bhattacharyya SS and Khuda-Bukhsh AR: Assessment of drug delivery
and anticancer potentials of nanoparticles-loaded siRNA targeting
STAT3 in lung cancer, in vitro and in vivo. Toxicol Lett.
225:454–466. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Paddison PJ, Caudy AA, Bernstein E, Hannon
G and Conklin DS: Short hairpin RNAs (shRNAs) induce
sequence-specific silencing in mammalian cells. Genes Dev.
16:948–958. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kumar A, Haque J, Lacoste J, Hiscott J and
Williams BR: Double-stranded RNA-dependent protein kinase activates
transcription factor NF-kappa B by phosphorylating I kappa B. Proc
Natl Acad Sci USA. 91:6288–6292. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gil J, Alcamí J and Esteban M: Activation
of NF-kappa B by the dsRNA-dependent protein kinase, PKR involves
the I kappa B kinase complex. Oncogene. 19:1369–1378. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Thompson JE, Phillips RJ,
Erdjument-Bromage H, Tempst P and Ghosh S: I kappa B-beta regulates
the persistent response in a biphasic activation of NF-kappa B.
Cell. 80:573–582. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lindgren H, Olsson AR, Pero RW and
Leanderson T: Differential usage of IkappaBalpha and IkappaBbeta in
regulation of apoptosis versus gene expression. Biochem Biophys Res
Commun. 301:204–211. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Haneji T, Hirashima K, Teramachi J and
Morimoto H: Okadaic acid activates the PKR pathway and induces
apoptosis through PKR stimulation in MG63 osteoblast-like cells.
Int J Oncol. 42:1904–1910. 2013.PubMed/NCBI
|
|
26
|
De Lucca FL, Serrano SV, Souza LR and
Watanabe MA: Activation of RNA-dependent protein kinase and nuclear
factor-kB by regulatory RNA from lipopolysaccharide-stimulated
macrophages: implications for cytokine production. Eur J Pharmacol.
450:85–89. 2002.PubMed/NCBI
|
|
27
|
Watanabe MA, Rodrigues Souza L, Murad JM
and De Lucca FL: Antitumor activity induced by regulatory RNA:
possible role of RNA-dependent protein kinase and nuclear
factor-kappa B. Eur J Pharmacol. 465:205–210. 2003. View Article : Google Scholar : PubMed/NCBI
|