Introduction
A manned mission to Mars constitutes a great vision
of mankind, and is supported by several space agencies worldwide.
However, the era of manned missions beyond the low Earth orbit from
1969 through 1972 ended over 40 years ago. Reasons for the
cessation are, among others, various uncertainties regarding
physiological effects of space radiation, isolation and
microgravity during prolonged space flights of approximately 3
years. Space radiation consists of protons (87%), α-particles (12%)
and heavy ions (1%) in solar particle events (SPEs) and galactic
cosmic rays (GCRs). The dose deposition of heavy-ion radiation in
SPEs and GCRs occurs inhomogeneously as the ions release their
energy localized at a very high density along their trajectory.
This distinguishes them from X-rays, which show a homogeneous dose
deposition. Furthermore, these radiation types differ in their
depth-dose profile. In contrast to X-rays, heavy-ion radiation is
characterized by an inverted dose profile, i.e. the dose increases
with the depth of penetration (1).
Particularly highly ionizing heavy ions of GCRs can be hardly
shielded exposing the crew members to a serious medical safety risk
(2,3), since the probability to be hit by a
heavy ion beam increases with prolonged stay in space. Previous
studies indicated that the risk of cancer-related mortality for
extended exploration missions to the Moon and Mars varies from 0.2
to 15% (4,5), which is a very wide range. The
uncertainties warrant further experimental analysis of the risk of
radiation-induced diseases and complications during extended space
flights. From experience in cancer therapy, we know that
inflammation of the mucogingival membrane (oral mucositis) is a
frequent complication of heavy-ion and X-ray radiotherapy (6–11).
Patients often complain about burning and intolerance of spicy
foods, xerostomia, bacterial, fungal or viral infections, dental
caries and loss of taste (12).
Furthermore, there is significant pain, odynodysphagia, subsequent
dehydration, malnutrition, anorexia, cachexia, neurocognitive
alterations and depression (13).
Due to restricted possibilities of medical treatment in a space
shuttle during prolonged missions, acute oral mucositis will most
likely exacerbate to chronic mucositis with an increased
probability of carcinogenesis (14–17).
It is known that in tight cell clusters the cells
communicate with each other via molecular signaling (18–21).
Since three-dimensional (3D) cultures mimic the in vivo
situation much closer than conventional monolayer cultures, we
chose this system as an adequate in vitro model for analyses
of radiation-induced pro-inflammatory effects in early stages of
oral mucositis. In the present study, we utilized a 3D mucosa model
consisting of human keratinocytes and fibroblasts, where the
keratinocytes interact with the fibroblasts in the layer below.
In vivo, peripheral blood mononuclear cells (PBMCs) are
indispensable for induction and progress of inflammatory processes.
On that account, we additionally established co-cultures containing
PBMCs to mimic the in vivo situation much closer, and
compared the results to the cultures without immune cells.
The focus of the present study was on immediate and
early pro-inflammatory effects after irradiation, where nuclear
factor κB (NFκB) activation and increased expression of the
cytokines and chemokines are precursors of oral mucositis (22). By way of comparison, we additionally
exposed the 3D mucosa model containing PBMCs to X-rays. Following
irradiation with X-rays or heavy ions (12C), we analyzed
the radiation impact on epithelium compactness, DNA damage,
activation of NFκB and the release of the cytokine interleukin 6
(IL6) and chemokine IL8.
Materials and methods
Cell culture
Immortalized human gingival keratinocytes (IHGK;
kindly provided by Professor Dr Tomakidi, Department of Oral
Biotechnology, University Medical Center, Freiburg, Germany) were
cultured in complete keratinocyte growth medium (PromoCell,
Heidelberg, Germany). Immortalized human dermal fibroblasts HH4ded,
kindly provided by Professor Dr Rodemann (Department of Radiation
Oncology, University Hospital and Faculty of Medicine, Tuebingen,
Germany) were cultured in Dulbecco’s modified Eagle’s medium (DMEM;
high glucose with L-glutamine; Gibco®, Life Technologies
GmbH, Darmstadt, Germany) supplemented with 10% (v/v) fetal bovine
serum Gold (PAA Laboratories GmbH, Coelbe, Germany). Cells were
cultured under standard tissue culture conditions and tested
routinely for mycoplasma contaminations (Venor®
GeM Mycoplasma Detection kit; Minerva Biolabs, Berlin,
Germany). PBMCs were isolated from buffy coats from healthy donors
(Blood Bank University Medical Center Mainz, Germany). Initially,
the buffy coat was diluted in sterile phosphate-buffered saline
(PBS; 1:4). For isolation, separation medium LSM1077 (PAA
Laboratories GmbH) was overlayed with 25 ml blood suspension. After
a 20 min centrifugation (1,200 × g; Megafuge 1.0; Heraeus, Hanau,
Germany) with breaks turned-off, the interphase was transferred to
a new reaction tube, washed twice with PBS, and centrifuged for 10
min with 300 × g. Numbers of viable PBMCs were determined upon
microscopic observation in a Neubauer chamber after staining with
0.4% trypan blue.
Preparation of organotypic 3D oral mucosa
model
Organotypic co-cultures were grown in
gelatine-coated ThinCerts™ tissue culture inserts in deep 12-well
plates (Greiner Bio-One GmbH, Frickenhausen, Germany). The inserts
consist of a polyester capillary pore membrane with 0.4-μm pore
size and a growth area of 1.131 cm2. Before initiating
the organotypic cultures, ThinCerts™ were coated with 0.2% gelatin
solution and incubated for 30 min at 37°C. Subsequently, the
fibroblasts were seeded in the 12-well plates with 4×104
cells/insert. After 3 days, the inserts were transferred into deep
well plates with 4.5 ml of medium in each well and 3×105
cells/insert keratinocytes were added onto the fibroblast layer.
After a further 3 days, the models were exposed to an air-liquid
interface and incubated for another 20 days with media changes
twice a week. Co-cultures with immune cells were generated by
seeding 106 PBMCs/ThinCert™24 h before irradiation.
Irradiation
Irradiation was performed at the GSI Helmholtz
Center for Heavy Ion Research in Darmstadt. Deep well plates with
inserts containing organotypic cultures were completely filled with
medium, sealed and transferred to the GSI. Due to a few
uncertainties regarding cumulative dose for the Mars mission crew,
we selected different doses for the irradiation. Control samples
were left untreated (0 Gy), other samples were irradiated with 2 or
4 Gy of 12C particle radiation with Emax
155.26 MeV/u, an LET of 300 keV/μm and a dose-rate of ~0.6 Gy/min.
To provoke a reliable radiation response in the mucosa model, we
additionally irradiated with 10 Gy 12C ions in one
experiment. For comparison, additional 3D cultures were irradiated
with equivalent physical doses of X-rays (250 kV, Isovolt DS1;
Seifert, Germany; dose-rate 1 Gy/min). After irradiation, samples
were transferred to new deep 12-well plates that were completely
filled with medium and transported back to the Mainz laboratory,
where the cultures were snap frozen 4, 8, 24 and 48 h after
irradiation. In the course of transit and radiation, samples were
kept at room temperature.
Immunohistochemistry and morphometric
characterization of 3D cultures
For histological and immunofluorescence stainings,
snap frozen samples were embedded in Tissue-Tek® O.C.T.
Compound (Sakura, Torrance, CA, USA) and cryosectioned (10 μm) with
a microtome (SLEE, Mainz, Germany). After overnight drying at room
temperature, the sections were stored at −20°C. Hematoxylin (Carl
Roth GmbH & Co. KG, Karlsruhe, Germany) and eosin (HE) (Merck
KGaA, Darmstadt, Germany) stainings were used for analyses of
structural changes in irradiated tissue. For discrimination of
epithelium and connective tissue as well as DNA damage and NFκB
p50, immunofluorescence stainings were carried out. Sections were
initially blocked with 2% donkey serum, following incubation with
primary antibody for 1 h and 3 washing steps with PBS.
Subsequently, samples were incubated with secondary antibody for 45
min, washed again 4 times and mounted in ImmunoSelect Antifading
Mounting Medium (Dianova GmbH, Hamburg, Germany) containing
4′,6-diamidino-2-phenylindole (DAPI). For anti-phospho-histone H2AX
(γH2AX) staining, a preceding permeabilization of the cell
membranes with a permeabilizing buffer [PBS + 0.5% (v/v) Triton-X
100] was required. The following antibodies were used: monoclonal
mouse anti-panCK (Dako, Hamburg, Germany), polyclonal rabbit anti
NFκB p50 (Santa Cruz Biotechnology, Inc., Heidelberg, Germany),
monoclonal mouse anti-phospho-histone H2AX Ser139 (Millipore
Corporation, Billerica, MA, USA). Secondary detection was carried
out with polyclonal donkey anti-rabbit DyLight 594 and polyclonal
donkey anti-mouse DyLight 488 antibodies (Dianova GmbH). Secondary
antibodies alone were used as negative controls. Sections were
registered photographically using a BZ-8000 fluorescence microscope
(Keyence Deutschland GmbH, Neu-Isenburg, Germany) for HE stainings
and a confocal laser scanning microscope (Zeiss) was used for other
stainings. For computer based analyses of epithelial proportion,
the binary images of panCK stained cultures were quantified by
ImageJ 1.46 using the plugin ‘Live Histogram’. The values obtained
were divided by the values obtained of binary images of
DAPI-stained cultures, which were quantified in the same way.
Compactness of oral mucosa models based on cell density and
integrity was quantified from binary HE-stained cultures using the
plugin ‘Live Histogram’ and related to the total area of each
sample.
Quantification of DNA damage and NFκB
activation
Computer-based analyses were carried out to quantify
DNA damage and NFκB activation. To analyze NFκB p50 activation,
only nuclear localized staining was quantified. For that purpose,
the binary image of DAPI-stained cell nuclei served as template for
NFκB p50 images. The intensity of the NFκB p50 stainings in nuclei
was quantified by ImageJ 1.46 using the plugin ‘Measure RGB’
(author, Wayne Rasband; version 1). The values obtained were
divided by the values of binary nuclei stainings and normalized to
untreated samples. For the evaluation of radiation-induced DNA
double strand breaks (DSB), the foci number/nucleus of the DSB
marker γH2AX was quantified. In order to do this, the 16 bit
greyscale images obtained from γH2AX staining were counted by
ImageJ using the plugin ‘FociPicker3D 3D and 2D particle counter’
(author, Guanghua Du; 16.04.2010, version 1). Subsequently, the
foci number was divided by the corresponding visually determined
cell number.
Cytokine quantification
Cell-free media supernatants of organotypic (co-)
cultures were collected at the time of cryopreservation. Remaining
solid particles were sedimented by centrifugation for 10 min with
400 × g at 4°C. The supernatants were stored at −80°C until the
samples were analyzed according to the manufacturer’s instructions
with human IL6 and IL8 ELISA MAX™ Standard Sets (BioLegend GmbH,
Fell, Germany), respectively.
Statistical analysis
Data are shown as average values ± standard error of
the mean (SEM) from at least 3 independent experiments. Only the
data regarding the epithelial component relative to respective
grown culture are shown as the means ± standard deviation.
Statistical significance (p≤0.05) was determined using a
two-tailed, unpaired Student’s t-test. Statistically significant
alterations marked with an asterisk (*) refer to the
untreated control samples and those with a hash (#),
refer to the corresponding dose at 4 h.
Results
Radiation impact on epithelium
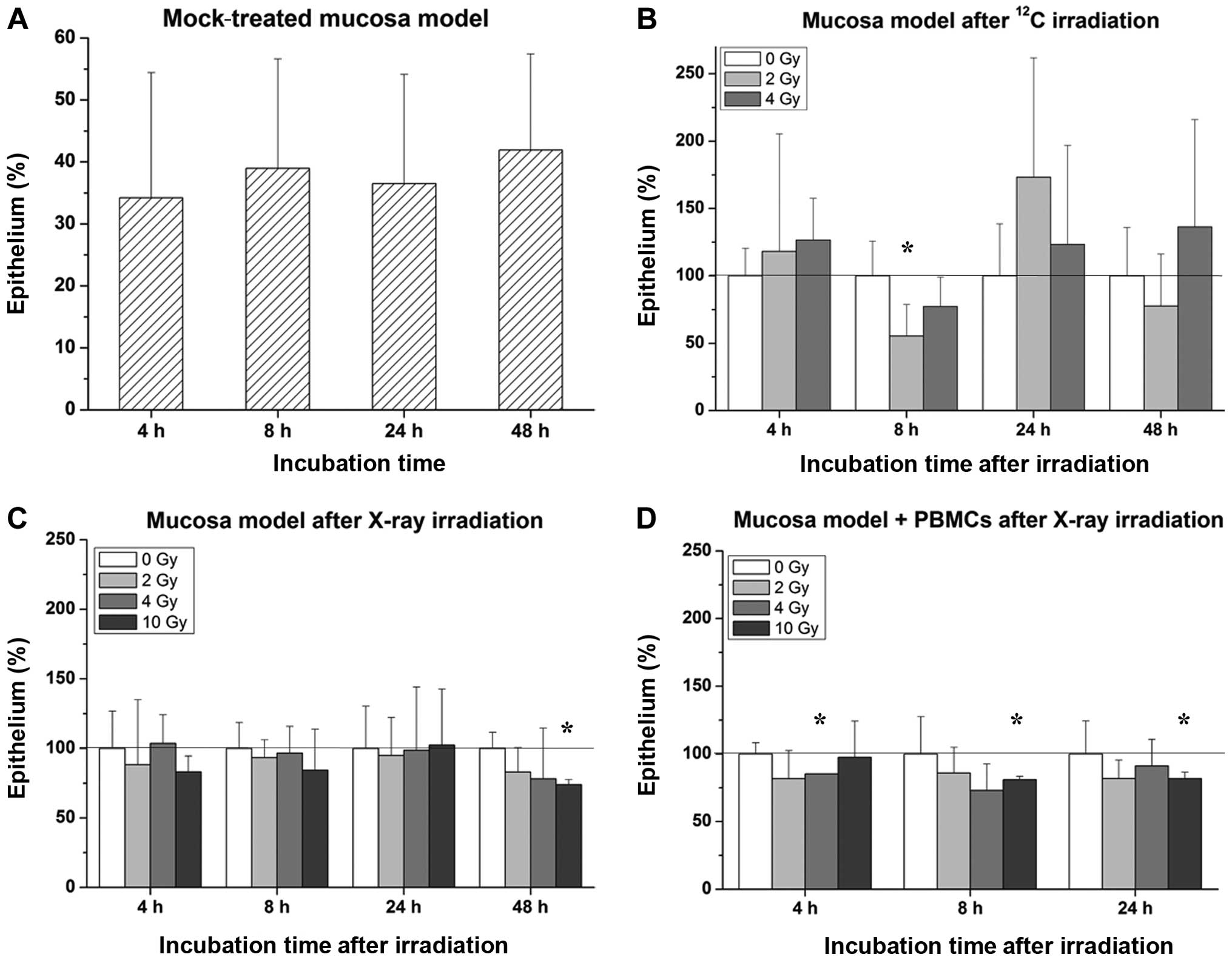
The epithelial proportion of oral mucosa models was
quantified from panCK stained cultures using at least 3 images of
the same 3D culture. In our mucosa model, the epithelial component
comprised ~30–40% (Fig. 1A).
Heavy-ion irradiation within a dose range of up to 4 Gy showed no
systematic effect on the proportion of the epithelium, as depicted
in Fig. 1B, whereas exposure to
X-rays at a dose of 10 Gy caused a shrinkage of the epithelium in
cultures without immune cells after 48 h and in co-cultures
containing immune cells after 8 and 24 h, respectively (Fig. 1C and D).
Radiation impact on compactness
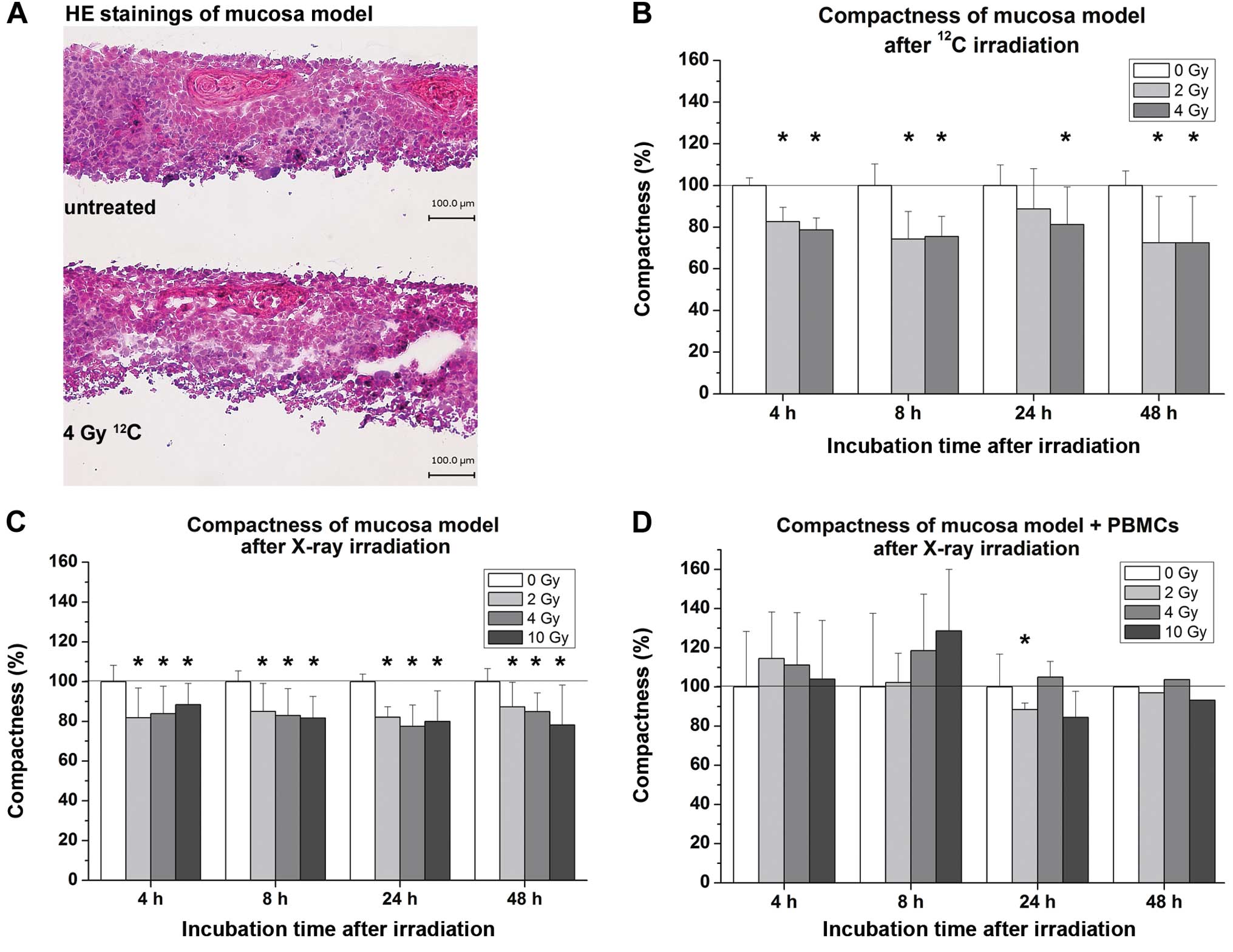
The compactness of oral mucosa models based on cell
density and integrity was quantified from HE-stained cultures using
at least 3 images of the same 3D culture. Radiation-induced loss of
compactness was clearly observed, as shown in Fig. 2A. Computer-based analyses revealed
that both heavy-ion irradiation with 12C particles and
X-rays caused a significant loss of compactness in almost all
irradiated samples (Fig. 2B and C).
Co-cultures containing immune cells exhibited no clear tendency of
changes in compactness, with the earliest effects occurring 24 h
after irradiation with 2 Gy (Fig.
2D).
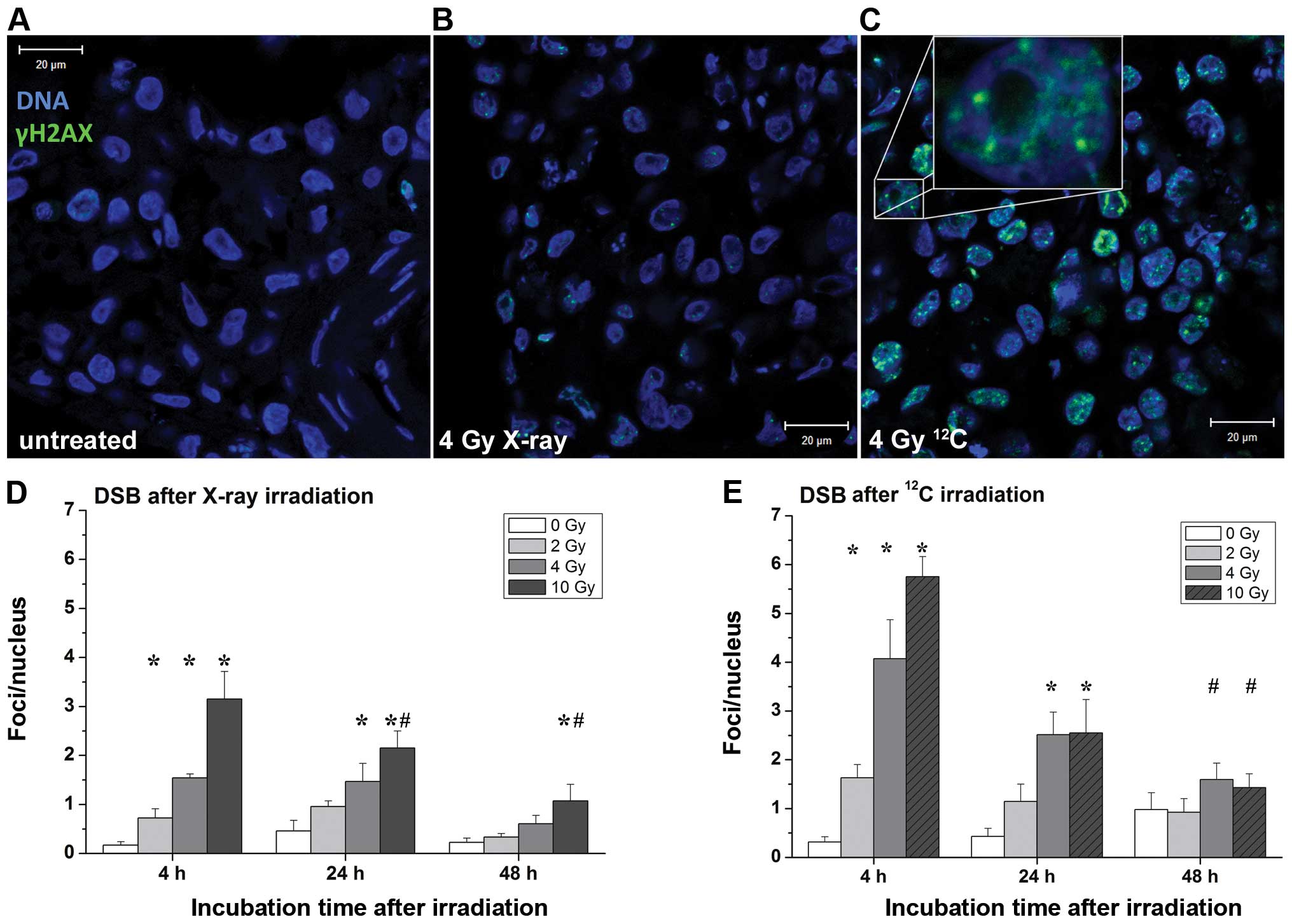
Radiation-induced DNA damage
Software-based evaluation of DSB was performed in
representative images of γH2AX stainings. Controls showed barely
visible DNA damage (Fig. 3A). In
contrast, analyses of cultures 4 h after irradiation showed twice
as many DSB after irradiation with heavy ions compared to X-rays
(Fig. 3B and C). Both irradiation
with heavy ions and with X-rays showed a dose-dependent increase of
DSB in all mucosa models, as shown in Fig. 3D and E. After an incubation time of
24 h, the number of foci/nucleus in both radiation types aligned
and decreased to the untreated level after 48 h. Samples, which
were irradiated with 10 Gy of X-rays showed, after 24 and 48 h,
significantly lower DSB levels compared to the corresponding dose
after 4 h. Similarly, 48 h after heavy ion irradiation with 4 or 10
Gy, the quantity of DSB decreased significantly, compared to the
corresponding doses after 4 h.
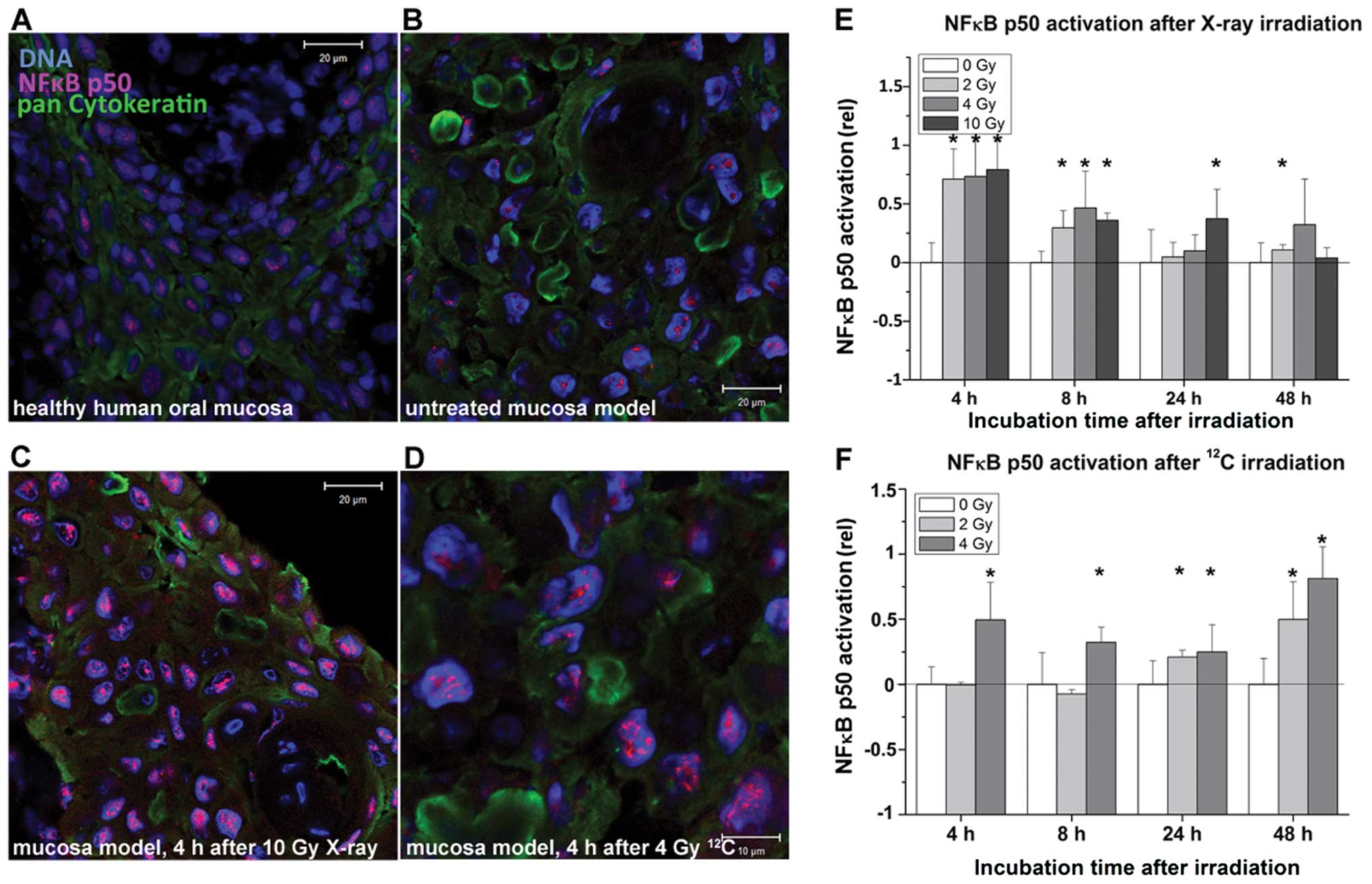
Radiation-induced activation of NFκB
p50
Fig. 4A–D show
representative images of NFκB stainings on cryosections of the
mucosa model. A clear accumulation of the transcription factor in
the nucleus according to its activation pattern was observed after
exposure to both types of radiation in all mucosa models (Fig. 4C and D). Software-based evaluations
confirmed these observations (Fig. 4E
and F). X-rays induced NFκB p50 activation almost immediately
after irradiation, with statistically significant changes at all
doses after 4 and 8 h. The NFκB p50 activation decreased after 24
and 48 h to the untreated level, significant changes were detected
only after exposure to 10 Gy after 24 h and to 2 Gy after 48 h. On
the other hand, 12C particles induced significant
changes in NFκB activation 24 h after 2 Gy and 48 h after 4 Gy.
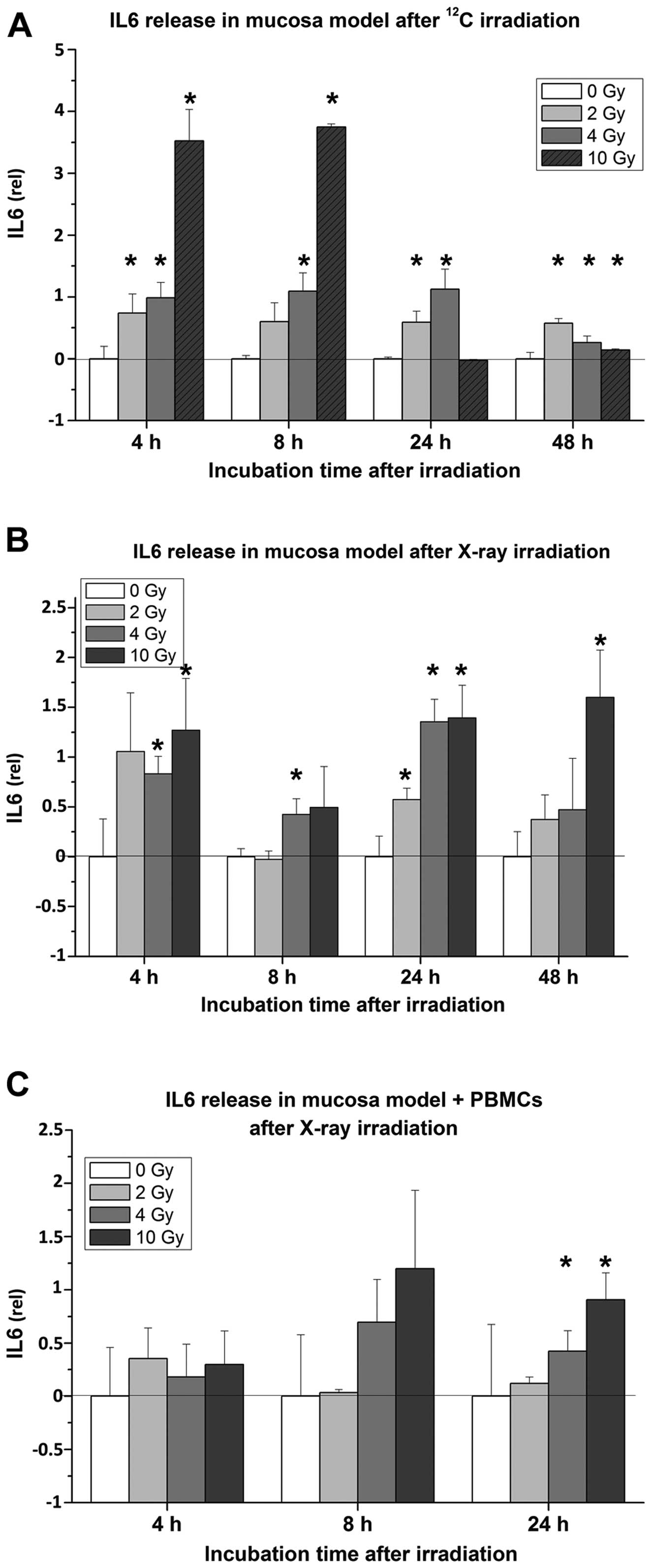
Radiation impact on IL6 and IL8
release
The 3D organotypic mucosa models immediately
responded to radiation of both qualities by secretion of cytokines
and chemokines, but there were still several differences in
cytokine release. Irradiation with the particle beam caused the
strongest effect after 4 and 8 h, while 24 h after exposure to 10
Gy, the levels of IL6 decreased, as depicted in Fig. 5A. The contrary effect was observed
in mucosa models exposed to X-rays, where the levels of IL6
increased over time and reached the maximum 24 h after treatment
(Fig. 5B). Co-cultures containing
immune cells revealed similar responses to X-ray treatment, but
significant increase of IL6 levels were detected only after 24 h
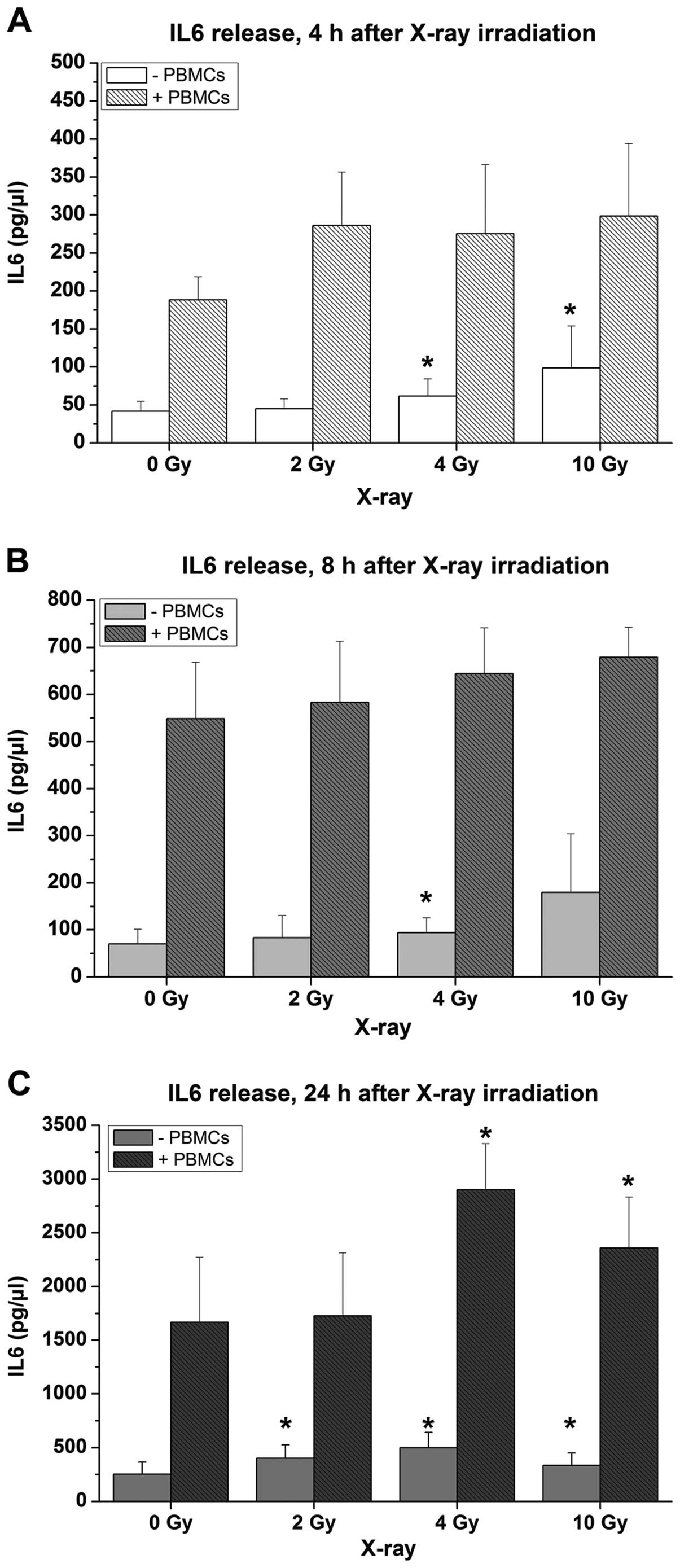
(Fig. 5C). Here, 2–3-fold higher
IL6 concentrations compared to mucosa models without PBMCs were
observed (Fig. 6).
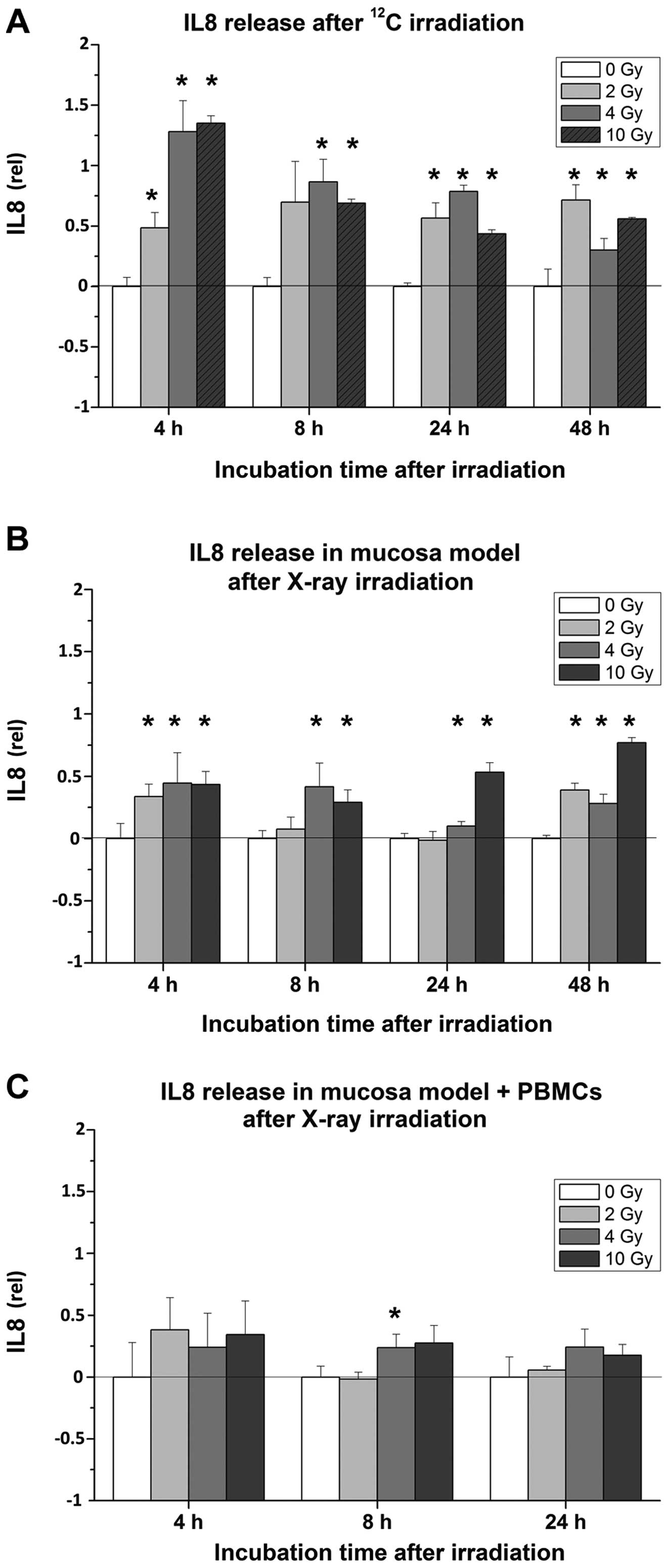
After irradiation with 12C particles,
organotypic cultures consistently released significantly more IL8,
compared to untreated samples, with the maximum after 4 h (Fig. 7A). Subsequently, the levels of IL8
decreased gradually over time. Mucosa models treated with X-rays
showed the strongest effects at a dose of 10 Gy and 48 h after
irradiation (Fig. 7B). Co-cultures
including PBMCs revealed a tendency to a higher release of IL8
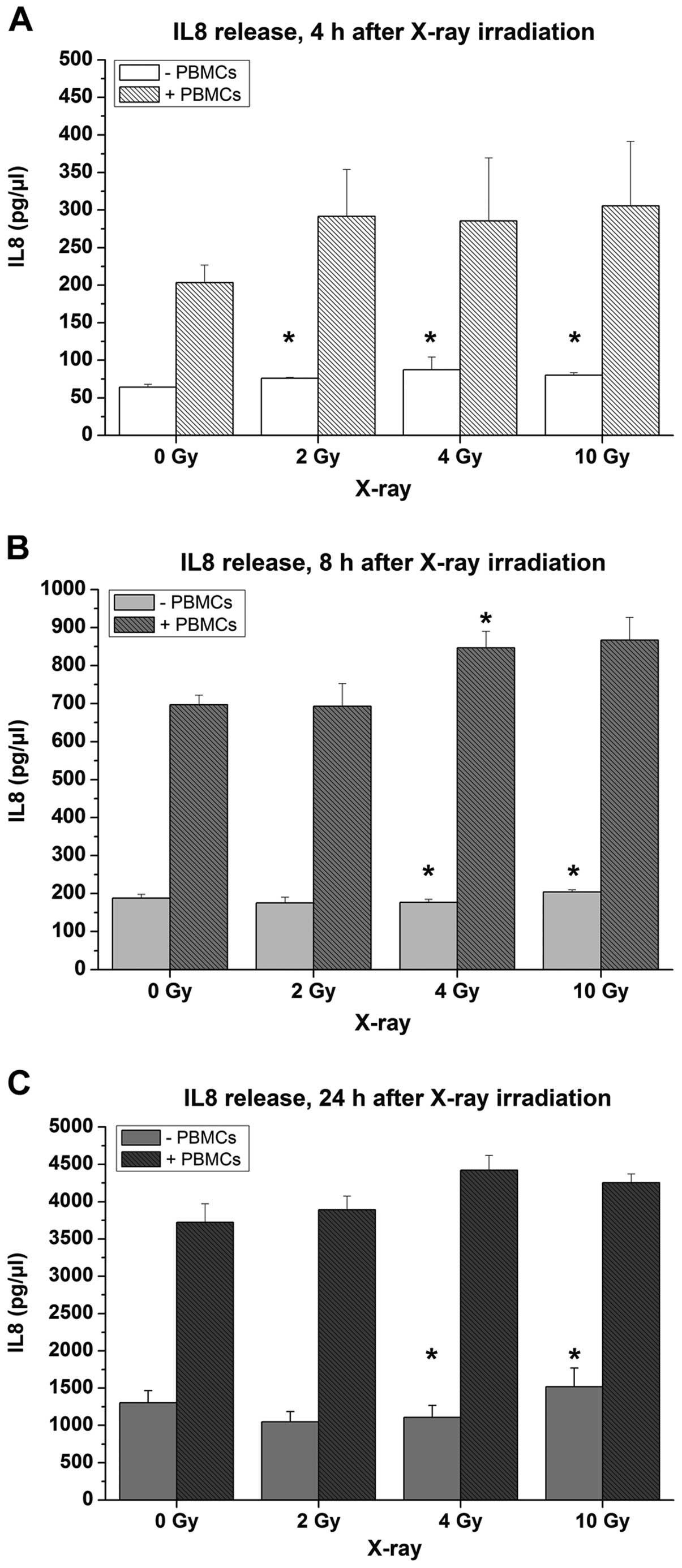
after X-ray irradiation compared to untreated samples (Fig. 7C) and 2–3-fold higher IL8
concentrations compared to mucosa models without PBMCs (Fig. 8).
Discussion
Although X-ray and heavy-ion radiotherapy have been
applied in cancer treatment, several adverse events restrict the
therapeutic success. Radiation-induced oral mucositis is a
well-documented painful side-effect of radiation therapy (6,7,9,11);
however, our understanding of the underlying cellular mechanisms is
limited. The present study investigated the radiation impact on
oral mucosa, using a 3D organotypic culture with and without human
immune cells, comparing X-rays and 12C heavy-ion
irradiation. In addition to the clinical aspects, the present study
assessed the risk of developing radiation-induced oral mucositis
during extended space flights.
Previous studies revealed increased cytokeratin
expression in the initial stage of oral mucositis in biopsies of
irradiated patients linked to the proliferation of epithelium cells
during the initial phases of radiotherapy (23). Biopsies were obtained from buccal
mucosa, when patients had already received a radiation dose of 36
Gy and had presented mucositis grade 1. In the present study,
however, irradiation of organotypic cultures with 12C
particles at a dose up to 4 Gy showed no effects on the proportion
of the epithelium within the 3D cultures, whereas X-ray irradiation
with up to 10 Gy appeared to reduce the epithelium. Furthermore,
12C ion irradiation caused a more pronounced loss of
compactness within the mucosa model cultures than X-ray
irradiation. These differences in the effect of 12C ions
and X-rays most likely result from the different physical qualities
of these radiation types.
It was also important to analyze the
radiation-induced DNA damage, since previous studies showed that
DSB may activate NFκB and the following signaling pathway by
mediator molecules such as ATM or DNA-dependent protein kinase
(17,24–26).
Analyses of radiation-induced DSB in our mucosa model revealed a
clear dose-dependent increase of γH2AX foci 4 h after irradiation
with carbon ions and X-rays. As a result of this DNA damage, the
foundation for induction of inflammatory response, such as
activation of NFκB p50 and the release of IL6 and IL8, was laid.
Subsequently, 48 h after irradiation, the DSB decreased to the
untreated level, although the incidence and severity of oral
mucositis is not necessarily correlated with DSB repair (10).
Activation of the transcription factor NFκB and
increased secretion of pro-inflammatory cytokines and chemokines
are involved not only in initiation of radiation-induced oral
mucositis (22,27), but also in linking inflammation to
cancer development and progression (14–17,28).
We observed carbon ion-induced activation of NFκB in the mucosa
model, which has been already documented for HEK cells (29). Cultures treated with X-rays showed a
distinct NFκB p50 translocation to the nucleus as well. Again, the
different qualities of radiation appear to affect mucosa cultures
in different ways following different kinetics. X-rays induced an
early activation of NFκB already 4 h after treatment, which
returned to control levels at 24 h after treatment. This is in
contrast to heavy-ion-induced effects, which reached their maximum
48 h after treatment.
Similar to other studies (30–32),
we observed an increased expression of IL6 and IL8 as a probable
response to preceding NFκB p50 activation. Notably, the analyses of
IL6 signal revealed a reverse behavior, compared to NFκB p50
activation. In the present study, carbon ion irradiation induced
the strongest effects 4 and 8 h after radiation, response to X-rays
was the highest 24 and 48 h after treatment. The kinetics of NFκB
activation and the IL6 release appear to occur with a time delay.
This may indicate a causative sequence of events. The
12C-induced segregation of the chemokine IL8 follows a
similar release profile as IL6. In contrast to IL6, the signal for
IL8 induced by X-rays reached its maximum at 8 h and decreased 24 h
after treatment. Co-cultures containing PBMCs responded
qualitatively similar to mucosa models without immune cells, but
showed much higher concentrations for IL6 and IL8, respectively,
which makes the mucosa model more sensitive for radiation-induced
responses. Since a correlation between increased expression of IL8
and carcinogenesis for several entities has been previously
described (15,16,33–35),
our results may indicate an increased risk of tumorigenesis for
astronauts during prolonged space missions.
In conclusion, reproducible inflammatory responses
induced by sparsely and densely ionizing radiation were detected as
a sign of the initiation stage of oral mucositis in 3D organotypic
mucosa models with and without immune cells. Furthermore, first
precursors of carcinogenesis were determined. Nevertheless, an
assessment of the risk of developing radiation-induced oral
mucositis during prolonged space flights proves to be rather
difficult, since the induction of inflammatory processes depends on
various determinants, such as the quality of the spacecraft
shielding or accumulation dose. To avoid these complications and to
ensure safety for crew members, ESA and other space agencies are
working on evaluating possible improvements in radiation shielding
approaches (3). For the consistent
reduction of risk, a cautious management and convenient timing for
such prolonged missions is required.
Acknowledgements
The present study was supported by the European
Space Agency (ESA), the German Aerospace Center (DLR), the Federal
Ministry of Economics and Technology (BMWi, no. 50 WB 0926), and
the GSI Helmholtz Center for Heavy Ion Research in Darmstadt. The
authors thank the laboratory of Dr Susanne Strand, University
Medical Center Mainz, particularly Dr Steffen Lorenz, for LSM
imaging and the GSI Biophysics accelerator team for the support
during beam times.
References
|
1
|
Hall EJ and Giaccia A: Radiobiology for
the Radiologist. 7th edition. Lippincott Williams & Wilkins;
Philadelphia, PA: 2012
|
|
2
|
Hawkey A: Physiological and biomechanical
considerations for a human Mars mission. J Br Interplanet Soc.
58:117–130. 2005.PubMed/NCBI
|
|
3
|
Cucinotta FA, Kim MHY and Chappell LJ:
Evaluating shielding approaches to reduce space radiation cancer
risks. NASA Center for AeroSpace Information; May. pp. 1–35.
2012
|
|
4
|
Cucinotta FA, Kim MHY and Ren L:
Evaluating shielding effectiveness for reducing space radiation
cancer risks. Radiat Meas. 41:1173–1185. 2006. View Article : Google Scholar
|
|
5
|
Durante M and Cucinotta FA: Heavy ion
carcinogenesis and human space exploration. Nat Rev Cancer.
8:465–472. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schulz-Ertner D, Haberer T, Scholz M, et
al: Acute radiation-induced toxicity of heavy ion radiotherapy
delivered with intensity modulated pencil beam scanning in patients
with base of skull tumors. Radiother Oncol. 64:189–195. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sonis ST and Fey EG: Oral complications of
cancer therapy. Oncology. 16:680–686. 2002.PubMed/NCBI
|
|
8
|
Vissink A, Jansma J, Spijkervet FK,
Burlage FR and Coppes RP: Oral sequelae of head and neck
radiotherapy. Crit Rev Oral Biol Med. 14:199–212. 2003. View Article : Google Scholar
|
|
9
|
Jensen AD, Nikoghosyan AV, Ecker S,
Ellerbrock M, Debus J and Münter MW: Carbon ion therapy for
advanced sinonasal malignancies: feasibility and acute toxicity.
Radiat Oncol. 6:302011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fleckenstein J, Kühne M, Seegmüller K, et
al: The impact of individual in vivo repair of DNA double-strand
breaks on oral mucositis in adjuvant radiotherapy of head-and-neck
cancer. Int J Radiat Oncol Biol Phys. 81:1465–1472. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sonis ST: Oral mucositis in head and neck
cancer: risk, biology, and management. Am Soc Clin Oncol Educ Book.
236–240. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Naidu MU, Ramana GV, Rani PU, Mohan IK,
Suman A and Roy P: Chemotherapy-induced and/or radiation
therapy-induced oral mucositis - complicating the treatment of
cancer. Neoplasia. 6:423–431. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hickok JT, Morrow GR, Roscoe JA, Mustian K
and Okunieff P: Occurrence, severity, and longitudinal course of
twelve common symptoms in 1129 consecutive patients during
radiotherapy for cancer. J Pain Symptom Manage. 30:433–442. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Porta C, Larghi P, Rimoldi M, Totaro MG,
Allavena P, Mantovani A and Sica A: Cellular and molecular pathways
linking inflammation and cancer. Immunobiology. 214:761–777. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Karin M and Greten FR: NF-κB: linking
inflammation and immunity to cancer development and progression.
Nat Rev Immunol. 5:749–759. 2005.
|
|
16
|
Karin M: NF-κB as a critical link between
inflammation and cancer. Cold Spring Harb Perspect Biol.
1:a0001412009.
|
|
17
|
Magné N, Toillon RA, Bottero V, Didelot C,
Houtte PV, Gérard JP and Peyron JF: NF-κB modulation and ionizing
radiation: mechanisms and future directions for cancer treatment.
Cancer Lett. 231:158–168. 2006.
|
|
18
|
Boxman IL, Ruwhof C, Boerman OC, Löwik CW
and Ponec M: Role of fibroblasts in the regulation of
proinflammatory interleukin IL-1, IL-6 and IL-8 levels induced by
keratinocyte-derived IL-1. Arch Dermatol Res. 288:391–398. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sawicki G, Marcoux Y, Sarkhosh K, Tredget
E and Ghahary A: Interaction of keratinocytes and fibroblasts
modulates the expression of matrix metalloproteinases-2 and -9 and
their inhibitors. Mol Cell Biochem. 269:209–216. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang Z, Wang Y, Farhangfar F, Zimmer M and
Zhang Y: Enhanced keratinocyte proliferation and migration in
co-culture with fibroblasts. PLoS One. 7:e409512012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Werner S, Krieg T and Smola H:
Keratinocyte-fibroblast interactions in wound healing. J Invest
Dermatol. 127:998–1008. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sonis ST: The pathobiology of mucositis.
Nat Rev Cancer. 4:277–284. 2004. View
Article : Google Scholar
|
|
23
|
Bonan PRF, Kaminagakura E, Pires FR,
Vargas PA and de Almeida OP: Cytokeratin expression in initial oral
mucositis of head and neck irradiated patients. Oral Surg Oral Med
Oral Pathol Oral Radiol Endod. 101:205–211. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Basu S, Rosenzweig KR, Youmell M and Price
BD: The DNA-dependent protein kinase participates in the activation
of NFκB following DNA damage. Biochem Biophys Res Commun.
247:79–83. 1998.
|
|
25
|
Lee SJ, Dimtchev A, Lavin MF, Dritschilo A
and Jung M: A novel ionizing radiation-induced signaling pathway
that activates the transcription factor NF-κB. Oncogene.
17:1821–1826. 1998.PubMed/NCBI
|
|
26
|
Piret B, Schoonbroodt S and Piette J: The
ATM protein is required for sustained activation of NF-κB following
DNA damage. Oncogene. 18:2261–2271. 1999.PubMed/NCBI
|
|
27
|
Sonis ST: New thoughts on the initiation
of mucositis. Oral Dis. 16:597–600. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ben-Neriah Y and Karin M: Inflammation
meets cancer, with NF-κB as the matchmaker. Nat Immunol.
12:715–723. 2011.
|
|
29
|
Hellweg CE, Baumstark-Khan C, Schmitz C,
et al: Carbon-ion-induced activation of the NF-κB pathway. Radiat
Res. 175:424–431. 2011.PubMed/NCBI
|
|
30
|
Colley HE, Eves PC, Pinnock A, Thornhill
MH and Murdoch C: Tissue-engineered oral mucosa to study
radiotherapy-induced oral mucositis. Int J Radiat Biol. 89:907–914.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tobita T, Izumi K and Feinberg SE:
Development of an in vitro model for radiation-induced effects on
oral keratinocytes. Int J Oral Maxillofac Surg. 39:364–370. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lambros MP, Parsa C, Mulamalla H, et al:
Identifying cell and molecular stress after radiation in a
three-dimensional (3-D) model of oral mucositis. Biochem Biophys
Res Commun. 405:102–106. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Brat DJ, Bellail AC and Van Meir EG: The
role of interleukin-8 and its receptors in gliomagenesis and
tumoral angiogenesis. Neuro Oncol. 7:122–133. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xie K: Interleukin-8 and human cancer
biology. Cytokine Growth Factor Rev. 12:375–391. 2001. View Article : Google Scholar
|
|
35
|
Yin Y, Si X, Gao Y, Gao L and Wang J: The
nuclear factor-κB correlates with increased expression of
interleukin-6 and promotes progression of gastric carcinoma. Oncol
Rep. 29:34–38. 2013.
|