Introduction
The presence of lymphatic metastasis to regional
lymph node is particularly important for cancer prognosis in common
human cancers, including gastric, colon and breast adenocarcinomas
(1). Thus, the lymphatic vascular
system correlates closely with tumor metastasis (2,3). The
recent identification of lymphatic endothelial specific markers,
such as hyaluronic acid receptor-1 (LYVE-1) (4) and podplanin (5), has greatly increased attention on how
lymphangiogenesis, the growth of lymphatic vessels, is regulated in
the tumor microenvironment. Tumor-induced lymphangiogenesis is
mediated by lymphangiogenic growth factors (2,3). The
involvement of vascular endothelial growth factor (VEGF)-C and
VEGF-D in cancer progression and in tumor-associated lymphatic
vessel growth has been demonstrated in several experimental systems
(6–8). A role for VEGF-A in tumor-mediated
lymphangiogenesis has also been reported (9,10). We
have developed a tumor xenograft experimental model using chimeric
nude mice with green fluorescent protein (GFP)-positive bone marrow
cells (11). In this xenograft
model, we showed that human gastric cancer MKN45 cells induced
tumor lymphangiogenesis via recruitment of lymphatic endothelial
progenitor cells from bone marrow. In the present study, using the
same xenograft model, human colorectal cancer LS174 and human
breast cancer SK-BR-3 cells also promoted lymphangiogenesis and the
recruitment of lymphatic endothelial progenitor cells from bone
marrow. These three human adenocarcinoma (gastric cancer MKN45,
colorectal cancer LS174 and breast cancer SK-BR-3) cells
predominantly produced VEGF-A with negligible secretion of VEGF-C
and VEGF-D. VEGF-A, as well as well-known lymphangiogenic factors,
VEGF-C and VEGF-D (2,3), induced the expression of LYVE-1, a
specific lymphatic endothelial marker, in bone marrow mononuclear
cells in culture. The present study shows the role of VEGF-A,
produced by adenocarcinomas, in the tumor lymphangiogenesis via
recruitment of lymphatic progenitor cells from bone marrow.
Materials and methods
Materials
LS174T colorectal cancer (12) and SK-BR-3 breast cancer cells
(13) were obtained from American
Type Culture Collection (ATCC) (Manassas, VA, USA). MKN45 gastric
cancer cells (14) were from RIKEN
BioResource Center (Tsukuba, Japan). C57BL/6J and
C57BL/6-Tg-CAG-EGFP mice were from SLC (Shizuoka, Japan).
CAnN.Cg-Foxn1nu/CrlCrlj nude mice were from Charles
River Laboratories (Yokohama, Japan). ELISA kits and recombinant
human VEGFs were from R&D Systems (Minneapolis, MN, USA).
Antibody against mouse LYVE-1 was from Santa Cruz Biotechnology
(Santa Cruz, CA, USA). Anti-mouse CD34 antibody was from Hycult
Biotech (Uden, The Netherlands). Alexa Fluor 594-conjugated
anti-rabbit secondary antibody, SuperScript VILO cDNA Synthesis kit
and 4′,6-diamidino-2-phenylindole (DAPI) were from Invitrogen
(Carlsbad, CA, USA). Histopaque-1083 was from Sigma (St. Louis, MO,
USA). PCR primers and SYBR Premix Ex Taq II were from Takara Bio,
Inc. (Osaka, Japan). RNeasy Plus Mini kit was from Qiagen (Tokyo,
Japan). Type I collagen-coated dishes were from Iwaki (Tokyo,
Japan). Tissue-Tek OCT compound was from Sakura Finetek (Tokyo,
Japan). EBM-2 medium was from Lonza (Walkersville, MD, USA).
RPMI-1640 medium, antibiotic-antimycotic and fetal bovine serum
(FBS) were from Life Technologies (Grand Island, NY, USA). Other
chemicals were of the highest quality available.
Culture of human adenocarcinoma
cells
MKN45 gastric cancer, LS174T colorectal cancer and
SK-BR-3 breast cancer cells were cultured in RPMI-1640 medium
supplemented with 10% FBS and 1% antibiotic-antimycotic.
ELISA assay
The culture media of human adenocarcinoma cells were
collected after 72 h in culture. Contents of VEGF-A, VEGF-C,
VEGF-D, epidermal growth factor (EGF) and hepatocyte growth factor
(HGF) were determined by species-specific ELISA kits as previously
described (15).
Bone marrow-transplanted chimera
mice
All animal procedures were carried out according to
a protocol approved by the Animal Care and Use Committee of Gifu
University Graduate School of Medicine. GFP-positive bone marrow
cells were obtained from age-matched 6 week-old C57BL/6-Tg-CAG-EGFP
mice by flushing the femurs and tibias with Hanks’ balanced salt
solution, and mononuclear cells were isolated by density gradient
centrifugation using Histopaque-1083 (16). The recipient nude mice,
CAnN.Cg-Foxn1nu/CrlCrlj, were lethally irradiated with
8.0 Gy, and received 6×106 donor GFP-positive bone
marrow mononuclear cells intravenously (11). Bone marrow cells from 6 week-old
C57BL/6J mice were used as a negative control for GFP-positive
donor cells. At 4 weeks after bone marrow transplantation, mice
were used for tumor implantation models. To determine the
transplantation efficiency of GFP-positive bone marrow, tail
peripheral blood (10 μl) or bone marrow from recipient nude mice
was collected 3 weeks after tumor implantation, and GFP-positive
cells spread on cover slides were analyzed by fluorescent
microscopy.
Murine tumor model
At four weeks after bone marrow transplantation,
2×106 cells were subcutaneously injected into chimeric
nude mice. At 3–8 weeks after tumor cell injection, mice were
anesthetized, perfused with 0.1 M phosphate-buffered saline (PBS;
pH 7.40) followed by 2% paraformaldehyde (PFA) in PBS and
sacrificed. The tumor area was removed, fixed for 2 h with 2% PFA
in PBS, and stored frozen in OCT compound (11).
Immunofluorescent staining
The cryostat sections (6 μm) of tumor tissues were
immunostained with the primary anti-body, rabbit anti-mouse LYVE-1
(1:100). The sections were next incubated with Alexa Fluor
594-conjugated anti-rabbit secondary antibody (1:200) and DAPI.
Images were analyzed using a BIOREVO immunofluorescence microscope
(Keyence, Osaka, Japan). To evaluate lymphatic vessel density or
the proportion of GFP/LYVE-1 double-positive cells, we used the
inverted-gray scale images of all data (nine fields in each
sections), and calculated the total area positive for LYVE-1 or the
area double-positive for GFP/LYVE-1 using NIH ImageJ software. To
elucidate the histological structure positively stained with LYVE-1
or GFP, light hematoxylin staining was performed on the sections
near the immunostained sections. Each result was obtained in at
least 4 separate experiments.
Bone marrow cell culture and
immunofluorescent cell staining
Bone marrow mononuclear cells were collected from
C57BL/6J mice as described above. Cells plated onto type 1
collagen-coated dishes and cultured in EBM-2 medium in the absence
or presence of recombinant human VEGF-A (50 ng/ml), human VEGF-C
(150 ng/ml) or mouse VEGF-D (150 ng/ml). For immunofluorescent
staining, cells were fixed with 2% PFA for 20 min at room
temperature and were stained with rabbit anti-mouse LYVE-1 (1:100)
primary antibody for 2 h at room temperature. Cells were next
incubated with Alexa Fluor 594-conjugated anti-rabbit secondary
antibody (1:200) at room temperature for 2 h, and also
counterstained with DAPI for 2 min. Images were analyzed using a
BIOREVO immunofluorescence microscope.
Quantitative real-time RT-PCR
Total RNA was extracted from bone marrow mononuclear
cells using an RNeasy Plus Mini kit and 0.5 μl of RNA was reverse
transcribed to cDNA using a SuperScript VILO cDNA Synthesis kit.
cDNA was amplified with SYBR Premix EX Taq II and the following
primers: murine LYVE-1 (sense, 5′-CAAAGCCTATTGCCACAACTCATC-3′ and
antisense, 5′-AGTAGGCGCTGCTGACA-3′) and mouse ACTB/β-actin.
ACTB/β-actin transcripts were measured simultaneously in all
reactions as internal controls. Up to 40 PCR cycles with an
annealing temperature of 60°C were performed for cDNA
amplification. Relative expression values were calculated from the
cycle threshold (Ct) as previously described (17).
Statistical analysis
Each result was obtained in at least 5 separate
experiments. All values are expressed as means ± SEM. Values were
analyzed using one-way analysis of variance, and then the
significance of differences in multiple comparisons was determined
using Scheffe’s multiple comparison test. P<0.01 was considered
statistically significant.
Results
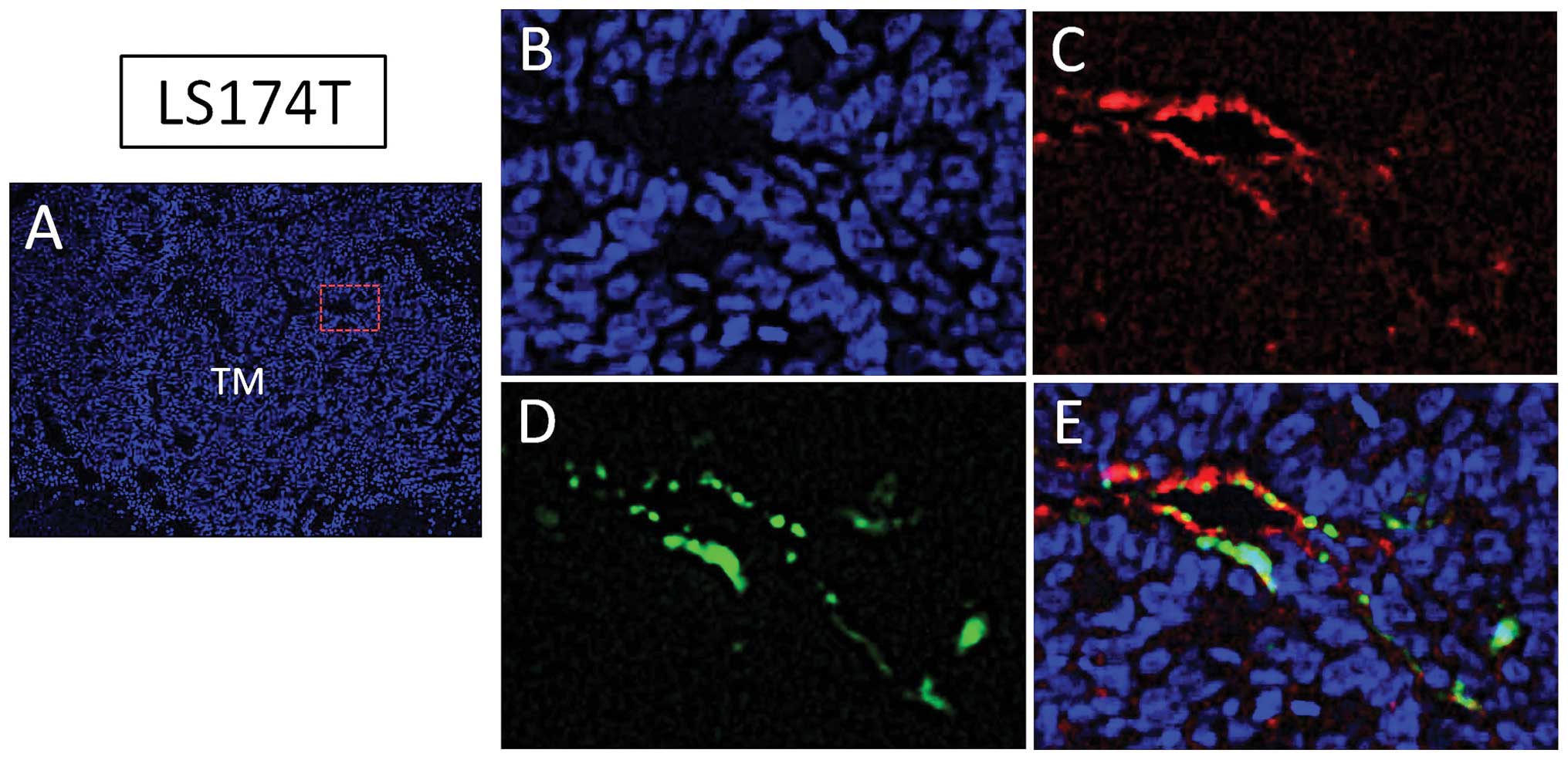
Human colorectal cancer LS174T cells
promote lymphangiogenesis and incorporation of bone marrow-derived
cells in tumor lymphatics
We developed a tumor xenograft experimental model
using chimeric nude mice with GFP-positive bone marrow cells in
order to identify the contribution of marrow-derived cells in tumor
lymphangiogenesis (11). Human
colorectal cancer LS174T cells were subcutaneously implanted in
this chimeric nude mouse model. LS174T cells formed tumor mass
(Fig. 1A) three weeks after
inoculation and a tumor lymphatic-rich microenvironment as detected
by LYVE-1 (Fig. 1B and C).
LYVE-1-positive tumor lymphatics were located not only in
peritumoral tissues but also in intratumoral tissue. Consistent
with the locations of tumor lymphatics, GFP-positive cells from
bone marrow were detected in peritumoral and intratumoral (Fig. 1D) tissues. Nearly 60% of
LYVE-1-positive cells co-expressed GFP (Fig. 1E). These results suggest the
possible involvement of bone marrow-derived cells in tumor
lymphangiogenesis.
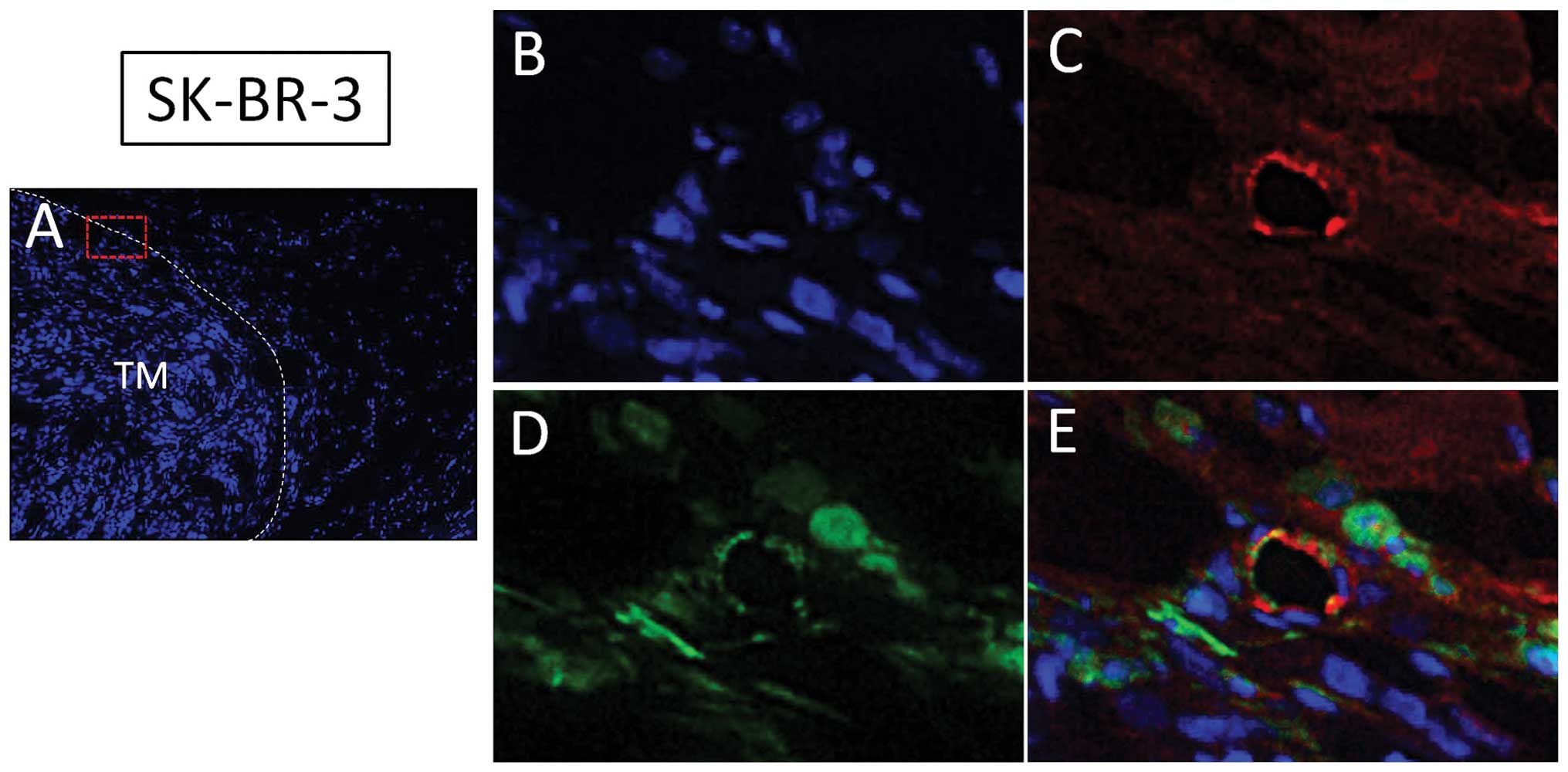
Human breast cancer SK-BR-3 cells promote
lymphangiogenesis and incorporation of bone marrow-derived cells in
tumor lymphatics
In the same chimeric nude mouse model, SK-BR-3 human
breast cancer cells were subcutaneously injected. At 8 weeks after
inoculation, SK-BR-3 human breast cancer cells formed a tumor mass
(Fig. 2A) and a tumor
lymphatic-rich microenvironment as detected by LYVE-1 (Fig. 2B and C). GFP-positive cells from
bone marrow were also located at peritumoral lymphatic tissues
(Fig. 2D). Nearly 50% of
LYVE-1-positive cells co-expressed GFP (Fig. 2E). These data also support the
hypothesis that bone marrow-derived lymphatic endothelial
progenitor cells are involved in the tumor lymphangiogenesis in
addition to pre-existing lymphatics.
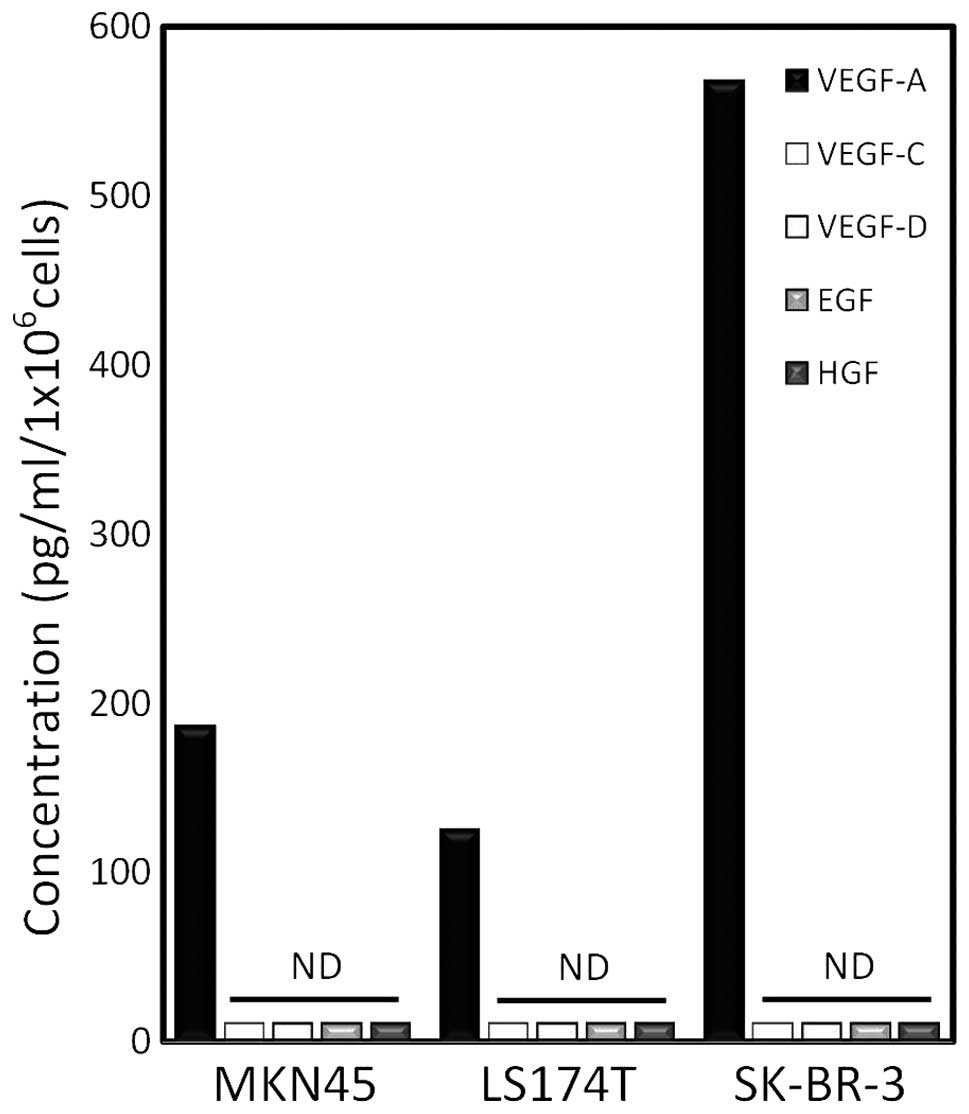
Human malignant adenocarcinoma cells
predominantly secrete VEGF-A
In order to identify the growth factor(s) involved
in the lymphangiogenesis in the peritumoral tissues, human gastric
cancer MKN45, colorectal cancer LS174 and breast cancer SK-BR-3
cells were cultured in vitro and growth factors in the
culture medium were measured by specific ELISA assay. Among the
growth factors tested, these three cell lines predominantly
produced VEGF-A at 72 h after incubation (Fig. 3). The well-characterized
lymphangiogenic factors (2,3,6–8),
VEGF-C and VEGF-D, were below the detectable levels in the present
ELISA assay. Furthermore, secretion of other growth factors, which
are implicated in lymphangiogenesis and angiogenesis (18,19),
EGF and HGF, was not detected in these cells.
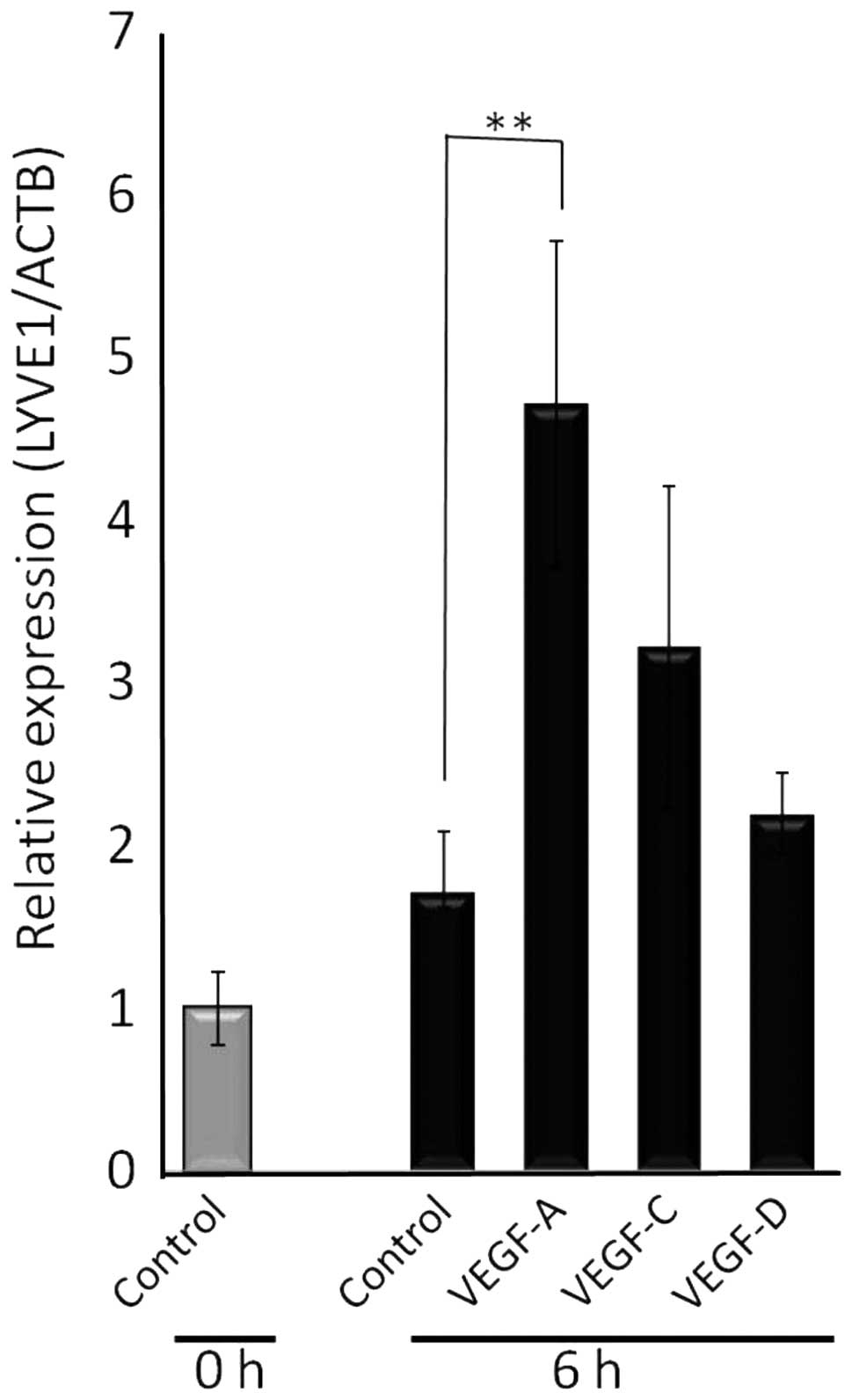
VEGF-A induces differentiation of bone
marrow mononuclear cells into lymphatic progenitor cells
The present and previous (11) results suggest that VEGF-A-rich human
adenocarcinomas recruit lymphatic epithelial progenitor cells from
bone marrow. We further examined whether VEGF-A directly induced
the differentiation of bone marrow mononuclear cells into lymphatic
epithelial progenitor cells. First, bone marrow mononuclear cells
were cultured in the absence or presence of VEGF family proteins
and the relative expression of LYVE-1, a specific lymphatic
endothelial marker, was analyzed. VEGF-A significantly increased
LYVE-1 mRNA in bone marrow cells at 6 h after administration
(Fig. 4). At the same time, VEGF-C
and VEGF-D tended to increase LYVE-1 mRNA, yet the increase was not
statistically significant. To further confirm VEGF-A-dependent
differentiation of bone marrow mononuclear cells into lymphatic
endothelial lineage cells, bone marrow cells were incubated with
VEGF-A and immunostained with antibody against LYVE-1. In the
control culture without VEGF family proteins, most of the attached
cells were positive for CD34, a marker of hematopoietic and
vascular endothelial progenitor cells (data not shown), but
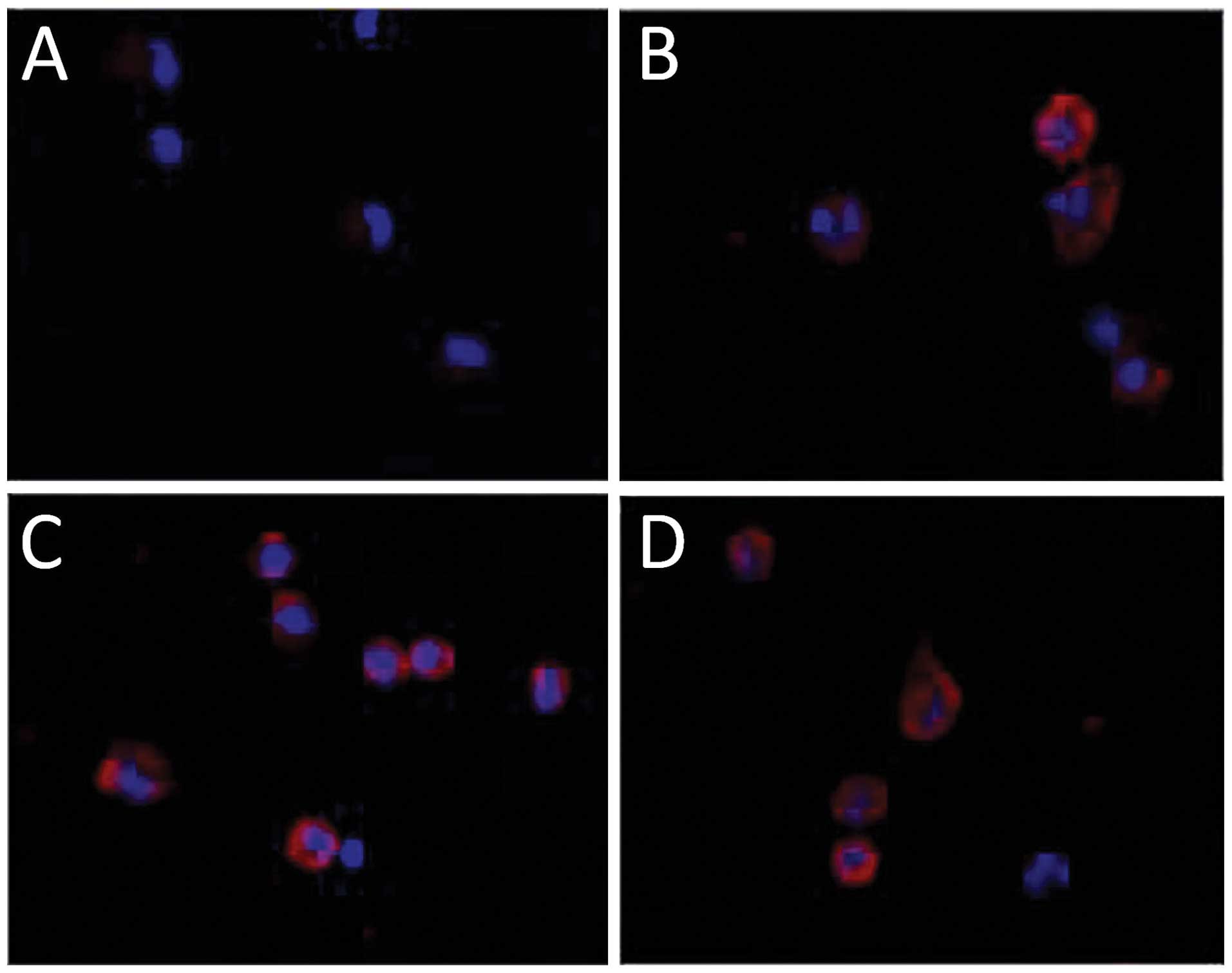
negative for LYVE-1 (Fig. 5A). The
well-characterized lymphangiogenic factors, VEGF-C (Fig. 5B) and VEGF-D (Fig. 5C), stimulated the expression of
LYVE-1 in bone marrow mononuclear cells in this analysis system
after incubation for 40 h; 50–60% cells were positive for LYVE-1.
VEGF-A also clearly induced the increase of LYVE-1 positive bone
marrow mononuclear cells (Fig. 5D).
Nearly 60% of cells became positive for LYVE-1 at 40 h after
incubation. These results suggest the differentiation of bone
marrow mononuclear cells into lymphatic endothelial lineage cells
in response to VEGF-A.
Discussion
The formation of a lymphatic vascular system is a
dynamic process during embryogenesis. In embryos, the blood
circulatory system is first to evolve, followed by specification of
lymphatic endothelial progenitor cells differentiated from blood
vascular endothelial cells and budding of these cells from the
cardinal vain (20). Under normal
physiological postnatal conditions, however, the formation of new
lymphatic vessels scarcely occurs. In adults, lymphangiogenesis
only takes place during certain pathological conditions such as
inflammation, tissue repair and tumor growth. Postnatal lymphatic
vessel formations have been assumed to occur mainly by hyperplasia
of lymphatic vessels themselves, or sprouting from preexisting
lymphatics. The relative contribution to the formation of new
vessels from circulating endothelial progenitor cells is not yet
clearly understood. Integration of bone marrow-derived cells into
tumor lymphatics was initially denied in a mouse xenograft model
with Lewis lung carcinoma or B16-F1 melanoma cells (21). In contrast, the involvement of bone
marrow-derived cells in lymphatic vessel formation has been
suggested in human kidney transplants (22) and mouse corneal lymphangiogenesis
model (23). Moreover, the role of
bone marrow progenitor cells in tumor lymphangiogenesis has
recently been reported (23–25).
We have previously shown that bone marrow-derived cells are
involved in the formation of tumor lymphatics in a mouse xenograft
model with human gastric cancer MKN45 cells (11). In the present study, we showed that
in the same experimental model with chimeric nude mice bearing
GFP-positive bone marrow, human colorectal cancer LS174T and breast
cancer SK-BR-3 cells also recruited bone marrow-derived cells in
the formation of tumor lymphatics. These results indicate that bone
marrow-derived lymphatic endothelial progenitor cells participate
in the human adenocarcinoma-induced lymphangiogenesis in addition
to pre-existing lymphatics.
Several factors with prolymphangiogenic activity
have been identified to date (2,3,20).
These include VEGF-A, VEGF-C, VEGF-D and HGF. Attention has been
focused on the role of VEGF-C and VEGF-D in recent years (2,3). Using
mouse models, overexpression of VEGF-C or VEGF-D has been shown to
increase lymphatic vessel density, vessel diameter and lymph node
and organ metastasis of many types of cancer (6–8). A
role for VEGF-A in tumor-mediated lymphangiogenesis and metastasis
has also been reported (9,10). In the present study, three malignant
adenocarcinomas examined predominantly secreted VEGF-A with minimal
productions of VEGF-C and VEGF-D. Various types of human cancer
cells extensively secrete VEGF-A and this ability is often
associated with poor prognosis (26–29).
VEGF-A is a key factor of new blood vessel formation (angiogenesis)
to support tumor growth and metastatic dissemination (26,27).
Moreover, recent reports have demonstrated the immunosuppressive
effects of VEGF-A by induction of regulatory T cells (29,30)
and by reduction of cytotoxic T cell activity (31).
Despite its initial recognition as a master
regulatory molecule in angiogenesis (26,27),
previous studies indicated that VEGF-A also plays a role in
lymphangiogenesis in tumor (8,9),
inflammation (33,34) and wound healing (35) models. VEGF-A may indirectly induce
lymphangiogenesis via recruitment of VEGF-C/D producing macrophages
(32). The role of VEGF-A appears
to vary depending on the tissue microenvironment (9,10,34,35).
The data obtained in the present study show that the expression of
a lymphatic endothelial-specific marker, LYVE-1, increased in in
vitro culture of mouse bone marrow mononuclear cells in the
presence of VEGF-A as assessed by immunostaining as well as
real-time PCR. These results indicate that VEGF-A is able to
directly promote differentiation of bone marrow mononuclear cells
into lymphatic endothelial lineage cells.
In summary, the present study indicated that human
malignant adenocarcinoma cells, which extensively secrete VEGF-A,
promote tumor lymphangiogenesis in part via recruitment and
incorporation of bone marrow-derived lymphatic endothelial
progenitor cells into tumor lymphatics. The mechanism involves
direct effect of VEGF-A on the differentiation of bone marrow
mononuclear cells into lymphatic lineage cells.
Acknowledgements
This study was supported in part by an NaSNeLC
research grant from Gifu University Graduate School of Medicine.
The authors thank Ryoko Hayashi for her technical assistance.
Abbreviations:
|
EGF
|
epidermal growth factor
|
|
GFP
|
green fluorescent protein
|
|
HGF
|
hepatocyte growth factor
|
|
LYVE-1
|
hyaluronic acid receptor-1
|
|
PFA
|
paraformaldehyde
|
|
VEGF
|
vascular endothelial growth factor
|
|
DAPI
|
4′,6-diamidino-2-phenylindole
|
References
|
1
|
Achen MG, Mann GB and Stacker SA:
Targeting lymphangiogenesis to prevent tumour metastasis. Br J
Cancer. 94:1355–1360. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Christiansen A and Detmar M:
Lymphangiogenesis and cancer. Genes Cancer. 2:1146–1158. 2011.
View Article : Google Scholar
|
|
3
|
Alitalo A and Detmar M: Interaction of
tumor cells and lymphatic vessels in cancer progression. Oncogene.
31:4499–4508. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Banerji S, Ni J, Wang SX, Clasper S, Su J,
Tammi R, Jones M and Jackson DG: LYVE-1, a new homologue of the
CD44 glycoprotein, is a lymph-specific receptor for hyaluronan. J
Cell Biol. 144:789–801. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Breiteneder-Geleff S, Soleiman A, Kowalski
H, Horvat R, Amann G, Kriehuber E, Diem K, Weninger W, Tschachler
E, Alitalo K and Kerjaschki D: Angiosarcomas express mixed
endothelial phenotypes of blood and lymphatic capillaries:
podoplanin as a specific marker for lymphatic endothelium. Am J
Pathol. 154:385–394. 1999. View Article : Google Scholar
|
|
6
|
Mandriota SJ, Jussila L, Jeltsch M,
Compagni A, Baetens D, Prevo R, Banerji S, Huarte J, Montesano R,
Jackson DG, Orci L, Alitalo K, Christofori G and Pepper MS:
Vascular endothelial growth factor-C-mediated lymphangiogenesis
promotes tumour metastasis. EMBO J. 20:672–682. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Stacker SA, Caesar C, Baldwin ME, Thornton
GE, Williams RA, Prevo R, Jackson DG, Nishikawa S, Kubo H and Achen
MG: VEGF-D promotes the metastatic spread of tumor cells via the
lymphatics. Nat Med. 7:186–191. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Skobe M, Hawighorst T, Jackson DG, Prevo
R, Janes L, Velasco P, Riccardi L, Alitalo K, Claffey K and Detmar
M: Induction of tumor lymphangiogenesis by VEGF-C promotes breast
cancer metastasis. Nat Med. 7:192–198. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nagy JA, Vasile E, Feng D, Sundberg C,
Brown LF, Detmar MJ, Lawitts JA, Benjamin L, Tan X, Manseau EJ,
Dvorak AM and Dvorak HF: Vascular permeability factor/vascular
endothelial growth factor induces lymphangiogenesis as well as
angiogenesis. J Exp Med. 196:1497–1506. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hirakawa S, Kodama S, Kunstfeld R, Kajiya
K, Brown LF and Detmar M: VEGF-A induces tumor and sentinel lymph
node lymphangiogenesis and promotes lymphatic metastasis. J Exp
Med. 201:1089–1099. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tawada M, Hayashi S, Osada S, Nakashima S
and Yoshida K: Human gastric cancer organizes neighboring lymphatic
vessels via recruitment of bone marrow-derived lymphatic
endothelial progenitor cells. J Gastroenterol. 47:1057–1060. 2012.
View Article : Google Scholar
|
|
12
|
Guadagni F, Witt PL, Robbins PF, Schlom J
and Greiner JW: Regulation of carcinoembryonic antigen expression
in different human colorectal tumor cells by interferon-γ. Cancer
Res. 50:6248–6255. 1990.
|
|
13
|
Imai Y, Leung CK, Friesen HG and Shiu RP:
Epidermal growth factor receptors and effect of epidermal growth
factor on growth of human breast cancer cells in long-term tissue
culture. Cancer Res. 42:4394–4398. 1982.PubMed/NCBI
|
|
14
|
Koshikawa N, Yasumitsu H, Umeda M and
Miyazaki K: Multiple secretion of matrix serine proteinases by
human gastric carcinoma cell lines. Cancer Res. 52:5046–5053.
1992.PubMed/NCBI
|
|
15
|
Hayashi S, Morishita R, Nakamura S,
Yamamoto K, Moriguchi A, Nagano T, Taiji M, Noguchi H, Matsumoto K,
Nakamura T, Higaki J and Ogihara T: Potential role of hepatocyte
growth factor, a novel angiogenic growth factor, in peripheral
arterial disease: downregulation of HGF in response to hypoxia in
vascular cells. Circulation. 100(Suppl 19): II301–II308. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Asahara T, Masuda H, Takahashi T, Kalka C,
Pastore C, Silver M, Kearne M, Magner M and Isner JM: Bone marrow
origin of endothelial progenitor cells responsible for postnatal
vasculogenesis in physiological and pathological
neovascularization. Cir Res. 85:221–228. 1999. View Article : Google Scholar
|
|
17
|
Hayashi S, Sato N, Yamamoto A, Ikegame Y,
Nakashima S, Ogihara T and Morishita R: Alzheimer
disease-associated peptide, amyloid β40, inhibits vascular
regeneration with induction of endothelial autophagy. Arterioscler
Thromb Vasc Biol. 29:1909–1915. 2009.
|
|
18
|
Kajiya K, Hirakawa S, Ma B, Drinnenberg I
and Detmar M: Hepatocyte growth factor promotes lymphatic vessel
formation and function. EMBO J. 24:2885–2895. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hogan BM, Bos FL, Bussmann J, Witte M, Chi
NC, Duckers HJ and Schulte-Merker S: ccbe1 is required for
embryonic lymphangiogenesis and venous sprouting. Nat Genet.
41:396–398. 2009. View
Article : Google Scholar
|
|
20
|
Alitalo K: The lymphatic vasculature in
disease. Nat Med. 17:1371–1380. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
He Y, Rajantie I, Ilmonen M, Makinen T,
Karkkainen MJ, Haiko P, Salven P and Alitalo K: Preexisting
lymphatic endothelium but not endothelial progenitor cells are
essential for tumor lymphangiogenesis and lymphatic metastasis.
Cancer Res. 64:3737–3740. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kerjaschki D, Huttary N, Raab I, Regele H,
Bojarski-Nagy K, Bartel G, Kröber SM, Greinix H, Rosenmaier A,
Karlhofer F, Wick N and Mazal PR: Lymphatic endothelial progenitor
cells contribute to de novo lymphangiogenesis in human renal
transplants. Nat Med. 12:230–234. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Maruyama K, Ii M, Cursiefen C, Jackson DG,
Keino H, Tomita M, Van Rooijen N, Takenaka H, D’Amore PA,
Stein-Streilein J, Losordo DW and Streilein JW:
Inflammation-induced lymphangiogenesis in the cornea arises from
CD11b-positive macrophages. J Clin Invest. 115:2363–2372. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Religa P, Cao R, Bjorndahl M, Zhou Z, Zhu
Z and Cao Y: Presence of bone marrow-derived circulating progenitor
endothelial cells in the newly formed lymphatic vessels. Blood.
106:4184–4190. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zumsteg A, Baeriswyl V, Imaizumi N,
Schwendener R, Ruegg C and Christofori G: Myeloid cells contribute
to tumor lymphangiogenesis. PLos One. 4:e70672009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ferrara N: VEGF and the quest for tumour
angiogenesis factors. Nat Rev Cancer. 2:795–803. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hicklin DJ and Ellis LM: Role of the
vascular endothelial growth factor pathway in tumor growth and
angiogenesis. J Clin Oncl. 23:1011–1027. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lyden D, Hattori K, Dias S, Costa C,
Blaikie P, Butros L, Chadburn A, Heissig B, Marks W, Witte L, Wu Y,
Hicklin D, Zhu Z, Hackett NR, Crystal RG, Moore MA, Hajjar KA,
Manova K, Benezra R and Rafii S: Impaired recruitment of
bone-marrow-derived endothelial and hematopoietic precursor cells
blocks tumor angiogenesis and growth. Nat Med. 7:1194–1201. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wada J, Suzuki H, Fuchino R, Yamasaki A,
Nagai S, Yanai K, Koga K, Nakamura M, Tanaka M, Morisaki T and
Katano M: The contribution of vascular endothelial growth factor to
the induction of regulatory T-cells in malignant effusions.
Anticancer Res. 29:881–888. 2009.PubMed/NCBI
|
|
30
|
Li B, Lalani AS, Harding TC, Luan B,
Koprivnikar K, Huan Tu G, Prell R, VanRoey MJ, Simmons AD and Jooss
K: Vascular endothelial growth factor blockade reduces intratumoral
regulatory T cells and enhances the efficacy of a GM-CSF-secreting
cancer immunotherapy. Clin Cancer Res. 12:6808–6816. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gavalas NG, Tsiatas M, Tsitsilonis O,
Politi E, Ioannou K, Ziogas AC, Rodolakis A, Vlahos G, Thomakos N,
Haidopoulos D, Terpos E, Antsaklis A, Dimopoulos MA and Bamias A:
VEGF directly suppresses activation of T cells from ascites
secondary to ovarian cancer via VEGF receptor type 2. Br J Cancer.
107:1869–1875. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cursiefen C, Chen L, Borges LP, Jackson D,
Cao J, Radziejewski C, D’Amore PA, Dana MR, Wiegand SJ and
Streilein JW: VEGF-A stimulates lymphangiogenesis and
hemangiogenesis in inflammatory neovascularization via macrophage
recruitment. J Clin Invest. 113:1040–1050. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Halin C, Tobler NE, Vigl B, Brown LF and
Detmar M: VEGF-A produced by chronically inflamed tissue induces
lymphangiogenesis in draining lymph nodes. Blood. 110:3158–3167.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kunstfeld R, Hirakawa S, Hong YK, Schacht
V, Lange-Asschenfeldt B, Velasco P, Lin C, Fiebiger E, Wei X, Wu Y,
Hicklin D, Bohlen P and Detmar M: Induction of cutaneous
delayed-type hypersensitivity reactions in VEGF-A transgenic mice
results in chronic skin inflammation associated with persistent
lymphatic hyperplasia. Blood. 104:1048–1057. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hong YK, Lange-Asschenfeldt B, Velasco P,
Hirakawa S, Kunstfeld R, Brown LF, Bohlen P, Senger DR and Detmar
M: VEGF-A promotes tissue repair-associated lymphatic vessel
formation via VEGFR-2 and the α1β1 and α2β1 integrins. FASEB J.
18:1111–1113. 2004.PubMed/NCBI
|