Introduction
Endometriosis is an estrogen-dependent gynecological
disorder that has many causes and is associated with many genes. It
is estimated that endometriosis may affect between 5 and 10% of
women who are of reproductive age (1). While endometriosis is a benign lesion,
it shares several characteristics with invasive cancer. Similar to
cancer, its behavior is characterized by proliferation, invasion,
metastasis and a high recurrence rate. Since Sampson first
theorized that endometriotic lesions could undergo malignant
transformation in 1925 (2),
histological and epidemiological studies have suggested that
ovarian endometriosis can transform into ovarian carcinoma. The
average rate of endometriotic malignant transformation is ~0.7%
(3). With an increase in the
morbidity associated with endometriosis, the malignant
transformation rate also has increased. Stern et al
(4) showed a 5% incidence of
ovarian malignancy associated with ovarian endometriosis. According
to Yoshikawa et al (5),
types of malignancies in ovarian endometriosis include clear cell
(39.2%), endometrioid (21.2%), serous (3.3%) and mucinous (3.0%).
Currently, endometrioid and clear cell carcinomas are the
predominant histological subtypes of endometriosis-associated
ovarian carcinoma (EAOC). Ovarian cancer is a malignant disease
with high global morbidity and mortality rates. Research on the
pathogenesis and candidate genes involved in EAOC development is
important for early detection, prevention and identification of new
therapeutic targets.
Research regarding the mechanisms involved in EAOC
has been limited by the low number of cases, difficulty obtaining
pathological materials and other factors. In recent years,
information concerning the pathogenesis of ovarian cancer has
significantly changed, including introduction of a dualistic model
in which the majority of what appear to be primary ovarian cancers,
namely serous, endometrioid and clear cell carcinomas, are thought
to be derived from the fallopian tube and endometrium rather than
directly from the ovary (6).
Understanding the associations between endometrioid and clear cell
carcinomas and malignant transformation of ovarian endometriosis
will require new approaches. At present, the rate of endometriotic
malignant transformation may be under-estimated, and evidence-based
medicine and epidemiology are necessary for revalidation.
Fundamental research regarding malignant transformation of
endometriosis will also require further attention.
Tumor-suppressor genes can be inactivated through a
variety of mechanisms. In addition to genetic mutations or
deletions, DNA methylation is widely considered one of the
mechanisms that leads to decreased genetic expression. Research has
shown that DNA methylation may represent an early, fundamental step
in the pathway through which normal tissues undergo neoplastic
transformation (7). Assessment of
methylation profiles within neoplastic tissues may provide key
information in enhancing diagnosis, predicting clinical behavior
and designing specific treatment plans for individual patients.
Martini et al (8)
demonstrated that hMLH1 promoter hypermethylation and resultant
gene silencing may be involved in the malignant transformation of
endometriosis. We previously screened candidate genes related to
malignant transformation of ovarian endometriosis for aberrant
methylation by evaluating CpG island methylation using enrichment
methods combined with representational difference analysis
(MCA-RDA). RUNX3 was one of the 9 candidate genes and is a
tumor-suppressor gene located on 1p36 (9). The largest, most recent genetic
studies of endometriosis and EAOC have identified 1p36 as a region
of paramount significance (10).
The runt domain family, consisting of RUNX1, RUNX2 and RUNX3, are
master regulators of gene expression in cell proliferation and
differentiation. All three RUNX family members play important roles
in normal developmental processes and carcinogenesis (11). Among the three RUNX family members,
RUNX3 was originally cloned as AML2 and was first reported to be
correlated with the genesis and progression of human gastric cancer
as a tumor suppressor (12). The
RUNX3 protein can mediate the growth suppressive effects of TGF-β
by associating with SMAD, a downstream protein in the signaling
pathway (13). RUNX3 has been shown
to be a target for epigenetic gene silencing via aberrant
hypermethylation of its CpG island promoter region in a variety of
estrogen-dependent tumors, such as breast cancer (14), endometrial cancer (15), ovarian granulosa cell tumors (GCTs)
(16) and testicular yolk sac
tumors in infants (17). Cheng
et al (18) found that RUNX3
promoter hypermethylation was induced by estrogen in
mammosphere-derived breast cancer cells. This estrogen-dependent
epigenetic silencing of RUNX3 is likely mediated by estrogen
receptor α (ERα) signaling. Estrogen, a major trigger for malignant
transformation of endometriosis, appears to play a role in this
process. Prolonged exposure to estrogen is known to increase the
risk of developing malignant transformation of endometriosis
(19). We hypothesized that RUNX3
inactivation by promoter hypermethylation is involved in malignant
transformation of endometriosis, which has not been previously
reported.
In the present study, we investigated the
association between RUNX3 inactivation primarily due to promoter
methylation and malignant transformation of ovarian EM. We also
investigated whether RUNX3 detection in the eutopic endometrium
could be used as a diagnostic marker in patients with endometriosis
who may be at an increased risk of developing EAOC. In addition, we
evaluated the relationship between RUNX3 promoter methylation and
ERα protein expression to define the role of estrogen in initiating
RUNX3 promoter methylation.
Materials and methods
Patients and materials
A total of 74 formalin-fixed and paraffin-embedded
(FFPE) samples were obtained from the Department of Pathology at
Shengjing Hospital, China Medical University from January 2005 to
December 2013. Malignant ovarian endometriotic cysts and the
corresponding eutopic endometrium from 30 patients who had
undergone total hysterectomy or salpingo-oophorectomy and surgical
staging of ovarian cancer (13 endometrioid and 17 clear cell
cancers) due to EAOC were collected as the EAOC group.
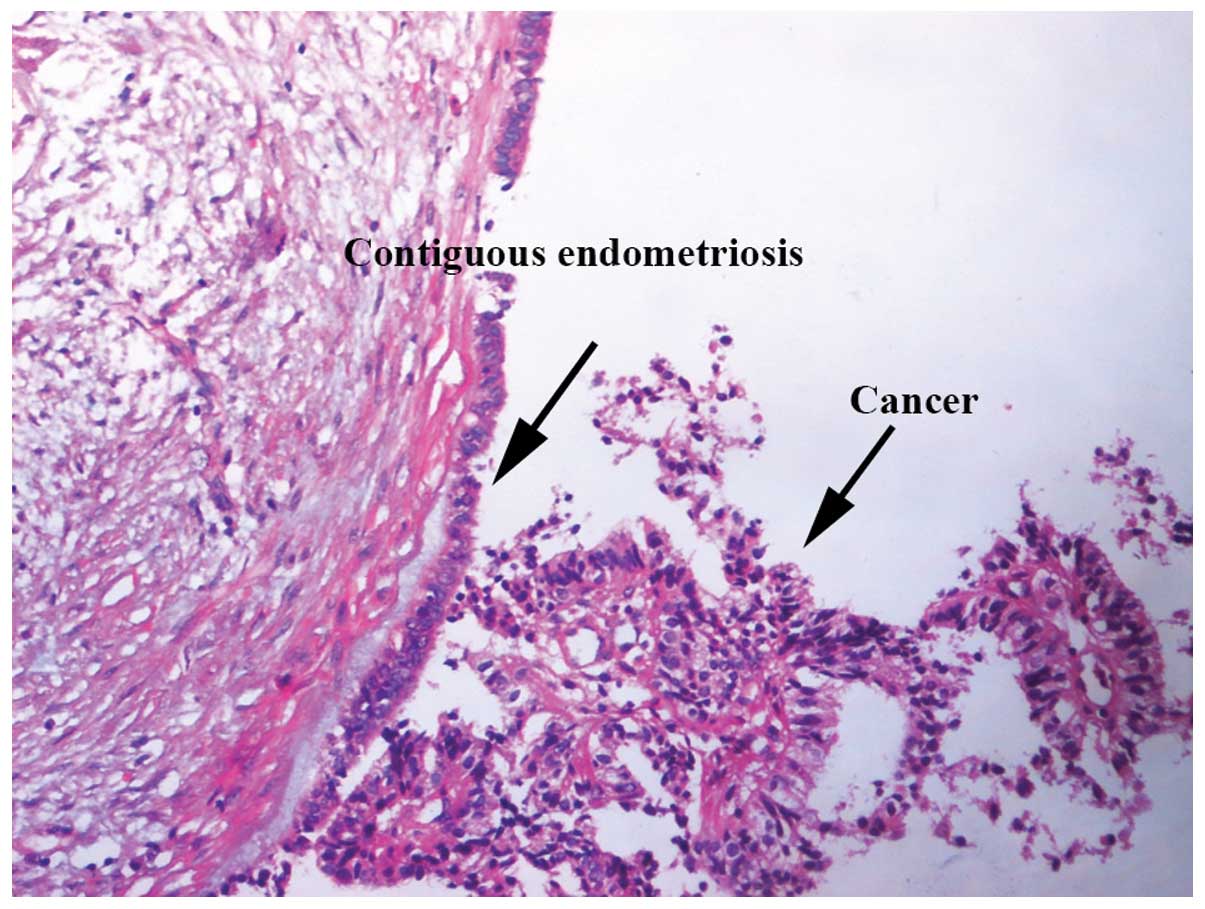
Histopathological examination revealed the coexistence and
continuity of endometriosis and cancer in patients with EAOC,
fulfilling the criteria proposed by Sampson (2) and Scott (20) (Fig.
1). Benign ovarian endometriotic cysts and the corresponding
eutopic endometrium from 22 patients who had undergone total
hysterectomy due to ovarian EM were collected as the EM group. The
normal eutopic endometrium from 22 patients diagnosed with benign
uterine conditions without endometriosis and estrogen-dependent
diseases who had undergone total hysterectomy were collected as the
CE group. The average patient age in the EAOC, EM and CE group was
51.17±1.50, 47.68±0.64 and 47.36±0.40 years, respectively, and
there was no statistical difference in age among the three groups
(P>0.05). None of the patients had received gonadotropin
releasing hormone (GnRH) analogs or other hormonal medications or
antibiotics during the 6 months prior to surgery. Histological
diagnoses for each patient were reviewed by a gynecological
pathologist following the criteria of the World Health Organization
classification.
Microdissection
All paraffin-embedded tissues were sliced at a 5-μm
thickness for hematoxylin and eosin (H&E) staining and
consecutively sliced into 4–6 sections at a 10-μm thickness
unstained for microscopic cutting. Microscopic cutting steps were
as follows. One slide of each specimen was randomly selected for
H&E staining as a template; the other slides were placed on the
H&E template and the target area was scraped using a disposable
sterile blade under a dissection microscope. Scraped tissues
included 30 malignant ovarian endometriotic cysts and 30
corresponding eutopic endometrium tissues from the EAOC group, 19
benign ovarian endometriotic cysts and 22 corresponding eutopic
endometrium tissues from the EM group and 22 normal eutopic
endometrium tissues from the CE group. Of the 22 patients who were
histologically confirmed as having ovarian EM, 3 patient samples
did not have sufficient tissue for microscopic cutting.
DNA extraction and bisulfite
modification
Total genomic DNA was extracted from 10-μm paraffin
tissue sections using the TIANamp Micro DNA kit (Tiangen Biotech
Co., Ltd., Beijing, China) according to the manufacturer’s
instructions, and 0.5 μg from each sample was modified via sodium
bisulfite treatment using the EZ DNA Methylation-Gold kit (Zymo
Research Corp., Orange, CA, USA) following the manufacturer’s
protocol.
Methylation-specific PCR (MSP) and
sequencing (BSP)
Methylation status in the RUNX3 promoter region was
determined by MSP. Modified genomic DNA was amplified using two
different sets of primers specific for unmethylated and methylated
RUNX3 promoters. Primers for the methylated reaction were as
follows (3): sense, 5′-ATA ATA GCG
GTC GTT AGG GCG TCG-3′ and antisense, 5′-GCT TCT ACT TTC CCG CTT
CTC GCG-3′. Primers for the unmethylated reaction were as follows:
sense, 5′-ATA ATA GTG GTT GTT AGG GTG TTG-3′ and antisense, 5′-ACT
TCT ACT TTC CCA CTT CTC ACA-3′ and the product size was 115 bp. PCR
was performed under the following conditions: initial denaturation
at 94°C for 5 min followed by 35 amplification cycles (denaturation
at 94°C for 30 sec, annealing at 55°C for 30 sec and extension at
72°C for 30 sec) and a final extension at 72°C for 7 min in a PCR
Thermal Cycler Dice (Takara Bio Inc., Otsu, Japan) using the Takara
EpiTap™ HS kit (Takara Biotechnology Co., Ltd., Dalian, China).
Normal lymphocyte DNA methylated in vitro with SssI
methylase (New England Biolabs, Beverly, MA, USA) was used as a
positive control (M+) and water was used as a blank control.
Products were subjected to electrophoresis on a 2% agarose gel. For
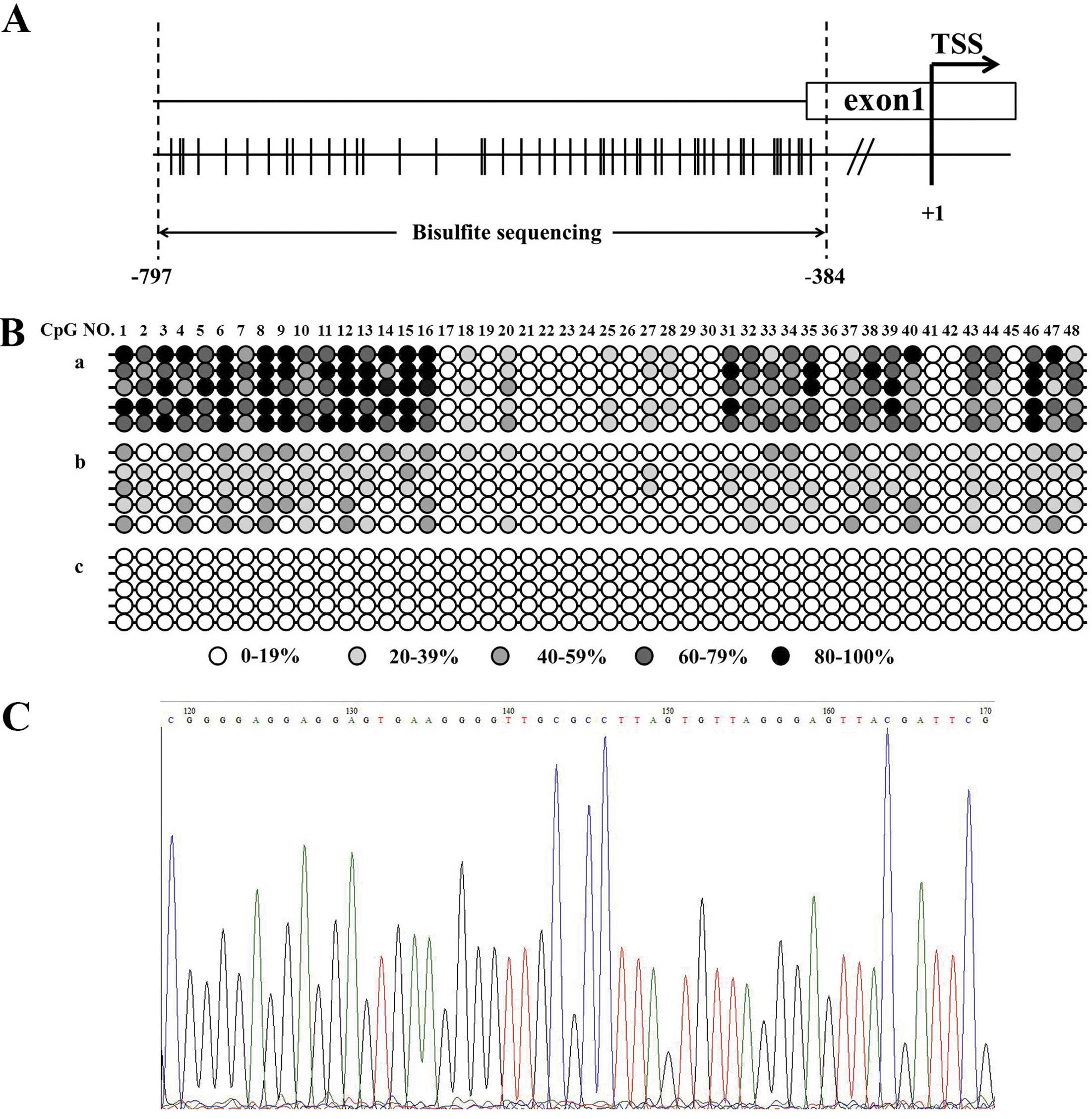
sequencing, a 414-nt region of the RUNX3 promoter between −797 and
−384 relative to the translation start site (TSS) was amplified
with the Takara EpiTap™ HS kit using primers: forward,
AGAAGTAGTAAGAGTTGGGGAAGTT and reverse, ACAAATCCTCCAAAATCAAATAAC.
Amplicons were gel purified using the TIANgel Midi Purification kit
(Tiangen Biotech Co., Ltd.) and subcloned into the PCR®
2.1 vector (Invitrogen). Five clones were randomly selected,
sequenced and analyzed using a BISMA analyzer.
Immunohistochemistry
All FFPE tissues were sectioned at a thickness of
5-μm for immunohistochemical staining using the standard
streptavidin-biotin method. Slides were dewaxed and rehydrated
following standard procedures. Antigen retrieval was performed to
enhance immunohistochemistry by microwaving the slides in citrate
buffer (pH 6.0) for 10 min. Endogenous peroxidase activity was
blocked for 10 min in 0.3% H2O2. Sections
were subsequently washed and blocked with normal goat serum for 30
min at room temperature and incubated with mouse monoclonal
antibody to RUNX3 (1:400; Enzo Life Sciences AG, Lausen,
Switzerland) and rabbit polyclonal antibody to ERα (1:100; Abcam,
Cambridge, UK) overnight at 4°C. Slides were washed in
phosphate-buffered saline and incubated with biotinylated goat
anti-mouse or goat anti-rabbit secondary antibodies for 30 min at
room temperature. Antigen-antibody complexes were detected with
avidin-biotin peroxidase for 30 min at room temperature. Slides
were stained with diaminobenzidine until the desired staining
intensity was developed and observed by light microscopy. Antibody
effectors of RUNX3 and ERα are brown granules expressed in the
nucleus and/or cell cytoplasm. RUNX3 and ERα expression was
evaluated by combining the estimated percentage of positive cells
with the estimated staining intensity. The frequency of positively
stained cells per 100 cells was scored as 0 (no positive cells), 1
(positive cells ≤10%), 2 (positive cells >10% and ≤50%), 3
(positive cells >50% and ≤75%) or 4 (positive >75%). Staining
intensity was scored as follows: 0, negative (no color); 1, weak
brown; 2, moderate brown and 3, strong brown. Raw data were
converted to an immunohistochemical score by multiplying the
quantity and staining intensity scores and graded from − to +++ (−,
0–4; +, 4–5; ++, 6–8; +++, 9–12). IHC scoring was performed
independently by two pathologists, and their consensus was
considered the final score in each case. The immunoreactivity
results − and + were considered negative, while ++ and +++ results
were considered positive.
Statistical analysis
Statistical analysis was performed with SPSS
software version 13.0 (SPSS Inc., Chicago, IL, USA). Results were
analyzed using the Chi-squared test and Fisher’s exact probability
test when appropriate. A two-sided value of P<0.05 was
considered statistically significant.
Results
Increased hypermethylation of the RUNX3
promoter
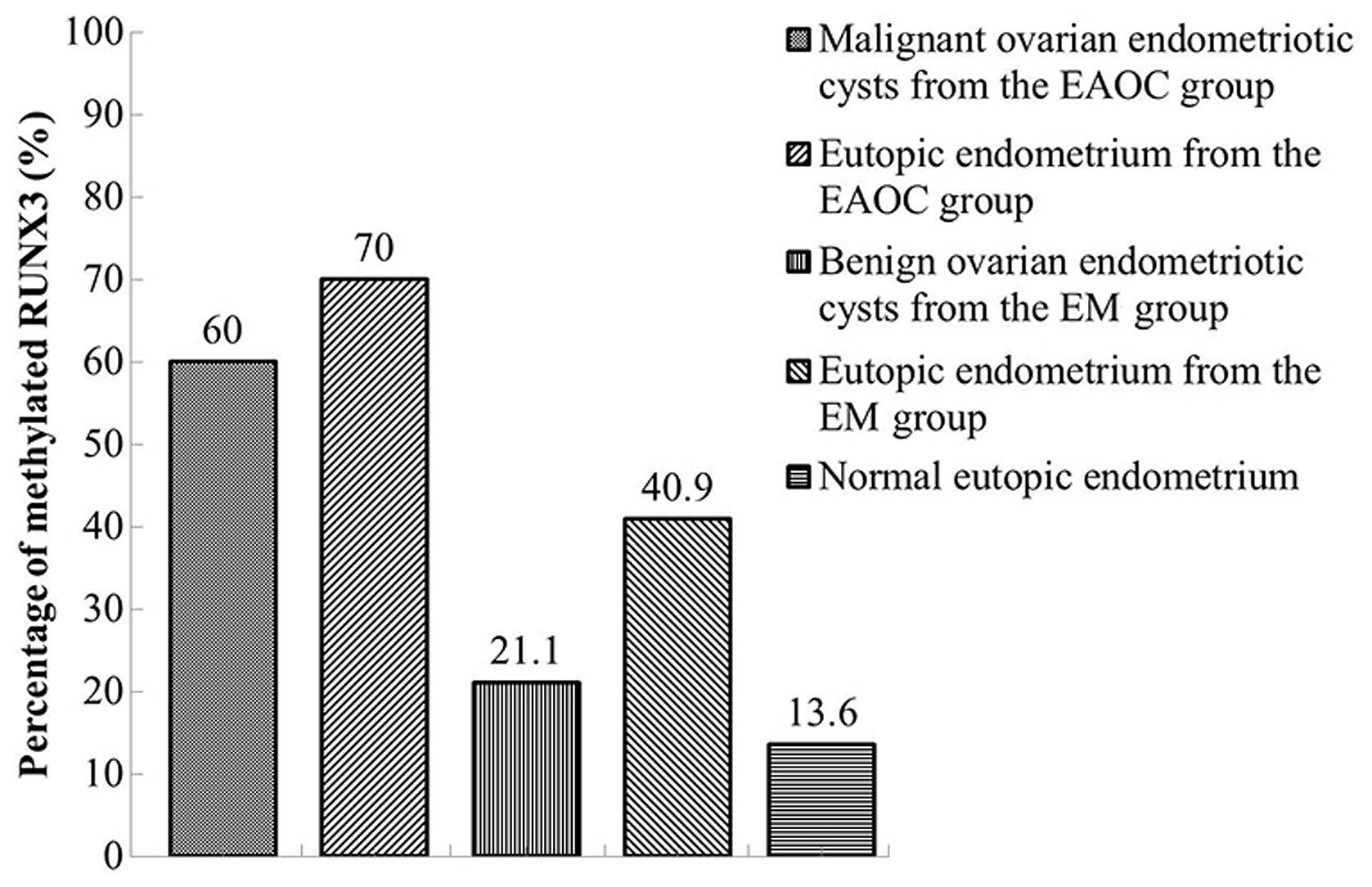
RUNX3 methylation was detected in 18 of 30 (60.0%)
malignant ovarian endometriotic cysts in the EAOC group and 4 of 19
(21.1%) benign ovarian endometriotic cysts in the EM group. The
percentage of RUNX3 gene methylation in the EAOC group was
significantly higher than that in the EM group (Fig. 2, P<0.05). Methylation frequencies
in the corresponding eutopic endometrium from the EAOC, EM and CE
groups were 70.0% (21/30), 40.9% (9/22) and 13.6% (3/22),
respectively. Statistically significant differences in the
frequency of RUNX3 gene methylation were observed, with gradually
decreasing levels among the three groups (Fig. 2, P<0.05). However, this
significant difference was not found among the malignant ovarian
endometriotic cysts, benign ovarian endometriotic cysts and their
corresponding eutopic endometrium (P>0.05). Representative
images from MSP analysis show methylated and unmethylated PCR
products (Fig. 3).
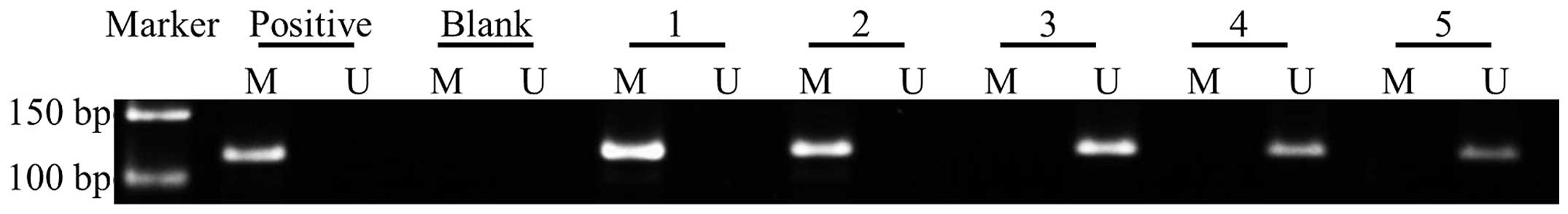 | Figure 3Representative image from MSP analysis
of the RUNX3 methylation status in the EAOC group, EM group and CE
group. M and U are the methylated and unmethylated primers,
respectively. Positive, methylation-positive control; Blank, blank
control. Lane 1, malignant ovarian endometriotic cyst tissues from
the EAOC group; lane 2, corresponding eutopic endometrium tissues
from the EAOC group; lane 3, benign ovarian endometriotic cyst
tissues from the EM group; lane 4, corresponding eutopic
endometrium tissues from the EM group; lane 5, normal eutopic
endometrium from the CE group. RUNX3, runt-related transcription
factor 3 gene; EAOC, endometriosis-associated ovarian carcinoma;
EM, endometriosis; CE, control endometrium. |
The methylation status of 48 CpGs across a 414-nt
region from −797 to −384 relative to the TSS was characterized by
bisulfite genomic sequencing, which is the gold standard for
mapping methylated CpG sites. We randomly selected 5 malignant
ovarian endometriotic cyst tissues from the EAOC group, 5 benign
ovarian endometriotic cyst tissues from the EM group and 5 normal
eutopic endometrium from the CE group. After subcloning the PCR
products generated from bisulfite-treated DNA using the
PCR® 2.1 vector, a selected clone was sequenced to
determine the precise methylation status of 48 CpG sites within the
RUNX3 gene. Malignant ovarian endometriotic cysts from the EAOC
group showed a dense methylation pattern in this region of the
RUNX3 gene compared to benign ovarian endometriotic cysts from the
EM group. No methylated CpG sites were detected in 5 samples from
the CE group (Fig. 4).
Relationship between RUNX3 methylation in
malignant ovarian endometriotic cysts from the EAOC group and
histological type and stage
As shown in Table I,
there was no significant correlation between the percentage of
methylation of the RUNX3 gene and histological type in the
malignant ovarian endometriotic cysts from the EAOC group
(P>0.05). However, the percentage of RUNX3 gene methylation was
significantly higher in the tissue samples from patients with
surgical stage IC EAOC than this percentage in the patients with
stage IA and IB disease (P<0.01).
 | Table IRelationship between RUNX3 methylation
in malignant ovarian endometriotic cysts from the EAOC group and
histological type and stage. |
Table I
Relationship between RUNX3 methylation
in malignant ovarian endometriotic cysts from the EAOC group and
histological type and stage.
| Characteristics | No. of cases | Methylated | Unmethylated | Methylated RUNX3
(%) | P-value |
|---|
| Histological
types |
| Endometrioid
carcinoma | 13 | 7 | 6 | 53.8 | 0.7106 |
| Clear cell
carcinoma | 17 | 11 | 6 | 64.7 | |
| Stage |
| IA | 12 | 5 | 7 | 41.7 | 0.6000a |
| IB | 5 | 1 | 4 | 20.0 | 0.0068b |
| IC | 13 | 12 | 1 | 92.3 | 0.0077c |
| ERα |
| Positive | 12 | 11 | 1 | 91.7 | 0.0068 |
| Negative | 18 | 7 | 11 | 38.9 | |
Association between RUNX3 methylation
status and protein expression
In the EAOC group, the percentage of positive RUNX3
protein expression was markedly higher in the malignant ovarian
endometriotic cysts (36.7%) than that in the corresponding eutopic
endometrium (10.0%) (P<0.05). In the EM group, the percentage of
positive RUNX3 protein expression was markedly higher in the benign
ovarian endometriotic cysts (84.2%) than that in the corresponding
eutopic endometrium (59.1%) (P<0.05). Compared to the benign
ovarian endometriotic cysts in the EM group (84.2%), the percentage
of positive RUNX3 protein expression in the malignant ovarian
endometriotic cysts from the EAOC group (36.7%) was significantly
downregulated (P<0.05). In the corresponding eutopic endometrium
from the EAOC, EM and CE groups, the incidence of positive RUNX3
protein expression was 10.0, 59.1 and 100.0%, respectively
(P<0.05) (Fig. 5, Table II).
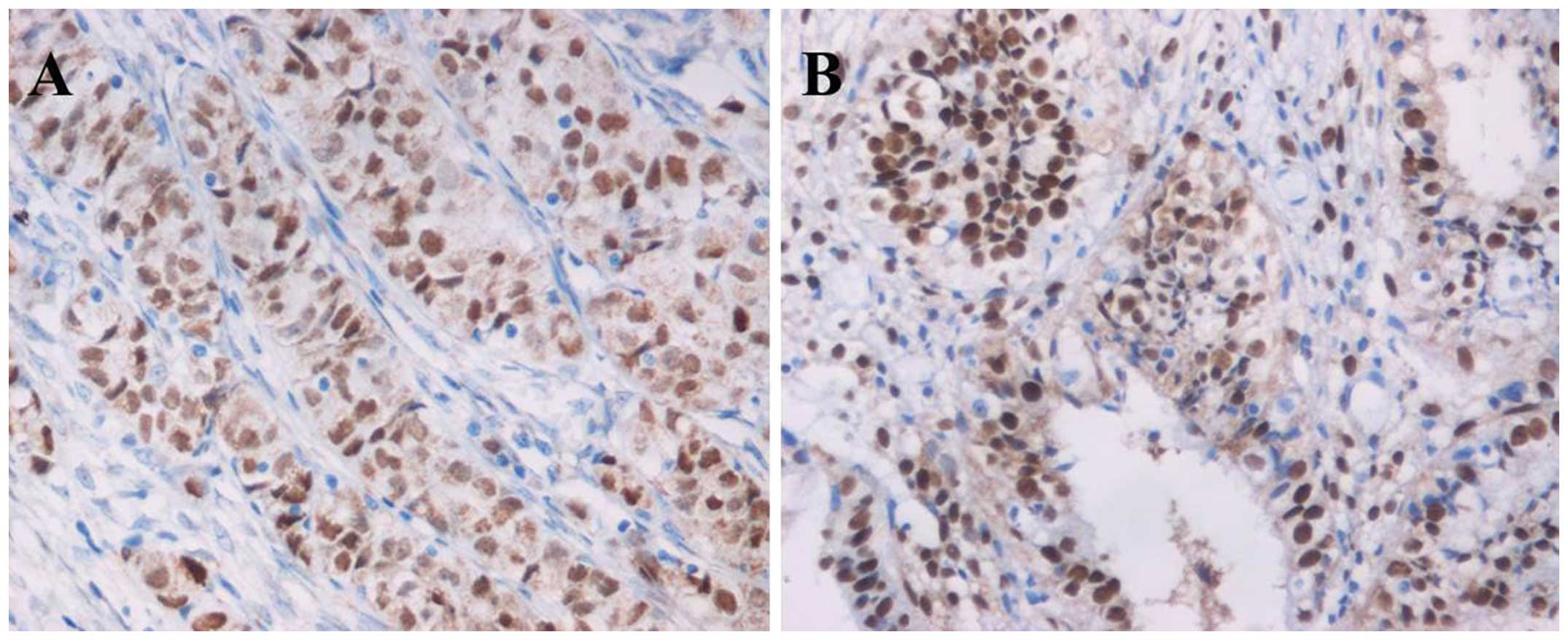
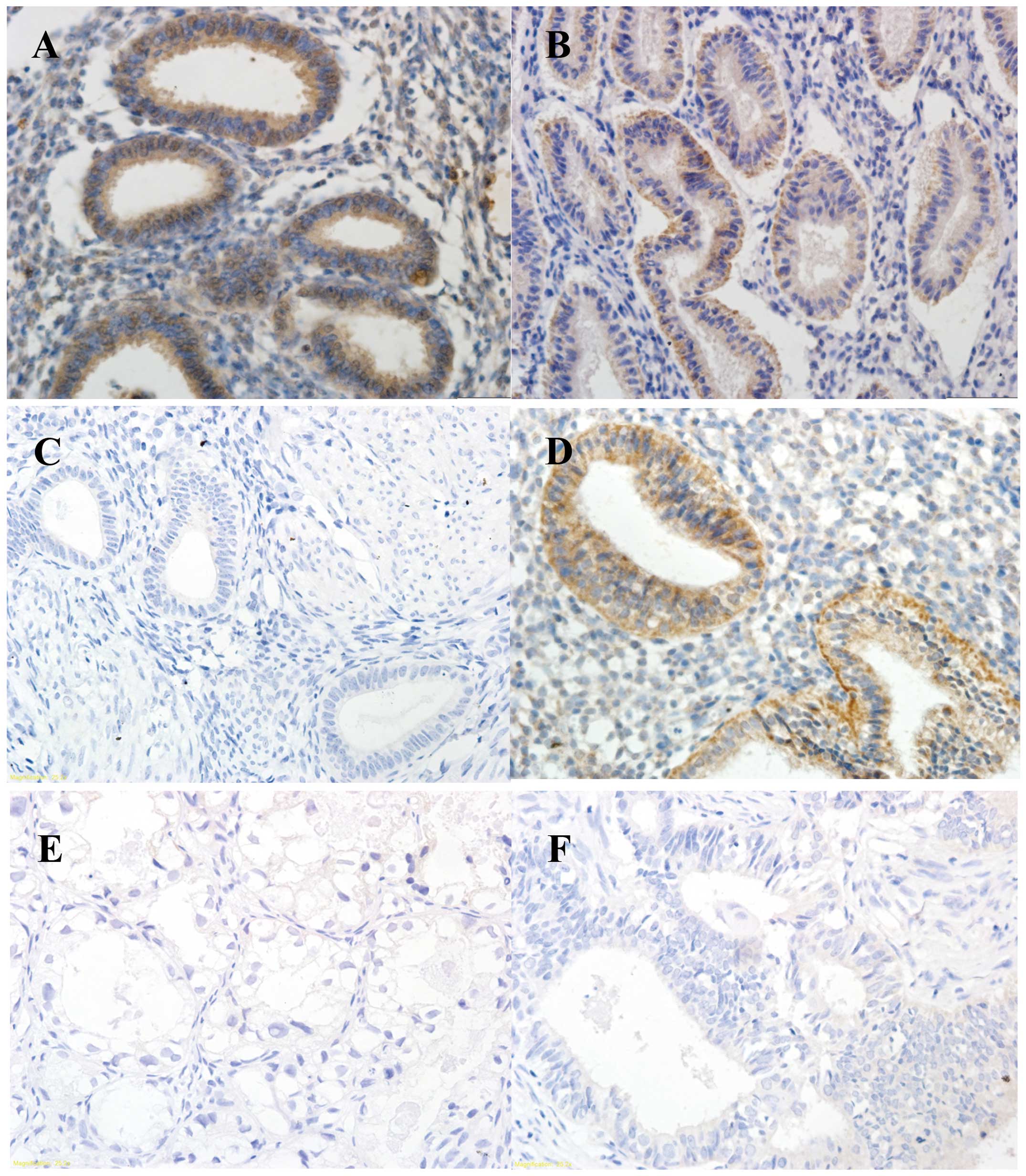 | Figure 5Immunohistochemical evaluation of
RUNX3 expression in the EAOC, EM and CE groups. (A) Normal eutopic
endometrium from the CE group, (B) corresponding eutopic
endometrium tissues from the EM group, (C) corresponding eutopic
endometrium tissues from the EAOC group, (D) benign ovarian
endometriotic cyst tissues from the EM group, (E) clear cell
carcinoma and (F) endometrioid carcinoma (original magnification,
×400). RUNX3, runt-related transcription factor 3 gene; EAOC,
endometriosis-associated ovarian carcinoma; EM, endometriosis; CE,
control endometrium. |
| Table IIAssociation between RUNX3 methylation
status and protein expression. |
Table II
Association between RUNX3 methylation
status and protein expression.
| | RUNX3
methylated | RUNX3
unmethylated |
|---|
| |
|
|
|---|
| No. of cases | Total | Positive RUNX3
protein expression (%) | Total | Positive RUNX3
protein expression (%) |
|---|
| EAOC group |
| Malignant ovarian
endometriotic cysts | 30 | 18 | 0 (0.0) | 12 | 11 (91.7) |
| Eutopic
endometrium | 30 | 21 | 1 (4.8) | 9 | 2 (22.2) |
| EM group |
| Benign ovarian
endometriotic cysts | 19 | 4 | 1 (25.0) | 15 | 15 (100.0) |
| Eutopic
endometrium | 22 | 9 | 3 (33.3) | 13 | 10 (76.9) |
| CE group |
| Normal eutopic
endometrium | 22 | 3 | 3 (100.0) | 19 | 19 (100.0) |
Overall, 47 of the 55 methylated samples were
negative for RUNX3 protein expression and only 11 of the 68 RUNX3
unmethylated samples were negative for RUNX3 protein expression. An
inverse correlation was noted between positive RUNX3 protein
expression and methylation (rs=−0.69, P<0.01)
(Table II).
Correlation between RUNX3 methylation
status and ERα protein expression in the malignant ovarian
endometriotic cysts from the EAOC group
In the malignant ovarian endometriotic cysts from
the EAOC group, the methylation frequency of the RUNX3 gene was
higher in the group with positive ERα protein expression compared
to the group with negative ERα protein expression (P<0.05).
Eleven of the 18 RUNX3 methylated samples had positive ERα protein
expression, while only 1 of 12 RUNX3 unmethylated samples were
positive. A positive correlation was shown between RUNX3
methylation and ERα protein expression (rs=0.53,
P<0.01) (Fig. 6, Table I).
Discussion
Malignant transformation of endometriosis is a long,
complicated process involving multiple genes and steps. Epigenetic
inactivation of cancer-associated genes has been observed
frequently in most human cancers and has been shown to be closely
associated with DNA hypermethylation. Silencing the promoter region
in CpGs of regulated genes, such as RUNX3, via hypermethylation is
recognized as one of three mechanisms that contributes to loss of
gene function in Knudson’s two-hit tumor progression hypothesis
(21). More importantly, silencing
due to hypermethylation is an epigenomic change in premalignant
lesions as part of the multistep carcinogenetic process in some
malignancies (7). DNA
methylation is a heritable, enzyme-induced modification to the DNA
structure without altering the specific sequence of base pairs
responsible for encoding the genome and the abnormal methylation
patterns can be reversed (22).
Consequently, systemic study of gene methylation is extremely
important in the early diagnosis and treatment of diseases.
Senthong et al (23)
demonstrated that LINE-1 hypomethylation occurs early in EAOC
development and indicated that epigenomic alterations may precede
development of ovarian cancer phenotypes. Therefore, epigenetic
modifications, particularly DNA methylation, provide exciting new
avenues in the study of the malignant transformation of
endometriosis.
Epigenetic inactivation of RUNX3 has been detected
in numerous cancers, particularly malignancies that involve
estrogen metabolism. Herein, we report the RUNX3 methylation status
in ovarian EM and EAOC for the first time. In the present study,
the frequency of RUNX3 methylation and negative RUNX3 protein
expression in malignant ovarian endometriotic cysts from the EAOC
group was significantly higher than that in the benign ovarian
endometriotic cysts from the EM group and normal eutopic
endometrium of the CE group. RUNX3 expression was inversely
correlated with aberrant hypermethylation of CpG islands in the
RUNX3 promoter region. Inactivation of RUNX3 expression primarily
due to frequent promoter hypermethylation may play an important
role in transformation of the endometrium to endometriosis and
EAOC. In addition, a relatively high percentage of RUNX3
methylation was detected in benign ovarian endometriotic cysts from
the EM group and normal eutopic endometrium from the CE group,
suggesting that epigenomic alterations in precancerous lesions can
result in individuals becoming carriers of abnormal methylation
patterns. Although the tissue morphology may appear normal, cells
have changed at a molecular level, which has been widely observed
in other tumors (24). Therefore,
RUNX3 methylation is an early event in the pathogenesis of EAOC and
may be a target for early EAOC detection.
Within the same tumor, methylation frequencies at
different CpG sites are not exactly the same (25). There are examples in which
methylation of individual CpG sites in promoter regions can
determine the transcriptional status of a gene by blocking access
of certain transcription factors that are sensitive to CpG
methylation at target binding sites. Alternatively, methylation of
sites in these regions may also silence genes by helping to recruit
chromatin protein complexes that repress transcription (26). In a previous study, we screened
different fragments of RUNX3 methylation using MCA-RDA at sites
located from −810 to −582 relative to the TSS. In the present
study, we tested the methylation status at a total of 48 CpG sites
across a 414-bp region from −797 to −384 relative to RUNX3 TSS.
Malignant ovarian endometriotic cysts from the EAOC group showed a
dense methylation pattern in this region compared to benign ovarian
endometriotic cysts from the EM group. No methylation CpG sites
were detected in 5 samples from the CE group. These results further
confirm that methylation of sites in this region play a significant
role in the malignant transformation of ovarian EM.
Hypermethylation of promoter CpG islands in tumor suppressors is
known to be a frequent, early reversible event in carcinogenesis
and can contribute to disease progression (27). Although the frequency of RUNX3
methylation increased with pathological stage, there was no
significant correlation with histological type, demonstrating that
methylation may play a role in both the occurrence and development
of EAOC regardless of histology. These data strongly imply early
epigenetic gene inactivation in EAOC and may assist in identifying
novel epigenetic biomarkers associated with EAOC risk.
Studies have shown that as many as 50% of women with
ovarian endometrioid carcinomas also have a simultaneous
endometrial adenocarcinoma, and ovarian cancer has been reported to
occur in 2–8.5% of patients with endometrial cancer. Almost 90% of
synchronous tumors identified in the ovary and endometrium were of
the endometrioid cell type. There is also a high incidence of
ovarian EM cases among synchronous adenocarcinomas originating in
the ovary and endometrium (28,29).
Based on a series of morphologic and molecular genetic studies,
Kurman et al (30) proposed
a dualistic model that categorizes various types of ovarian cancer
into two groups, type I and type II. The cellular origins of
ovarian cancer and the mechanisms by which cancer develops have
been long debated. The traditional view of ovarian carcinogenesis
is that various tumors are all derived from the ovarian surface
epithelium (mesothelium) and that subsequent metaplastic changes
lead to development of different cell types (e.g., serous,
endometrioid, clear cell, mucinous and transitional cell). However,
compelling recent evidence suggests that the vast majority of what
appear to be primary ovarian cancers, including serous,
endometrioid and clear cell carcinomas, are derived from the
fallopian tubes and endometrium and not directly from the ovary.
Endometrioid and clear cell carcinomas may develop from malignant
transformation of ovarian endometriotic cysts, with secondary
involvement from the ovary (31).
Studies have reported that malignant transformation of ovarian
endometriosis is a continual process. Similar to the eutopic
endometrium, ectopic endometrial cells can also transform into
atypical hyperplasia and cancer (32). In addition, other authors have
demonstrated a higher prevalence of endometrial pathology (e.g.,
endometrial adenocarcinoma, hyperplasia and polyps) when ovarian
tumors were considered to originate from endometriosis (33). This indicates a shared molecular
pathogenesis in the process of malignant transformation of the
eutopic and ectopic endometrium. Although the precise mechanism
leading to endometriosis is not clear, it is generally accepted
that the eutopic endometrium in women with endometriosis has
intrinsic molecular abnormalities (1). Moreover, Rosenblatt et al
observed that the strongest apparent protective effects of tubal
ligation occurred for clear cell and endometrioid tumors, but there
were no differences in the risk for borderline and malignant tumors
(34). In a previous study, we
detected a higher methylation frequency of hMLH1 in the eutopic
endometrium from the EAOC group compared to the EM and CE groups,
suggesting that abnormal hMLH1 methylation in the eutopic
endometrium from EAOC and malignant transformation of the ectopic
endometrium are interrelated (35).
Consequently, intrinsic molecular abnormalities in the eutopic
endometrium may be the source of malignant transformation of the
ectopic endometrium. In the present study, we evaluated the RUNX3
methylation status and protein expression in the eutopic
endometrium from three groups. The frequency of epigenetic RUNX3
inactivation in the eutopic endometrium from the EAOC group was
higher than in the EM and CE groups. These results verify the
hypothesis that intrinsic molecular abnormalities in the eutopic
endometrium may be the source of malignant transformation of the
ectopic endometrium and that the eutopic and ectopic endometrium
may be synchronously affected by RUNX3 hypermethylation. Epigenomic
RUNX3 alterations in the eutopic endometrium promoted malignant
transformation of the ectopic endometrium. Based on the above
conclusion, RUNX3 methylation status and protein expression in the
eutopic endometrium could provide early diagnosis of malignant
transformation of ovarian EM. Furthermore, the eutopic endometrium
is readily available and gene alteration in the eutopic endometrium
can be easily detected, which solves difficulties of basic
experimentation due to a shortage of malignantly transformed
ovarian EM samples.
EM is an estrogen-dependent disease and
hyperestrogenism has been associated with malignant transformation
of endometriotic cysts. The microenvironment provided by
endometriosis facilitates accumulation of excess estrogen. There
have been several recent reports that estrogen replacement therapy
could lead to malignant transformation of endometrial ectopic foci.
Recent studies imply that estrogen imprinting can be mediated by
epigenetic mechanisms via ERα. Suzuki et al (36) found that the frequency of RUNX3
promoter methylation was higher in ER-positive samples than
ER-negative samples. Subramaniam et al (14) also reported that RUNX3 promoter
hypermethylation was significantly correlated with positive ER
expression in invasive carcinomas. Our data showed that 91.67%
(11/12) of ERα-positive samples had RUNX3 promoter
hypermethylation, while only 38.9% (7/18) of ERα-negative samples
had detectable hypermethylation of the RUNX3 promoter in malignant
ovarian endometriotic cysts from the EAOC group. There was a
positive correlation between RUNX3 methylation and ERα protein
expression. We postulate that activation of ER signaling by high
local concentrations of estrogen and inactivation of RUNX3 protein
expression due to promoter methylation are involved in the process
of malignant transformation of endometriotic cysts. However, it
remains to be determined how estrogen receptor signaling initiates
methylation of the RUNX3 promoter. Correlation between RUNX3
methylation and estrogen is a main future research direction in
malignant transformation of ovarian EM.
In conclusion, we provide evidence that RUNX3
inactivation by promoter hypermethylation constitutes an early
event in the malignant transformation of ovarian EM and is closely
related to estrogen metabolism. Our results also suggest that
epigenetic inactivation of RUNX3 plays a vital role in the
occurrence and development of malignant transformation of ovarian
EM. Corresponding changes to the eutopic endometrium indicate that
detection of RUNX3 protein expression and methylation status in the
eutopic endometrium may be valuable tools for identifying patients
with ovarian EM who are at an elevated risk of malignant
transformation and could lead to preventative strategies and novel
treatment approaches for women who have been diagnosed with
EAOC.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation of China (nos. 81070467 and
81270675) and the Plan of Free Researchers in Shengjing
Hospital.
References
|
1
|
Bulun SE: Endometriosis. N Engl J Med.
360:268–279. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sampson JA: Endometrial carcinoma of the
ovary arising in endometrial tissue in that organ. Arch Surg.
10:1–72. 1925. View Article : Google Scholar
|
|
3
|
Kobayashi H, Sumimoto K, Moniwa N, et al:
Risk of developing ovarian cancer among women with ovarian
endometrioma: a cohort study in Shizuoka, Japan. Int J Gynecol
Cancer. 17:37–43. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Stern RC, Dash R, Bentley RC, Snyder MJ,
Haney AF and Robboy SJ: Malignancy in endometriosis: frequency and
comparison of ovarian and extraovarian types. Int J Gynecol Pathol.
20:133–139. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yoshikawa H, Jimbo H, Okada S, et al:
Prevalence of endometriosis in ovarian cancer. Gynecol Obstet
Invest. 50(Suppl 1): 11–17. 2000. View Article : Google Scholar
|
|
6
|
Shih Ie M and Kurman RJ: Ovarian
tumorigenesis: a proposed model based on morphological and
molecular genetic analysis. Am J Pathol. 164:1511–1518.
2004.PubMed/NCBI
|
|
7
|
Park SY, Kwon HJ, Lee HE, et al: Promoter
CpG island hypermethylation during breast cancer progression.
Virchows Arch. 458:73–84. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Martini M, Ciccarone M, Garganese G, et
al: Possible involvement of hMLH1, p16(INK4a) and
PTEN in the malignant transformation of endometriosis. Int J
Cancer. 102:398–406. 2002.
|
|
9
|
Levanon D, Negreanu V, Bernstein Y, Bar-Am
I, Avivi L and Groner Y: AML1, AML2, and AML3, the human members of
the runt domain gene-family: cDNA structure, expression, and
chromosomal localization. Genomics. 23:425–432. 1994.PubMed/NCBI
|
|
10
|
Uno S, Zembutsu H, Hirasawa A, et al: A
genome-wide association study identifies genetic variants in the
CDKN2BAS locus associated with endometriosis in Japanese.
Nat Genet. 42:707–710. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Habtemariam B, Anisimov VM and MacKerell
AD Jr: Cooperative binding of DNA and CBFβ to the Runt domain of
the CBFα studied via MD simulations. Nucleic Acids Res.
33:4212–4222. 2005.
|
|
12
|
Li QL, Ito K, Sakakura C, et al: Causal
relationship between the loss of RUNX3 expression and
gastric cancer. Cell. 109:113–124. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zaidi SK, Sullivan AJ, van Wijnen AJ,
Stein JL, Stein GS and Lian JB: Integration of Runx and Smad
regulatory signals at transcriptionally active subnuclear sites.
Proc Natl Acad Sci USA. 99:8048–8053. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Subramaniam MM, Chan JY, Soong R, et al:
RUNX3 inactivation by frequent promoter hypermethylation and
protein mislocalization constitute an early event in breast cancer
progression. Breast Cancer Res Treat. 113:113–121. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yoshizaki T, Enomoto T, Fujita M, et al:
Frequent inactivation of RUNX3 in endometrial carcinoma.
Gynecol Oncol. 110:439–444. 2008. View Article : Google Scholar
|
|
16
|
Dhillon VS, Shahid M and Husain SA:
CpG methylation of the FHIT, FANCF,
cyclin-D2, BRCA2 and RUNX3 genes in Granulosa
cell tumors (GCTs) of ovarian origin. Mol Cancer. 3:332004.
View Article : Google Scholar
|
|
17
|
Kato N, Tamura G, Fukase M, Shibuya H and
Motoyama T: Hypermethylation of the RUNX3 gene promoter in
testicular yolk sac tumor of infants. Am J Pathol. 163:387–391.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cheng AS, Culhane AC, Chan MW, et al:
Epithelial progeny of estrogen-exposed breast progenitor cells
display a cancer-like methylome. Cancer Res. 68:1786–1796. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Oxholm D, Knudsen UB, Kryger-Baggesen N
and Ravn P: Postmenopausal endometriosis. Acta Obstet Gynecol
Scand. 86:1158–1164. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Scott RB: Malignant changes in
endometriosis. Obstet Gynecol. 2:283–289. 1953.
|
|
21
|
Jones PA: The DNA methylation paradox.
Trends Genet. 15:34–37. 1999. View Article : Google Scholar
|
|
22
|
Kulis M and Esteller M: DNA methylation
and cancer. Adv Genet. 70:27–56. 2010. View Article : Google Scholar
|
|
23
|
Senthong A, Kitkumthorn N, Rattanatanyong
P, Khemapech N, Triratanachart S and Mutirangura A: Differences in
LINE-1 methylation between endometriotic ovarian cyst and
endometriosis-associated ovarian cancer. Int J Gynecol Cancer.
24:36–42. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zochbauer-Muller S, Fong KM, Maitra A, et
al: 5′ CpG island methylation of the FHIT gene is correlated
with loss of gene expression in lung and breast cancer. Cancer Res.
61:3581–3585. 2001.
|
|
25
|
Baylin SB and Herman JG: DNA
hypermethylation in tumorigenesis: epigenetics joins genetics.
Trends Genet. 16:168–174. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jones PA and Baylin SB: The fundamental
role of epigenetic events in cancer. Nat Rev Genet. 3:415–428.
2002.PubMed/NCBI
|
|
27
|
Wang H, Feng Y, Bao Z, et al: Epigenetic
silencing of KAZALD1 confers a better prognosis and is associated
with malignant transformation/progression in glioma. Oncol Rep.
30:2089–2096. 2013.PubMed/NCBI
|
|
28
|
Falkenberry SS, Steinhoff MM, Gordinier M,
Rappoport S, Gajewski W and Granai CO: Synchronous endometrioid
tumors of the ovary and endometrium. A clinicopathologic study of
22 cases. J Reprod Med. 41:713–718. 1996.PubMed/NCBI
|
|
29
|
Zaino R, Whitney C, Brady MF, DeGeest K,
Burger RA and Buller RE: Simultaneously detected endometrial and
ovarian carcinomas - a prospective clinicopathologic study of 74
cases: a Gynecologic Oncology Group study. Gynecol Oncol.
83:355–362. 2001. View Article : Google Scholar
|
|
30
|
Kurman RJ and Shih IeM: Pathogenesis of
ovarian cancer: lessons from morphology and molecular biology and
their clinical implications. Int J Gynecol Pathol. 27:151–160.
2008.PubMed/NCBI
|
|
31
|
Martin DC: Cancer and endometriosis: do we
need to be concerned? Semin Reprod Endocrinol. 15:319–324. 1997.
View Article : Google Scholar
|
|
32
|
Vigano P, Somigliana E, Chiodo I, Abbiati
A and Vercellini P: Molecular mechanisms and biological
plausibility underlying the malignant transformation of
endometriosis: a critical analysis. Hum Reprod Update. 12:77–89.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Van Gorp T, Amant F, Neven P, Vergote I
and Moerman P: Endometriosis and the development of malignant
tumours of the pelvis. A review of literature. Best Pract Res Clin
Obstet Gynaecol. 18:349–371. 2004.PubMed/NCBI
|
|
34
|
Rosenblatt KA and Thomas DB: Reduced risk
of ovarian cancer in women with a tubal ligation or hysterectomy.
The World Health Organization Collaborative Study of Neoplasia and
Steroid Contraceptives. Cancer Epidemiol Biomarkers Prev.
5:933–935. 1996.
|
|
35
|
Ren F, Wang D, Jiang Y and Ren F:
Epigenetic inactivation of hMLH1 in the malignant transformation of
ovarian endometriosis. Arch Gynecol Obstet. 285:215–221. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Suzuki M, Shigematsu H, Shames DS, et al:
DNA methylation-associated inactivation of TGFβ-related
genes DRM/Gremlin, RUNX3, and HPP1 in human
cancers. Br J Cancer. 93:1029–1037. 2005. View Article : Google Scholar
|