Introduction
Lung cancer is one of the most prevalent cancers and
is the leading cause of death-related worldwide (1,2). As
non-small cell lung cancers (NSCLCs) account for almost 85% of all
cases of human lung cancers, many therapeutic efforts have been
made to treat NSCLCs (3,4). During the last decade, receptor
tyrosine kinases (RTKs) such as epidermal growth factor receptor
(EGFR) have been regarded as appropriate targets for NSCLC
treatment (5–7). Actually, aberrant overexpression of
EGFR has been detected in ~50% of NSCLC patients and is known to be
correlated with tumor growth and poor prognosis of various types of
human cancers (8). Therefore,
EGFR-tyrosine kinase inhibitors (EGFR-TKIs) such as gefitinib or
erlotinib have been developed for patients with NSCLCs or other
human cancers (9,10). However, despite the beneficial
effect of these TKIs, EGFR-TKI therapies are now confronting the
problem of acquired resistance (11). Recent studies have identified the
two representative molecular mechanisms for this resistance: a
secondary T790M mutation on exon 20 of the EGFR gene and
amplification of the Met proto-oncogene or overexpression of
hepatocyte growth factor (HGF) (12–16).
Several EGFR-TKIs have been investigated to overcome this acquired
resistance including irreversible EGFR-TKIs and TKIs selective for
mutant EGFR (17). Moreover,
combined treatment with EGFR-TKIs and Met-TKIs has been proposed to
overcome resistance induced by c-Met amplification (18). Nonetheless, more effective and safer
therapeutic agents for overcoming resistance to EGFR-TKIs are still
needed.
Traditional natural compounds provide highly fertile
ground for drug development, and a number of studies have proven
that numerous purified natural products possess anticancer
activities (19). Notably,
Pulsatilla koreana is a traditional Korean herb which
belongs to the family Ranunculaceae. Its roots have been widely
used in traditional herbal medicine for the treatment of several
diseases, in particular, malaria and amoebic dysentery in Korea
(20). Notably, 17 types of
saponins have been isolated from Pulsatilla koreana, among
which, Pulsatilla saponin D (hereafter designated as SB365)
has been reported to exert anticancer effects in several types of
cancer cells (21–24). In particular, the present study
demonstrated that SB365 inhibits c-Met phosphorylation by docking
at its allosteric site, thereby suppressing the growth of gastric
cancers (22). Based on this, it
may be promising to assess the anticancer effects of SB365 in
gefitinib-resistant Met-amplified NSCLCs.
In the present study, we assessed the anticancer
efficacy of SB365 in Met-amplified HCC827GR cells. Here, we found
that SB365 exerts its ability to inhibit anchorage-(in) dependent
cell growth, migration and invasion by attenuating the Met-Akt
signaling pathway in gefitinib-resistant HCC827GR cells. Moreover,
our data also revealed that SB365 is sufficient for inducing
apoptosis in gefitinib-resistant HCC827GR cells.
Materials and methods
Materials
Antibodies for phospho-EGFR (Tyr1068), Met,
phospho-Met (Tyr1234/1235), Akt, phospho-Akt (Ser473), Erk1/2,
phospho-Erk1/2 (Thr202/Tyr204) and β-actin were purchased from Cell
Signaling Technology (Beverly, MA, USA). Antibodies specific for
EGFR and the secondary antibody were purchased from Santa Cruz
Biotechnology (Santa Cruz, CA, USA). Gefitinib was purchased from
LC Laboratories (Woburn, MA, USA). Halt™ Protease and phosphatase
inhibitor cocktail (100X), EDTA (100X) and the BCA protein assay
kit were purchased from Thermo Fisher Scientific (Rockford, IL,
USA). 3-(4,5-Dimethylthiazol-2yl)-2,5-diphenyltetrazolium bromide
(MTT) was purchased from Sigma-Aldrich (St. Louis, MO, USA). PVDF
membranes were purchased from Bio-Rad (Hercules, CA, USA).
SuperSignal West Dura Extended Duration Substrate was purchased
from Thermo Scientific (Waltham, MA, USA). A human phospho-RTK
array kit was purchased from R&D Systems (Minneapolis, MN,
USA).
Cell culture
The human NSCLC cell line HCC827 was cultured in
RPMI-1640 with L-glutamine supplemented with 10% fetal bovine serum
(FBS) and penicillin/streptomycin. The gefitinib-resistant cell
line HCC827GR was generated upon continuous exposure of parental
HCC827 cells to gefitinib for over 6 months. In detail, starting
with a gefitinib concentration of 10 nM, the exposure dose was
doubled every 2 weeks until a final concentration of 10 μM was
achieved. HCC827GR cells were cultured in RPMI-1640 with
L-glutamine supplemented with 10% FBS, penicillin/streptomycin and
1 μM gefitinib. All cells were maintained at 37°C with 5%
CO2 in a humidifier incubator.
Preparation of SB365
SB365 was isolated from the roots of P.
koreana, as previously described by Kim et al (25). Briefly, powered roots of P.
koreana (50 g) were extracted 3 times with 50% aqueous ethanol
(500 ml), and the resulting extracts were combined and concentrated
in vacuo to yield a light brown residue. The fraction was
chromatographed using a Sephadex LH-20 column (200 g, 60×4 cm) with
an 80:20 mixture of methanol and H2O, to give four
fractions: SPX1 (139 mg, 24.8%), SPX2 (344 mg, 61.4%), SPX3 (61 mg,
10.9%) and SPX4 (15.7 mg, 2.8%). After heating, the chromatogram
was sprayed with 10% H2SO4. The third
fraction, which exhibited the most potent activity, was again
chromatographed by solid phase HPLC [solid phase, RP-C18, 250 × 10
mm; mobile phase, MeOH-H2O (82:20); UV wavelength, 210
nm; flow rate, 1 ml/min] to yield 3 major fractions. Among them,
the third fraction (SPX3), which exhibited the most potent
activity, was purified by HPLC to yield saponin D, SB365. For more
precise analysis of purified SB365, we also used mass spectrometry
and NMR spectroscopy.
Cell proliferation assay
Cell proliferation was measured using MTT assay.
HCC827 and HCC827GR cells were seeded at 2.5×103
cells/well in 96-well plates. Following 24 h of incubation, the
cells were treated with the indicated concentrations of the
compounds, and subsequently incubated for 24, 48 and 72 h at 37°C.
After being incubated with the compounds, the medium was then
replaced with medium containing MTT (0.5 mg/ml MTT, 100 μl/well)
and incubated for 4 h. The medium was aspirated from the wells, and
100 μl of dimethyl sulfoxide (DMSO) was added into each well and
agitated for 180 sec. Absorbance at 565 nm was then read on a Tecan
Infinite F200 Pro plate reader (Promega, Madison, WI, USA), and
values are expressed as the percentage of absorbance from cells
incubated in DMSO alone.
Western blot analysis
HCC827 and HCC827GR cells were seeded in a 100-mm
culture dish at 2×106 cells and incubated for 24 h. The
cells were treated with the indicated concentrations of the
compounds or DMSO, and incubated for 24 h. The cells were harvested
in cold lysis buffer (50 mM Tris-HCl pH 8.0, 150 mM NaCl, 1% NP-40)
with Halt™ Protease and phosphatase inhibitor cocktail and EDTA.
The cells were lysed on ice for 30 min and centrifuged at 14,000
rpm for 20 min at 4°C. The proteins were quantified using the BCA
method. Protein (20 μl) was separated by SDS-PAGE and transferred
to PVDF membranes. The membranes were blocked with 5% skim milk in
0.1% TBS-T at room temperature for 2 h with rocking and then
incubated with the specific primary antibodies overnight at 4°C.
After washing the membrane with 0.1% TBS-T 3 times for 10 min each,
the membranes were incubated with the HRP-conjugated secondary
antibody at room temperature for 1 h. The membranes were detected
by SuperSignal West Dura Extended Duration Substrate. The membranes
were imaged with LAS-3000 (Fuji, Japan) according to the
manufacturer’s instructions.
Phospho-RTK array analysis
Phospho-RTK array analysis was carried out using a
human phospho-RTK array kit (R&D Systems). Phospho-RTK array
was performed according to the product manual. HCC827GR cells were
seeded in a 100-mm culture dish at 2×106 cells. The
cells were treated with SB365 (10 μM) and incubated for 24 h. The
cell lysates were prepared using NP40-lysis buffer containing the
protease-phosphatase inhibitor cocktail and EDTA. After blocking
for 1 h with Array Buffer 1, the arrays were incubated with 300 μg
of protein lysates overnight at 4°C. The arrays were then washed
and incubated with a HRP-conjugated phosphotyrosine detection
antibody. The arrays were detected by chemiluminescence and imaged
using LAS-3000 according to the manufacturer’s instructions. The
intensity of the average signal of the pair of duplicated spots was
calculated relative to the negative control spots.
Migration and invasion assay
Cell migration and invasion capacity in HCC827GR
cells were assessed using BD cell-culture inserts and/or BD BioCoat
Matrigel Invasion Chamber, respectively. The cells were suspended
in culture media (1×105 cells/ml) and then 0.5 ml cell
suspension (5×104 cells/ml) was added to the upper
chambers. After incubating for 24 h (for migration) or 48 h (for
invasion), culture media in the inserts were carefully removed, and
the membrane containing the cells on the lower surface of the
inserts was fixed using methanol and stained with H&E. The
upper surface of the membrane was gently scrubbed with a cotton
swab to remove the cells. The stained cells were quantified under a
light microscope.
Soft agar colony formation assay
HCC827GR cells (8×103 cells/well) were
suspended in BME (1 ml with 10% FBS, 0.33% agar) and plated over a
layer of solidified bottom agar mixture (BME with 10% FBS, 0.5%
agar) with the indicated concentrations of the drugs. The cultures
were maintained at 37°C in a 5% CO2 incubator for 6–7
days, and the number of colonies was counted under a light
microscope.
Flow cytometry analysis
For assessing cell cycle distribution, the cells
were detached and fixed overnight in ice-cold 70% ethanol. Fixed
cells were washed with phosphate-buffered saline (PBS), and
incubated with RNase A and propidium iodide (PI) at 37°C for 30
min, and then cells were subjected to flow cytometric analysis to
determine the percentage of cells at specific phases of the cell
cycle. Apoptosis in HCC827GR cells was also measured using the
Annexin V-FITC Apoptosis Detection Kit I (BD Biosciences
Pharmingen) according to the product protocols. The cells were
seeded in a 60-mm culture dish at 5×105 cells. The cells
were treated with various concentrations of the compounds and
incubated for 24 h. The cells were detached and then washed twice
with cold PBS. The cells were suspended in 1× binding buffer
(5×105 cells/ml) and Annexin V-FITC and PI (5 μl
stock/100 μl buffer) were added. After incubation at room
temperature for 15 min in the dark, 400 μl of 1× binding buffer was
added to each tube. Flow cytometric analysis was performed using a
BD FACSVerse flow cytometer and BD FACSuite software.
Statistical analysis
All quantitative data are presented as mean value ±
SD unless indicated otherwise. The statistical significance of
compared measurements was measured using the Student’s t-test, and
p<0.05 was considered to indicate a statistically significant
result.
Results
Characterization of the
gefitinib-resistant NSCLC cell line, HCC827GR
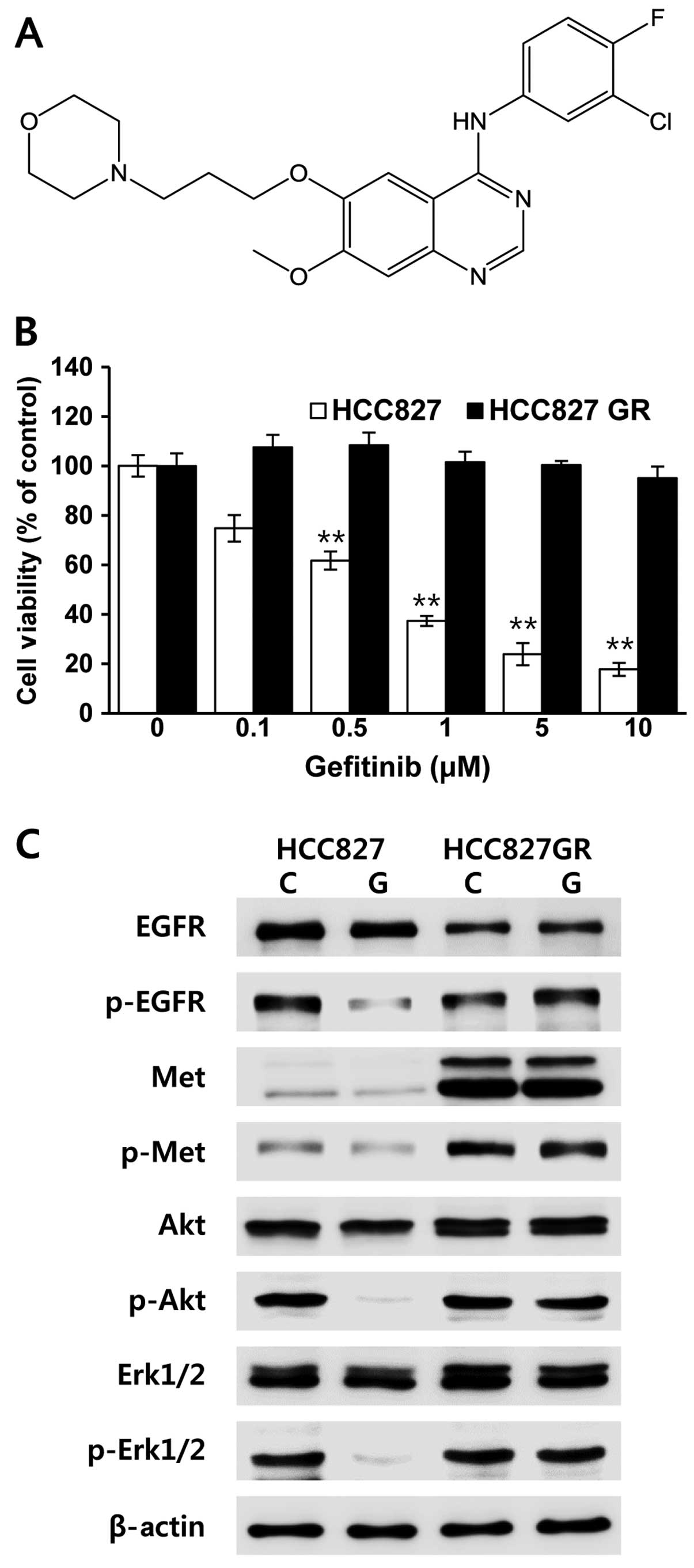
Gefitinib-resistant HCC827 cells (HCC827GR) were
generated by continuously exposing the cells to increasing
concentrations of gefitinib as reported (14,26).
The resultant HCC827GR cells were characterized by quantifying cell
viability at different concentrations of gefitinib (Fig. 1A). In order to obtain this, the
cells were incubated with the indicated concentration of gefitinib
up to 72 h, and cell proliferation was determined by MTT assay. As
expected, our data showed that HCC827GR cells were resistant to
gefitinib treatment when compared with the parental HCC827 cells
(Fig. 1B). We further analyzed the
protein expression and phosphorylation status of EGFR, Met and
downstream signaling molecules in the HCC827 and HCC827GR cells.
Our data clearly revealed that an abundant expression of Met
protein was observed in the gefitinib-resistant HCC827GR cells,
which was associated with emergence of acquired resistance
(Fig. 1C). Unlike HCC827 cells, the
phosphorylation status of EGFR, Met, Akt and Erk persisted at
higher levels in the HCC827GR cells even following treatment with
gefitinib (Fig. 1C). Therefore, our
results indicate that gefitinib-resistant HCC827GR cells were
successfully generated, and the bypass signaling mediated by Met
led to persistent activation of downstream signaling in the
HCC827GR cells in the presence of gefitinib.
SB365 inhibits anchorage-dependent and
-independent growth of HCC827GR cells
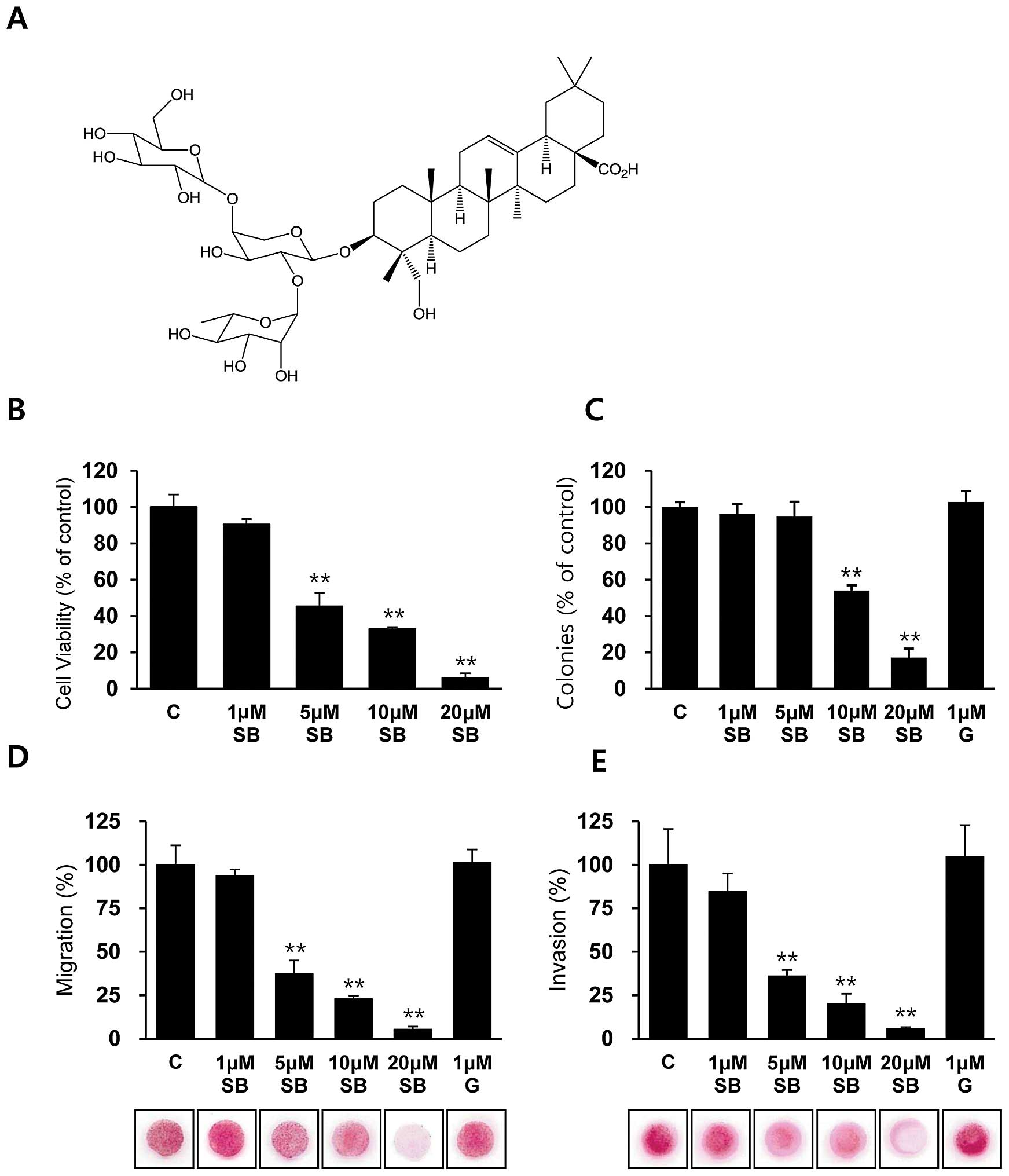
To assess the inhibitory activity of SB365 (Fig. 2A) on the proliferation of
gefitinib-resistant HCC827GR cells, HCC827GR cells were treated
with the indicated concentrations of SB365 up to 72 h, and cell
viability was determined by the MTT assay. Notably, treatment with
SB365 decreased the cell viability of the HCC827GR cells in a
dose-dependent manner (Fig. 2B),
whereas co-treatment with gefitinib did not show any addictive or
synergistic effects (data not shown). Next, we investigated the
antitumor effect of SB365 in HCC827GR cells by performing an
anchorage-independent colony formation assay, which is an in
vitro indicator and a key characteristic of the transformed
cell phenotype (27). Our data
revealed that treatment with SB365 resulted in fewer colonies being
formed in soft agar at concentrations of 10 and 20 μM SB365
compared with the control cells or gefitinib-treated cells
(Fig. 2C). These data indicate that
treatment with SB365 inhibits both anchorage-dependent and
-independent growth of HCC827GR cells, suggesting that the
malignant potential of HCC827GR cells is reduced by SB365 which may
be a potential natural compound for overcoming acquired resistance
to gefitinib.
Effects of SB365 on the migration and
invasion of gefitinib-resistant HCC827GR cells
To investigate whether SB365 has antimetastatic
activity in vitro, we performed migration and invasion
assays in gefitinib-resistant HCC827GR cells. Our data showed that
treatment with 1 μM gefitinib did not affect the migratory and
invasive capacities, whereas dose-dependent treatment with SB365
strongly inhibited the migratory and invasive capacities of the
gefitinib-resistant HCC827GR cells up to 90% at a dose of 20 μM
(Fig. 2D and E). Therefore, these
data revealed that SB365 has an antimetastatic effect on
gefitinib-resistant HCC827GR cells.
SB365 inhibits the phosphorylation of Met
and downstream signaling in HCC827GR cells
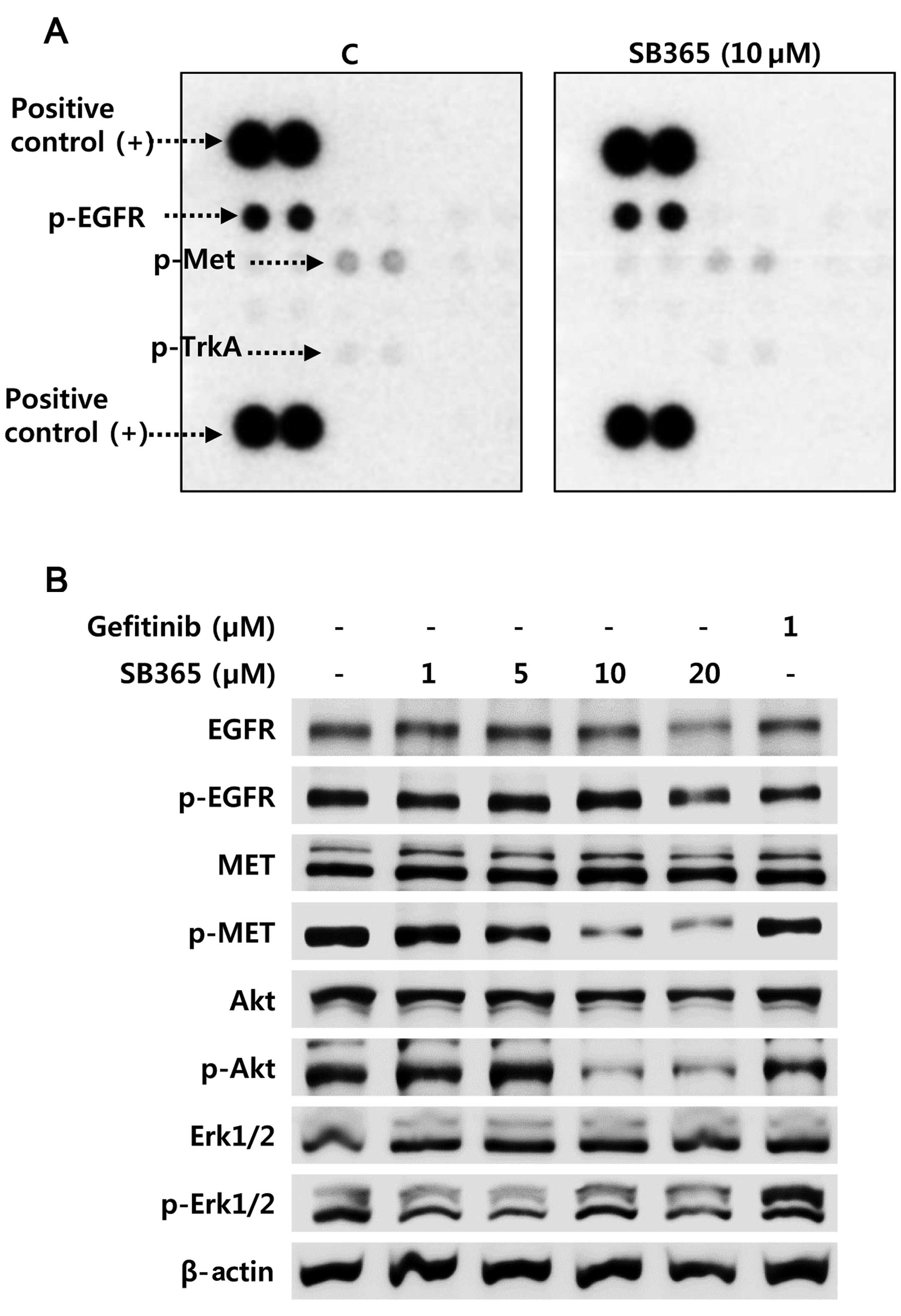
To explore the underlying mechanism by which SB365
inhibits anchorage-independent cell growth and in vitro
metastatic capacities in gefitinib-resistant HCC827GR cells, we
next performed a human phospho-RTK array to investigate the
relative levels of phosphorylation of different RTKs. In agreement
with a previous report (22), our
data also revealed that SB365 may be a possible inhibitor of c-Met
in NSCLCs. Notably, a decrease in phosphorylated-Met was observed
in the HCC827GR cells after SB365 treatment, whereas the
phosphorylation level of EGFR was not changed in the 10 μM
SB365-treated HCC827GR cells (Fig.
3A). To better understand the mechanism responsible for c-Met
inhibition by SB365, we examined the protein expression and
phosphorylation status of RTK proteins including EGFR, Met and
downstream signaling molecules in the HCC827GR cells. In order to
obtain this, cells were treated with 1 μM of gefitinib or various
concentrations of SB365 at 1, 5, 10 and 20 μM for 24 h. As
expected, treatment with 1 μM gefitinib did not affect the
phosphorylation status of EGFR, Met, Akt and Erk1/2 in the
gefitinib-resistant HCC827GR cells (Fig. 3B). When HCC827GR cells were treated
with various concentrations of SB365, the phosphorylation levels of
Met and downstream signaling molecule Akt but not Erk1/2 were
effectively suppressed at doses of 10 or 20 μM SB365 (Fig. 3B). Therefore, these findings
indicate that SB365 suppressed cancer progression through the
Met-Akt signaling pathway in Met-amplified NSCLC HCC827GR
cells.
SB365 induces apoptosis in the HCC827GR
cells
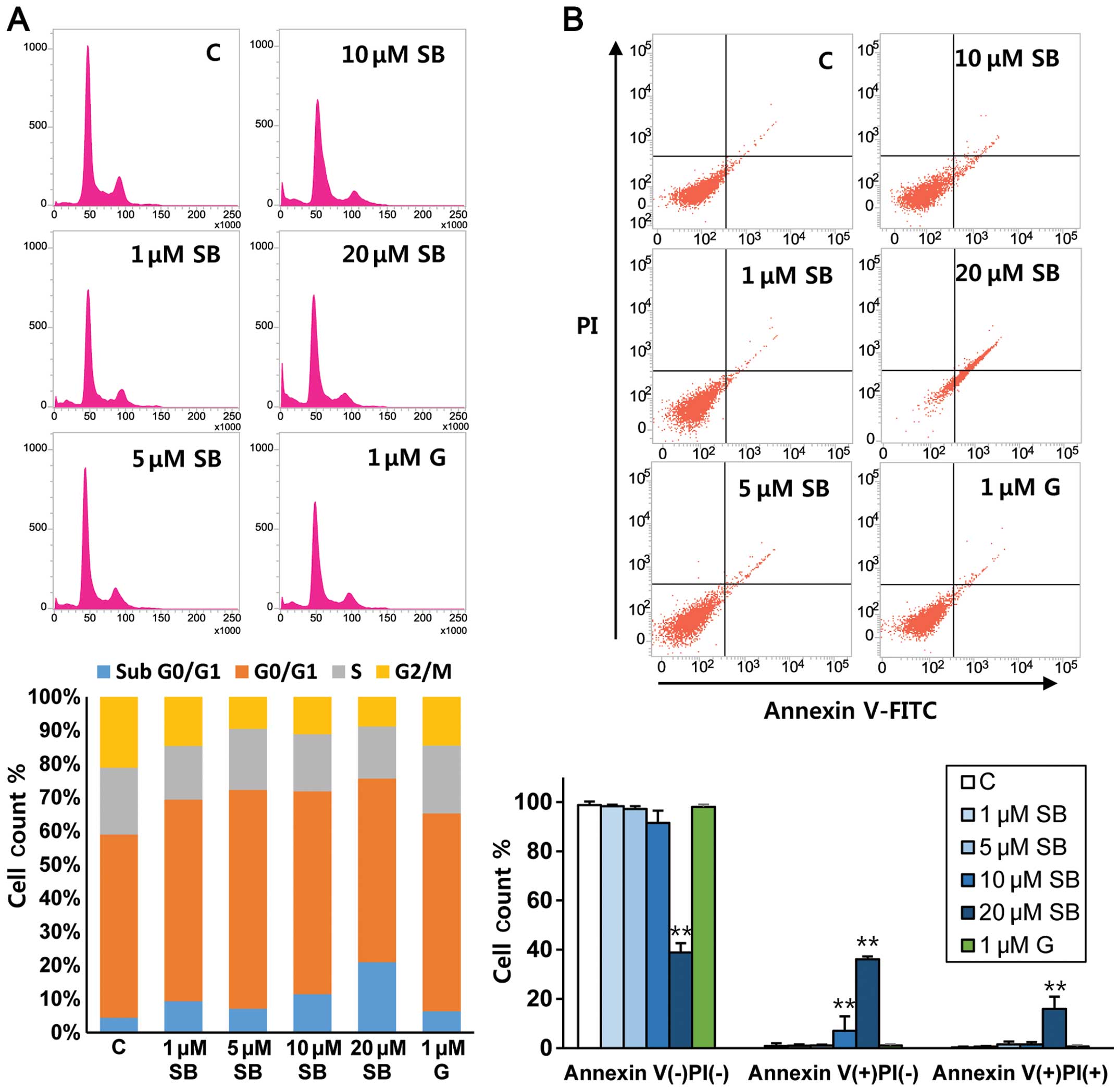
To determine the pro-apoptotic effect of SB365 on
gefitinib-resistant HCC827GR cells, flow cytometric analysis was
performed. An increased proportion of apoptotic cells in the sub-G1
phase was strongly detected following dose-dependent treatment with
SB365 (4.9–13.5%) compared with the control (2.5%), indicating a
pro-apoptotic effect of SB365 in the HCC827GR cells (Fig. 4A). Moreover, flow cytometric
analysis with Annexin V showed that treatment with SB365 induced
early apoptosis (Annexin V+/PI−) in the
HCC827GR cells, yet not with gefitinib (Fig. 4B). A few late apoptotic cells
(Annexin V+/ PI+) were also observed in the
cells treated with 20 μM SB365 (Fig.
4B). More importantly, its apoptotic effect was accompanied by
increased levels of cleaved caspase-3 and poly(ADP ribose)
polymerase (Fig. 4C). Moreover,
treatment with SB365 increased the protein expression levels of
cleaved caspase-3 and PARP and decreased Bcl-2 expression (Fig. 4C). These findings clearly suggest
that treatment with SB365 is sufficient for inducing death
signaling even in gefitinib-resistant NSCLC HCC827GR cells with Met
amplification.
Discussion
To date, reversible small-molecule EGFR-TKIs,
including gefitinib (Iressa®) and erlotinib
(Tarceva®), have been recommended as molecular-targeted
anticancer drugs for patients with NSCLC with EGFR-activating
mutations (9,10). However, despite excellent initial
responses to EGFR-TKIs in these NSCLC patients, nearly all patients
eventually develop drug resistance after a median period of ~10
months through two genetically conferred mechanisms; a secondary
T790M mutation in EGFR and c-Met gene amplification (12–16).
For this reason, a strategy using irreversible EGFR-TKIs or the
combined use of EGFR- and Met-TKIs has been proposed to overcome
resistance to EGFR-TKIs. Considering this, it is crucial to
establish an alternative treatment strategy overcoming resistance
to EGFR-TKIs to improve the survival of patients with NSCLC.
A recent trend in anticancer therapy has been
centered on the search for anticancer drugs which are safer and
highly patient acceptable. Consistently, the use of medicinal
compounds from herbal/natural sources is becoming increasingly
popular over synthetic drugs as an alternative treatment choice for
cancer patients (28). Based on
this notion, we isolated SB365, a saponin D from Pulsatilla
koreana and explored its anticancer effects on
gefitinib-resistant NSCLC cells. Previous results suggest that
saponins have been known to possess a wide range of anticancer
properties such as chemopreventive or chemotherapeutic effects on
various types of cancer cells (29,30).
We also found that SB365, a saponin D, exerted remarkable
anticancer properties in several types of cancer cells including
hepatocellular carcinoma, gastric, colon and pancreatic cancers
(21–24). In the present study, we showed that
SB365 has an inhibitory effect on the anchorage-independent
proliferation, migration and invasion of HCC827GR NSCLC cells
resistant to gefitinib. Moreover, SB365 was sufficient for inducing
apoptosis in gefitinib-resistant HCC827GR cells. These effects of
SB365 against gefitinib resistance may be attributed to its ability
to inhibit both Met phosphorylation and subsequent Akt activation
in Met-amplified HCC827GR cells.
Accumulated evidence suggests that identification of
natural compounds with high anticancer efficacy and defined
molecular targets could be a prerequisite for successful drug
discovery. For example, various phytochemicals, including
[6]-gingerol (31), resveratrol
(32) and myricetin (33), have been reported to directly
interact with their targets in cells, thereby inhibiting cancer
cell proliferation. Consistent with this, we previously reported
that SB365 effectively inhibited the phosphorylation of Met by
binding to an allosteric site of Met in MKN-45 gastric cancers,
indicating that SB365 may be a potential natural inhibitor of Met
(22). Met is a well-established
mediator of carcinogenesis and induces an invasive program
consisting of cell proliferation, migration, invasion and survival
(34). In addition, Met activation
prevents cancer cell apoptosis through the PI3K-Akt signaling
pathway (35). Based on the notion
that EGFR-TKI resistance can be accompanied by Met amplification,
SB365 may be effectively used for the treatment of Met-amplified
NSCLCs.
In the present study, we generated
gefitinib-resistant HCC827GR cells with Met amplification and then
evaluated the anticancer activity of SB365. The present data
demonstrated that SB365 inhibited anchorage-independent
proliferation, migration, and invasion of HCC827GR cells and
induced cancer cell apoptosis by a caspase-dependent manner.
Moreover, our phospho-array data revealed that the phosphorylation
of Met, not EGFR, was robustly decreased by treatment with SB365 in
HCC827GR cells. Collectively, our findings suggest that SB365 is a
new therapeutic candidate to overcome resistance to EGFR-TKIs in
NSCLCs harboring Met amplification.
Acknowledgements
This study was conducted by the Settlement Research
Grant of Keimyung University in 2011 (to C-H.J.).
References
|
1
|
Jung KW, Park S, Kong HJ, et al: Cancer
statistics in Korea: incidence, mortality and survival in
2006–2007. J Korean Med Sci. 25:1113–1121. 2010.
|
|
2
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar
|
|
3
|
Owonikoko TK, Ragin CC, Belani CP, et al:
Lung cancer in elderly patients: an analysis of the surveillance,
epidemiology, and end results database. J Clin Oncol. 25:5570–5577.
2007. View Article : Google Scholar
|
|
4
|
Sher T, Dy GK and Adjei AA: Small cell
lung cancer. Mayo Clin Proc. 83:355–367. 2008. View Article : Google Scholar
|
|
5
|
Fukuoka M, Yano S, Giaccone G, et al:
Multi-institutional randomized phase II trial of gefitinib for
previously treated patients with advanced non-small-cell lung
cancer (The IDEAL 1 Trial) [corrected]. J Clin Oncol. 21:2237–2246.
2003.
|
|
6
|
Hotta K, Matsuo K, Ueoka H, Kiura K,
Tabata M and Tanimoto M: Addition of platinum compounds to a new
agent in patients with advanced non-small-cell lung cancer: a
literature based meta-analysis of randomised trials. Ann Oncol.
15:1782–1789. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shepherd FA, Rodrigues Pereira J, Ciuleanu
T, et al: Erlotinib in previously treated non-small-cell lung
cancer. N Engl J Med. 353:123–132. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Laskin JJ and Sandler AB: Epidermal growth
factor receptor: a promising target in solid tumours. Cancer Treat
Rev. 30:1–17. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Maemondo M, Inoue A, Kobayashi K, et al:
Gefitinib or chemotherapy for non-small-cell lung cancer with
mutated EGFR. N Engl J Med. 362:2380–2388. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mitsudomi T, Morita S, Yatabe Y, et al:
Gefitinib versus cisplatin plus docetaxel in patients with
non-small-cell lung cancer harbouring mutations of the epidermal
growth factor receptor (WJTOG3405): an open label, randomised phase
3 trial. Lancet Oncol. 11:121–128. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rosell R, Moran T, Queralt C, et al:
Screening for epidermal growth factor receptor mutations in lung
cancer. N Engl J Med. 361:958–967. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pao W, Miller VA, Politi KA, et al:
Acquired resistance of lung adenocarcinomas to gefitinib or
erlotinib is associated with a second mutation in the EGFR kinase
domain. PLoS Med. 2:e732005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bean J, Brennan C, Shih JY, et al:
MET amplification occurs with or without T790M
mutations in EGFR mutant lung tumors with acquired
resistance to gefitinib or erlotinib. Proc Natl Acad Sci USA.
104:20932–20937. 2007. View Article : Google Scholar
|
|
14
|
Engelman JA, Zejnullahu K, Mitsudomi T, et
al: MET amplification leads to gefitinib resistance in lung
cancer by activating ERBB3 signaling. Science. 316:1039–1043. 2007.
View Article : Google Scholar
|
|
15
|
Yano S, Wang W, Li Q, et al: Hepatocyte
growth factor induces gefitinib resistance of lung adenocarcinoma
with epidermal growth factor receptor-activating mutations. Cancer
Res. 68:9479–9487. 2008. View Article : Google Scholar
|
|
16
|
Sequist LV, Waltman BA, Dias-Santagata D,
et al: Genotypic and histological evolution of lung cancers
acquiring resistance to EGFR inhibitors. Sci Transl Med.
3:75ra262011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Engelman JA, Zejnullahu K, Gale CM, et al:
PF00299804, an irreversible pan-ERBB inhibitor, is effective in
lung cancer models with EGFR and ERBB2 mutations that
are resistant to gefitinib. Cancer Res. 67:11924–11932. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tang Z, Du R, Jiang S, et al: Dual
MET-EGFR combinatorial inhibition against T790M-EGFR-mediated
erlotinib-resistant lung cancer. Br J Cancer. 99:911–922. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Saha SK and Khuda-Bukhsh AR: Molecular
approaches towards development of purified natural products and
their structurally known derivatives as efficient anti-cancer
drugs: current trends. Eur J Pharmacol. 714:239–248. 2013.
View Article : Google Scholar
|
|
20
|
Youn HJ, Lakritz J, Kim DY, Rottinghaus GE
and Marsh AE: Anti-protozoal efficacy of medicinal herb extracts
against Toxoplasma gondii and Neospora caninum. Vet
Parasitol. 116:7–14. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hong SW, Jung KH, Lee HS, et al: SB365
inhibits angiogenesis and induces apoptosis of hepatocellular
carcinoma through modulation of PI3K/Akt/mTOR signaling pathway.
Cancer Sci. 103:1929–1937. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hong SW, Jung KH, Lee HS, et al: SB365,
Pulsatilla saponin D, targets c-Met and exerts
antiangiogenic and antitumor activities. Carcinogenesis.
34:2156–2169. 2013.PubMed/NCBI
|
|
23
|
Son MK, Jung KH, Hong SW, et al: SB365,
Pulsatilla saponin D suppresses the proliferation of human
colon cancer cells and induces apoptosis by modulating the AKT/mTOR
signalling pathway. Food Chem. 136:26–33. 2013.PubMed/NCBI
|
|
24
|
Son MK, Jung KH, Lee HS, et al: SB365,
Pulsatilla saponin D suppresses proliferation and induces
apoptosis of pancreatic cancer cells. Oncol Rep. 30:801–808.
2013.PubMed/NCBI
|
|
25
|
Kim Y, Bang SC, Lee JH and Ahn BZ:
Pulsatilla saponin D: the antitumor principle from
Pulsatilla koreana. Arch Pharm Res. 27:915–918. 2004.
View Article : Google Scholar
|
|
26
|
Ghosh G, Yan X, Lee AG, Kron SJ and
Palecek SP: Quantifying the sensitivities of EGF receptor (EGFR)
tyrosine kinase inhibitors in drug resistant non-small cell lung
cancer (NSCLC) cells using hydrogel-based peptide array. Biosens
Bioelectron. 26:424–431. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Freedman VH and Shin SI: Cellular
tumorigenicity in nude mice: correlation with cell growth in
semi-solid medium. Cell. 3:355–359. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhao J: Nutraceuticals, nutritional
therapy, phytonutrients, and phytotherapy for improvement of human
health: a perspective on plant biotechnology application. Recent
Pat Biotechnol. 1:75–97. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Taylor WG, Elder JL, Chang PR and Richards
KW: Micro-determination of diosgenin from fenugreek (Trigonella
foenum-graecum) seeds. J Agric Food Chem. 48:5206–5210. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Price KR, Johnson IT and Fenwick GR: The
chemistry and biological significance of saponins in foods and
feedingstuffs. Crit Rev Food Sci Nutr. 26:27–135. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jeong CH, Bode AM, Pugliese A, et al:
[6]-Gingerol suppresses colon cancer growth by targeting
leukotriene A4 hydrolase. Cancer Res. 69:5584–5591.
2009.
|
|
32
|
Oi N, Jeong CH, Nadas J, et al:
Resveratrol, a red wine polyphenol, suppresses pancreatic cancer by
inhibiting leukotriene A4 hydrolase. Cancer Res.
70:9755–9764. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jung SK, Lee KW, Byun S, et al: Myricetin
inhibits UVB-induced angiogenesis by regulating PI-3 kinase in
vivo. Carcinogenesis. 31:911–917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Birchmeier C, Birchmeier W, Gherardi E and
Vande Woude GF: Met, metastasis, motility and more. Nat Rev Mol
Cell Biol. 4:915–925. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xiao GH, Jeffers M, Bellacosa A, Mitsuuchi
Y, Vande Woude GF and Testa JR: Anti-apoptotic signaling by
hepatocyte growth factor/Met via the phosphatidylinositol
3-kinase/Akt and mitogen-activated protein kinase pathways. Proc
Natl Acad Sci USA. 98:247–252. 2001. View Article : Google Scholar : PubMed/NCBI
|