Introduction
Gastric carcinoma is one of the most common
malignant tumors. Recently, research has shown that the genesis and
development of gastric carcinoma is a multi-step and multi-gene
process. Therefore, it is extremely important to investigate the
pathogenesis of gastric cancer and search for new targets of
prevention and therapy and effective drugs for decreasing morbidity
and mortality.
Receptor tyrosine kinases (RTKs) transmit
extracellular signals to the nucleus, and then initiates a cascade
of signaling events that modulate cellular responses. The
erythropoietin-producing hepatocellular (EPH) gene family
represents the largest known family of receptor tyrosine kinases.
EphA2 was the first gene identified with tyrosine kinase activity
in the EPH family. In normal cells, EphA2 appears to be restricted
to intercellular junctions between epithelial cells, where it binds
ligands and anchors to the membrane of adjacent cells. It then
initiates tyrosine kinases in the cytoplasm and participates in
embryonic development, cell migration and vasiformation (1). A normal EphA2 expression signal
weakens the Ras/MAPK cascade reaction, which is activated by PDEF,
EGF, VEGF, and restrains cell growth. It then negatively regulates
cellular adherin through breaking spot-adherin. EphA2 has been
given more and more attention in carcinogenesis and development in
recent years. EphA2 is overexpressed in many human cancers such as
breast, colon, prostate and non-small cell lung cancer (NSCLC)
(2–4), whose growth, transformation and
metastasis correlate with EphA2 mutation, overexpression, abnormal
location and (or) abnormal inactivation. Moreover, EphA2 is a new
target for clinical diagnosis and therapy, while there is no exact
experimental base for abormal expression of EphA2 in gastric
carcinoma.
Geldanamycin (GA), a benzoquinoid ansamycin
antibiotic, targets heat shock protein 90 (Hsp90) (5). It is an inhibitor of Hsp90. Hsp90 is a
group of highly conservative molecular chaperones, which is widely
found in organisms. EphA2 is abnormally highly overexpressed in
tumors, and its downstream proteins take part in key points of
tumor progression. EphA2, as a member of the RTK family, is
believed to be a downstream protein of Hsp90. Recently, GA has
become a novel target for anticancer drug development due to its
inhibition of cell proliferation and induction of apoptosis.
However, whether GA suppresses cell proliferation of gastric
carcinoma and the possible mechanisms remain undetermined.
Therefore, in this study, we detected the role of
the expression of EphA2 in the carcinogenesis and progression of
gastric carcinoma in order to explore the relationship between
EphA2 expression and proliferation and apoptosis in gastric
carcinoma. Furthermore, RNAi was used to silence EphA2 expression
to investigate the precise effect of EphA2 on cell proliferation
and apoptosis of gastric cancer cells. Moreover we determined the
effect of GA, an inhibitor of Hsp90, on the proliferation and
apoptosis of human gastric carcinoma MGC803 cells, which may
elucidate the mechanism of the carcinogenesis and progression of
gastric carcinoma as well.
Materials and methods
Reagents and antibodies
TRIzol reagent and Lipofectamine 2000 were obtained
from Invitrogen (Carlsbad, CA, USA). Horseradish peroxidase (HRP)
Affinipure goat anti-mouse/rabbit IgG (H+L) and SP9000 were
purchased from Zhongshan Golden Bridge Biotechnology Co., Ltd.
(Beijing, China). FITC Affinipure goat anti-mouse IgG was from
Jackson ImmunoResearch Co. The pCDNA3.1 vector and pCDNA3.1/EphA2
were obtained from Invitrogen Co. The rabbit anti-EphA2 antibodies
were from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Clinical materials
Eighty-two cases of surgically resected primary
gastric carcinoma tissues, paired adjacent mucosa (2–5 cm from the
margin of the gastric carcinoma) and normal mucosa at the surgical
margin (at least 5 cm from the margin of the gastric carcinoma and
histologically proven) tissues collected between January and July
2007 at the Inpatient Department of the Fourth Hospital of Hebei
Medical University were used in this study. These gastric carcinoma
tissues included 30 cases of well-differentiated gastric carcinoma
and 52 cases of moderately and poorly differentiated gastric
carcinoma [superficial muscle invasive (n=20), deep muscle invasive
(n=62), lymph node metastasis positive (n=51), lymph node
metastasis negative (n=31)]. Thirty cases were randomly chosen to
detect EphA2 mRNA and protein expression [9 cases of well
differentiated tissues and 21 cases of moderately and poorly
differentiated tissues; superficial muscle invasive (n=8), deep
muscle invasive (n=22), lymph node metastasis positive (n=21),
lymph node metastasis negative (n=9)]. Experimental protocols were
approved by the Institutional Human Care and Use Committee of Hebei
Medical University.
Cell lines and treatment groups
Human gastric cancer cells (MGC803) were grown in
humidified air with 5% CO2 in an incubator at 37°C in
RPMI-1640 supplemented with 10% fetal bovine serum, 100 μg/ml
streptomycin and 100 U/ml penicillin. In order to investigate the
effect of GA on cell proliferation, the cells were divided into 20,
40, 200, 400 and 2,000 nmol/l groups. In order to explore the
mechanism of GA on apoptosis of MGC803 cells, the cells were
divided into six groups: control, GA (200 nmol/l), GA +
pYr-3.1-shNC, GA + pYr-3.1-shNC; GA + pcDNA3.1-EphA2-IRES-EGFP, and
GA + pcDNA3.1-NC-IRES-EGFP; and were collected at 48 h.
Immunohistochemistry and
immunocytochemistry for detection of EphA2 protein
The tissues were fixed in 4% formaldehyde. Antigen
recovery was performed using a microwave. The sections were
incubated with primary antibodies against EphA2 (1:100) overnight
at 4°C. On the following day, the sections were incubated with
polyperoxidase anti-mouse IgG at 37°C, and finally stained with
diaminobenzidine. The sections were imaged with an Olympus
microscope. Positive staining of EphA2 protein was located in the
cytoplasm or/and membrane of epithelial cells. The results were
divided into 4 grades as previously described (6).
The cells were fixed in a 4% paraformaldehyde
solution and permeabilized with 0.1% Triton X-100. Following
incubation with anti-EphA2 (1:100; Santa Cruz) antibodies and the
secondary antibody conjugated with HRP, the slices were stained
with diaminobenzidine using the method introduced by Feng et
al (7).
RT-PCR
Total RNA was extracted from gastric cancer tissues
or MGC830 cells with TRIzol reagent according to the instructions
of the manufacturer. Total RNA (2 μg) was reverse transcribed into
cDNA by AMV reverse transcriptase at 42°C for 1 h and then heated
to 94°C for 5 min in a total reaction volume of 20 μl. The PCR
condition used for EphA2 and the internal reference GAPDH was 94°C
for 5 min followed by 33 cycles of 94°C for 45 sec, 60°C for 45 sec
and 72°C for 45 sec, and 72°C for 7 min. The specific primers were
as follows: EphA2 sense, 5′-CCAAGTTCGCTGACATCGT-3 and anti-sense,
5′-GCCATGAAGTGCTCCGTAT-3′; GAPDH sense, 5′-ACCACAGTCCATGCCATCAC-3′
and antisense, 5′-TCC ACCACCCTGTTGCTGTA-3′. The expected PCR
products were 196 and 486 bp for EphA2 and GAPDH, respectively. The
amplicons were analyzed by electrophoresis, imaged using UVI gel
imaging system and quantified using Gel-Pro Analyzer 3.1 software.
Expression levels of EphA2 were normalized to the internal
reference GAPDH.
Total protein extraction and western
blotting
Total protein extraction from the collected tissues
and cultured cells was performed as described previously (8). The protein extracts were separated by
10% SDS-PAGE and then transferred to PVDF membranes. The membranes
were incubated overnight at 4°C with anti-EphA2 (1:100; Santa Cruz)
and β-actin (1:200; Santa Cruz) antibodies. Subsequently, the
membranes were incubated with HRP-conjugated goat anti-rabbit IgG
(1:5,000) and then exposed to X-ray film using an enhanced
chemiluminescence system (Pierce Biotechnology, Rockford, IL, USA).
EphA2 protein expression was quantified by comparison with
β-actin.
Measurement of cell proliferation by
MTT
MGC803 cells in a logarithmic phase were inoculated
into a 96-well plate with 1×105 cells in each well. Cell
viability at 24 h was examined using the MTT method.
OD490nm values were detected in six duplicate wells and
their averages were used to plot the growth curve and calculate the
growth inhibition rate of each treatment using the following
formula: Growth inhibition rate (IR) = OD490nm of the
control group − OD490nm of the treatment
group/OD490nm of the control group × 100%.
Flow cytometric analysis
The tissues and the cell pellets treated by GA were
fixed in 70% ethanol, and stained with 500 μg/ml propidium iodide
(Sigma) containing 3 Kunitz units of RNase and 1% fetal bovine
serum in PBS. Then, the stained cells were analyzed in an
Epics-XLII flow cytometer (Beckman Coulter, Miami, FL, USA) and the
number of the cells in each phase of the cell cycle was calculated
using the ModFit LT cell cycle analysis program according to the
manufacturer’s instructions. The proliferation index (PI)
represents the number of proliferating cells and was calculated
using the following equation: PI = (S + G2M)/(S +
G2M + G0/G1) × 100%.
Transient transfection
The sh-EphA2 vector and negative control sh-Scramble
vector were designed and produced by Invitrogen. The expression
vector of EphA2 (pcDNA3.1-EphA2-IRES-EGFP) and the control vector
(pcDNA3.1-IRES-EGFP) were designed and produced by the Beijing
Fungenome Technology Co. (Beijing, China). Transient transfection
of MGC803 cells was carried out using Lipofectamine 2000 according
to the manufacturer’s instructions. At ~80% confluency, the cells
were transfected with 2.0 μg of vector DNA with 4 μl of
Lipofectamine 2000 in 2 ml of serum-free RPMI-1640 medium. Six
hours after the transfection, the cells were incubated for 48 h
with normal RPMI-1640 medium containing 10% fetal bovine serum.
Then, 200 nmol/l GA was added to the medium, and the cells were
cultured for 48 h. Subsequently, EphA2 expression, PI, cell cycle
distribution and apoptosis were determined.
Statistical analysis
Data are expressed as mean ± standard variation and
were analyzed using SPSS 15.0 statistical software package.
Differences between samples were tested using single factor
analysis of variance and LSD method for multiple comparisons. A
p-value <0.05 was considered as a significant difference. Before
comparison, data homogeneity of variance was first examined using
the F test. In the case of heterogeneity of variance, the
approximate variance F test/Welch method was used.
Results
EphA2 protein expression is increased in
the gastric carcinoma tissues and is positively correlated with the
degree of differentiation and apoptosis
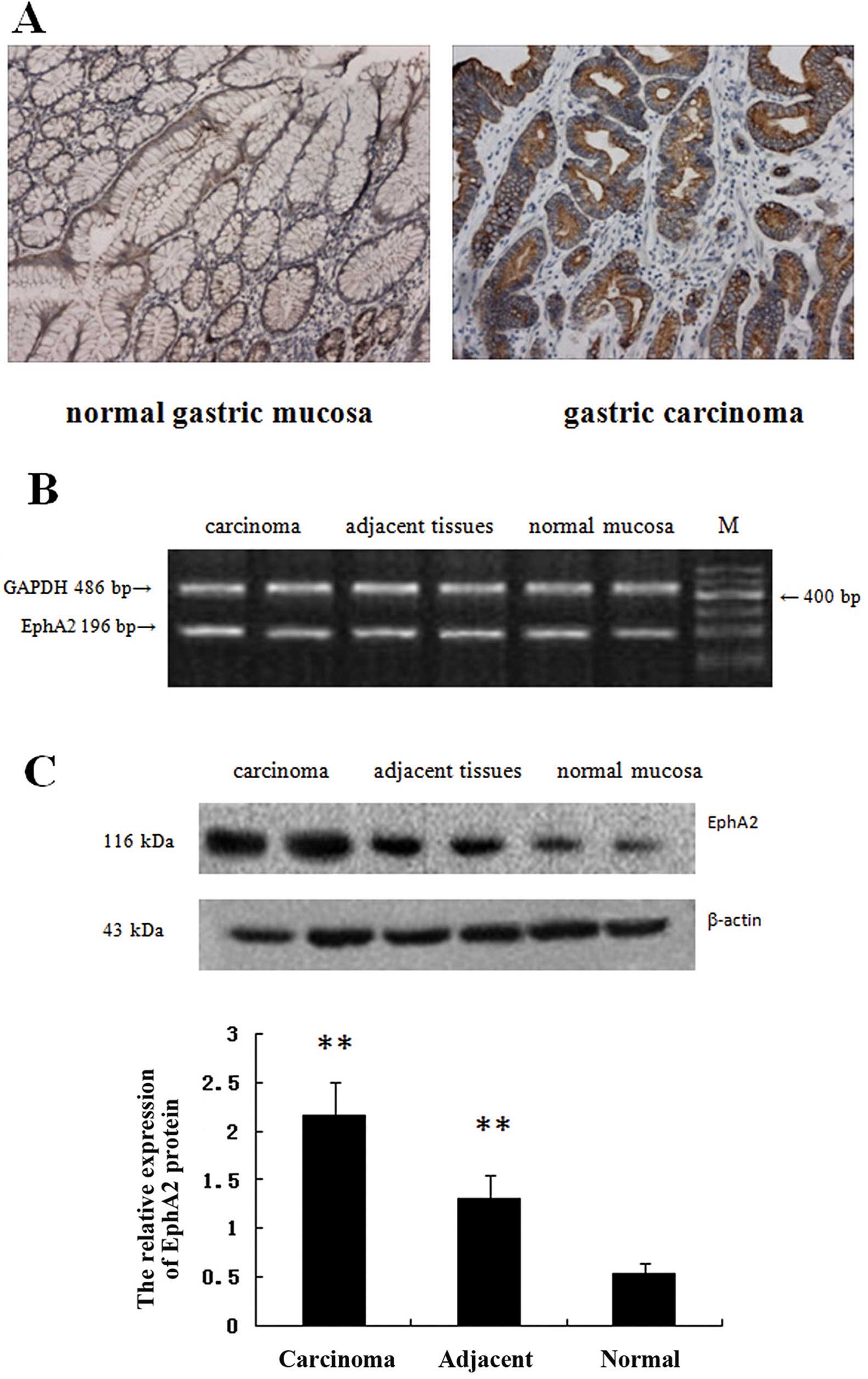
As shown in Fig. 1A
and Table I, positive expression of
EphA2 protein was noted in the cytoplasm and/or membrane and was
higher in the carcinoma tissues than that in the adjacent and
normal tissues (P<0.01). Moreover, the positive expression rate
was higher in moderately and poorly differentiated tissues than
that in the well-differentiated tissues (P<0.05). In addition,
the expression of EphA2 in tumors with deep infiltration was higher
than that in tumors with superficial layer invasion (P<0.05).
Higher expression was noted in cases with lymph node metastasis
than in non-lymph node metastasis (P<0.05, Tables II and III). There was no difference in EphA2
mRNA among the tissue groups as shown by RT-PCR (P>0.05,
Fig. 1B), while the results of the
western blot analysis were identical with IHC and the EphA2 protein
expression was increased in the gastric carcinoma tissues.
 | Table IExpression of EphA2 in the different
tissue groups (IHC). |
Table I
Expression of EphA2 in the different
tissue groups (IHC).
| Tissue groups | n | − | + | ++ | +++ |
|---|
| Carcinoma | 82 | 6 | 16 | 28 | 32 |
| Adjacent | 82 | 35 | 26 | 19 | 2a |
| Normal | 82 | 49 | 22 | 11 | 0a |
| Table IIExpression of EphA2 in gastric
carcinoma tissues and the correlation with clinicopathological
features (IHC). |
Table II
Expression of EphA2 in gastric
carcinoma tissues and the correlation with clinicopathological
features (IHC).
| Features | n | − | + | ++ | +++ | P-value |
|---|
| Differentiation | | | | | | |
| High | 30 | 3 | 9 | 12 | 6 | 0.019 |
| Moderate/low | 52 | 3 | 7 | 16 | 26 | |
| Depth of
invasion | | | | | | |
| Superficial | 20 | 4 | 8 | 3 | 5 | 0.017 |
| Deep | 62 | 12 | 20 | 29 | 1 | |
| Lymphatic
metastasis | | | | | | |
| Yes | 51 | 4 | 5 | 16 | 26 | 0.009 |
| No | 31 | 2 | 11 | 12 | 6 | |
| Table IIIRelationship of EphA2 mRNA and
protein expression (western blot analysis) with clinicopathological
features of the gastric cancer cases. |
Table III
Relationship of EphA2 mRNA and
protein expression (western blot analysis) with clinicopathological
features of the gastric cancer cases.
| Features | n | EphA2 mRNA (mean ±
SD) | P-value | EphA2 protein (mean
± SD) | P-value |
|---|
| Tumor
differentiation |
| High | 9 | 1.420±0.121 | 0.912 | 1.792±0.194 | 0.026 |
| Moderate/low | 21 | 1.462±0.406 | | 2.329±0.652 | |
| Depth of
invasion |
| Superficial | 8 | 1.384±0.109 | 0.354 | 1.576±0.372 | 0.020 |
| Deep | 22 | 1.473±0.242 | | 2.383±0.890 | |
| Lymphatic
metastasis |
| Yes | 21 | 1.450±0.369 | 0.978 | 2.201±0.787 | 0.596 |
| No | 9 | 1.418±0.131 | | 1.959±0.360 | |
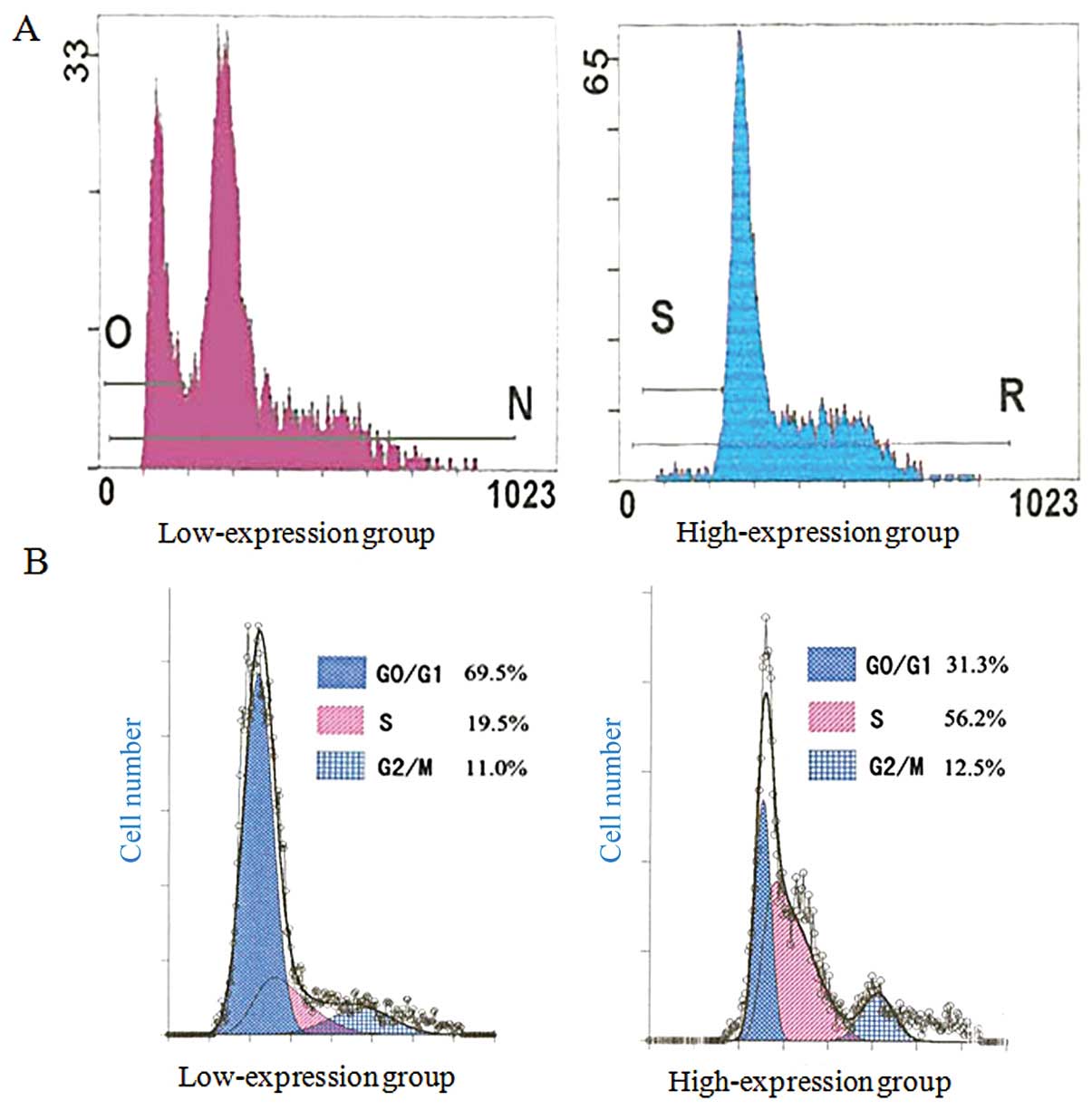
According to the results of IHC, the carcinoma cases
were divided into two groups: EphA2 high-expression group (++~+++)
and EphA2 low-expression group (−~+). The apoptosis rate was lower
in the EphA2 high-expression group than that in the low-expression
groups (P=0.018) and PI had a contrasting trend (P=0.002) as shown
by FCM (Fig. 2 and Table IV).
| Table IVApoptotic rate and proliferative
index (PI) in the gastric carcinoma cases according to EphA2
expression. |
Table IV
Apoptotic rate and proliferative
index (PI) in the gastric carcinoma cases according to EphA2
expression.
| | Apoptotic rate | PI |
|---|
| |
|
|
|---|
| EphA2 | n | Mean ± SD (%) | P-value | Mean ± SD (%) | P-value |
|---|
| −~+ | 22 | 12.27±4.35 | 0.018 | 41.83±19.44 | 0.002 |
| ++~+++ | 60 | 10.51±3.49 | | 56.72±15.91 | |
Effect of RNAi silencing of EphA2 on the
proliferation and apoptosis of human gastric carcinoma cell line
MGC803
In order to explore the precise effect of EphA2 on
the cell proliferation and apoptosis of gastric cancer cells, the
MGC803 cell line was used, which overexpressed the EphA2 protein.
In addition, RNAi was used to knock down the expression of EphA2.
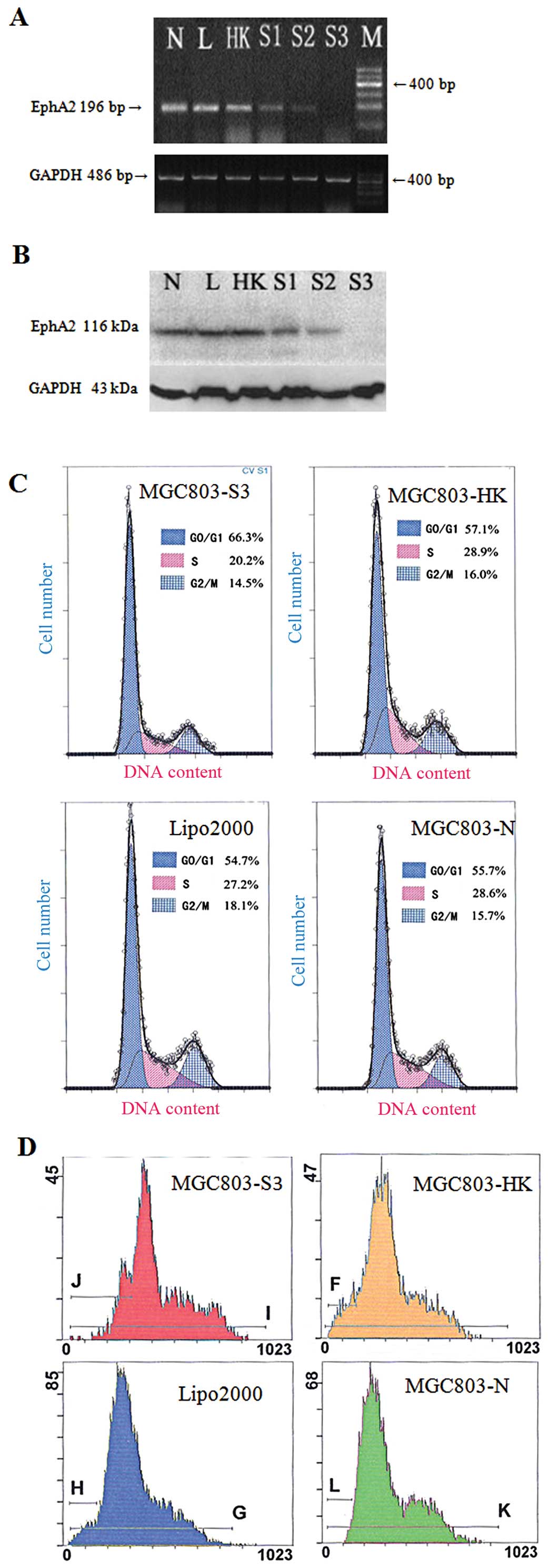
As shown in Fig. 3A and B, MGC803
cells transfected with the sh-EphA2 vector showed low EphA2 mRNA
and protein expression, particularly in the S3 vector group. On the
contrary, no change in EphA2 protein expression was found between
cells transfected with the blank control vector and the
untransfected MGC803 cells.
FCM revealed that the number of cells in the S phase
decreased and the number of cells in the
G0/G1 phase was increased. Therefore, the PI
decreased following transfection of the MGC803 cells with the
sh-EphA2 vector (Table V and
Fig. 3C). Furthermore, silencing of
EphA2 expression induced cell apoptosis (Table VI and Fig. 3D).
| Table VChanges in the cell cycle distribute
and proliferative index (PI) in the cell groups after transfection
at 48 h (FCM). |
Table V
Changes in the cell cycle distribute
and proliferative index (PI) in the cell groups after transfection
at 48 h (FCM).
| Cells |
G0/G1 | S | G2M | PI |
|---|
| MGC803-S3 | 62.70±2.39 | 22.27±2.10 | 15.03±0.55 | 37.30±2.39 |
| MGC803-HK | 57.40±0.98a | 26.10±1.47 | 16.53±0.61 | 42.60±0.98a |
| Lipo2000 | 54.30±1.73a | 29.93±3.74a | 15.77±2.35 | 45.70±1.73a |
| MGC803-N | 56.33±1.18a | 27.77±3.33a | 15.90±2.21 | 43.67±1.18a |
| Table VIApoptosis rate of the 4 cell groups
after transfection at 48 h (FCM). |
Table VI
Apoptosis rate of the 4 cell groups
after transfection at 48 h (FCM).
| Cells | Apoptosis rate
(Mean ± SD) | P-value |
|---|
| MGC803-S3 | 19.53±0.80 | 0.0001 |
| MGC803-HK | 11.05±1.34a,b | |
| Lipo2000 | 9.83±0.44a,b | |
| MGC803-N | 2.28±0.61a | |
GA inhibits cell proliferation and
induces cell apoptosis in the human gastric carcinoma cell line
MGC803 and decreases EphA2 protein expression
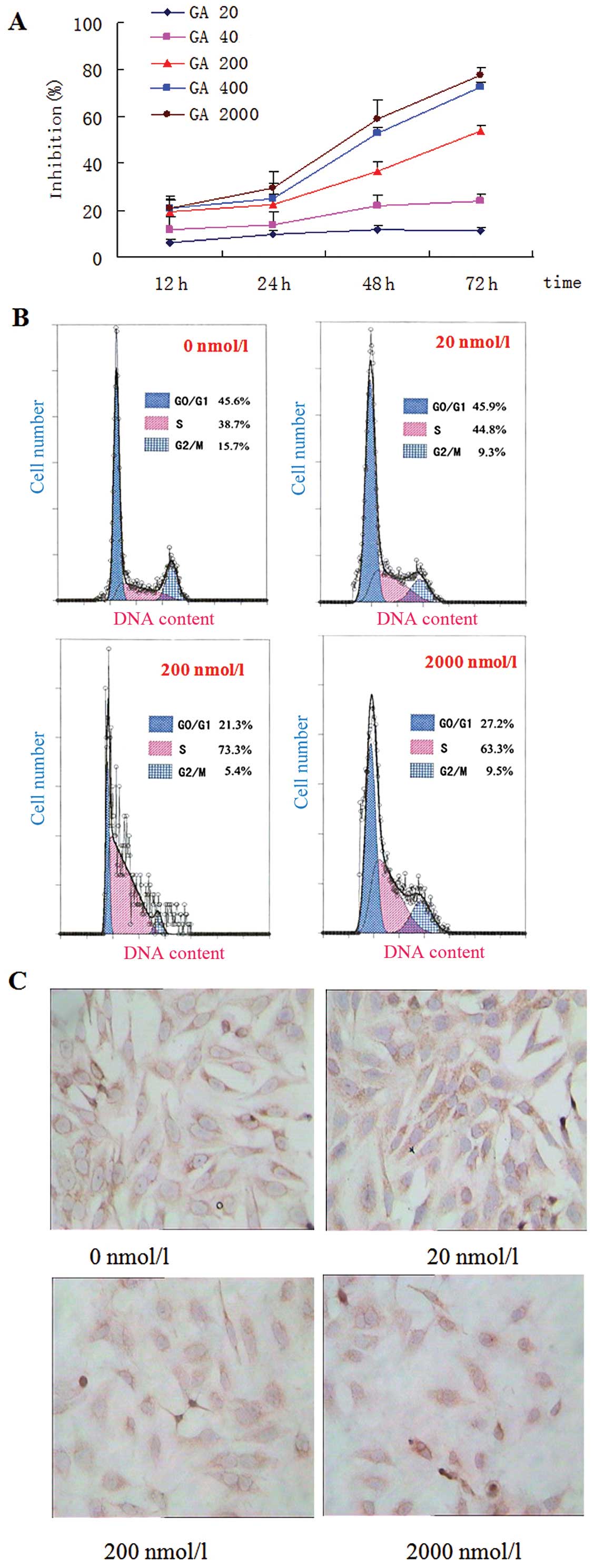
The MTT results showed that GA inhibited the
proliferation of MGC803 cells significantly in a time-dependent and
dose-dependent manner and the highest inhibition rate reached
77.69±0.91%; IC50 was 558.94 nmol/l after 48 h of GA
treatment (Fig. 4A).
FCM analysis showed that the percentage of cells in
the S phase was increased while the percentage of cells in the
G2/M phase was decreased in the GA-stimulated cells
compared with the control group, and the percentage of cells in the
S phase was 36.35±3.04% in the control group, 45.00±1.41% in the
middle-dose group, and 45.25±3.04% in the high-dose group,
respectively. At same time, the percentage of cells in the
G0/G1 phase increased (Fig. 4B and Table VII). As shown in Table VIII, GA induced cell apoptosis of
MGC803 cells in a dose-dependent manner (P<0.01). As shown in
Fig. 4C, GA treatment inhibited the
expression of EphA2 protein.
| Table VIIEffect of geldanamycin (GA) on MGC803
cell cycle distribution as assayed by FCM. |
Table VII
Effect of geldanamycin (GA) on MGC803
cell cycle distribution as assayed by FCM.
| Treatment
groups | Concentration
(nmol/l) |
G0/G1 | S |
G2/M |
|---|
| A (control) | 0 | 34.00±1.56 | 36.35±3.04 | 29.65±4.60 |
| B | 20 | 39.85±3.89 | 36.65±2.19 | 23.50±1.70 |
| C | 200 | 42.50±2.12 | 45.00±1.41a | 12.57±1.71a |
| D | 2,000 | 49.20±4.81a | 45.25±3.04a | 5.55±2.19a |
| Table VIIIInfluence of geldanamycin (GA) on the
apoptosis of MGC-803 cells (FCM). |
Table VIII
Influence of geldanamycin (GA) on the
apoptosis of MGC-803 cells (FCM).
| Treatment
groups | Concentration
(nmol/l) | Apoptotic rate
(%) | P-value |
|---|
| A (control) | 0 | 1.76±0.36 | 0.002 |
| B | 20 | 5.94±1.25a | |
| C | 200 | 15.44±6.48b | |
| D | 2,000 | 26.91±3.30b | |
In order to investigate whether GA induces MGC803
cell apoptosis through downregulation of EphA2 protein, transient
transfection experiments were performed. Compared with the control
group, pcDNA3.1-EphA2-IRES-EGFP effectively upregulated the EphA2
protein and mRNA levels in the MGC803 cells by 2.3 and 2.5-fold. As
detected by FCM, GA increased the apoptosis ratio of MGC803 cells;
however, the apoptosis ratio in the GA+EphA2 vector group was
decreased compared with the GA group. Importantly, knockdown of
EphA2 expression increased the apoptosis ratio induced by GA in the
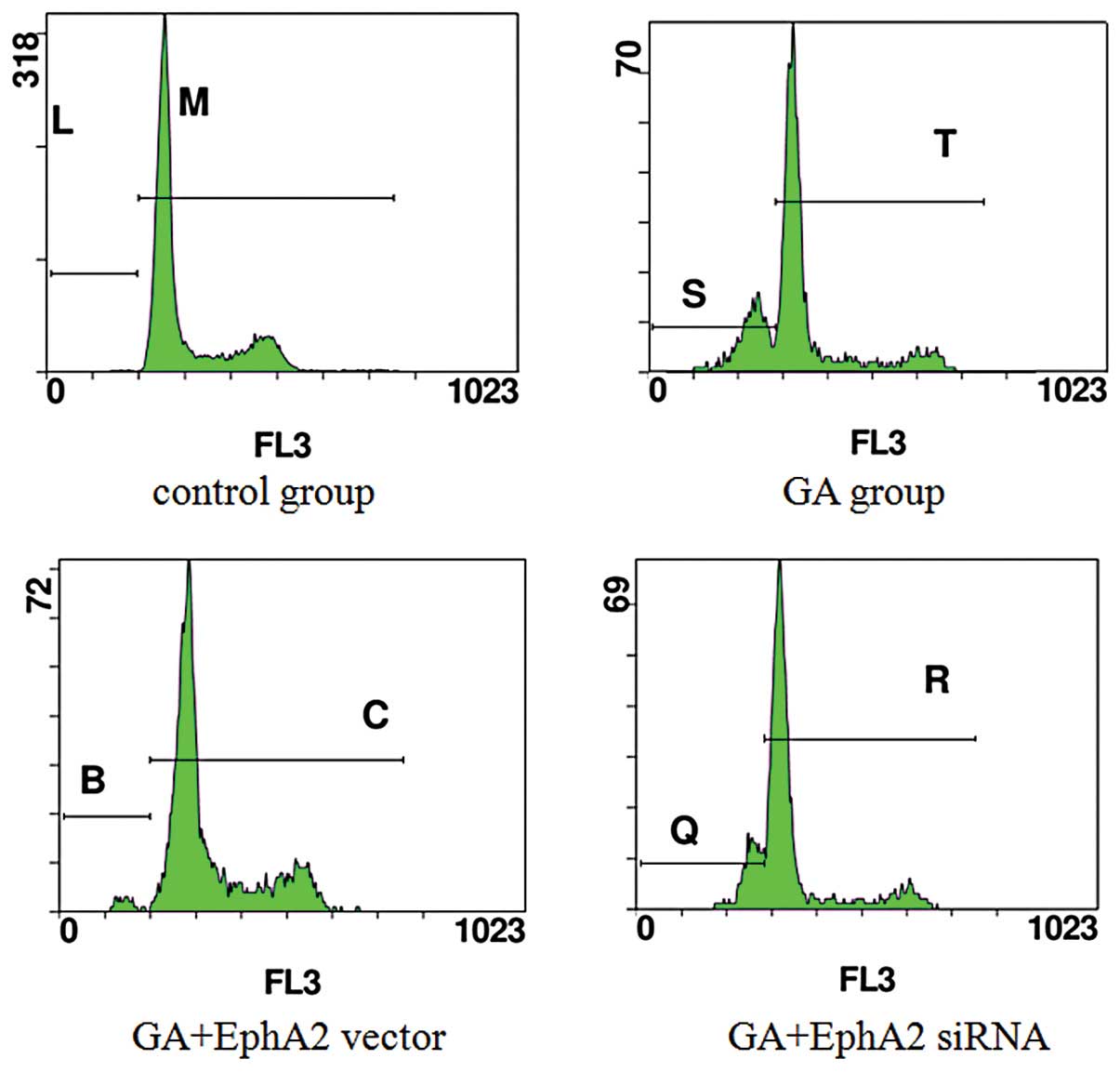
MGC803 cells (Fig. 5).
Discussion
EphA2 is overexpressed in a variety of human
malignancies, such as breast, colon, ovarian and pancreatic
(9–13). EphA2 mediates angiogenesis, tumor
growth, invasion and metastasis (9,14–16)
and its expression level is correlated with patient prognosis
(17–21). Although the kinase domain is
necessary for EphA2-driven cell migration and tumorigenicity
(22), the outcome of
ligand-dependent receptor phosphorylation is controversial.
Furthermore, the activation of oncogenic signaling pathways further
disrupts the delicate balance of EphA2 bi-directional signaling
(23). Our results showed that
EphA2 protein expression was increased in the gastric carcinoma
tissues and was positively correlated with the histologic degree of
differentiation. In addition, apoptosis rate was higher in the
EphA2 high-expression group than that in the low-expression group
while PI had an inverse trend, which suggests that EphA2 plays an
important role in angiogenesis of gastric carcinoma, especially
related with abnormal growth of gastric carcinoma cells.
RNAi has become an important method in tumor
research (24). In order to further
investigate whether EphA2 is involved in the cell proliferation or
cell apoptosis of gastric cancer cells, RNAi method was used to
knock down the expression of EphA2. Moreover, EphA2 knockdown
inhibited MGC803 cell growth, and blocked cells in the
G0/G1 stage, inducing apoptosis, suggesting
that silencing of EphA2 inhibits tumorigenesis and development
possibly through cell cycle arrest and induction of cell
apoptosis.
GA is a benzoquinone ansamycin antibiotic and a
potent tyrosine kinase inhibitor that was first isolated from
Streptomyces hygrocopicus in 1970. GA is also known as a
potent inhibitor of Hsp90 (25).
Research has shown that GA inhibits cell growth in many types of
cancers (26,27). However, it is unknown whether or not
GA inhibits cell growth of gastric cancer. In the present study, we
found that GA inhibited the cell proliferation of MGC803 cells,
arrested the cell cycle and induced cell apoptosis in a dose- and
time-dependent manner, which suggests that GA has an anticancer
effect on gastric cancer.
In order to investigate whether GA inhibited cell
growth through inhibition of EphA2 expression, we detected the
effect of EphA2 by GA and analyzed the correlation between EphA2
expression and apoptosis. The results showed that GA decreased the
expression of EphA2 protein and EphA2 expression was negatively
correlated with cell apoptosis. Overexpression of EphA2 inhibited
cell apoptosis induced by GA and knockdown of EphA2 protein
increased the apoptosis rate induced by GA in MGC803 cells. From
the above results, we believe that GA inhibits cell proliferation
and induces apoptosis partly through inhibition of EphA2 protein
expression.
In summary, our data demonstrated that EphA2
over-expression plays an important role in the carcinogenesis of
gastric epithelial cells, with a decrease in apoptosis and cell
cycle arrest. Knockdown of EphA2 blocked MGC803 cell proliferation
and inducd cell apoptosis. GA inhibited MGC803 cell proliferation
and induced cell apoptosis by upregulating expression of EphA2.
These findings suggest that EphA2 may be a new target for
preventing gastric carcinogenesis and treating gastric carcinoma.
However, further studies are needed to examine the precise role of
EphA2 in the pathogenesis of gastric cancer.
Acknowledgements
This research was supported by the Hebei Natural
Science Foundation (H2013206446).
References
|
1
|
Nakamoto M and Bergemann AD: Diverse roles
for Eph family of receptor tyrosine kinases in carcinogenesis.
Microsc Res Tech. 59:58–67. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fang WB, Brantley-Sieders DM, Parker MA,
Reith AD and Chen J: A kinase-dependent role for EphA2 receptor in
promoting tumor growth and metastasis. Oncogene. 24:7859–7868.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Miyazaki T, Kato H, Fukuchi M, Nakajima M
and Kuwano H: EphA2 overexpression correlates with poor prognosis
in esophageal squamous cell carcinoma. Int J Cancer. 103:657–663.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Orsulic S and Kemler R: Expression of Eph
receptors and ephrins is differentially regulated by E-cadherin. J
Cell Sci. 113:1793–1802. 2000.PubMed/NCBI
|
|
5
|
Trojandt S, Reske-Kunz AB and Bros M:
Geldanamycin-mediated inhibition of heat shock protein 90 partially
activates dendritic cells, but interferes with their full
maturation, accompanied by impaired upregulation of RelB. J Exp
Clin Cancer Res. 33:162014. View Article : Google Scholar
|
|
6
|
Zeng G, Hu Z, Kinch MS, Pan CX, Flockhart
DA, Kao C, Gardner TA, Zhang S, Li L, Baldridge LA, Koch MO,
Ulbright TM, Eble JN and Cheng L: High-level expression of EphA2
receptor tyrosine kinase in prostatic intraepithelial neoplasia. Am
J Patho1. 163:2271–2276. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Feng X, Hao J, Liu Q, Yang L, Lv X, Zhang
Y, Xing L, Xu N and Liu S: HMGB1 mediates IFN-γ-induced cell
proliferation in MMC cells through regulation of cyclin D1/CDK4/p16
pathway. J Cell Biochem. 113:2009–2019. 2012.
|
|
8
|
Hao J, Zhang YJ, Lv X, Xu N, Liu QJ, Zhao
S, Feng XJ, Xing LL, Kang PP, Li GY and Liu SX: IFN-γ induces
lipogenesis in mouse mesangial cells via the JAK2/STAT1 pathway. Am
J Physiol Cell Physiol. 304:c760–c767. 2013.
|
|
9
|
Dodelet VC and Pasquale EB: Eph receptors
and ephrin ligands: embryogenesis to tumorigenesis. Oncogene.
19:5614–5619. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Duxbury MS, Ito H, Zinner MJ, Ashley SW
and Whang EE: Ligation of EphA2 by Ephrin A1-Fc inhibits pancreatic
adenocarcinoma cellular invasiveness. Biochem Biophys Res Commun.
320:1096–1102. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Thaker PH, Deavers M, Celestino J,
Thornton A, Fletcher MS, Landen CN, Kinch MS, Kiener PA and Sood
AK: EphA2 expression is associated with aggressive features in
ovarian carcinoma. Clin Cancer Res. 10:5145–5150. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wykosky J, Gibo DM, Stanton C and Debinski
W: EphA2 as a novel molecular marker and target in glioblastoma
multiforme. Mol Cancer Res. 3:541–551. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu F, Park PJ, Lai W, Maher E,
Chakravarti A, Durso L, Jiang X, Yu Y, Brosius A, Thomas M, Chin L,
Brennan C, DePinho RA, Kohane I, Carroll RS, Black PM and Johnson
MD: A genome-wide screen reveals functional gene clusters in the
cancer genome and identifies EphA2 as a mitogen in glioblastoma.
Cancer Res. 66:10815–10823. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zelinski DP, Zantek ND, Stewart JC,
Irizarry AR and Kinch MS: EphA2 overexpression causes tumorigenesis
of mammary epithelial cells. Cancer Res. 61:2301–2306.
2001.PubMed/NCBI
|
|
15
|
Hess AR, Seftor EA, Gruman LM, Kinch MS,
Seftor RE and Hendrix MJ: VE-cadherin regulates EphA2 in aggressive
melanoma cells through a novel signaling pathway: implications for
vasculogenic mimicry. Cancer Biol Ther. 5:228–233. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Brantley-Sieders DM, Zhuang G, Hicks D,
Fang WB, Hwang Y, Cates JM, Coffman K, Jackson D, Bruckheimer E,
Muraoka-Cook RS and Chen J: The receptor tyrosine kinase EphA2
promotes mammary adenocarcinoma tumorigenesis and metastatic
progression in mice by amplifying ErbB2 signaling. J Clin Invest.
118:64–78. 2008. View
Article : Google Scholar
|
|
17
|
Fox BP and Kandpal RP: Invasiveness of
breast carcinoma cells and transcript profile: Eph receptors and
ephrin ligands as molecular markers of potential diagnostic and
prognostic application. Biochem Biophys Res Commun. 318:882–892.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Herrem CJ, Tatsumi T, Olson KS, Shirai K,
Finke JH, Bukowski RM, Zhou M, Richmond AL, Derweesh I, Kinch MS
and Storkus WJ: Expression of EphA2 is prognostic of disease-free
interval and overall survival in surgically treated patients with
renal cell carcinoma. Clin Cancer Res. 11:226–231. 2005.PubMed/NCBI
|
|
19
|
Merritt WM, Thaker PH, Landen CN Jr,
Deavers MT, Fletcher MS, Lin YG, Han LY, Kamat AA, Schmandt R,
Gershenson DM, Kinch MS and Sood AK: Analysis of EphA2 expression
and mutant p53 in ovarian carcinoma. Cancer Biol Ther. 5:1357–1360.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Abraham S, Knapp DW, Cheng L, Snyder PW,
Mittal SK, Bangari DS, Kinch M, Wu L, Dhariwal J and Mohammed SI:
Expression of EphA2 and Ephrin A-1 in carcinoma of the urinary
bladder. Clin Cancer Res. 12:353–360. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Larsen AB, Pedersen MW, Stockhausen MT,
Grandal MV, van Deurs B and Poulsen HS: Activation of the EGFR gene
target EphA2 inhibits epidermal growth factor-induced cancer cell
motility. Mol Cancer Res. 5:283–293. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fang WB, Brantley-Sieders DM, Parker MA,
Reith AD and Chen J: A kinase-dependent role for EphA2 receptor in
promoting tumor growth and metastasis. Oncogene. 24:7859–7868.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Carles-Kinch K, Kilpatrick KE, Stewart JC
and Kinch MS: Antibody targeting of the EphA2 tyrosine kinase
inhibits malignant cell behavior. Cancer Res. 62:2840–2847.
2002.PubMed/NCBI
|
|
24
|
Zantek ND, Azimi M, Fedor-Chaiken M, Wang
B, Brackenbury R and Kinch MS: E-cadherin regulates the function of
the EphA2 receptor tyrosine kinase. Cell Growth Differ. 10:629–638.
1999.PubMed/NCBI
|
|
25
|
Macrae M, Neve RM, Rodriguez-Viciana P,
Haqq C, Yeh J, Chen C, Gray JW and McCormick F: A conditional
feedback loop regulates Ras activity through EphA2. Cancer Cell.
8:111–118. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ui T, Morishima K, Saito S, Sakuma Y,
Fujii H, Hosoya Y, Ishikawa S, Aburatani H, Fukayama M, Niki T and
Yasuda Y: The HSP90 inhibitor 17-N-allylamino-17-demethoxy
geldanamycin (17-AAG) synergizes with cisplatin and induces
apoptosis in cisplatin-resistant esophageal squamous cell carcinoma
cell lines via the Akt/XIAP pathway. Oncol Rep. 31:619–24.
2014.PubMed/NCBI
|
|
27
|
Jilani K, Qadri SM and Lang F:
Geldanamycin-induced phosphatidylserine translocation in the
erythrocyte membrane. Cell Physiol Biochem. 32:1600–1609.
2013.PubMed/NCBI
|