Introduction
Chelidonium majus is commonly known as
greater celandine. It belongs to the family Papaveraceae and is
widely distributed in Europe and western Asia. Extracts of
Chelidonium majus L. (C. majus L.) have been shown to
exhibit a variety of biological effects including
anti-inflammatory, antimicrobial, antiviral and antitumor
activities (1,2). The antitumor, antigenotoxic and
hepatoprotective effects of C. majus L. extract suggest that
it may be potentially useful as an anticancer therapeutic agent
(3).
Apoptosis is mediated through at least 3 major
pathways, which are regulated by death receptors, mitochondria and
the endoplasmic reticulum. Activation of the apoptosis pathway is a
key mechanism by which cytotoxic drugs kill tumor cells. Defects in
apoptosis signaling contribute to the drug resistance of tumor
cells (4,5). The Bcl-2 family consists of important
apoptotic regulators of programmed cell death. This family of
proteins includes both anti-apoptotic molecules such as Bcl-2 and
pro-apoptotic molecules such as Bax (6). Bcl-2 and Bcl-XL are members
of the Bcl-2 family and are regulated by nuclear factor-(NF)-κB.
These proteins can prevent release of cytochrome c and
activation of caspases (7).
Mitogen-activated protein kinase (MAPK) cascades are
involved in the signaling pathways that regulate various cellular
responses such as inflammation, proliferation and cell death
(8). Three major mammalian MAPK
subfamilies have been described: extracellular signal-regulated
kinases (ERKs), c-Jun N-terminal kinases and p38 kinases.
Each MAPK is activated through a specific phosphorylation cascade
(9,10). The ERK cascade is activated through
receptor-mediated signaling stimuli and is associated with cell
proliferation, differentiation and survival (11). However, in some cases, ERK
activation contributes to cell death (12).
In the present study, we examined how the effects of
C. majus L. extract on human epidermoid carcinoma A431 cells
are mediated through multiple mechanisms, including activation of
cell cycle arrest and the caspase-dependent pathway, blocking of
NF-κB activation and mediation of the MAPK pathway in human
epidermoid carcinoma A431 cells.
Materials and methods
Preparation of C. majus L. extract
Five hundred grams of C. majus L. and 3 l of
distilled water were placed into a round-bottomed flask fitted with
a condenser and a heating mantle. The contents of the flask were
then boiled for 2 h. The resulting water extract was filtered
through a Whatman no. 1 filter paper and its volume was reduced
using a rotary evaporator (Buchi, Flawil, Switzerland). The
concentrated extract was freeze-dried (EYELA, Tokyo, Japan) and
stored at 4°C in a vacuum container until it was used. The final
weight of the extract was 100 g, of which 20% was natural
product.
Cell culture
The human epidermoid carcinoma A431 cell line was
purchased from the American Tissue Culture Collection (Manassas,
VA, USA). The cells were cultured in Dulbecco’s modified
Eagle’s medium (Gibco, Grand Island, NY, USA), which
contained 10% fetal bovine serum, 100 U/ml penicillin and 100 μg/ml
streptomycin at 37°C in a humidified atmosphere of 5%
CO2 and 95% air.
MTT assay
Cell proliferation was evaluated using the CellTiter
96 Aqueous One Solution (Promega, Madison, WI, USA). Cells
(1×104) were incubated with several concentrations of
C. majus L. extract (0–500 μg/ml) at 37°C for 24, 48 and 72
h. Cell viability was determined through a colorimetric assay by
using PMS/MTS solution. The absorbance was determined at 492 nm
with background subtraction at 650 nm.
Cell cycle analysis
Cells (5×105) were treated with C.
majus L. extracts for 72 h. At the end of the treatment period,
the cells were harvested and washed with PBS. They were then fixed
with 70% ethanol for 1 h, treated with RNase A (20 μg/ml) at 37°C
for 1 h and stained with propidium iodide (50 μg/ml). The DNA
content at each cell cycle stage was analyzed using a FACSCalibur
flow cytometer and CellQuest software (Becton Dickinson, Franklin
Lakes, NJ, USA).
Apoptosis assay
The number of apoptotic A431 cells was determined
using the Cell Death Detection ELISAplus kit (Roche
Molecular Biochemicals, Mannheim, Germany). Cells
(1×104) were incubated with several concentrations of
C. majus L. extract for 72 h. They were then lysed with the
cell lysis buffer (200 μl). The cell lysates were assayed for DNA
fragments by using the Cell Death Detection ELISAplus
kit according to the manufacturer’s protocol. DNA fragmentation was
evaluated at 405 nm against an untreated control.
Caspase inhibitor assay
Cells (1×104) were pre-incubated with
several concentrations of the pan-caspase inhibitor Z-VAD-FMK
(R&D Systems, Minneapolis, MN, USA) or the caspase-3-specific
inhibitor Z-DEVD-FMK for 2 h. Then, 500 μg/ml C. majus L.
extract was added to the cells and the cells were cultured for a
further 72 h. Cell viability was determined through a colorimetric
assay using the PMS/MTS solution. The absorbance was determined at
492 nm with background subtraction at 650 nm.
RNA extraction and real-time PCR
Total RNA was purified from cultured cells by using
an RNA-Bee solution kit following the manufacturer’s protocol
(Tel-Test, Friendswood, TX, USA). First-strand cDNA synthesis was
performed with 1 μg of total RNA and transcribed into cDNA using a
reverse transcription system with random hexamers according to the
manufacturer’s protocol. The primer sequences used were as
follows: cyclin D1 (5′-CCGTCCATGCGGAAGATC-3′, 5′-ATGGCCAGCG
GGAAGAC-3′; 86 bp), p21 (5′-CAGACCAGCATGACAGAT TTC-3′,
5′-TTAGGGCTTCCTCTTGGAGA-3′; 66 bp), Bcl-2
(5′-GATTGATGGGATCGTTGCCTTA-3′, 5′-CCTTGGCAT GAGATGCAGGA-3′; 200
bp), Bax (5′-GGATGCGTCCACC AAGAAG-3′, 5′-GCCTTGAGCACCAGTTTGC-3′;
216 bp), Mcl-1 (5′-CTCATTTCTTTTGGTGCCTTT-3′, 5′-CCAGTCC
CGTTTTGTCCTTAC-3′; 117 bp), survivin (5′-GGCCCA GTGTTTCTTCTGCTT-3′,
5′-GCAACCGGACGAATGC TTT-3′; 91 bp), β-actin
(5′-GCGAGAAGATGACCCAGATC-3′, 5′-GGATAGCACAGCCTGGATAG-3′; 77 bp).
Real-time PCR was performed using a StepOnePlus Real-Time PCR
system with the Power SYBR Green PCR Master Mix (Applied
Biosystems, Foster, CA, USA). The PCRs were performed with 1 μl of
cDNA in 20 μl reaction mixtures that consisted of 10 μl Power
SYBR-Green PCR Master Mix, 2 μl of primers and 7 μl of PCR-grade
water. The reactions were performed with a denaturation step at
95°C for 10 min, followed by 40 cycles at 95°C for 15 sec and at
60°C for 1 min. The crossing point of each target gene with β-actin
was calculated by using the formula 2−(target gene −
β-actin) and the relative amounts of the PCR products were
quantified.
Immunoblot analysis
Cells (2×106) were treated with various
concentrations of C. majus L. for 72 h. After treatment, the
cells were washed with cold PBS and lysed with the lysis buffer [20
mM Tris-HCl (pH 7.5), 150 mM NaCl, 1 mM Na2EDTA, 1 mM
EGTA, 1% Triton, 2.5 mM sodium pyrophosphate, 1 mM
β-glycerophosphate, 1 mM Na3VO4 and 1 μg/ml
leupeptin] containing 1 mM PMSF. The protein concentration was
determined by means of the BCA protein assay according to the
manufacturer’s protocol. Thirty micrograms of protein was
fractionated by performing SDS-PAGE on a 12% gel and then
transferred through electrophoresis onto nitrocellulose membranes.
The membranes were blocked with 5% nonfat dry milk for 1 h at room
temperature and then incubated overnight with anti-NF-κB p65,
anti-p38 MAPK, anti-phospho-p38 MAPK, anti-ERK, anti-phospho-ERK,
anti-MEK, anti-phospho-MEK (Cell Signaling Technology, Danvers, MA,
USA) and anti-β-actin antibodies (Sigma-Aldrich) diluted to 1:1,000
with Tris-buffered saline containing 0.05% Tween-20 (TBS-T). After
washing with TBS-T for 1 h, the membranes were treated for 1 h at
room temperature with horseradish peroxidase-conjugated secondary
antibody diluted to 1:2,500 with TBS-T. Then, the membranes were
washed with TBS-T for 1 h and the proteins were detected using an
enhanced chemiluminescence kit (Santa Cruz Biotechnology, Santa
Cruz, CA, USA). Protein expression was analyzed using a
Chemiluminescence Imaging System (Davinch-Chemi™, Seoul,
Korea).
Statistical analysis
The values are expressed as the mean ± SD.
Student’s t-test was used to evaluate differences between
the control and C. majus L. extract-treated samples. The
effect of the caspase inhibitors on cell viability was estimated by
calculating the differences between the C. majus L.
extract-treated samples and the C. majus L. extract/caspase
inhibitor-treated samples. *p<0.05 and
**p<0.01 were considered to indicate statistically
significant results.
Results
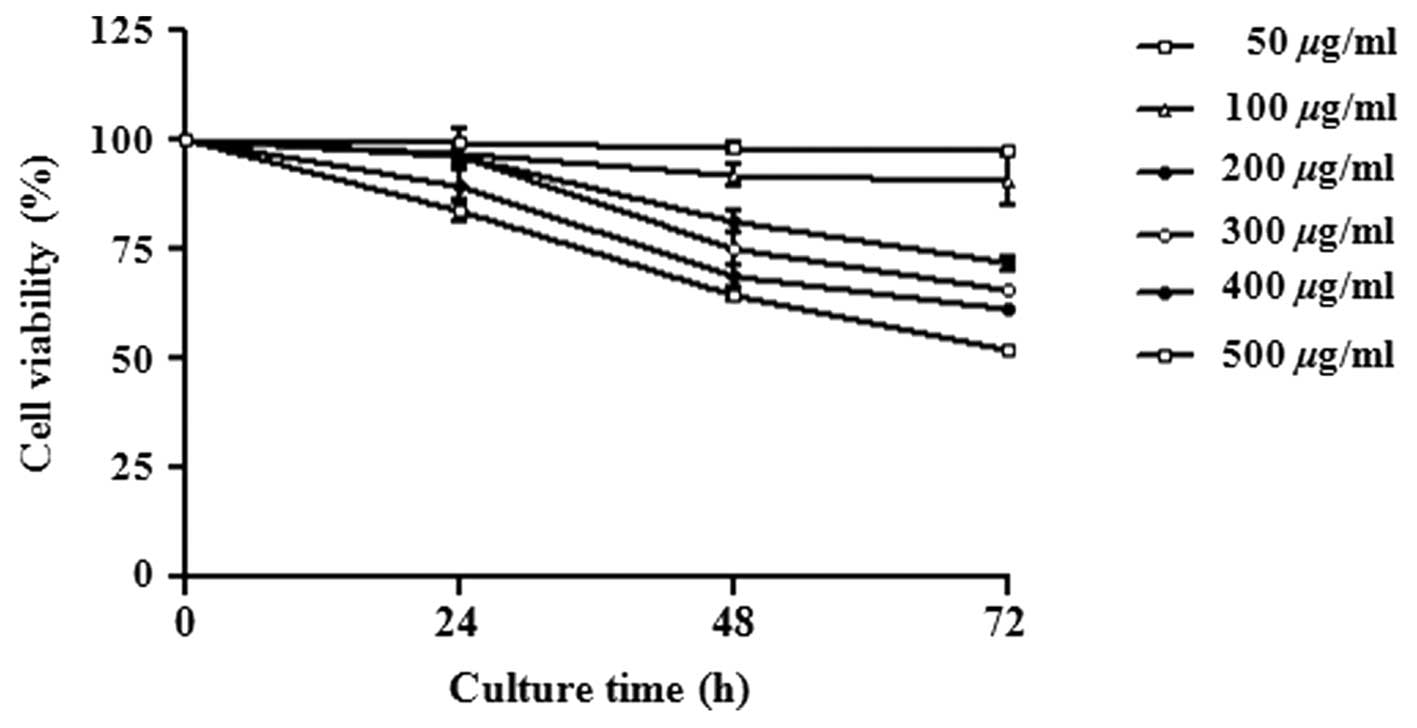
C. majus L. extract inhibits cell
proliferation
A431 cells were treated with various concentrations
of C. majus L. extract (0–500 μg/ml) for 24, 48 and 72 h,
respectively. The effect of C. majus L. extract on cell
proliferation was evaluated through an MTT assay. C. majus
L. extract inhibited proliferation of the cells in a dose- and
time-dependent manner (Fig. 1).
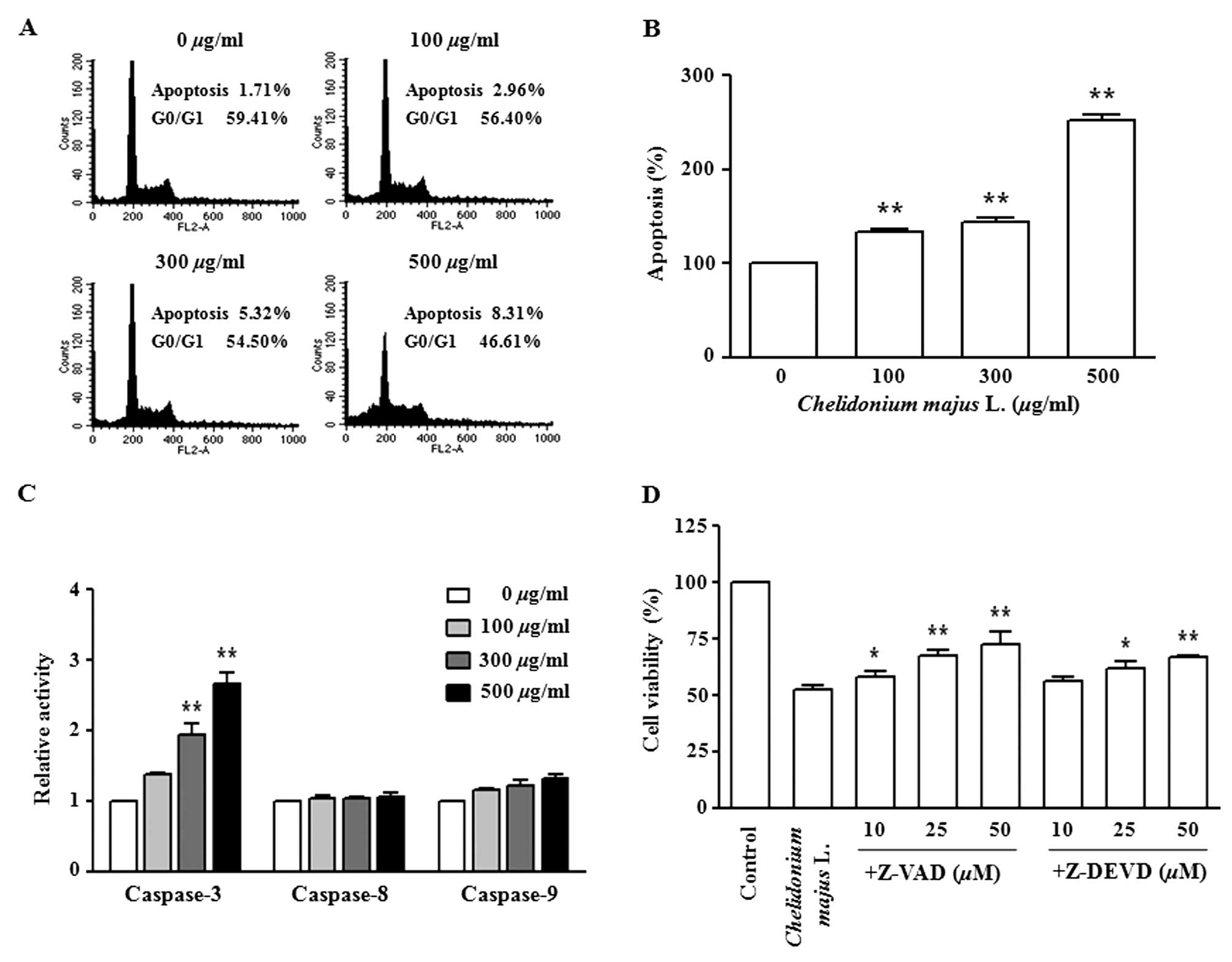
C. majus L. extract induces
apoptosis
A431 cells were treated with various concentrations
of C. majus L. extract for 72 h. Flow cytometric analysis
showed a dose-dependent increase in the number of cells in the
apoptosis phase and a decrease in the number of cells in the G0/G1
phase of the cell cycle (Fig. 2A).
Apoptotic cells were then detected using a Cell Death Detection
ELISA (Fig. 2B). The number of
apoptotic cells increased in a dose-dependent manner following
treatment with C. majus L. extract at various
concentrations. Caspase activity was assayed using a colorimetric
ELISA. Caspase-3 and caspase-9 activities increased in a
dose-dependent manner following treatment with C. majus L.
extract (Fig. 2C). To confirm
whether caspase activation was involved in C. majus L.
extract-induced apoptosis, MTT assay was performed to determine
A431 cell proliferation due to treatment with C. majus
L. extract. Pretreatment of the cells with the pan-caspase
inhibitor Z-VAD-FMK and the caspase-3 inhibitor Z-DEVD-FMK
increased the C. majus L. extract-induced cell proliferation
(Fig. 2D).
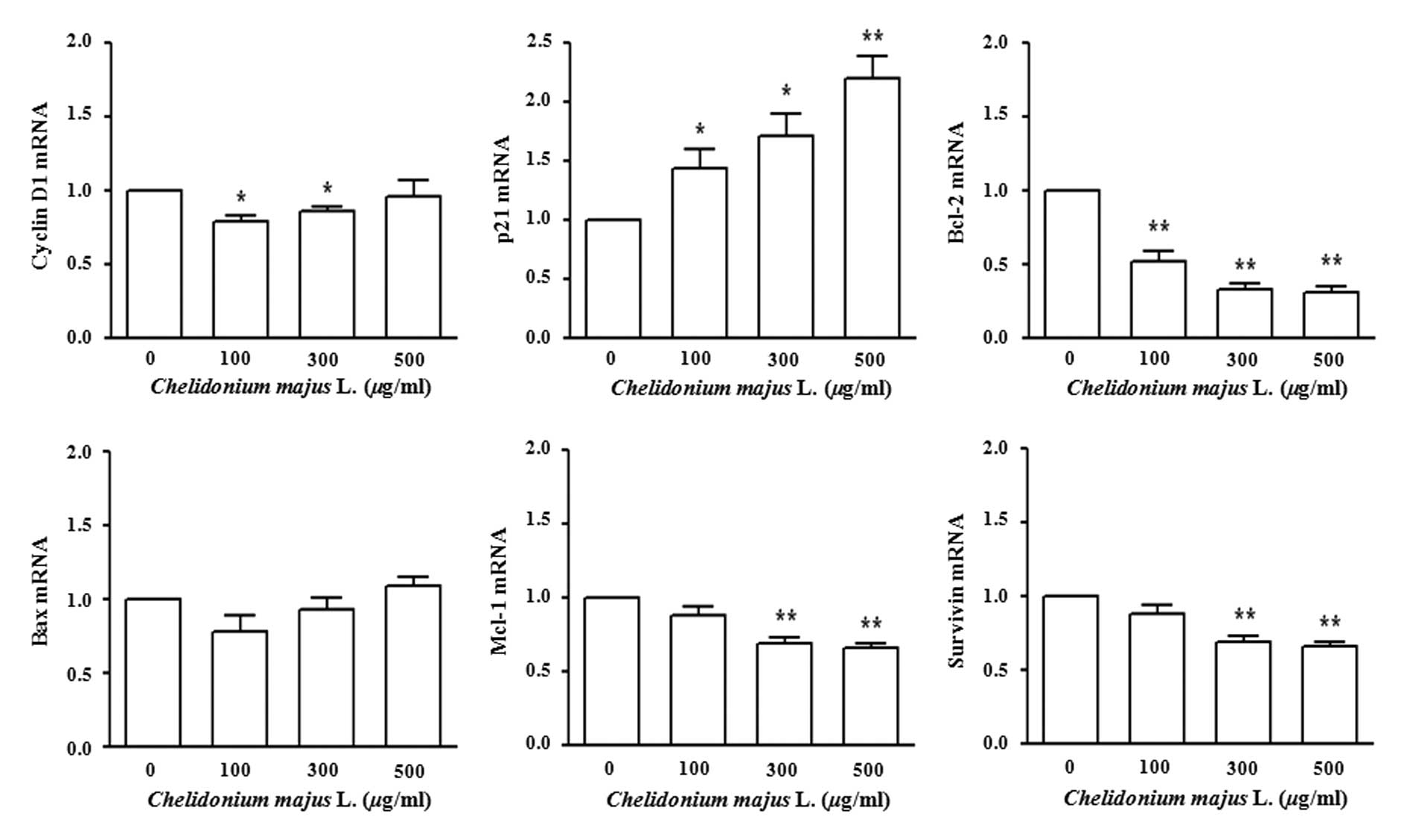
C. majus L. extract regulates mRNA
expression
A431 cells were treated with various concentrations
of C. majus L. extract for 72 h. Then, the mRNA levels of
the cell cycle-related genes cyclin D and p21 and the
apoptosis-related genes Bcl-2, Mcl-1, Bax and survivin were
examined using real-time PCR. The mRNA levels of cyclin D1, Bcl-2,
Mcl-1 and survivin decreased, while those of p21 and Bax increased
in a dose-dependent manner (Fig.
3).
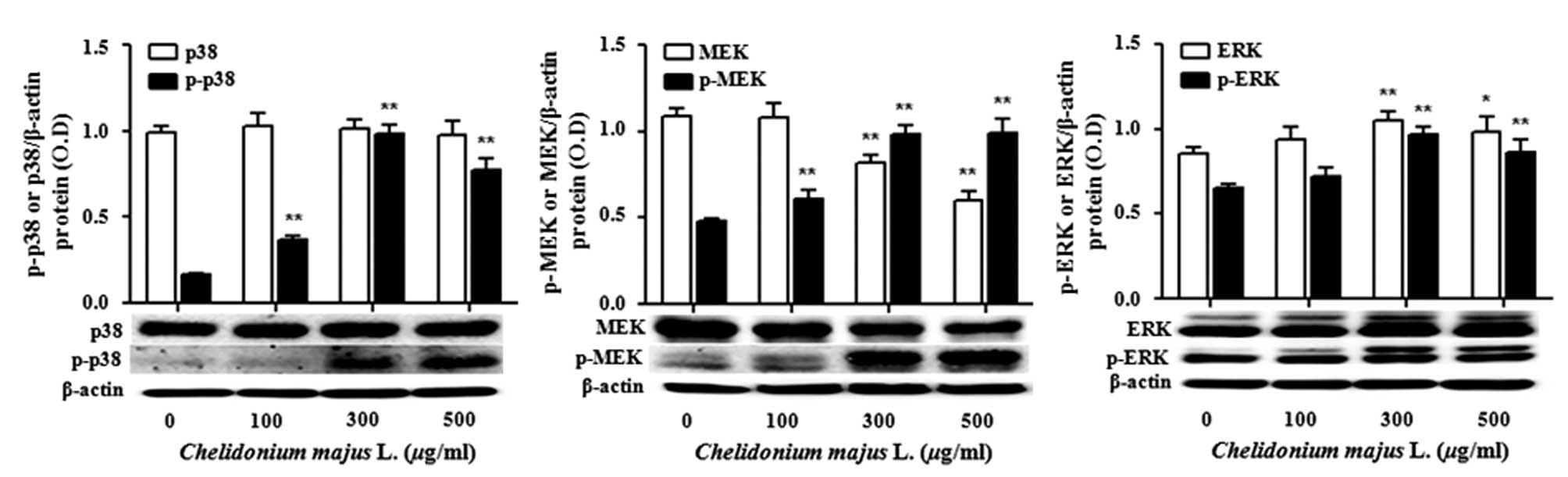
C. majus L. extract inhibits NF-κB and
increases the activities of p38 MAPK and MEK/ERK
A431 cells were treated with various concentrations
of C. majus L. extract for 72 h. Changes in the activation
and phosphorylation levels of key proteins in the cells were
determined by performing western blot analysis. The levels of NF-κB
p65 were significantly inhibited (Fig.
4). However, high concentrations (300 and 500 μg/ml) of C.
majus L. extract induced activities in the levels of
phosphorylated p38 MAPK and MEK/ERK1/2 (Fig. 5).
Discussion
The cytotoxic activity of C. majus L.
on human and animal tumor cell cultures in vitro are of
great interest and can be considered promising for use in cancer
therapy (13). We investigated the
effect of C. majus L. extract on the viability of A431
cells. C. majus L. extract inhibited A431 cell
proliferation and this result was consistent with those of Chor
et al (14), who reported on
the activities of several herbs. Our results showed that treatment
with C. majus L. extract increased the percentage of cells
in the apoptotic phase of the cell cycle and enhanced apoptotic
death of A431 cells. Taken together, these data suggest that C.
majus L. extract not only inhibits A431 cell growth and blocks
cell cycle progression at the G1 phase, but also induces apoptosis.
A recent study reported a decrease in the number of human hepatic
stellate LX2 cells in the G1 phase and accumulation of cells in the
sub-G0/G1 phase after incubation with Orostachys japonicas
(15).
To investigate the molecular mechanism that
underlies the apoptosis of A431 cells by C. majus L., we
assessed the caspase activity of A431 cells during treatment with
C. majus L. extract. Caspases are central mediators in
apoptosis. We found that treatment of A431 cells with C.
majus L. extract increased their intracellular caspase-3 and
caspase-9 activities. This finding was confirmed by experiments
using the pan-caspase inhibitor Z-VAD-FMK and the caspase
3-specific inhibitor Z-DEVD-FMK, which enhanced C. majus L.
extract-mediated cell proliferation. These results suggest that
C. majus L. extract-induced apoptosis of A431 cells was
mediated by caspase activation. The inhibitors of caspase-3 and
caspase-9 were previously found to almost completely suppress
Houttuynia cordata Thunb-induced activities of caspase-3 and
caspase-9 in human colon adenocarcinoma HT-29 cells (16). The activities of caspase-3 and
caspase-9 and release of mitochondrial cytochrome c
increased in ovarian cancer cell lines with increasing doses of
Antrodia camphorate (17).
We investigated the mRNA levels of pro-apoptotic and
anti-apoptotic genes in A431 cells that were treated with C.
majus L. and detected enhanced expression of pro-apoptotic Bax
and reduced expression of anti-apoptotic Bcl-2. In addition, the
RNA levels of cyclin D1, Mcl-1 and survivin decreased, while that
of p21 increased. These results suggest that C. majus L.
regulated transcription factors of cell cycle-related genes and
apoptosis-related genes. Tanshinone IIA was found to exert
significant anti-proliferation effects on THP-1 cells through
induction of apoptosis. Tanshinone IIA-induced apoptosis of THP-1
cells was found to be mainly related to activation of caspase-3,
downregulation of anti-apoptotic proteins Bcl-2 and survivin and
upregulation of pro-apoptotic protein Bax (18). Shikonin was previously found to
cause cell apoptosis by upregulating p27 and Bax and downregulating
Bcl-2 and Bcl-XL in human colorectal carcinoma COLO 205
cells (19). Wogonin-induced
apoptosis was found to be accompanied by significant decreases in
Bcl-2 and survivin levels and an increase in Bax levels. In
addition, wogonin treatment was found to significantly increase the
active apoptotic forms of caspase-3, -8 and -9. Z-DEVD-FMK, which
is a specific caspase-3 inhibitor, significantly inhibited
wogonin-induced apoptosis of MCF-7 human breast cancer cells
(20).
In the present study, we found that C. majus
L. extract inhibited NF-κB activity in A431 cells. C. majus
L. extract induced cell apoptosis by inhibiting NF-κB via
downregulation of the Bcl-2 gene. NF-κB is a common transcription
factor that can modulate the expression of apoptosis-related
proteins (21). Our results suggest
that C. majus L. extract functions as an effective apoptosis
inducer through an NF-κB-dependent mechanism. Thus, suppression of
the NF-κB pathway should be effective at inducing apoptosis of A431
cells. Triptolide was found to induce the apoptosis of human
anaplastic thyroid carcinoma cells by downregulating NF-κB
expression. It also downregulated the anti-apoptotic proteins Bcl-2
and Bcl-XL, which are transcriptionally regulated by the
NF-κB signaling pathway (22). In
the present study, treatment with C. majus L. extract led to
inhibition of the NF-κB pathway. Thus, suppression of the NF-κB
pathway should be effective at inducing the apoptosis of tumor
cells.
MAPKs are important regulators of apoptosis, cell
proliferation and cell differentiation. Dysregulation of the MAPK
pathway is associated with tumor development and progression
(23). We demonstrated that high
concentrations of C. majus L. extract activated p38 MAPK,
MEKs and ERKs in A431 cells. Emerging evidence suggests that
activation of ERKs contributes to apoptosis. Recent studies showed
that kaempferol causes cancer cells to undergo apoptosis through an
ERK-dependent pathway (24).
Shikonin decreases the levels of phosphorylated EGFR, ERKs and
protein tyrosine kinases and increases intracellular levels of
apoptosis-related proteins, which leads to apoptosis of epidermoid
carcinoma cells (25). The
pro-apoptotic effect of wogonin was found to be mediated via
activation of ERKs and caspases and is correlated with interruption
of the PI3K/Akt/survivin signaling pathways in MCF-7 cells
(20). A number of studies have
shown the importance of ERK signaling in the regulation of
apoptosis (26,27). Although the ERK pathway delivers a
survival signal, studies using chemopreventive and chemotherapeutic
agents have linked activation of ERKs with induction of apoptosis
(28,29). Scutellaria baicalensis
extract was found to prevent hepatic fibrosis by promoting ERK-p53
pathways, which in turn caused G2/M cell cycle arrest and
activation of the caspase system resulting in apoptosis. It may
also induce Bax expression with concomitant decrease in Bcl-2 and
MAPK signaling in HSC-T6 cells (30). Here, we also found that sustained
activation of ERKs is involved in the C. majus L.
extract-induced growth inhibition and apoptosis in A431 cells. In
conclusion, the present study demonstrated that A431 cells are
susceptible to C. majus L. extract-induced inhibition of
proliferation and induction of apoptosis, which is mediated in part
by caspase activation and NF-κB inhibition via MAPK-independent
pathways. These results indicate the need for further research on
the in vivo efficacy of C. majus L. extract as a
possible anticancer agent.
Acknowledgements
This research was supported by the Bio-industry
Technology Development Program, Ministry for Food, Agriculture,
Forestry and Fisheries, Republic of Korea (grant no. 311059-4) and
partially supported by the Rural Development Administration,
Republic of Korea (grant no. PJ008475022012).
References
|
1
|
Fik E, Dalgalarrondo M, Haertle T and
Gozdzicka-Jozefiak A: Comparative biochemical analysis of lectin
and nuclease from Chelidonium majus L. Acta Biochim Pol.
47:413–420. 2000.PubMed/NCBI
|
|
2
|
Fik E, Wołun-Cholewa M, Kistowska M,
Warchoł JB and Gozdzicka-Jozefiak A: Effect of lectin from
Chelidonium majus L. on normal and cancer cells in culture. Folia
Histochem Cytobiol. 39:215–216. 2001.PubMed/NCBI
|
|
3
|
Biswas SJ, Bhattacharjee N and
Khuda-Bukhsh AR: Efficacy of a plant extract (Chelidonium majus L.)
in combating induced hepatocarcinogenesis in mice. Food Chem
Toxicol. 46:1474–1487. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tsuruo T, Naito M, Tomida A, Fujita N,
Mashima T, Sakamoto H and Haga N: Molecular targeting therapy of
cancer: drug resistance, apoptosis and survival signal. Cancer Sci.
94:15–21. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Debatin KM: Apoptosis pathways in cancer
and cancer therapy. Cancer Immunol Immunother. 53:153–159. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Levine B, Sinha S and Kroemer G: Bcl-2
family members: dual regulators of apoptosis and autophagy.
Autophagy. 4:600–606. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liang H, Gu MN, Yang CX, Wang HB, Wen XJ
and Zhou QL: Sevoflurane inhibits proliferation, induces apoptosis,
and blocks cell cycle progression of lung carcinoma cells. Asian
Pac J Cancer Prev. 12:3415–3420. 2011.PubMed/NCBI
|
|
8
|
Dong C, Davis RJ and Flavell RA: MAP
kinases in the immune response. Annu Rev Immunol. 20:55–72. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Johnson GL and Vaillancourt RR: Sequential
protein kinase reactions controlling cell growth and
differentiation. Curr Opin Cell Biol. 6:230–238. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Robinson MJ and Cobb MH: Mitogen-activated
protein kinase pathways. Curr Opin Cell Biol. 9:180–186. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cobb MH: MAP kinase pathways. Prog Biophys
Mol Biol. 71:479–500. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Stanciu M, Wang Y, Kentor R, et al:
Persistent activation of ERK contributes to glutamate-induced
oxidative toxicity in a neuronal cell line and primary cortical
neuron cultures. J Biol Chem. 275:12200–12206. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Colombo ML and Bosisio E: Pharmacological
activities of Chelidonium majus L. (Papaveraceae). Pharmacol Res.
33:127–134. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chor SY, Hui AY, To KF, et al:
Anti-proliferative and pro-apoptotic effects of herbal medicine on
hepatic stellate cell. J Ethnopharmacol. 100:180–186. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kim YI, Park SW, Choi IH, Lee JH, Woo HJ
and Kim Y: Effect of Orostachys japonicus on cell growth and
apoptosis in human hepatic stellate cell line LX2. Am J Chin Med.
39:601–613. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tang YJ, Yang JS, Lin CF, et al:
Houttuynia cordata Thunb extract induces apoptosis through
mitochondrial-dependent pathway in HT-29 human colon adenocarcinoma
cells. Oncol Rep. 22:1051–1056. 2009.PubMed/NCBI
|
|
17
|
Liu FS, Yang PY, Hu DN, Huang YW and Chen
MJ: Antrodia camphorata induces apoptosis and enhances the
cytotoxic effect of paclitaxel in human ovarian cancer cells. Int J
Gynecol Cancer. 21:1172–1179. 2011.PubMed/NCBI
|
|
18
|
Liu JJ, Zhang Y, Lin DJ and Xiao RZ:
Tanshinone IIA inhibits leukemia THP-1 cell growth by induction of
apoptosis. Oncol Rep. 21:1075–1081. 2009.PubMed/NCBI
|
|
19
|
Hsu PC, Huang YT, Tsai ML, Wang YJ, Lin JK
and Pan MH: Induction of apoptosis by shikonin through coordinative
modulation of the Bcl-2 family, p27, and p53, release of cytochrome
c, and sequential activation of caspases in human colorectal
carcinoma cells. J Agric Food Chem. 52:6330–6337. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Huang KF, Zhang GD, Huang YQ and Diao Y:
Wogonin induces apoptosis and down-regulates survivin in human
breast cancer MCF-7 cells by modulating PI3K-AKT pathway. Int
Immunopharmacol. 12:334–341. 2012. View Article : Google Scholar
|
|
21
|
Reed JC: Apoptosis-targeted therapies for
cancer. Cancer Cell. 3:17–22. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhu W, Hu H, Qiu P and Yan G: Triptolide
induces apoptosis in human anaplastic thyroid carcinoma cells by a
p53-independent but NF-κB-related mechanism. Oncol Rep.
22:1397–1401. 2009.PubMed/NCBI
|
|
23
|
Rubinfeld H and Seger R: The ERK cascade:
a prototype of MAPK signaling. Mol Biotechnol. 31:151–174. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jeong JC, Kim MS, Kim TH and Kim YK:
Kaempferol induces cell death through ERK and Akt-dependent
down-regulation of XIAP and survivin in human glioma cells.
Neurochem Res. 34:991–1001. 2009. View Article : Google Scholar
|
|
25
|
Singh F, Gao D, Lebwohl MG and Wei H:
Shikonin modulates cell proliferation by inhibiting epidermal
growth factor receptor signaling in human epidermoid carcinoma
cells. Cancer Lett. 200:115–121. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ma Y, Yu WD, Kong RX, Trump DL and Johnson
CS: Role of nongenomic activation of phosphatidylinositol
3-kinase/Akt and mitogen-activated protein kinase/extracellular
signal-regulated kinase kinase/extracellular signal-regulated
kinase 1/2 pathways in 1,25D3-mediated apoptosis in squamous cell
carcinoma cells. Cancer Res. 66:8131–8138. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jeon ES, Lee MJ, Sung SM and Kim JH:
Sphingosyl-phosphorylcholine induces apoptosis of endothelial cells
through reactive oxygen species-mediated activation of ERK. J Cell
Biochem. 100:1536–1547. 2007. View Article : Google Scholar
|
|
28
|
Yu W, Liao QY, Hantash FM, Sanders BG and
Kline K: Activation of extracellular signal-regulated kinase and
c-Jun-NH2-terminal kinase but not p38 mitogen-activated protein
kinases is required for RRR-α-tocopheryl succinate-induced
apoptosis of human breast cancer cells. Cancer Res. 61:6569–6576.
2001.PubMed/NCBI
|
|
29
|
Xu C, Shen G, Yuan X, et al: ERK and JNK
signaling pathways are involved in the regulation of activator
protein 1 and cell death elicited by three isothiocyanates in human
prostate cancer PC-3 cells. Carcinogenesis. 27:437–445. 2006.
View Article : Google Scholar
|
|
30
|
Pan TL, Wang PW, Leu YL, Wu TH and Wu TS:
Inhibitory effects of Scutellaria baicalensis extract on hepatic
stellate cells through inducing G2/M cell cycle arrest and
activating ERK-dependent apoptosis via Bax and caspase pathway. J
Ethnopharmacol. 139:829–837. 2012. View Article : Google Scholar : PubMed/NCBI
|