Introduction
Lung cancer is the leading cause of cancer-related
mortality among males worldwide, and the second leading cause of
cancer-related deaths among females, ranking second to only breast
cancer (1). Non-small cell lung
cancer (NSCLC) and small cell lung cancer are the main types of
lung cancer according to histologic classification, with NSCLC
accounting for ~80–85% of all lung cancer cases (2). Cigarette smoking and air pollution
have been proven to be the main causative factors of lung cancer.
Patients with lung cancer usually are diagnosed in advanced stages,
and most of lung cancer commonly metastasizes to the brain, bones,
liver and adrenal glands (3).
Although there are many common treatments, including surgery,
chemotherapy and radiotherapy, the survival rates of lung cancer
patients are still very low and recurrence risk is high.
Genetic and epigenetic changes have been shown to
play important roles in the formation and progression of multiple
types of cancers, including the silencing of tumor suppressors,
overexpression of oncogenes and changes in microRNAs (miRNAs). Over
the past several decades, the molecular mechanisms of
tumorigenesis, progression and metastasis of lung cancer remain
unclear, in spite of a large number of research studies. New and
effective therapies are still lacking. Given the high mortality of
lung cancer, a better understanding of the mechanisms that underlie
lung carcinogenesis is needed.
miRNAs are a class of small endogenous
post-transcriptional regulatory non-coding RNAs (containing ~22
nucleotides), first discovered by Lee et al (4) in 1993. Primary miRNAs capped with a
specially modified nucleotide at the 5′ end and polyadenylated with
multiple adenosines [a poly(A) tail] are transcribed by RNA
polymerases II (5,6) and then processed in the nucleus by
Drosha and Pasha to generate hairpin loop pre-miRNAs composed of
~70 nucleotides each. Subsequently, the pre-miRNA is exported out
of the nucleus into the cytoplasm in a process involving the
nucleocytoplasmic shuttler Exportin-5, which is dependent on
RAN-GTP (7). In the cytoplasm, the
pre-miRNA hairpin is cleaved by the RNase III enzyme Dicer,
yielding an miRNA:miRNA* duplex ~22 nucleotides in
length (8). One strand is then
incorporated into the RNA-induced silencing complex (RISC), where
the miRNA and its mRNA target interact. The interaction of miRNAs
with mRNAs normally results in the cleavage or translation
inhibition of the target mRNA by binding to the 3′ untranslated
regions.
miRNAs are involved in a large number of biological
processes, including cell proliferation, apoptosis, metabolism,
cell differentiation and others (9–11).
Moreover, abundant evidence has shown that dysfunction of miRNAs is
associated with various human diseases, including cancer (12), and a recent study showed that miRNAs
can function not only as oncogenes but also as tumor suppressors
(13). Over the past several years,
several experiments have quantified miRNAs in lung cancer tissues
and several dysregulated miRNAs have been revealed: let-7,
miR-17–92 cluster, miR-155, miR-145 and hsa-miR-21 (14).
The miR-212/132 family is highly conserved in
vertebrates. miR-132 and miR-212 genes are arrayed in tandem on
chromosome 17p13.3 in humans, which is referred to as the
miR-212/132 cluster. Therefore, miR-132 and miR-212 are transcribed
simultaneously to the primary product pri-miRNA-212/132 in the form
of a gene cluster. Then, pri-miRNA-212/132 is processed and cleaved
to the mature miRNAs (15). Mature
miR-132 and miR-212 possess similar sequences and share the same
seed region (a region located between nucleotides 2 and 8 in the 5′
region of the miRNA), suggesting that they target the same mRNAs.
Nevertheless, this ‘double-targeting’ by both miRNAs has been
reported for only a few mRNAs to date and most of the mRNAs are
singly targeted (16).
Numerous studies have shown that the miR-212/132
cluster is necessary for the proper development, maturation and
function of neurons. Therefore, its dysregulation is the cause of
several neurological disorders, such as Alzheimer’s disease. In
addition, it can regulate circadian rhythms and is involved in
inflammation, immune progress, and drug addiction (16). Recently, miR-212/132 was found to be
dysregulated in many cancers and many studies have begun to focus
on its biological roles in cancers. Zhang et al (17) discovered that miR-132 was decreased
in pancreatic cancer and that it could suppress the proliferation
of pancreatic cancer cells through the Akt pathway. However, there
are few reports on the potential function of miR-212/132 in the
progression of other human cancers, including lung cancer.
Therefore, in the present study, we investigated the role of the
miR-212/132 cluster in modulating lung cancer progression. Our
study may provide insight into the biological function of the
miR-212/132 cluster in lung cancer.
Materials and methods
miR-212/132 overexpression vector and
inhibitors
The miR-212/132 overexpression vector
(pLMP-miR-212/132) and control vector plasmid (pLMP) were
constructed and verified by Huaan Pingkang Biotech Co., Ltd.
(Shenzhen, China). The miRNA inhibitor targeting miR-132 and
miR-212 (miR-212/132 inhibitor) and the negative inhibitor (NC
inhibitor) were obtained from GenePharma (Shanghai, China).
Cell culture
Human lung cancer cell lines A549 and H1299 were
cultured in Dulbecco’s modified Eagle’s medium (DMEM) supplemented
with 10% fetal bovine serum (FBS), 100 μg/ml streptomycin, and 100
U/ml penicillin (Gibco, Grand Island, NY, USA). Cells were grown in
a 37°C incubator with 5% CO2.
Cell viability assay
Cell viability was evaluated using the
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide
(MTT) assay. Cells were plated in 96-well plates. The next day, the
cells were transfected with plasmids or inhibitors according to the
experimental design. After 48 h, the cells were then incubated with
20 μl MTT (5 mg/ml) for 4 h. Next, the medium was carefully removed
and 100 μl DMSO was added to the plates. The optical density (OD)
was measured at 570 nm with a reference wavelength 650 nm using a
microplate reader (Bio-Rad, Hercules, CA, USA).
Focus formation
A549 or H1299 cells were seeded onto 6-well plates
and were then transfected with the indicated vectors. After
incubation for ~10 days, the cells were fixed and stained with
crystal violet. Colonies consisting of more than 50 cells were
counted using a microscope.
Clonogenic assay of radiosensitivity
Cells transfected with plasmids or inhibitors were
plated at different densities. Then, cells were irradiated with 160
kV X-rays at a dose rate of 1.15 Gy/min by a biological research
irradiator (RadSource Technologies, Suwanee, GA, USA) at various
doses. The cells were then grown for ~10 days to allow for colony
formation and were subsequently fixed with methanol and stained
using 1% crystal violet. Colonies containing 50 or more cells were
designated as a clone and counted.
Western blotting
Cells were lysed by RIPA lysis buffer on ice for 40
min and centrifuged at 13,000 rpm for 10 min at 4°C. The
supernatant was collected and subjected to western-blotting. Cell
lysates containing equal amount of protein were fractionated by 10%
SDS-PAGE, and electrically transferred to PVDF membranes
(Millipore, Bedford, MA, USA). Then, non-specific binding was
blocked with Tris-buffered saline containing 5% nonfat milk for 1–2
h at room temperature. Next, the membranes were incubated with the
appropriate primary antibody against p21 or cyclin D1 (both from
Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA) overnight at a
1:1,000 dilution. After 4 TBST washes, the membranes were then
incubated with corresponding horseradish peroxidase-conjugated
secondary antibodies at a 1:1,000 dilution for 1 h at room
temperature. Bound secondary antibodies were detected using
enhanced chemiluminescence method.
Flow cytometric analysis
A549 or H1299 cells were transfected with the
indicated vectors. After 24 h, cells were harvested and fixed
overnight with 70% ice-cold ethanol. Cells were stained with
propidium iodide (PI) containing RNase A for 30 min. The
distribution of the cell cycle was detected using a flow cytometer
(Beckman Coulter Inc., Fullerton, CA, USA). For apoptosis analysis,
cells were cultured for 48 h after transfection. Next, the cells
were harvested and washed with ice-cold phosphate-buffered saline
(PBS), and then stained using the Annexin
V-phycoerythrin/7-aminoactinomycin D apoptosis detection kit (BD
Biosciences, San Jose, CA, USA). The percentage of apoptotic cells
was also measured by a flow cytometer within 1 h.
Wound healing assay
H1299 cells were transfected with the indicated
vectors. Similar sized wounds were made by scraping a conventional
10-μl micropipette tip across the monolayer. The cells were washed
three times with PBS and then DMEM was added without FBS to the
plates. The distance between the wound edges was observed and
photographed immediately after wounding and 24 h later. The
distance between the wound edges was evaluated using ImageJ
software (NIH, Bethesda, MD, USA) and expressed as a percentage of
the initial wound distance.
Statistical analysis
Data are expressed as the mean ± standard error of
the mean (SEM) of at least three independent experiments. Analysis
was performed using the Student’s t-test. The sensitizer
enhancement ratios (SER) were measured according to the
multi-target single hit model. Statistical analysis was performed
using SPSS software (Release 18.0; SPSS Inc., San Rafael, CA, USA).
Data were considered statistically significant at P<0.05.
Results
miR-212/132 suppresses the proliferation
of lung cancer cells in vitro
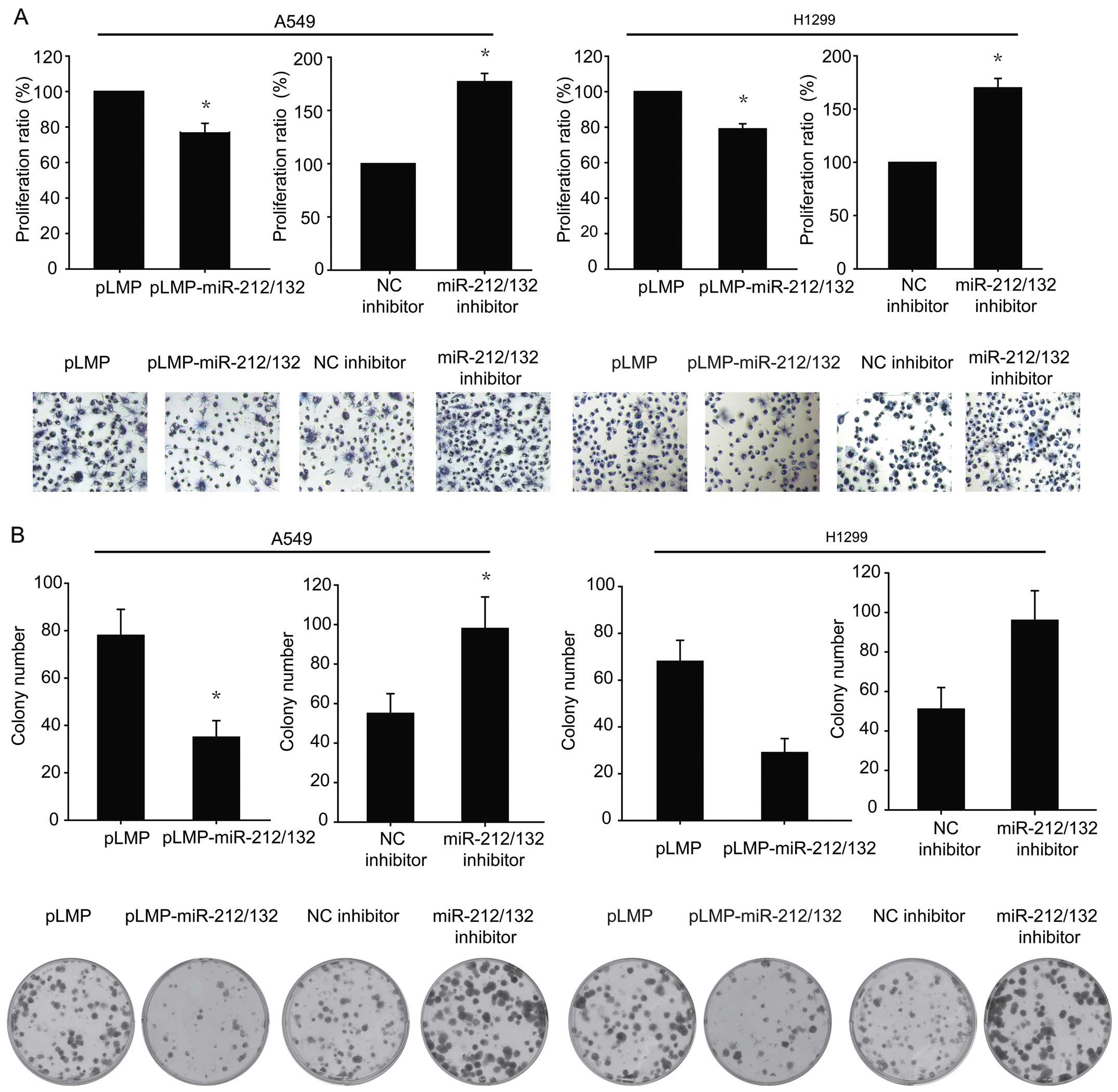
To investigate the effect of miR-212/132 on the
proliferation of lung cancer cells, an miR-212/132-overexpressing
vector (pLMP-miR-212/132) or a control vector (pLMP) were
transiently transfected into A549 and H1299 cells, and the
resulting growth was measured with MTT and colony formation assays.
The results of MTT and colony formation assays revealed that
upregulation of miR-212/132 suppressed the growth and focus
formation of A549 and H1299 cells, compared to the corresponding
controls (Fig. 1). We further
inhibited miR-212/132 expression using an inhibitor. The
downregulation of miR-212/132 promoted cell proliferation and
colony formation (Fig. 1).
miR-212/132 increases the cell apoptosis
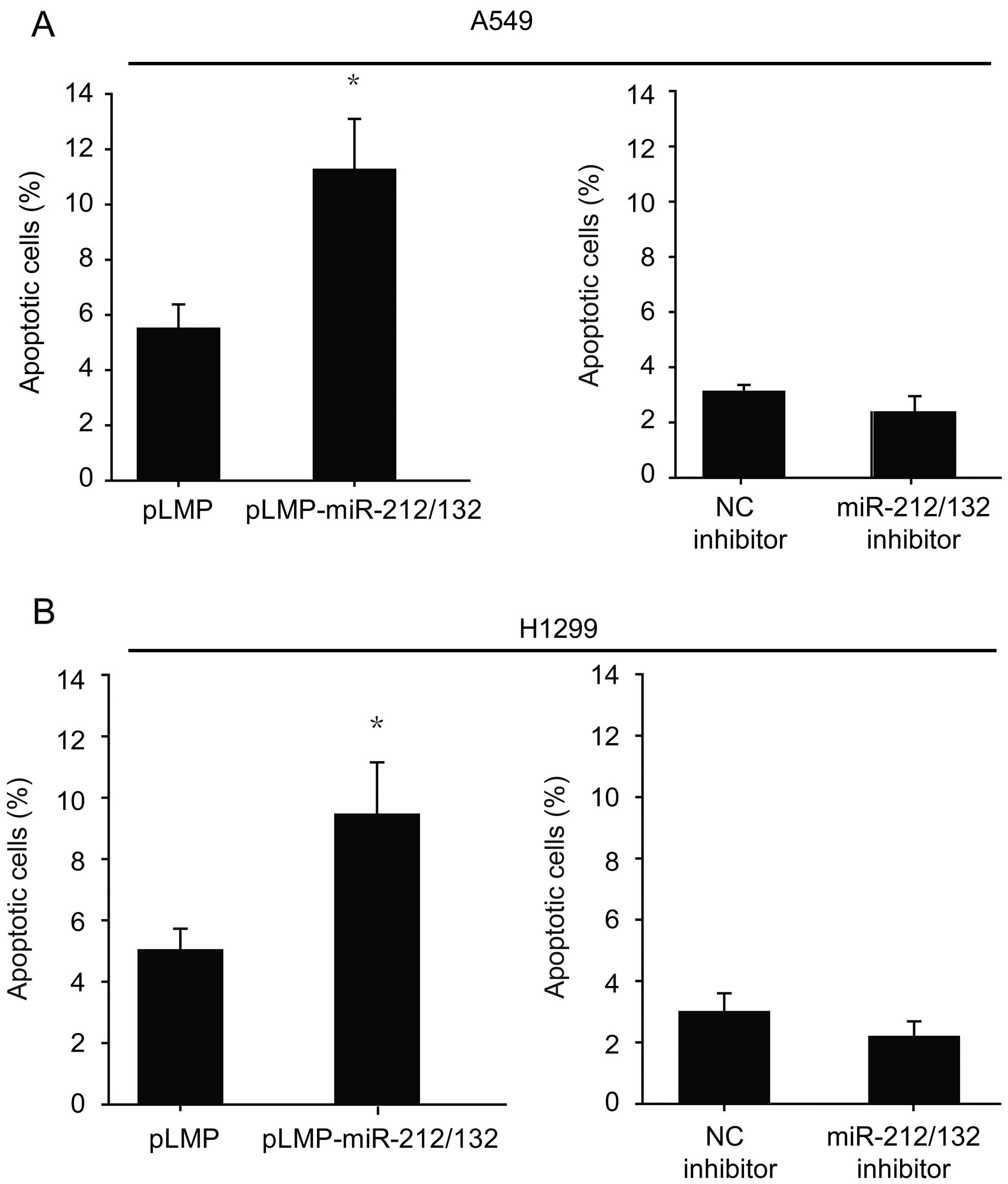
of lung cancer cells
To investigate the changes in cell apoptosis of lung
cancer cells treated with miR-212/132, cells were stained with
Annexin V-phycoerythrin/7-aminoactinomycin D and subjected to flow
cytometric analysis. As shown in Fig.
2, treatment of miR-212/132 induced a substantial increase in
the apoptotic rate when compared to this rate in the corresponding
control groups. The downregulation of miR-212/132 decreased cell
apoptosis but the difference was not statistically significant
(Fig. 2).
miR-212/132 induces cell cycle
arrest
To investigate the effects of miR-212/132 on cell
cycle progression, flow cytometric analysis was used to observe the
cell cycle distribution 24 h after transfection. miR-212/132 led to
an increase in the percentage of cells in G1 phase compared with
the control group, and this cell cycle arrest was relieved when the
miR-212/132 inhibitor was transfected (Fig. 3A). Next, we analyzed the expression
of p21 and cyclin D1, which are implicated in cell cycle
progression. As shown in Fig. 3B,
cells transfected with miR-212/132 exhibited upregulated p21
expression and reduced cyclin D1 expression. Conversely, cells
transfected with the miR-212/132 inhibitor displayed reduced
expression of p21 and upregulated expression of cyclin D1,
indicating that miR-212/132 inhibits the cell cycle progression of
lung cancer.
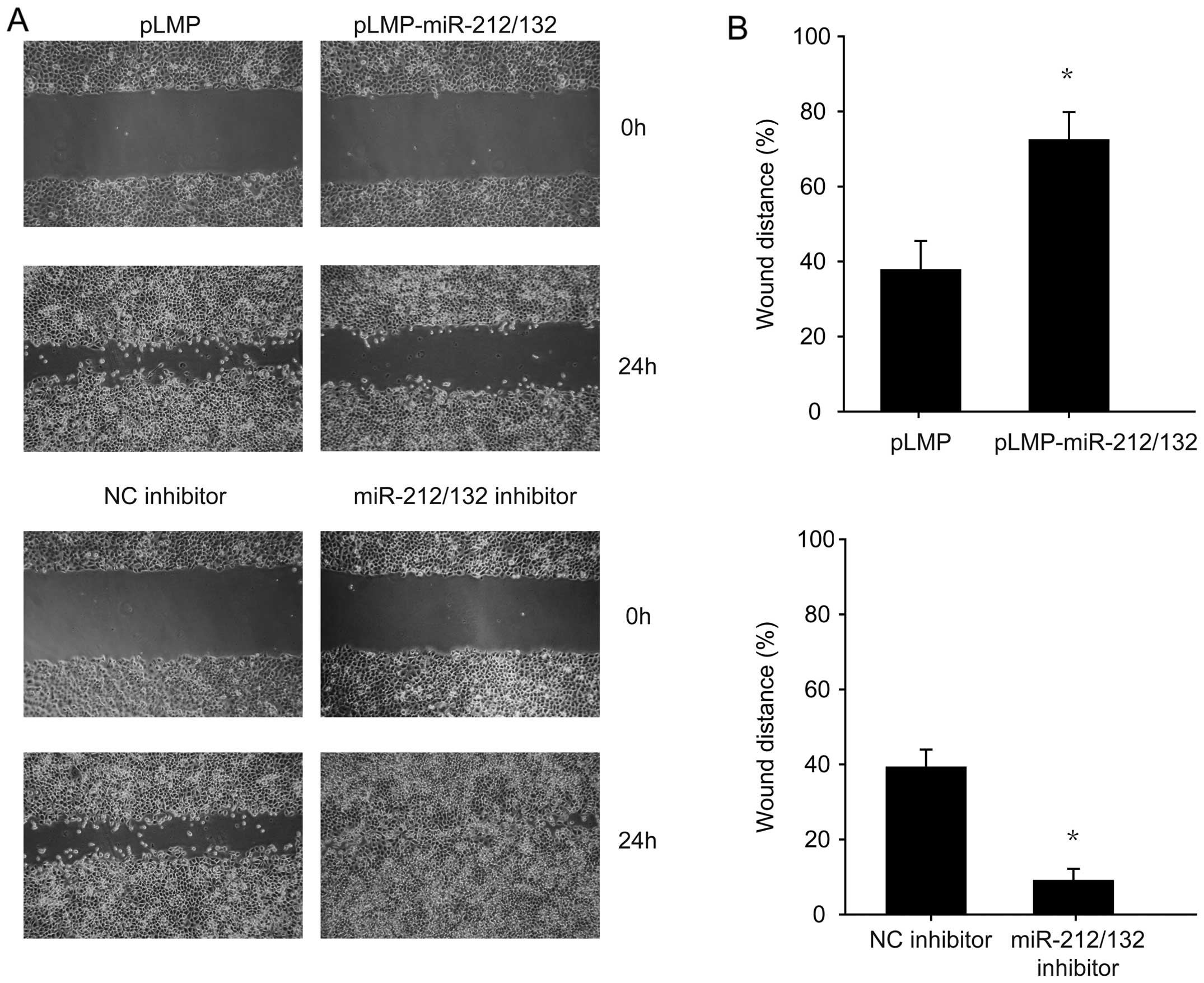
miR-212/132 transfection diminishes the
migration of H1299 cells
The wound healing assay was employed to investigate
the correlation between miR-212/132 and H1299 cell migration. The
results indicated that miR-212/132 inhibited the migration of H1299
cells compared to the control group (Fig. 4). Conversely, cells transfected with
the miR-212/132 inhibitor showed enhanced migration (Fig. 4). These data suggest that
miR-212/132 inhibited the migration, which could be reversed by the
miR-212/132 inhibitor.
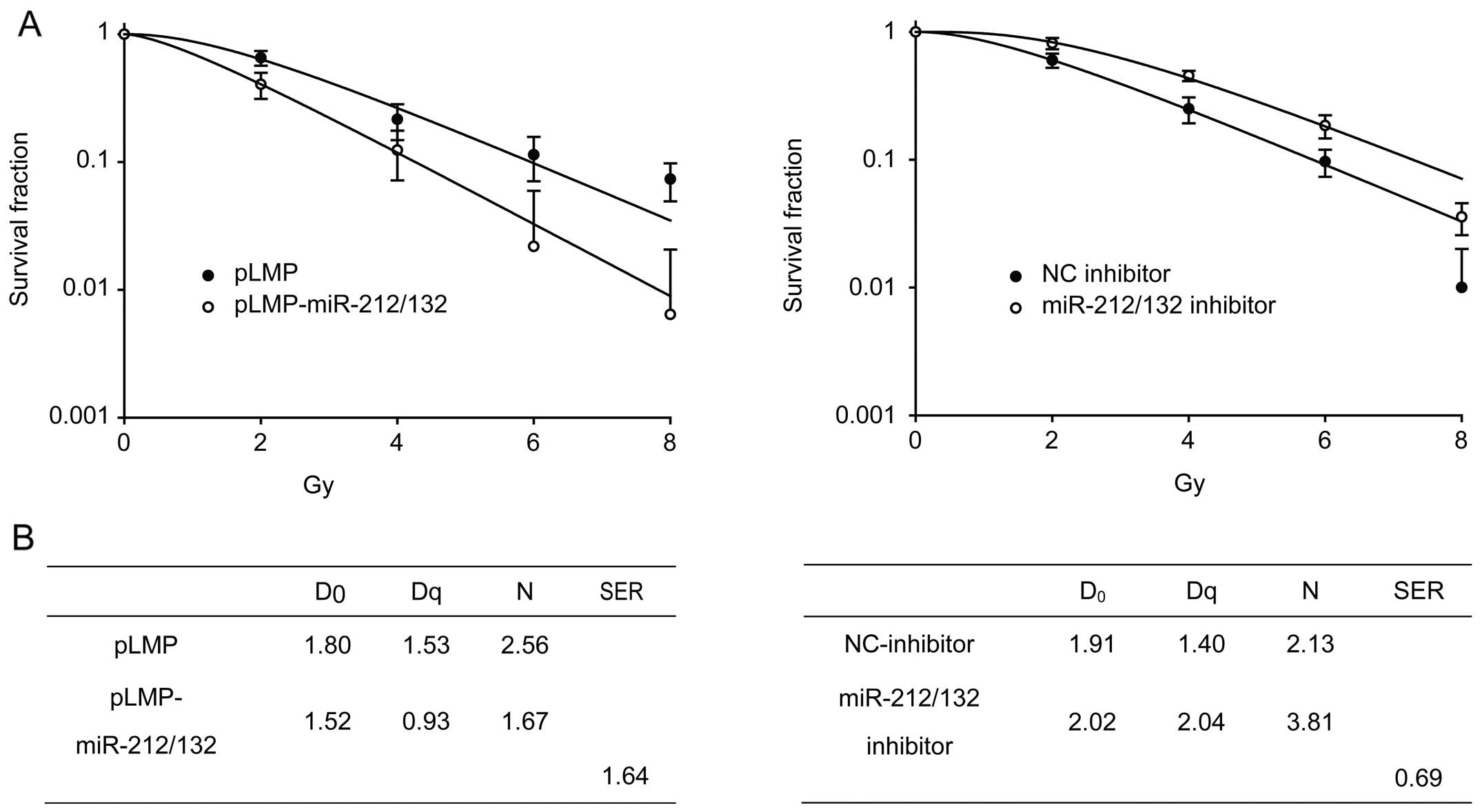
miR-212/132 enhances the radiosensitivity
of H1299 cells
To investigate the effect of miR-212/132 on the
radiosensitivity of H1299 cells, cells transfected with the
different vectors were irradiated with different X-ray doses, and
clonogenic assays were performed. As shown in Fig. 5, cells that were transiently
transfected with pLMP-miR-212/132 exhibited lower clonogenic
survival rates than cells treated with radiation alone, indicating
that miR-212/132 enhanced the radiosensitivity of H1299 cells.
Next, we transfected the miR-212/132 inhibitor into H1299 cells,
and found that the cells became resistant to radiation (Fig. 5).
Discussion
In recent years, many groups have focused on the
molecular mechanisms of lung cancer development, including miRNAs.
miRNAs are involved in the formation and progression of lung cancer
and their expression profiles have been used to classify cancers
and their signature predicts survival and relapse in lung cancer
(18,19). In addition, it was revealed that
miRNAs participate in multiple biological processes of lung cancer,
including proliferation, apoptosis, development and metastasis.
Song et al (20) found that
miR-483-5p promotes the invasion and migration of lung
adenocarcinoma by targeting RhoGDI1 and ALCAM. Yoo et al
(21) reported that miR-9500
inhibited the proliferation and migration of human lung cancer
cells by targeting Akt1. Shi et al (22) showed that miR-34a was a negative
regulator of the tumorigenic properties of NSCLC cells and
inhibited their growth.
miR-132 expression has been reported to be increased
in several types of cancers including lung cancer (14), squamous cell carcinoma of the tongue
(23), colorectal carcinoma
(24) and esophageal cancer
(25). However, decreased miR-132
expression has been observed in osteosarcoma (26) and liver cancer (27). Increased miR-212 expression was
reported in oral squamous cell carcinoma (28) and colorectal carcinoma (23), and miR-212 has been shown to be
downregulated in gastric cancer (29), NSCLC (30) and head and neck squamous cell
carcinoma (31). The different
levels of expression of these miRNAs in various cancers indicate
that they may exert diverse functions in cancer cells. To our
knowledge, no report has taken an in-depth look into the role of
miR-212/132 in lung cancer cells.
In the present study, we found that upregulation of
miR-212/132 significantly suppressed the growth and focus formation
of A549 and H1299 cells and conferred radiosensitivity to H1299
cells, whereas downregulation of miR-212/132 reversed these
effects. Furthermore, miR-212/132 overexpression induced cell cycle
arrest at the G1/S phase transition of A549 and H1299 cells, and
the miR-212/132 inhibitor abrogated this arrest. Therefore, the
inhibition of cellular proliferation by overexpression of
miR-212/132 may be due to restrained cancer cell division. In
addition, the miR-212/132 overexpression group exhibited a higher
percentage of cells that underwent apoptosis, which may also lead
to the inhibiiton of growth and colony formation. To identify the
molecular mechanism, p21 and cyclin D1 were detected at the protein
level in each group and we discovered that cells transfected with
miR-212/132 had upregulated p21 expression and reduced cyclin D1
expression. Conversely, cells transfected with the miR-212/132
inhibitor displayed reduced expression of p21 and upregulated
expression of cyclin D1, suggesting that miR-212/132 may mediate
proliferation and cell cycle arrest through p21 and cyclin D1
(32).
You et al (33) found that miR-132 suppressed the
migration of lung cancer cells by targeting the EMT regulator ZEB2.
Therefore, we hypothesized that miR-212/132 may also suppress the
migration of lung cancer cells. As expected, upregulation of
miR-212/132 diminished the migration of H1299 cells and
downregulation of miR-212/132 enhanced their migration.
The predicted targets of miR-132 and miR-212 include
hundreds of genes implicated in multiple pathways. These targets
may be the upstream of Akt, including ACVR2B, ACVR1, connective
tissue growth factor, HB-EGF (31,34),
GTPase-activating proteins [p120RasGAP (35) and p250GAP (36,37)],
Rb1 (38), STAT4 (39) and ZO-1 (40). We did not investigate the direct
targets of miR-132 and miR-212, in part because the roles of the
predicted targets listed above in lung cancer progression are not
clear, which warrants further investigation. Additionally, since
each miRNA can regulate numerous protein-coding genes, its
functions can be interpreted as the sum of the functions of the
genes it regulates (41).
Therefore, we considered it more important to analyze the
consequences of miR-212/132 dysregulation in lung cancer
progression.
In conclusion, we illustrated the biological role of
miR-212/132 in lung cancer cells. Our findings suggest that
miR-212/132 may be a novel tumor-suppressor miRNA. miR-212/132
blocked proliferation and migration, and led to cell cycle arrest
of lung cancer cells through modulating the expression of p21 and
cyclin D1. This study may provide a potential therapeutic target
for the treatment of lung cancer.
Acknowledgements
This study as funded by the National Natural Science
Foundation of China (81402518 and 81472920), the Jiangsu Provincial
Special Program of Medical Science (BL2012046), the Changzhou
Social Development Project (CE20125026 and CE20135050) and the
Changzhou Scientific Program (ZD201315 and CY20130017).
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ramalingam S and Belani C: Systemic
chemotherapy for advanced non-small cell lung cancer: recent
advances and future directions. Oncologist. 13(Suppl 1): s5–s13.
2008. View Article : Google Scholar
|
|
3
|
Chaffer CL and Weinberg RA: A perspective
on cancer cell metastasis. Science. 331:1559–1564. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lee RC, Feinbaum RL and Ambros V: The C.
elegans heterochronic gene lin-4 encodes small RNAs with antisense
complementarity to lin-14. Cell. 75:843–854. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lee Y, Kim M, Han J, et al: MicroRNA genes
are transcribed by RNA polymerase II. EMBO J. 23:4051–4060. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cai X, Hagedorn CH and Cullen BR: Human
microRNAs are processed from capped, polyadenylated transcripts
that can also function as mRNAs. RNA. 10:1957–1966. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Murchison EP and Hannon GJ: miRNAs on the
move: miRNA biogenesis and the RNAi machinery. Curr Opin Cell Biol.
16:223–229. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lund E and Dahlberg JE: Substrate
selectivity of exportin 5 and Dicer in the biogenesis of microRNAs.
Cold Spring Harb Symp Quant Biol. 71:59–66. 2006. View Article : Google Scholar
|
|
9
|
Bueno MJ, Pérez de Castro I and Malumbres
M: Control of cell proliferation pathways by microRNAs. Cell Cycle.
7:3143–3148. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang Y and Lee CG: MicroRNA and cancer -
focus on apoptosis. J Cell Mol Med. 13:12–23. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen CZ, Li L, Lodish HF and Bartel DP:
microRNAs modulate hematopoietic lineage differentiation. Science.
303:83–86. 2004. View Article : Google Scholar
|
|
12
|
Wang D, Qiu C, Zhang H, Wang J, Cui Q and
Yin Y: Human microRNA oncogenes and tumor suppressors show
significantly different biological patterns: from functions to
targets. PLoS One. 5:e130672010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen CZ: microRNAs as oncogenes and tumor
suppressors. N Engl J Med. 353:1768–1771. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yanaihara N, Caplen N, Bowman E, et al:
Unique microRNA molecular profiles in lung cancer diagnosis and
prognosis. Cancer Cell. 9:189–198. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tognini P and Pizzorusso T:
MicroRNA212/132 family: molecular transducer of neuronal function
and plasticity. Int J Biochem Cell Biol. 44:6–10. 2012. View Article : Google Scholar
|
|
16
|
Wanet A, Tacheny A, Arnould T and Renard
P: miR-212/132 expression and functions: within and beyond the
neuronal compartment. Nucleic Acids Res. 40:4742–4753. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang S, Hao J, Xie F, et al:
Downregulation of miR-132 by promoter methylation contributes to
pancreatic cancer development. Carcinogenesis. 32:1183–1189. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lu J, Getz G, Miska EA, et al: MicroRNA
expression profiles classify human cancers. Nature. 435:834–838.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yu SL, Chen HY, Chang GC, et al: MicroRNA
signature predicts survival and relapse in lung cancer. Cancer
Cell. 13:48–57. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Song Q, Xu Y, Yang C, et al: miR-483-5p
promotes invasion and metastasis of lung adenocarcinoma by
targeting RhoGDI1 and ALCAM. Cancer Res. 74:3031–3042. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yoo JK, Jung HY, Lee JM, et al: The novel
miR-9500 regulates the proliferation and migration of human lung
cancer cells by targeting Akt1. Cell Death Differ. 21:1150–1159.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shi Y, Liu C, Liu X, Tang DG and Wang J:
The microRNA miR-34a inhibits non-small cell lung cancer (NSCLC)
growth and the CD44hi stem-like NSCLC cells. PLoS One.
9:e900222014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wong TS, Liu XB, Wong BY, Ng RW, Yuen AP
and Wei WI: Mature miR-184 as potential oncogenic microRNA of
squamous cell carcinoma of Tongue. Clin Cancer Res. 14:2588–2592.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Schetter AJ, Leung SY, Sohn JJ, et al:
MicroRNA expression profiles associated with prognosis and
therapeutic outcome in colon adenocarcinoma. JAMA. 299:425–436.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ogawa R, Ishiguro H, Kuwabara Y, et al:
Expression profiling of micro-RNAs in human esophageal squamous
cell carcinoma using RT-PCR. Med Mol Morphol. 42:102–109. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gougelet A, Pissaloux D, Besse A, et al:
Micro-RNA profiles in osteosarcoma as a predictive tool for
ifosfamide response. Int J Cancer. 129:680–690. 2011. View Article : Google Scholar
|
|
27
|
Meng F, Henson R, Wehbe-Janek H, Ghoshal
K, Jacob ST and Patel T: MicroRNA-21 regulates expression of the
PTEN tumor suppressor gene in human hepatocellular cancer.
Gastroenterology. 133:647–658. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Scapoli L, Palmieri A, Lo Muzio L, et al:
MicroRNA expression profiling of oral carcinoma identifies new
markers of tumor progression. Int J Immunopathol Pharmacol.
23:1229–1234. 2010.
|
|
29
|
Wada R, Akiyama Y, Hashimoto Y, Fukamachi
H and Yuasa Y: miR-212 is downregulated and suppresses
methyl-CpG-binding protein MeCP2 in human gastric cancer. Int J
Cancer. 127:1106–1114. 2010. View Article : Google Scholar
|
|
30
|
Incoronato M, Garofalo M, Urso L, et al:
miR-212 increases tumor necrosis factor-related apoptosis-inducing
ligand sensitivity in non-small cell lung cancer by targeting the
antiapoptotic protein PED. Cancer Res. 70:3638–3646. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hatakeyama H, Cheng H, Wirth P, et al:
Regulation of heparin-binding EGF-like growth factor by miR-212 and
acquired cetuximab-resistance in head and neck squamous cell
carcinoma. PLoS One. 5:e127022010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gartel AL and Radhakrishnan SK: Lost in
transcription: p21 repression, mechanisms and consequences. Cancer
Res. 65:3980–3985. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
You J, Li Y, Fang N, et al: MiR-132
suppresses the migration and invasion of lung cancer cells via
targeting the EMT regulator ZEB2. PLoS One. 9:e918272014.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Molnár V, Érsek B, Wiener Z, et al:
MicroRNA-132 targets HB-EGF upon IgE-mediated activation in murine
and human mast cells. Cell Mol Life Sci. 69:793–808. 2012.
View Article : Google Scholar
|
|
35
|
Anand S, Majeti BK, Acevedo LM, et al:
MicroRNA-132-mediated loss of p120RasGAP activates the endothelium
to facilitate pathological angiogenesis. Nat Med. 16:909–914. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Vo N, Klein ME, Varlamova O, et al: A
cAMP-response element binding protein-induced microRNA regulates
neuronal morphogenesis. Proc Natl Acad Sci USA. 102:16426–16431.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wayman GA, Davare M, Ando H, et al: An
activity-regulated microRNA controls dendritic plasticity by
downregulating p250GAP. Proc Natl Acad Sci USA. 105:9093–9098.
2008. View Article : Google Scholar
|
|
38
|
Park JK, Henry JC, Jiang J, et al: miR-132
and miR-212 are increased in pancreatic cancer and target the
retinoblastoma tumor suppressor. Biochem Biophys Res Commun.
406:518–523. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Huang Y, Lei Y, Zhang H, Hou L, Zhang M
and Dayton AI: MicroRNA regulation of STAT4 protein expression:
rapid and sensitive modulation of IL-12 signaling in human natural
killer cells. Blood. 118:6793–6802. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tang Y, Banan A, Forsyth CB, et al: Effect
of alcohol on miR-212 expression in intestinal epithelial cells and
its potential role in alcoholic liver disease. Alcohol Clin Exp
Res. 32:355–364. 2008. View Article : Google Scholar
|
|
41
|
Sonkoly E, Wei T, Janson PC, et al:
microRNAs: novel regulators involved in the pathogenesis of
psoriasis? PLoS One. 2:e6102007. View Article : Google Scholar : PubMed/NCBI
|