Introduction
Colorectal cancer is one of the leading causes of
cancer-related mortality in most industrialized nations (1), and the effectiveness of conventional
treatments for colorectal cancer is limited. Despite the fact that
advances in the adjuvant treatment of colorectal cancer have been
achieved for patients with lymph node-positive disease (2,3), the
risk of recurrence is significantly high. Hence, extensive research
has been conducted to find an effective adjuvant therapy for
colorectal cancer (4), such as
non-steroidal anti-inflammatory drugs, immunotherapy and gene
therapy (5).
Thioredoxin (Trx) and thioredoxin reductase 1 (TR1)
are among the major redox regulators in mammalian cells and control
many cellular processes, including proliferation, defense against
oxidative stress and apoptosis (6,7). Thio
redoxin-1 (Trx-1), an ubiquitously expressed low-molecular-weight
redox protein, is overexpressed in several types of human cancers
(8–10) and has multiple effects in cells that
include transactivating activity of a number of redox-sensitive
transcription factors (11,12), and the regulation of DNA binding
(13). Trx-1 has been shown to play
a critical role in a series of human diseases including cancer of
the cervix, pancreas, lung, colorectal and squamous cell cancer
(8–10,14–16).
Trx-1-transfected cells exhibit increased colony formation and cell
growth in soft agar (17).
Retrospective analyses in colorectal cancer indicate that increased
expression of Trx-1 in cancer cells may be an independent
prognostic factor (10).
1-Methylpropyl 2-imidazolyl disulfide (PX-12) is a
potent inhibitor of Trx-1 by irreversible thioalkylation of
Cys73 which lies outside the conserved redox catalytic
site of Trx-1, causing inhibition of Trx-dependent cell growth
(18,19). PX-12 has been shown to have in
vivo antitumor activity against human tumor xenografts
including HT-29 colon cancer in SCID mice and has been tested in a
phase I clinical trial in patients (20). In the present study, we investigated
the antitumor effects of the Trx-1 redox signaling inhibitor PX-12
on colorectal cancer in vitro, including induction of
apoptosis, growth arrest, inhibition of cell migration and
invasion.
Materials and methods
Cell lines and cell culture
Two colorectal cancer cell lines SW620 and DLD-1
were used in the present study. They were cultured in Dulbecco’s
modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine
serum (FBS; Invitrogen, Carlsbad, CA, USA), 100 μg/ml streptomycin
and 100 U/ml penicillin in a humidified 5% CO2 incubator
at 37°C.
Cell viability and colony formation
assays
Absorbance of
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT;
Sigma-Aldrich) in living cells was measured to determine the effect
of PX-12 on cell viability. Briefly, 1×104 cells/well
were seeded in 96-well plates and exposed to the designated
concentrations of PX-12 for the indicated times, MTT solution was
added to each well. After the incubating time (4 h at 37°C) was
completed, the medium was withdrawn from the plates and 200 μl DMSO
was added to each well to solubilize the formazan crystals. The
optical density was measured at 490 nm using a Universal Microplate
Reader (ELx800; BioTek Instruments, Inc., Winooski, VT, USA).
For the colony formation assay, 5,000 cells were
seeded into a 10-cm culture dish and incubated at 37°C in a
humidified 5% CO2 incubator for 14 days to form
colonies, which were then stained with Coomassie blue. The rate of
colony formation was calculated with the following equation: Colony
formation rate = (number of colonies/number of seeded cells) ×
100%.
Cell cycle analysis
Cell cycle analysis was performed using propidium
iodide (PI) (Ex/Em=488/617 nm; Sigma-Aldrich) staining. Briefly,
2×105 cells/well in 6-well plates were treated with 5 μM
PX-12. At 24 h after the incubation time was completed, total
cells, including floating cells, were then washed with
phosphate-buffered saline (PBS) and fixed in 70% (v/v) ethanol. The
washing step was repeated again with PBS, and then staining with 50
mg/l PI which contained 0.1% Triton X-100 and RNase (100 mg/l) was
carried out. Cellular DNA content was measured using a FACSCalibur
flow cytometer and analyzed using ModFit LT 2.0 software (both from
Becton-Dickinson, Franklin Lakes, NJ, USA).
Apoptosis assay
For the apoptosis assay, apoptotic cell death was
determined by Annexin V-FITC Apoptosis Detection Kit II (BD
Pharmingen™, San Diego, CA, USA) according to the manufacturer’s
protocols. Briefly, 2×105 cells/well were seeded in
6-well plates and treated with 5 μM PX-12 for 48 h. After the
incubation time was completed, total cells, including floating
cells, were washed twice with cold PBS and then resuspended in
binding buffer at a density of 1×106 cells/ml. Annexin
V-FITC (5 μl) and PI (1 μg/ml) were then added, and the cells were
analyzed with the FACSCalibur flow cytometer.
Migration and invasion assay
The effect of PX-12 on the migration and invasive
abilities of the colorectal cancer cells in vitro was
examined using Transwell assays. PX-12 (1 μM) which has little
effect on cell growth at 24 h was used to treat the colorectal
cancer cells. Migration and invasion assays were performed in
Costar Transwell cell culture chamber inserts (Corning Costar
Corp., Cambridge, MA, USA) with an 8-μm pore size. For the
migration assay, a total of 1×105 cells were collected
and seeded to the upper part of the chamber in 100 μl of serum-free
medium. Medium with 10% FBS was added to the bottom chamber as a
chemoattractant. Cells were supplemented with PX-12 during the
incubation time (24 h at a concentration of 1 mM). After 24 h, the
cells that migrated to the lower surface of the membrane were fixed
with 4% of paraformaldehyde, and stained with 0.1% of crystal
violet. The number of cells that migrated to the lower surface of
the membrane was counted in five random selected fields under a
microscope at a magnification of ×100. For the invasion assays, the
polycarbonate membrane filters were coated with 1:3 diluted
Matrigel (BD Biosciences, USA). All experiments were carried out in
triplicate.
Reverse transcription (RT)-PCR
Total RNA was extracted from cells using TRIzol
(Invitrogen), and reverse-transcribed into cDNA using a SuperScript
II First Strand DNA Synthesis kit (Promega, Madison, WI, USA). The
quantification of mRNA levels was performed using
SYBR®-Green PCR Master Mix and an ABI 7500 Real-Time PCR
System (both from Applied Biosystems, Warrington, UK). The
sequences of the forward and reverse primers for the NOX1, CDH17,
KLF17, S100P, S100A4 and GAPDH genes used in the real-time RT-PCR
are listed in Table I. The relative
expression of these genes was normalized to that of GAPDH, an
endogenous housekeeping gene.
 | Table IPrimers for the NOX1, CDH17, KLF17,
S100P, S100A4 and GAPDH genes. |
Table I
Primers for the NOX1, CDH17, KLF17,
S100P, S100A4 and GAPDH genes.
| Gene name | Forward primer | Reverse primer |
|---|
| NOX1 |
5′-GGATGATCGTGACTCCCACT-3′ |
5′-TTTGGATGGTGCATAACAA-3′ |
| CDH17 |
5′-GACAATCGACCCACGTTTCT-3′ |
5′-GCTCCCGTTTTGTTGTTGAT-3′ |
| KLF17 |
5′-CTCCATGGCTCAGATGTTGC-3′ |
5′-TAGTTGCAGCAGTAAGGCCT-3′ |
| S100P |
5′-AAGGTGCTGATGGAGAAGGA-3′ |
5′-ACTTGTGACAGGCAGACGTG-3′ |
| S100A4 |
5′-GGTGTCCACCTTCCACAAGT-3′ |
5′-GCTGTCCAAGTTGCTCATCA-3′ |
| GAPDH |
5′-CCAGCCGAGCCACATCGCTC-3′ |
5′-ATGAGCCCCAGCCTTCTCCAT-3′ |
Western blotting
At 24 h after cells were treated with PX-12 at the
concentrations of 0, 5 and 10 μM, ~2×106 cells were
homogenized with lysis buffer containing a protease inhibitor
cocktail (P8340; Sigma-Aldrich, St. Louis, MO, USA). The protein
concentration was determined using the BCA protein assay kit (Qcbio
Science & Technologies Co., Ltd., Shanghai, China). Total
proteins (30 μg) were electrophoresed on a 12% SDS-PAGE and
transferred onto PVDF membranes. The membranes were then incubated
with an appropriate dilution of the primary antibody for 1 h
followed by incubation with the horseradish peroxidase-conjugated
secondary antibody (Amersham, Arlington Heights, IL, USA), and
visualized using the enhanced chemiluminescence substrate kit
(Amersham Biosciences, Inc.). The primary antibodies used in the
present study were polyclonal antibodies against S100A4 and actin
(80–50; Abcam, Cambridge, UK).
Statistical analysis
The results in the present study are expressed as
means ± SD. Statistical analysis was performed by the independent
samples t-test (SPSS, Inc., Chicago, IL, USA). A P-value of
<0.05 was considered to indicate a statistically significant
result.
Results
PX-12 inhibits colorectal cancer cell
proliferation and colony formation
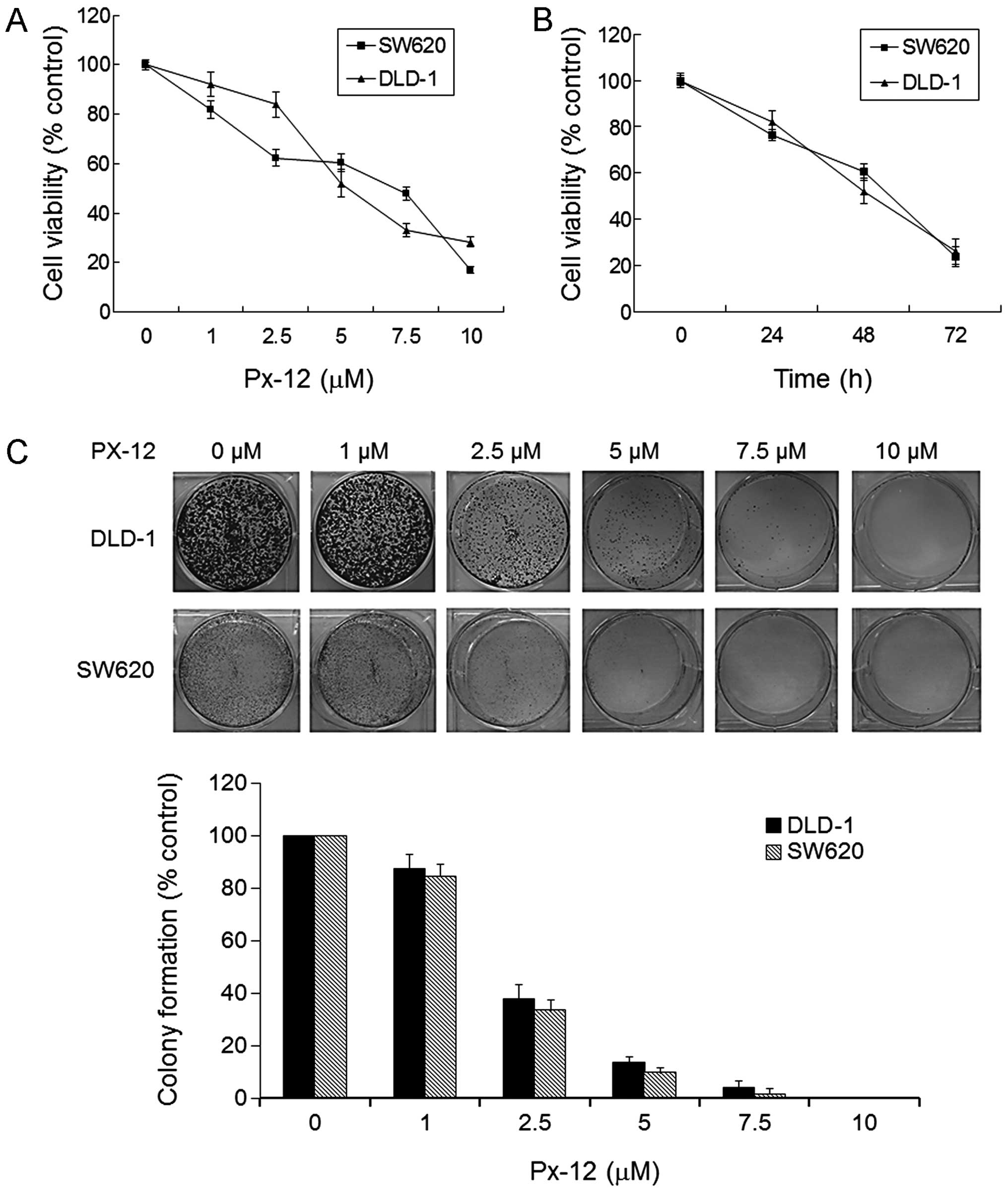
We first examined the effect of PX-12 on the cell
viability of human colorectal cancer cell lines DLD-1 and SW620
using an MTT assay. After exposure to a range of concentrations of
PX-12 (0, 1, 2.5, 5, 7.5, 10 μM) for 48 h, the viability of the
DLD-1 and SW620 cells was reduced in a dose-dependent manner
(Fig. 1A). In addition, a
time-dependent decrease in cell viability was also observed when
cells were treated with 5 μM PX-12 (Fig. 1B). As shown in Fig. 1C, DLD-1 and SW620 cells treated with
the indicated concentrations of PX-12 for 14 days showed a
significant reduction in the colony formation in a dose-dependent
manner.
PX-12 alters the cell cycle distribution
and induces cell apoptosis in colorectal cancer cells
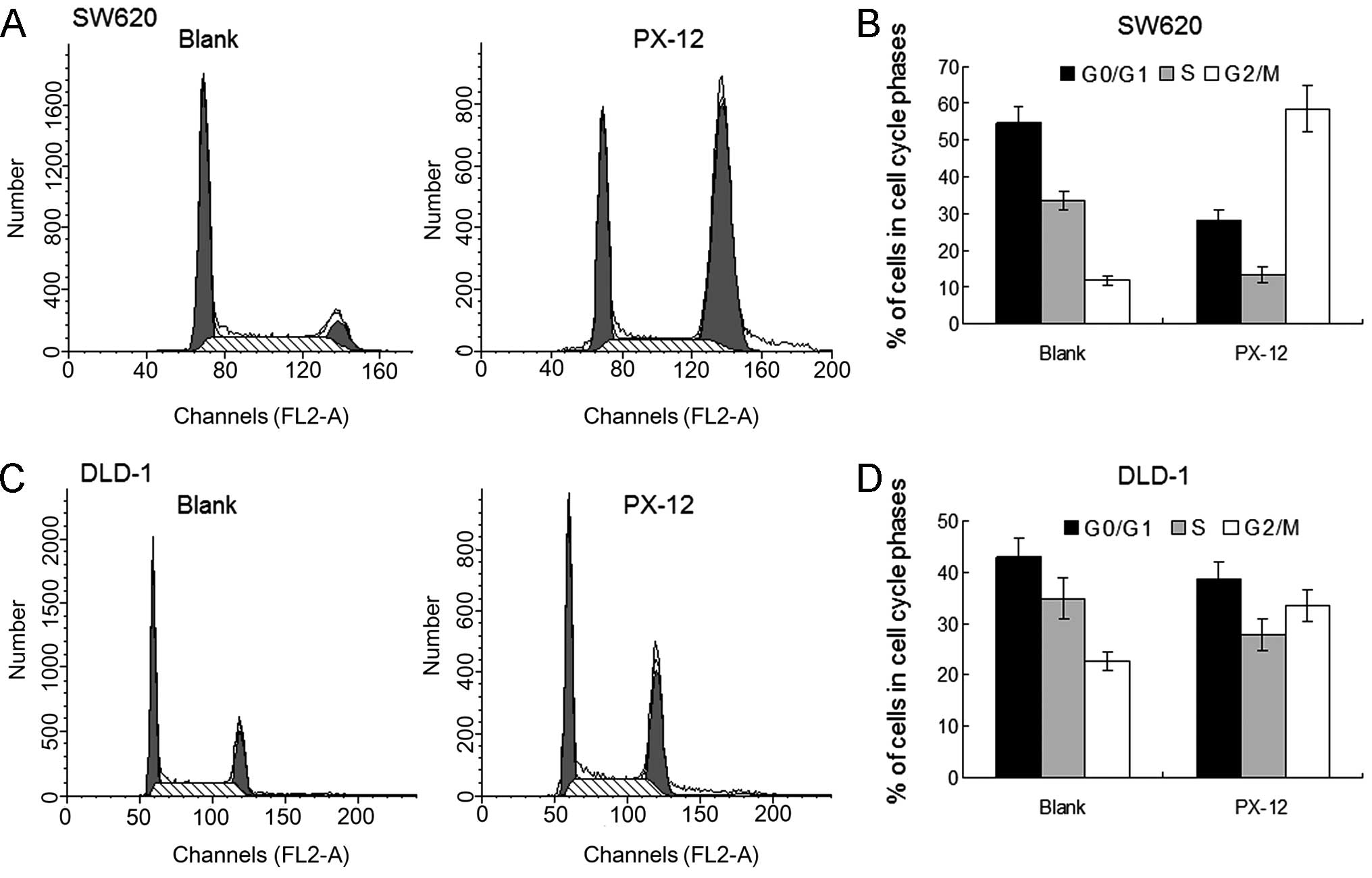
To evaluate the effects of PX-12 on the cell cycle,
cell cycle distributions were determined using flow cytometry.
After the indicated PX-12 treatments for 24 h, PX-12 significantly
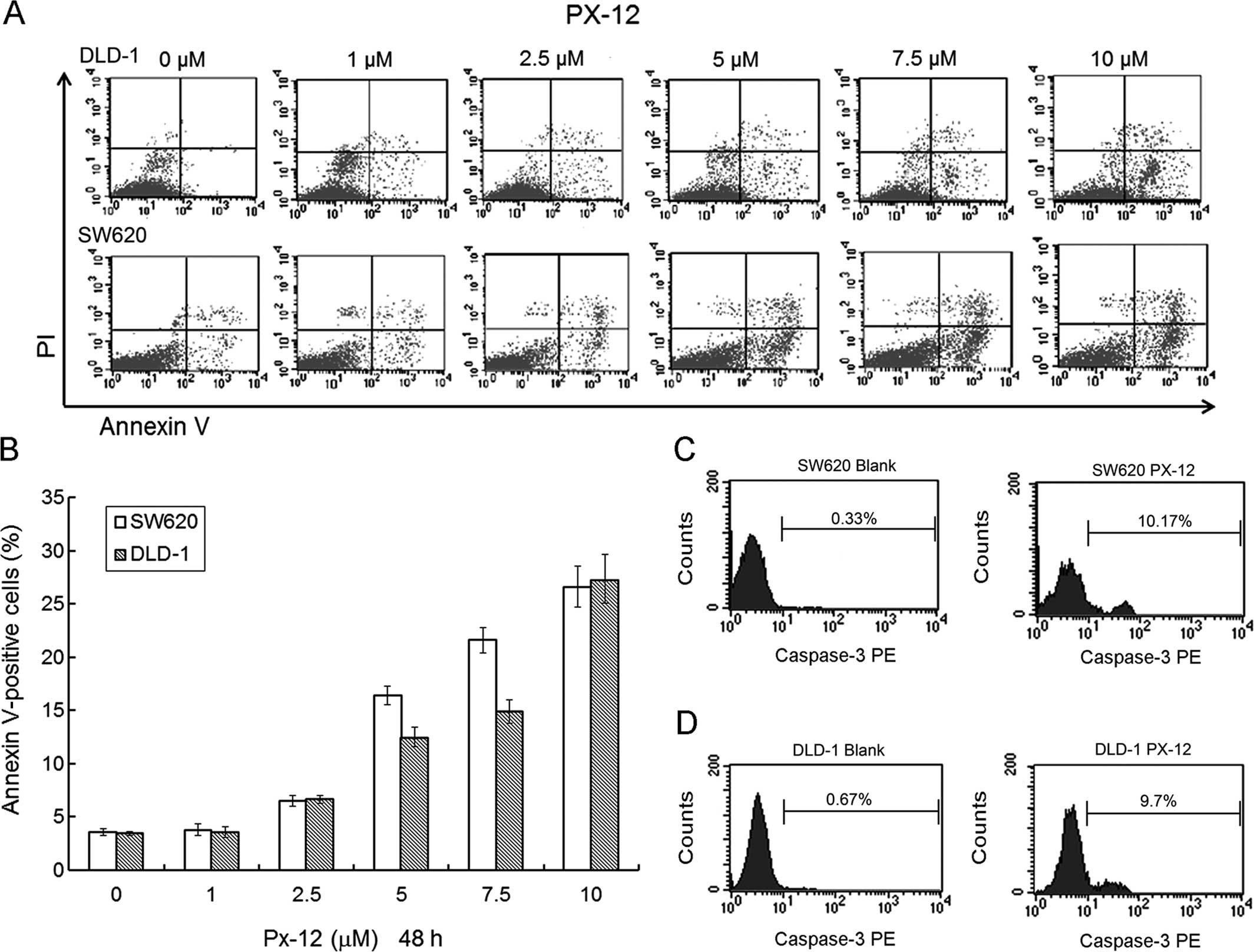
induced G2/M phase arrest of the DLD-1 and SW620 cells (Fig. 2). Cell apoptosis was detected by
Annexin V-FITC/PI staining. We found that PX-12 induced DLD-1 and
SW620 cell apoptosis in a dose-dependent manner (Fig. 3A and B). In additional, PX-12
increased the levels of activated caspase-3 expression in the SW620
and DLD-1 cells at the concentration of 10 μM PX-12 for 48 h
(Fig. 3C and D).
PX-12 inhibits colorectal cancer cell
migration and invasion
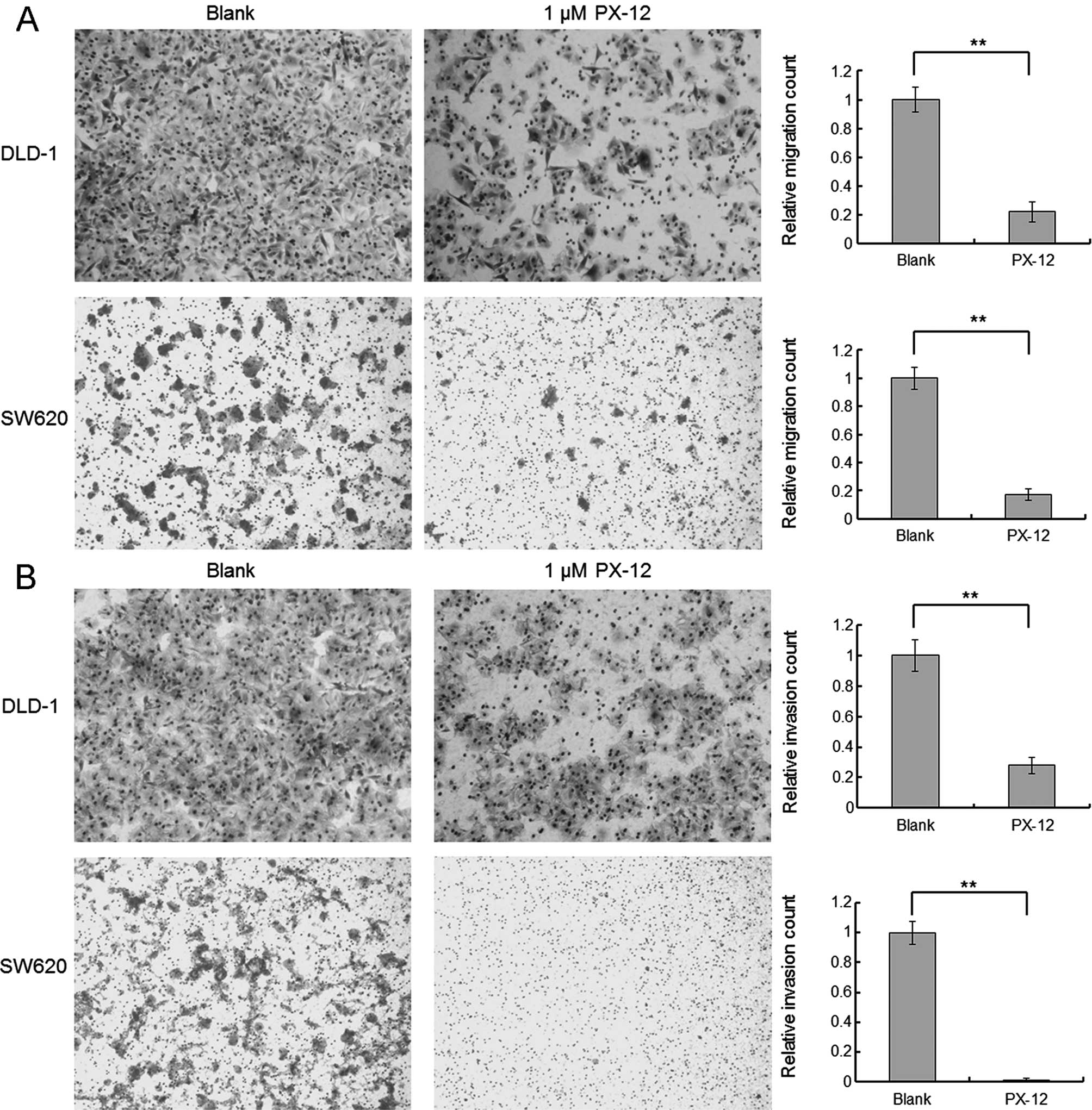
The effects of PX-12 on the migration and invasion
of the colorectal cancer cells were evaluated using Transwell
filter membrane chambers. PX-12 (1 μM), a dose with no significant
effect on the rate of apoptosis, was used in the migration and
invasion study. As shown by the representative images in Fig. 4A and B, the migration and invasion
abilities were inhibited in the PX-12-treated colorectal cancer
DLD-1 and SW620 cells. PX-12 treatment of the DLD-1 and SW620 cells
decreased the cell counts in the migration assay by 78 and 83%
respectively, and in the invasion assay by 72 and 98.6%,
respectively.
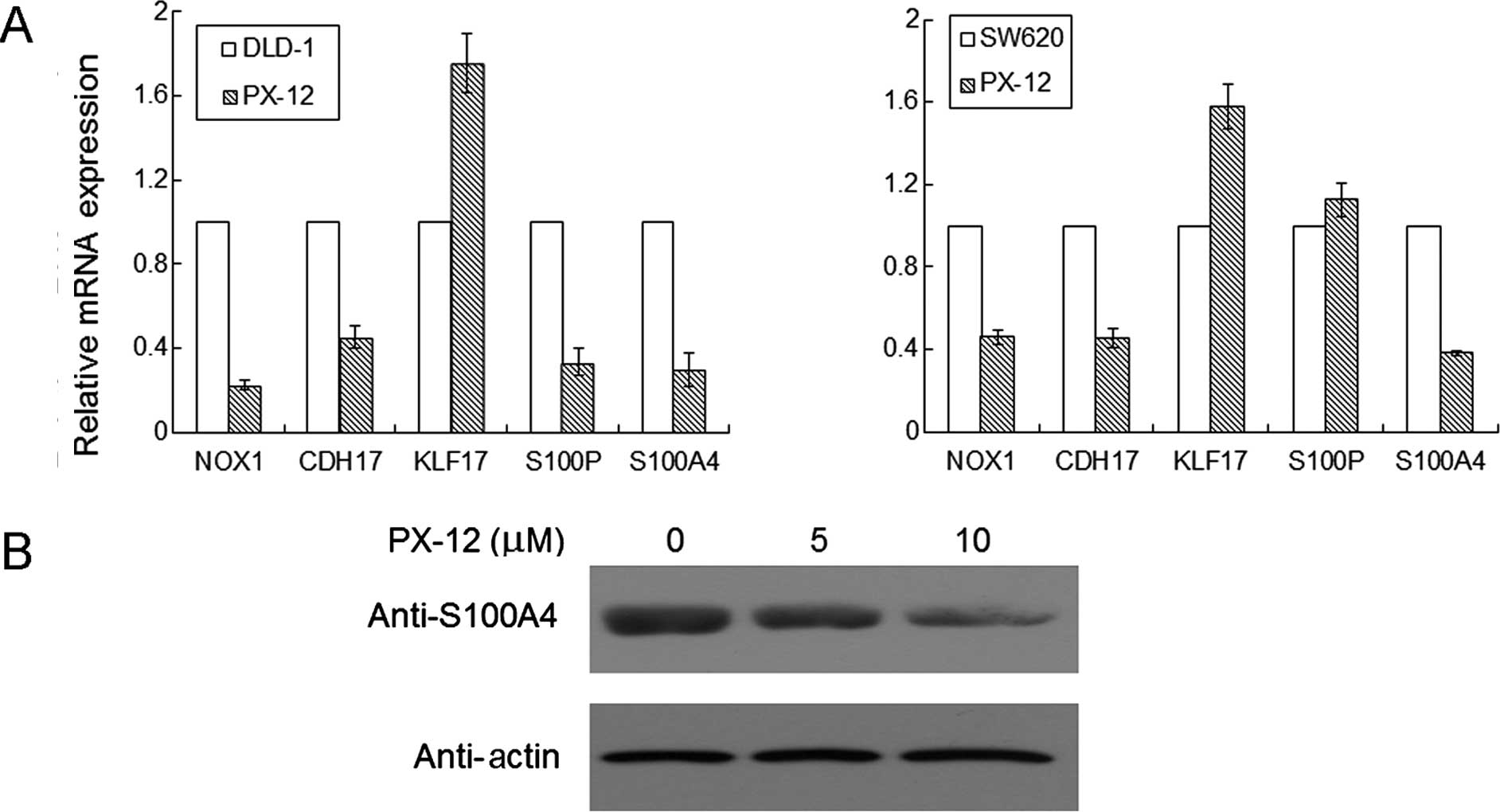
Effects of PX-12 on the NOX1, CDH17,
KLF17, S100P and S100A4 mRNA expression in colorectal cancer
cells
To examine the effect of PX-12 on NOX1, CDH17,
KLF17, S100P and S100A4 mRNA levels in colorectal cancer DLD-1 and
SW620 cells, real-time PCR analysis was performed. As shown in
Fig. 5A, levels of NOX1, CDH17,
S100P and S100A4 mRNA expression were reduced by >50%, and KLF17
mRNA expression was increased after treatment with 2.5 μM PX-12 for
48 h in the DLD-1 cells. In another colorectal cancer cell line
SW620, most of the results were similar as those in the DLD-1 cells
except for S100P mRNA expression which had no significance change
after treatment with PX-12.
PX-12 decreases S100A4 protein expression
in colorectal cancer cells
To examine the effect of PX-12 on S100A4 protein in
colorectal cancer cells, western blot analysis was performed. As
shown in Fig. 5B, the S100A4
protein expression was diminished as the concentration of PX-12
increased in the SW620 cells.
Discussion
The differentiation and proliferation of colorectal
cancer cells are complex events and the relevant mechanisms have
been universally investigated (21–23).
Trx-1 expression was reported to be increased in several types of
human cancers, including colorectal carcinoma (10,24).
Trx-1 has various biological activities, such as antioxidant,
growth control, anti-apoptotic properties and inflammation
regulation that provide reducing equivalents and a transcriptional
regulator (6,25,26).
PX-12, an inhibitor of Trx-1, has shown excellent in vitro
antitumor activity, and Ramanathan et al reported the
first-in-human phase I trial of PX-12 in patients with advanced
solid tumors (20). However, the
antitumor effect of PX-12 in colorectal cancer cells, particularly
the effect of PX-12 on cancer cell migration and invasion, is still
obscure.
The aim of the present study was to assess the
effects of PX-12 on colorectal cancer cells in vitro.
Following exposure to a range of concentrations of PX-12 (0, 1,
2.5, 5, 7.5 and 10 μM) for 48 h, cell growth was inhibited as the
concentration of PX-12 increased. The same result was shown when
the exposure time was prolonged. We also demonstrated that PX-12
inhibited colorectal cancer cell colony formation. The
susceptibility of colorectal cancer cells to PX-12 appeared to
significantly increase after the incubation time was prolonged or
the concentration of PX-12 was increased. Namely the inhibitory
effects of PX-12 on colorectal cancer cells were dose- and
time-dependent. DNA flow cytometric analysis indicated that PX-12
significantly induced G2/M phase arrest of the cell cycle in
colorectal cancer DLD-1 and SW620 cells. Similarly, PX-12 induced
G2/M phase arrest in breast cancer and lung cancer cells (27,28).
Therefore, this body of evidence suggests that the G2/M phase
arrest in PX-12-treated cells is an underlying mechanism to
suppress the growth of cancer cells, including colorectal cancer
cells.
In the apoptosis assay, PX-12 also increased the
number of Annexin V-positive cells at the designated concentrations
of PX-12 for 48 h, suggesting that PX-12-induced colorectal cancer
cell death occurred via apoptosis. Our results indicated that PX-12
induced apoptosis in human colorectal cancer cells through
enhancement of caspase-3 activation. In additional, the migration
and invasion abilities were inhibited in the PX-12-treated
colorectal cancer cells. To further elucidate the molecular
mechanisms of the antitumor effects of PX-12, we performed gene
array studies and comparative gene profiling analysis of DLD-1
cells treated with PX-12 (data not shown). NOX1, CDH17, KLF17,
S100P and S100A4 were found to be regulated by PX-12 in the DLD-1
cells. NOX1, an NADPH oxidase, which is known to enhance reactive
oxygen species (ROS), is expressed in all types of tissues
including osteoclasts, vascular smooth muscle, and normal gastric
and colonic mucosa (29); there is
growing evidence that Nox1 is overexpressed in many human colonic
adenocarcinomas (30). CDH17 was
found to increase cell adhesion and proliferation in colon cancer
cells, by regulating α2β1 integrin signaling to induce particular
focal adhesion kinase and Ras activation (31). KLF17 plays vital roles in many
oncogenic processes, such as tumor growth and metastasis. Studies
have reported that KLF17 acts as an inhibitor of
epithelial-to-mesenchymal transition and metastasis in breast
cancer (32). More and more
evidence suggests that S100P plays a critical role in tumors. This
gene was found to been expressed in several different types of
cancer, including colon, pancreas, breast and lung cancer (33–36).
Recent studies have also shown that S100A4, a small Ca2+
binding protein, is highly expressed in different tumor tissues
(37). High S100A4 expression is
related to increased metastasis formation in tumors (38), including colorectal cancer (39,40).
We further confirmed a change in the expression of genes by
quantitative RT-PCR. After exposure of DLD-1 and SW620 cells to
PX-12 for 48 h, NOX1, CDH17 and S100A4 mRNA expression was reduced,
while KLF17 mRNA expression was increased. This indicated that
PX-12 may trigger the inhibition of NOX1, CDH17 and S100A4 mRNA
expression and an increase in KLF17 mRNA expression. S100A4 is
overexpressed in a range of different tumor types, and plays an
important role in the process of cancer metastasis (38,41).
S100A4 expression is significantly associated with positive lymph
node metastasis in colorectal cancer (39), and targeting the expression of
S100A4 with calcimycin was found to inhibit colon cancer cell
migration and invasion (42,43).
As shown in Fig. 5B, the S100A4
protein expression was diminished in the PX-12-treated SW620 cells.
The mechanism involved in the decrease in S100A4 protein by PX-12
is not known. It is known that PX-12 is an inhibitor of Trx-1 and
has antitumor activity. One may predict that inhibition of the
Trx-1 redox system leads to decreased S100A4 protein and
trans-activation of downstream targets. Therefore, future research
is required to investigate the correlation of the antitumor effect
of PX-12 on colorectal cancer cells with NOX1, CDH17, S100A4, S100P
and KLF17 expression.
In summary, we demonstrated that PX-12 inhibited the
growth of colorectal cancer cells via G2/M phase arrest and
induction of apoptosis. Notably, a low dose of PX-12 inhibited
colorectal cancer cell migration and invasion. PX-12 has been shown
to have significant antitumor activity in colorectal cancer in
vitro. This antitumor effect may be associated with NOX1,
CDH17, S100A4 and KLF17 mRNA expression. The present study provides
important insight into the antitumor effects of PX-12 on colorectal
cancer cells and represents a novel and effective strategy for
treating cancer.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (81101823), the Medicine and Health
Technology Program of Zhejiang Province (2013KYA129), and the
Wenzhou Science and Technology Bureau Program (Y20120181).
References
|
1
|
Khuhaprema T and Srivatanakul P: Colon and
rectum cancer in Thailand: an overview. Jpn J Clin Oncol.
38:237–243. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Moertel CG, Fleming TR, Macdonald JS, et
al: Fluorouracil plus levamisole as effective adjuvant therapy
after resection of stage III colon carcinoma: a final report. Ann
Intern Med. 122:321–326. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sargent D, Sobrero A, Grothey A, et al:
Evidence for cure by adjuvant therapy in colon cancer: observations
based on individual patient data from 20,898 patients on 18
randomized trials. J Clin Oncol. 27:872–877. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chau I and Cunningham D: Adjuvant therapy
in colon cancer - what, when and how? Ann Oncol. 17:1347–1359.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chung-Faye GA and Kerr DJ: ABC of
colorectal cancer: innovative treatment for colon cancer. BMJ.
321:1397–1399. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Powis G and Montfort WR: Properties and
biological activities of thioredoxins. Annu Rev Biophys Biomol
Struct. 30:421–455. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lim JY, Yoon SO, Hong SW, Kim JW, Choi SH
and Cho JY: Thioredoxin and thioredoxin-interacting protein as
prognostic markers for gastric cancer recurrence. World J
Gastroenterol. 18:5581–5588. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Powis G and Montfort WR: Properties and
biological activities of thioredoxins. Annu Rev Pharmacol Toxicol.
41:261–295. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nakamura H, Bai J, Nishinaka Y, et al:
Expression of thioredoxin and glutaredoxin, redox-regulating
proteins, in pancreatic cancer. Cancer Detect Prev. 24:53–60.
2000.PubMed/NCBI
|
|
10
|
Raffel J, Bhattacharyya AK, Gallegos A, et
al: Increased expression of thioredoxin-1 in human colorectal
cancer is associated with decreased patient survival. J Lab Clin
Med. 142:46–51. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ueno M, Masutani H, Arai RJ, et al:
Thioredoxin-dependent redox regulation of p53-mediated p21
activation. J Biol Chem. 274:35809–35815. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Welsh SJ, Bellamy WT, Briehl MM and Powis
G: The redox protein thioredoxin-1 (Trx-1) increases
hypoxia-inducible factor 1α protein expression: Trx-1
overexpression results in increased vascular endothelial growth
factor production and enhanced tumor angiogenesis. Cancer Res.
62:5089–5095. 2002.PubMed/NCBI
|
|
13
|
Abate C, Patel L, Rauscher FJ III and
Curran T: Redox regulation of fos and jun DNA-binding activity in
vitro. Science. 249:1157–1161. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kakolyris S, Giatromanolaki A, Koukourakis
M, et al: Thioredoxin expression is associated with lymph node
status and prognosis in early operable non-small cell lung cancer.
Clin Cancer Res. 7:3087–3091. 2001.PubMed/NCBI
|
|
15
|
Grogan TM, Fenoglio-Prieser C, Zeheb R, et
al: Thioredoxin, a putative oncogene product, is overexpressed in
gastric carcinoma and associated with increased proliferation and
increased cell survival. Hum Pathol. 31:475–481. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jordan BF, Runquist M, Raghunand N, et al:
The thioredoxin-1 inhibitor 1-methylpropyl 2-imidazolyl disulfide
(PX-12) decreases vascular permeability in tumor xenografts
monitored by dynamic contrast enhanced magnetic resonance imaging.
Clin Cancer Res. 11:529–536. 2005.PubMed/NCBI
|
|
17
|
Gallegos A, Gasdaska JR, Taylor CW, et al:
Transfection with human thioredoxin increases cell proliferation
and a dominant-negative mutant thioredoxin reverses the transformed
phenotype of human breast cancer cells. Cancer Res. 56:5765–5770.
1996.PubMed/NCBI
|
|
18
|
Wipf P, Hopkins TD, Jung JK, et al: New
inhibitors of the thioredoxin-thioredoxin reductase system based on
a naphthoquinone spiroketal natural product lead. Bioorg Med Chem
Lett. 11:2637–2641. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kirkpatrick DL, Kuperus M, Dowdeswell M,
et al: Mechanisms of inhibition of the thioredoxin growth factor
system by antitumor 2-imidazolyl disulfides. Biochem Pharmacol.
55:987–994. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ramanathan RK, Kirkpatrick DL, Belani CP,
et al: A phase I pharmacokinetic and pharmacodynamic study of
PX-12, a novel inhibitor of thioredoxin-1, in patients with
advanced solid tumors. Clin Cancer Res. 13:2109–2114. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Uchiyama S, Itoh H, Naganuma S, et al:
Enhanced expression of hepatocyte growth factor activator inhibitor
type 2-related small peptide at the invasive front of colon
cancers. Gut. 56:215–226. 2007. View Article : Google Scholar
|
|
22
|
Yamazaki K, Shimizu M, Okuno M, et al:
Synergistic effects of RXRα and PPARγ ligands to inhibit growth in
human colon cancer cells - phosphorylated RXRα is a critical target
for colon cancer management. Gut. 56:1557–1563. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shen D, Deng C and Zhang M: Peroxisome
proliferator-activated receptor γ agonists inhibit the
proliferation and invasion of human colon cancer cells. Postgrad
Med J. 83:414–419. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Noike T, Miwa S, Soeda J, Kobayashi A and
Miyagawa S: Increased expression of thioredoxin-1, vascular
endothelial growth factor, and redox factor-1 is associated with
poor prognosis in patients with liver metastasis from colorectal
cancer. Hum Pathol. 39:201–208. 2008. View Article : Google Scholar
|
|
25
|
Lillig CH and Holmgren A: Thioredoxin and
related molecules - from biology to health and disease. Antioxid
Redox Signal. 9:25–47. 2007. View Article : Google Scholar
|
|
26
|
Arnér ES: Focus on mammalian thioredoxin
reductases - important selenoproteins with versatile functions.
Biochim Biophys Acta. 1790:495–526. 2009. View Article : Google Scholar
|
|
27
|
Vogt A, Tamura K, Watson S and Lazo JS:
Antitumor imidazolyl disulfide IV-2 causes irreversible
G2/M cell cycle arrest without hyperphosphorylation of
cyclin-dependent kinase Cdk1. J Pharmacol Exp Ther. 294:1070–1075.
2000.PubMed/NCBI
|
|
28
|
Shin HR, You BR and Park WH: PX-12-induced
HeLa cell death is associated with oxidative stress and GSH
depletion. Oncol Lett. 6:1804–1810. 2013.PubMed/NCBI
|
|
29
|
Krause KH: Tissue distribution and
putative physiological function of NOX family NADPH oxidases. Jpn J
Infect Dis. 57:S28–S29. 2004.PubMed/NCBI
|
|
30
|
Juhasz A, Ge Y, Markel S, et al:
Expression of NADPH oxidase homologues and accessory genes in human
cancer cell lines, tumours and adjacent normal tissues. Free Radic
Res. 43:523–532. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bartolomé RA, Barderas R, Torres S, et al:
Cadherin-17 interacts with α2β1 integrin to regulate cell
proliferation and adhesion in colorectal cancer cells causing liver
metastasis. Oncogene. 33:1658–1669. 2014. View Article : Google Scholar
|
|
32
|
Gumireddy K, Li A, Gimotty PA, et al:
KLF17 is a negative regulator of epithelial-mesenchymal transition
and metastasis in breast cancer. Nat Cell Biol. 11:1297–1304. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fuentes MK, Nigavekar SS, Arumugam T, et
al: RAGE activation by S100P in colon cancer stimulates growth,
migration, and cell signaling pathways. Dis Colon Rectum.
50:1230–1240. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Logsdon CD, Simeone DM, Binkley C, et al:
Molecular profiling of pancreatic adenocarcinoma and chronic
pancreatitis identifies multiple genes differentially regulated in
pancreatic cancer. Cancer Res. 63:2649–2657. 2003.PubMed/NCBI
|
|
35
|
Guerreiro Da Silva ID, Hu YF, Russo IH, et
al: S100P calcium-binding protein overexpression is associated with
immortalization of human breast epithelial cells in vitro and early
stages of breast cancer development in vivo. Int J Oncol.
16:231–240. 2000.PubMed/NCBI
|
|
36
|
Diederichs S, Bulk E, Steffen B, et al:
S100 family members and trypsinogens are predictors of distant
metastasis and survival in early-stage non-small cell lung cancer.
Cancer Res. 64:5564–5569. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mishra SK, Siddique HR and Saleem M:
S100A4 calcium-binding protein is key player in tumor progression
and metastasis: preclinical and clinical evidence. Cancer
Metastasis Rev. 31:163–172. 2012. View Article : Google Scholar
|
|
38
|
Boye K and Maelandsmo GM: SS100A4 and
metastasis: a small actor playing many roles. Am J Pathol.
176:528–535. 2010. View Article : Google Scholar :
|
|
39
|
Huang LY, Xu Y, Cai GX, et al: S100A4
over-expression underlies lymph node metastasis and poor prognosis
in colorectal cancer. World J Gastroenterol. 17:69–78. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Stein U, Arlt F, Walther W, et al: The
metastasis-associated gene S100A4 is a novel target of
β-catenin/T-cell factor signaling in colon cancer.
Gastroenterology. 131:1486–1500. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Orre LM, Panizza E, Kaminskyy VO, et al:
S100A4 interacts with p53 in the nucleus and promotes p53
degradation. Oncogene. 32:5531–5540. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sack U, Walther W, Scudiero D, et al:
S100A4-induced cell motility and metastasis is restricted by the
Wnt/β-catenin pathway inhibitor calcimycin in colon cancer cells.
Mol Biol Cell. 22:3344–3354. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jiang L, Lai YK, Zhang J, et al: Targeting
S100P inhibits colon cancer growth and metastasis by
lentivirus-mediated RNA interference and proteomic analysis. Mol
Med. 17:709–716. 2011. View Article : Google Scholar : PubMed/NCBI
|