Introduction
Lung cancer is one of the most common neoplasms
worldwide, and approximately 85–90% of lung cancer cases are
non-small cell lung cancer (NSCLC) (1). Despite the fact that chemotherapy and
radiotherapy have been widely used in the treatment of advanced
NSCLC, the outcome remains poor, with less than 15% of patients
surviving beyond 5 years due to the lack of early diagnosis and
effective treatment methods (2,3).
Therefore, it is urgent to identify new therapeutics to combat this
disease.
With the recent advances in the understanding of the
molecular pathways involved in NSCLC progression, targeted
therapies that are designed to interfere with the mechanisms of
cancer cell growth and survival offer new hope in NSCLC
therapeutics. X-linked inhibitor of apoptosis protein (XIAP) has
been identified as one of the most potent inhibitors of caspases
and apoptosis to date and is known to play an important role in
cell apoptosis, cell migration and cell invasion (4,5). It is
highly expressed in various malignancies, while its expression is
very low or absent in normal cells, which makes it an attractive
target for cancer therapeutics (6–8). A
large number of studies have demonstrated that inhibition of XIAP
expression using antisense oligonucleotides or small interfering
RNA (siRNA) could suppress the proliferation of tumor cells, induce
cell apoptosis, and sensitize tumor cells to chemotherapeutic
agents (9–14). In addition, Harlin et al
found that XIAP-knockout mice have normal survival with no
significant pathological features, consistent with XIAP-targeted
therapeutics exerting minimal toxicity to normal tissues (15). Therefore, in the present study, we
explored the possibility of XIAP as a therapeutic target for the
treatment of human NSCLC.
Celecoxib (CXB), a selective cyclooxygenase-2
(COX-2) inhibitor, has been used in the treatment of a wide range
of cancers including NSCLC (2,16).
Previous studies have shown that the antitumor mechanisms of CXB
include different pathways, such as the death receptor,
mitochondria-mediated pathways, Akt phosphorylation inhibition,
endoplasmic reticulum (ER) stress, and autophagy (17–21).
Despite its excellent antitumor activity, CXB has a relatively low
therapeutic index, and its clinical usage is limited due to acute
and chronic toxicities such as cardiovascular thrombosis,
congestive heart failure, gastrointestinal ulceration, renal or
hepatic injury and platelet aggregation (22). In addition, resistance to this agent
is common, representing a major obstacle to successful treatment.
The toxicity of doxorubicin can be reduced if it is used in
conjunction with another, more tumor-specific treatment, in order
to reduce the dosage. Currently, CXB is widely being tested in
clinical trials for its therapeutic activity against various
cancers in combination with other agents (23–25).
For example, Morisaki et al found that the combination of
sorafenib (SOR) and CXB has synergistic anti-proliferative and
pro-apoptotic effects on human liver cancer cells (25). Our recent study also showed that CXB
in combination with SOR suppressed VEGFR and COX-2 expression and
inhibited the tumor growth of NSCLC (3). Huang et
al demonstrated that downregulation of glucose-regulated
protein (GRP) 78 potentiated by RNAi enhanced CXB-induced cell
apoptosis in human urothelial carcinoma cells (26). These findings suggest that CXB in
combination with other agents could allow a decrease in the dose of
the drug, and increase antitumor activity.
In the present study, we aimed to investigate the
efficacy of a combination of XIAP-targeted short hairpin RNA
(XIAP-shRNA) and CXB in the treatment of NSCLC in vitro and
in vivo on the basis of the hypothesis that this combination
exhibits a synergistic anticancer effect compared with either
treatment alone. We found that this combination treatment led to
high antitumor activity in vitro and in vivo. This
combination is a promising drug candidate for the treatment of
NSCLC.
Materials and methods
Reagents and antibodies
CXB, one type of COX-2 inhibitor, was purchased from
Pfizer Corporation Inc. (New York, NY, USA) and was dissolved in
dimethyl sulfoxide (DMSO).
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
was obtained from Sigma Aldrich (St. Louis, MO, USA), and was
prepared by dissolving 1 mg of each compound in 1 ml of
phosphate-buffered saline (PBS, pH 7.2). For western blot analysis,
the following antibodies were used: mouse monoclonal anti-β-actin
(Sigma Aldrich), mouse monoclonal anti-Bcl-2, mouse monoclonal
anti-survivin, mouse monoclonal anti-PI3K, mouse monoclonal
anti-phosphorylated (p)-PI3K, mouse monoclonal anti-AKT, mouse
monoclonal anti-p-AKT and anti-mouse IgG were purchased from Santa
Cruz Biotechnology (Santa Cruz, CA, USA).
shRNA design and plasmid
construction
The cDNA sequence of XIAP was obtained from GenBank
(accession number NM-001167). The siRNA target design tools from
Ambion were used to design XIAP-shRNA, and non-specific shRNA
sequences were designed according to the siRNA target design tools
from Ambion (Austin, TX, USA). XIAP-shRNA and non-specific shRNA
were designed and synthesized as follows: XIAP-shRNA sense, 5′-GAT
CCGGTGAAGGTGATAAAGTAATCAAGAGATACAAA TTCTTCTTCCTTAAGA-3′ and
antisense, 5′-AGCTTCT GATCCGGTGAAGGTGATAAAGTAACTCTTGAATACA
AATTCTTCCTTAG-3′; control shRNA sense,
5′-GATCCTTCTCCGAACGTGTCACGTTCAAGAGAAAACTACTTC TTTTACCTTAGA-3′ and
antisense, 5′-AGCTTCTTCTTT CTCCGAACGTGTCACGTCTCTTGAAAAACTACTTCT
TTTACCTTG-3′; the sequence does not target any gene product and has
no significant sequence similarity to human gene sequences. The
oligonucleotides were annealed and then ligated into the
BamHI and HindIII sites of pSilencer 4.1-CMV neo
expression vector according to the manufacturer’s instructions
(Ambion). The recombinant vectors were confirmed by the digestion
analysis of restriction endonuclease, and all inserted sequences
were verified by DNA sequencing. Each vector contained the SV40
early promoter to provide G418 resistance in mammalian cells. The
shRNA insert template oligonucleotides encoding XIAP are referred
to as XIAP-shRNA, and the plasmid encoding non-specific siRNA,
which was used as a negative control, is referred to as
NC-shRNA.
Cell culture and transfection
The human NSCLC cell line A549 was purchased from
the Cell Bank of the Type Culture Collection of the Chinese Academy
of Sciences, Shanghai Institute of Cell Biology, Chinese Academy of
Sciences (Shanghai, China). A549 cells were cultured in RPMI-1640
medium (Invitrogen, Carlsbad, CA, USA) supplemented with
heat-inactivated 10% fetal bovine serum (FBS; Biochrom AG) at 37°C
in a humidified atmosphere containing 5% CO2.
A549 cells were seeded in 6-well plates at
2.0×104 cells/well, respectively and cultured overnight
to 80% confluency prior to transfection. Transfection was performed
using Lipofectamine Plus (Grand Island, NY, USA), and the ratio of
the plasmids and the transfection reagent was 1 mg:2 ml. Cells were
transfected with the different plasmids (XIAP-shRNA, NC-shRNA)
according to the protocol specifications. G418 (800 μg/ml, Sigma
Aldrich) was used to screen stably transfected clones. Stable
transfectants were named A549/XIAP-shRNA and A549/NC-shRNA.
Quantification by real-time polymerase
chain reaction
Total cellular RNA was isolated using TRIzol reagent
according to the manufacturer’s protocol (Invitrogen). M-MLV
reverse transcriptase (Fermentas MBI, Amherst, NY, USA) was used to
create cDNA following the manufacturer’s instructions. Quantitative
real-time polymerase chain reaction (RT-PCR) assays were carried
out using SYBR-Taq real-time kits (Takara Biotechnology, Otsu,
Japan) and RT-PCR amplification equipment ABI Prism 7900HT (Applied
Biosystems, Foster City, CA, USA). The PCR primers used to detect
XIAP and GAPDH were as follows: XIAP forward, 5′-GACAGTATGCAAGAT
GAGTCAAGTCA-3′ and reverse, 5′-GCAAAGCTTCTCCTC TTGCAG-3′, with a
product length of 93 bp; GAPDH forward, 5′-TGTGGGCATCAATGGATTTGG-3′
and reverse, 5′-ACA CCATGTATTCCGGGTCAAT-3′, with a product length
of 171 bp. The PCR conditions were as follows: pre-denaturation at
94°C for 5 min, followed by 40 cycles of denaturation at 94°C for
10 sec, annealing/extension at 58°C for 15 sec and a final
extension at 72°C for 10 min. The amplification specificity was
checked by melting curve analysis. The expression of the genes of
interest was determined by normalization of the threshold cycle
(Ct) of these genes to that of the control GAPDH. The
2−ΔΔCt method was used to calculate the relative
abundance of target gene expression generated using Rotor-Gene
Real-Time analysis software 6.1.81.
Western blot analysis
The A549 cells were then homogenized in a lysis
buffer (Tris-HCl 50 mmol/l, EDTA 5 mmol/l, NaCl 150 mmol/l, sodium
deoxycholate 1%, Na3VO4 500 μmol/l, Triton
X-100 0.5%, AEBSF 10 μmol/l, NaF 10 mmol/l) on ice. The homogenates
were then centrifuged at 14,000 rpm at 4°C for 30 min, and the
supernatants were collected for protein concentration determination
using the BCA assay kit (Sigma Aldrich). Cell extracts (50 μg of
protein) were separated on a sodium dodecyl sulfate-polyacrylamide
electrophoretic gel (SDS-PAGE) and transferred to nitrocellulose
membranes, which were blocked in 3% bovine serum albumin (BSA) for
2 h. After blocking, the membranes were incubated with the primary
antibodies overnight at 4°C for 2 h, and subseqnently with the
horseradish peroxidase-conjugated secondary antibody for 2 h at
room temperature. Protein bands were visualized with enhanced
chimioluminescence reagent (ECL; Amersham, GE Healthcare,
Velizy-Villacoublay, France). Densitometry was performed by
Quantity One image analysis software.
Cell viability assay
To measure the effect of shRNA-XIAP in combination
with CXB on cell proliferation, an MTT assay was used. The cell
density of A549 cells was adjusted to 5×104/ml, and
cells were added to a 96-well plate (100 μl/well). In the blank
controls, 100 μl of medium alone was added. At 24 h after culture,
cells were treated with the indicated concentrations of CXB and
transfected with the XIAP-shRNA plasmid, or both. Cells were
divided into five groups: normal control, negative control
(NC-shRNA), XIAP-shRNA group, CXB group and XIAP-shRNA combined
with CXB group. There were 8 wells in each group/subgroup. At 48 h
after culture, 200 μl of MTT (5 mg/ml) was added to each well
followed by incubation at 37°C for 4 h. Then, centrifugation was
performed at 2,000 × g for 10 min. The supernatant was removed, and
200 μl of DMSO was added to each well followed by shaking for 10
min. Absorbance was measured at 490 nm with a microplate reader
(Molecular Devices Corp., Sunnyvale, CA, USA), and growth
inhibition was calculated. The mean proliferation of cells without
any treatment was expressed as 100%. All experiments were performed
in triplicate and repeated at least three times.
TUNEL assay
To measure the effect of XIAP-shRNA in combination
with CXB on cell apoptosis, TUNEL assay was carried out. In
briefly, cellular DNA fragmentation was measured with the ApoTag
Red In Situ Apoptosis Detection kit (Chemicon Inc.,
Temecula, CA, USA) according to the manufacturer’s instructions
when A549 cells were treated with XIAP-shRNA, CXB alone or both for
24 h. To quantify the apoptotic cells, terminal deoxynucleotidyl
transferase-mediated nick end labeling (TUNEL)-positive cells were
counted using confocal microscopy (Olympus, Tokyo, Japan).
In addition, at the molecular level, we also
detected other anti-apoptotic molecules, such as survivin, and
Bcl-2 protein expression by western blotting as an additional
indicator of apoptosis.
Caspase activity
The activities of caspase-3 and -9 were measured
using Caspase Colorimetric Protease Assay kits (Millipore Corp.,
Billerica, MA, USA) according to the manufacturer’s instructions.
In brief, A549 cells were treated with the indicated plasmid for 24
h, and washed twice with ice-cold PBS (pH 7.2) and harvested by
centrifugation. The cell pellets were then lysed in 150 μl buffer
provided in the kit (Millipore). An aliquot of lysates (80 μl) was
incubated with 10 μl substrate of each caspase at 37°C for 2 h.
Samples were analyzed at 405 nm in a microplate reader (Thermo
Fisher Scientific Inc., Waltham, MA, USA). The relative caspase
activity of the control group was taken as 100. Each assay was
conducted in triplicate.
Wound healing assay
To assess the effect of XIAP-shRNA in combination
with CXB on cell migration, a wound-healing assay was performed. In
brief, 2×105 A549 cells were plated in 6-well plates in
RMP-1640 medium containing 10% FBS. After 24 h, a scratch was made
through the confluent cell monolayer, and the cells were treated
with the indicated agent, respectively, in 3 ml of complete medium.
After a 48-h treatment, cells were stained with hematoxylin and
eosin (H&E). Cells invading the wound line were observed under
an inverted phase-contrast microscope (Leica DMR, Germany).
Triplicates were repeated in all experiments.
Matrigelinvasion assay
The migration capacity of NSCLC cells was determined
in vitro using Transwell chambers (Corning, Tewksbury, MA,
USA) in which the two chambers were separated by a Matrigel-coated
polycarbonate membrane (8-μm pore size). Cells were treated with
CXB, XIAP-shRNA, or the combination of the two agents for 24 h in
FBS-free RPMI-1640 medium. The lower chamber was then filled with
500 ml of RPMI-1640 medium supplemented with 10% FBS and
5×105 cells/ml were placed in the upper chamber in a
total volume of 200 ml. The Transwell chamber was then assembled,
and the cells were incubated for 24 h at 37°C in a 5%
CO2 environment. After a 24-h incubation, cells invading
the bottom surface of the filter were fixed and stained with 0.1%
crystal violet in 20% methanol. Invasiveness was determined by
counting the penetrating cells under a Nikon phase-contrast
microscope and counted in >10 fields of view at ×200
magnification. The invasion of cells without any treatment was
determined as 100%. Each cell migration experiment was repeated
three times.
In addition, we also detected MMP-9 and MMP-2
protein expression by western blotting as an additional indicator
of invasion and migration.
Measurement of prostaglandin-E2 (PGE2)
production and VEGF expression
Prostaglandin-E2 (PGE2) production was determined by
competitive enzyme-linked immunosorbent assay (ELISA). In brief,
A549 cells were treated with XIAP-shRNA, CXB, alone or both for 48
h in 12-well plates, and then the culture media were centrifuged to
remove the cell debris. Cell-free culture media were collected at
the indicated times. Then PGE2 levels were measured by using Human
Prostaglandin E2 ELISA kits (Kemin, Shanghai, China) according to
the manufacturer’s instructions.
A549 cells were treated with XIAP-shRNA, CXB or both
for 48 h in 24-well plates, and then the culture media were
centrifuged to remove cell debris. Cell-free culture media were
collected at the indicated times. The VEGF level in the cell
supernatant was determined by using the Human VEGF ELISA kit
(Yanyu, Shanghai, China) according to the manufacturer’s
instructions.
Tumor xenograft experiments
Specific pathogen-free (SPF) nude mice (BALB/c) used
in the experiments were provided by the Center for Experimental
Animals at Jilin University (Jilin, China). All experiments
involving animals were performed in accordance with institutional
guidelines, following a protocol approved by the Ethics Committees
of the Disease Model Research Center, The First Hospital of Jilin
University. Forty female BALB mice (aged 6–7 weeks) were maintained
under specific pathogen-free conditions and provided with food and
water ad libitum.
Stable A549/XIAP-shRNA clones were harvested, and a
tumorigenic dose of 2.5×106 cells was injected
intraperitoneally in 20 mice. The remaining 20 mice were injected
with untreated A549 cells as the control group. When progression
was evident in ~28 days, 10 mice from each group were randomly
selected and were treated with CXB (4.56 mg/kg body weight)
intraperitoneally on alternative days for 2 weeks, respectively.
Tumor growth was recorded twice a week, starting with the first day
of treatment. The mice were euthanized 14 days after CXB treatment,
and the subcutaneous tumors were isolated, weighed and volume
measured. Parts of each tumor tissue were wax embedded for H&E
staining to study cell apoptosis in vivo by TUNEL assay.
In situ TUNEL assay
TUNEL analysis for apoptotic cells in the tumor
tissues was performed with the Dead End™ Fluorometric TUNEL System
(Promega, Madison, WI, USA) according to the manufacturer’s
protocol.
Statistical analysis
All data are expressed as mean values ± SD.
Statistical analysis between two samples was performed using the
Student’s t-test. Statistical comparison of more than two groups
was performed using one-way ANOVA followed by a Tukey post hoc
test. The Graphpad Prism 5.0 software (GraphPad Software, San
Diego, CA, USA) was used for statistical analyses. P<0.05 was
considered to indicate a statistically significant difference.
Results
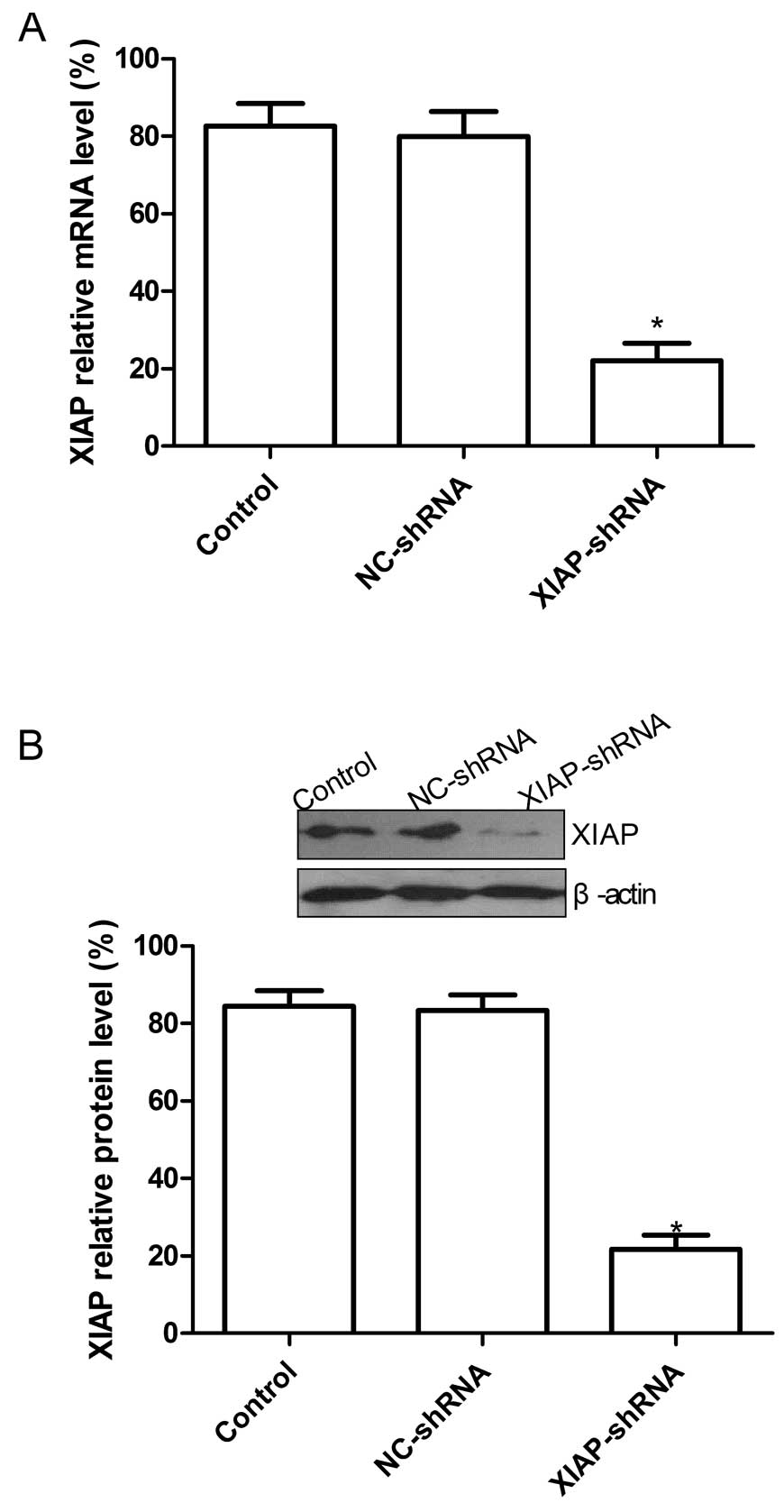
Specific downregulation of XIAP
expression by XIAP-shRNA
The mRNA and protein expression levels of the XIAP
gene, inhibited by specific shRNAs in the NSCLC cells, were
analyzed by real-time quantitive RT-PCR and western blotting. As
shown in Fig. 1A, the level of XIAP
mRNA in the XIAP-shRNA group was significantly decreased compared
with these levels in the control group and the NC-shRNA group,
respectively (P<0.05); no significant difference was noted
between the NC-shRNA group and the control group. Additionally, at
the protein expression level, no significant inhibition of XIAP
protein expression was found in the NC-shRNA group and control
group (P>0.05), while the band density decreased markedly in the
XIAP-shRNA group as compared with the NC-shRNA and control groups
(P<0.05, Fig. 1B). The above
results indicate that the expression of the XIAP gene in the A549
cells was specifically and effectively downregulated by the shRNA
targeting XIAP.
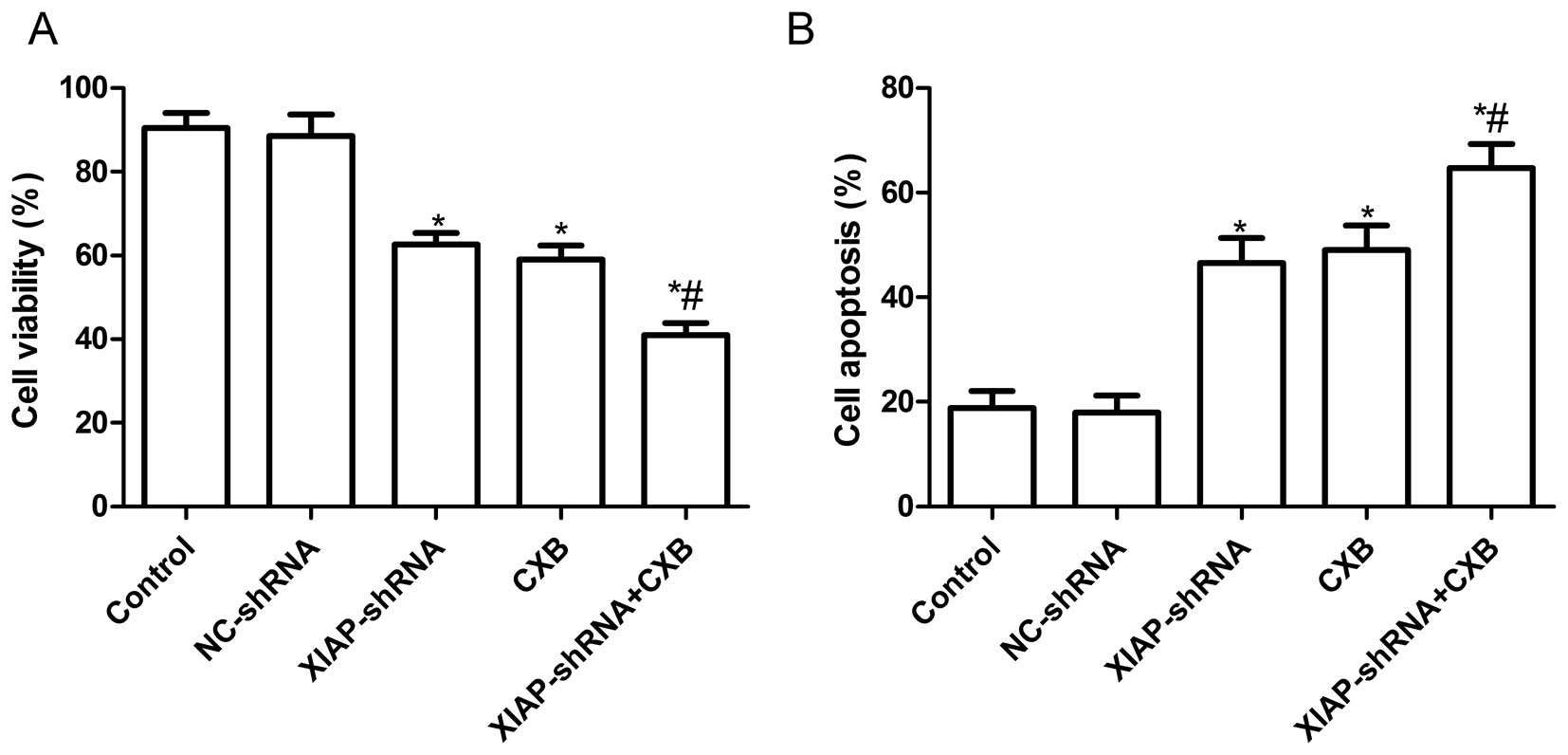
Effects of CXB and XIAP-shRNA alone or in
combination on A549 cell proliferation and apoptosis
To evaluate the effect of XIAP-shRNA and CXB alone
and in combination on the cell proliferation of NSCLC cells in
vitro, MTT assay was performed in A549 cells 48 h after
treatment with XIAP-shRNA and CXB alone or the combination. It was
found that the inhibitory rates of the cells treated with
XIAP-shRNA and CXB alone or the combination were higher than those
of the control group and NC-shRNA group (P<0.05, Fig. 2A). No significant difference was
noted between the control group and the NC-shRNA group (P>0.05,
Fig. 2A). In addition, the
inhibitory rate of the combination group was higher than the rates
in the single treatment groups (P<0.05, Fig. 2A).
The effects of XIAP-shRNA and CXB alone or the
combination on A549 cell apoptosis were then analyzed by TUNEL
assay. Compared with control group and NC-shRNA group, XIAP-shRNA
and CXB alone or the combination groups had significantly induced
cell apoptosis (P<0.05, Fig.
2B). The combination group had a greater percentage of cell
apoptosis than either drug alone treated group (P<0.05, Fig. 2B).
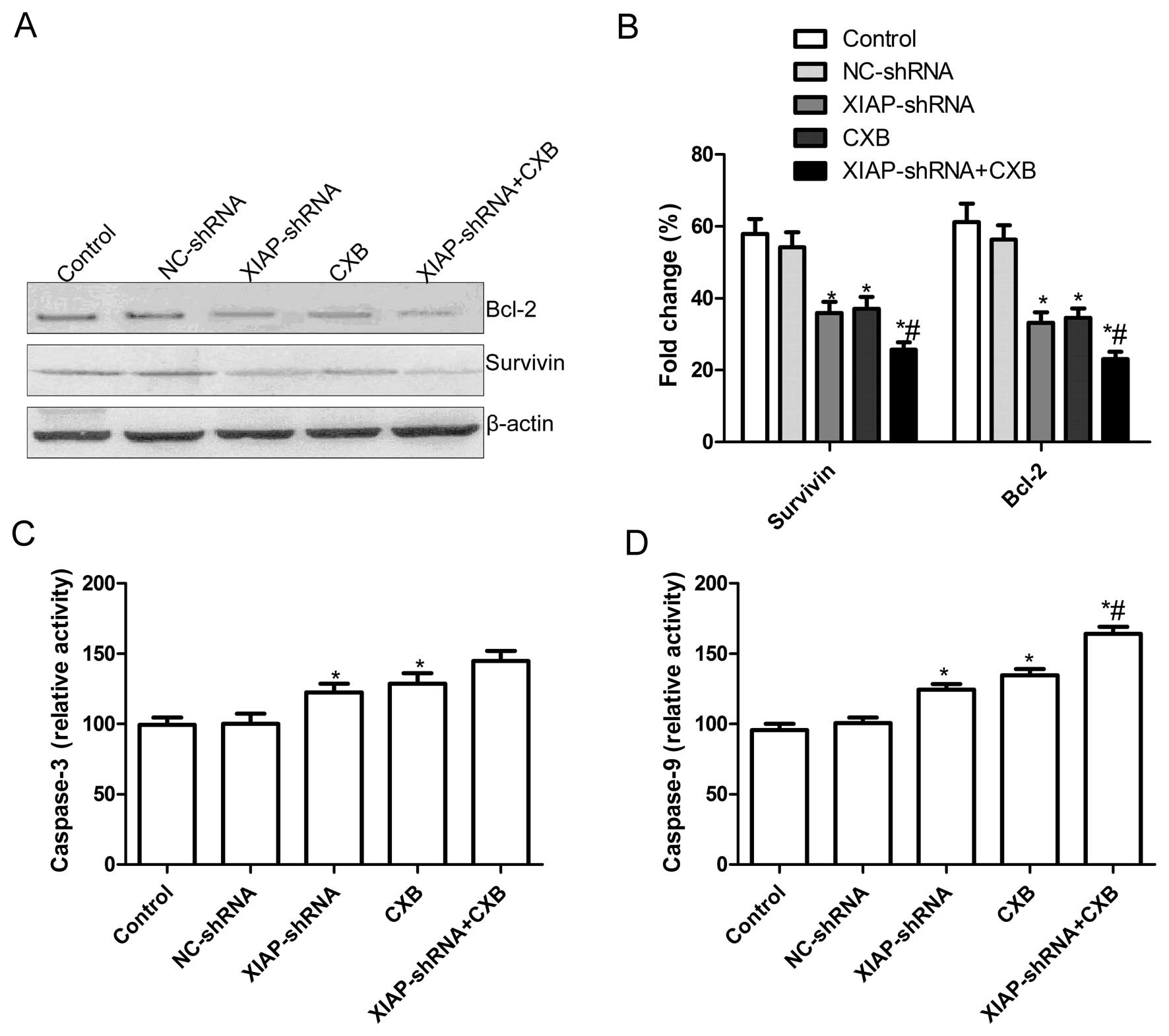
Effects of CXB and XIAP-shRNA alone or in
combination on the expression of anti-apoptotic molecules and
caspase activity in A549 cells
To explore the possible mechanism of induction of
cell apoptosis by CXB and XIAP-shRNA alone or the combination in
A549 cells, expression patterns of survivin, and Bcl-2 were
determined by western blotting. The results showed that XIAP-shRNA
and CXB alone or the combination significantly decreased the
expression of apoptosis inhibiting genes survivin and Bcl-2 in the
A549 cells compared to levels in the control group and the NC-shRNA
group (P<0.05, Fig. 3A and B).
Compared to the single treatment groups, the combination treatment
led to markedly decreased survivin and Bcl-2 protein expression
(P<0.05, Fig. 3A and B).
Additionally, caspase-3 and caspase-9 activity were then determined
by ELISA. The results showed that XIAP-shRNA and CXB alone or the
combination significantly increased caspase-3 and caspase-9
activity compared to the control group and the NC-shRNA group
(P<0.05, Fig. 3C and D), and the
combination group resulted in a greater added increase compared to
the single drug treatment groups.
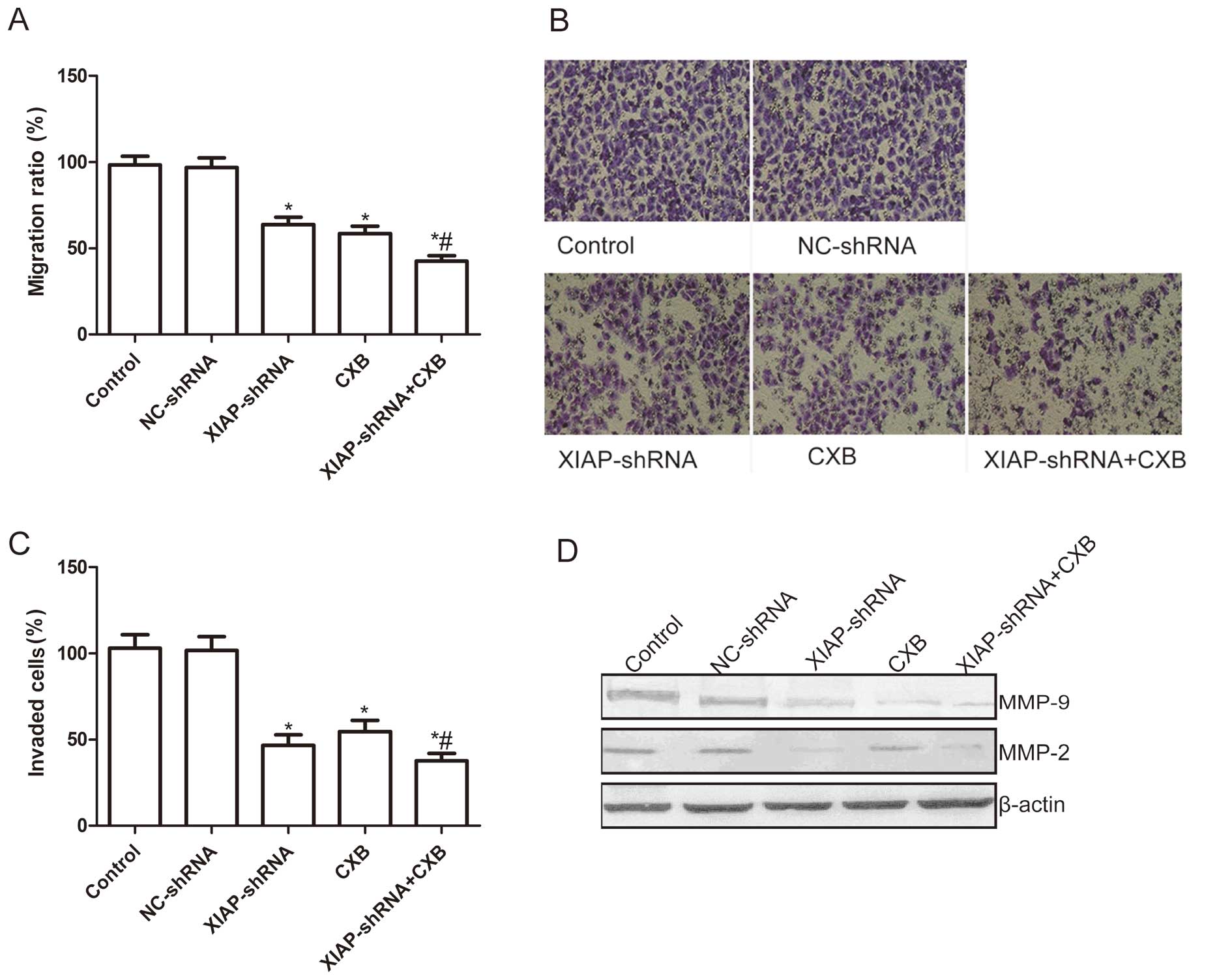
Effects of CXB and XIAP-shRNA alone or
the combination on cell migration and invasion
To ascertain the inhibitory effect of XIAP-shRNA and
CXB alone or the combination on NSCLC cell motility in
vitro, a wound-healing assay was performed. As shown in
Fig. 4A, cells in the XIAP-shRNA
and CXB alone and the combination group migrated significantly less
than those of the control group and the NC-shRNA group when A549
cells were treated with XIAP-shRNA and CXB alone or combination
treatment for 48 h (P<0.05). In addition, compared to the single
drug treatments, the migratory ability was significantly reduced in
the CXB and XIAP-shRNA combination group (P<0.05, Fig. 4A).
The invasion potential of treated cells was
evaluated by a Transwell assay. As shown in Fig. 4B, the result of the cell invasion
assay showed that there was no significant difference in the number
of cells that passed through the simulated basement membrane
between the control group and the NC-shRNA group. However, the
number of cells that passed through the simulated basement membrane
in the XIAP-shRNA and CXB alone or the combination group was
significantly reduced when compared with the control group and
NC-shRNA group. Moreover, the invasiveness of A549 cells was
further attenuated when the cells received the combination
treatment as compared with the single treatment groups.
To determine the potential mechanism of the
inhibition of cell invasion and migration by the combination
treatment of CXB and XIAP-shRNA in vitro, levels of MMP-2
and MMP-9 protein expression were determined by western blotting.
Western blot analysis displayed a significant decrease in MMP-2 and
MMP-9 proteins in the XIAP-shRNA and CXB alone and in the
combination group when compared to these levels in the control
group and the NC-shRNA group (P<0.05, Fig. 4C and D). As compared to the single
treatment groups, the combination group exhibited obviously
decreased MMP-2 and MMP-9 protein expression (P<0.05, Fig. 4C and D).
Effects of CXB and XIAP-shRNA alone or
the combination on PGE2 production and the VEGF level
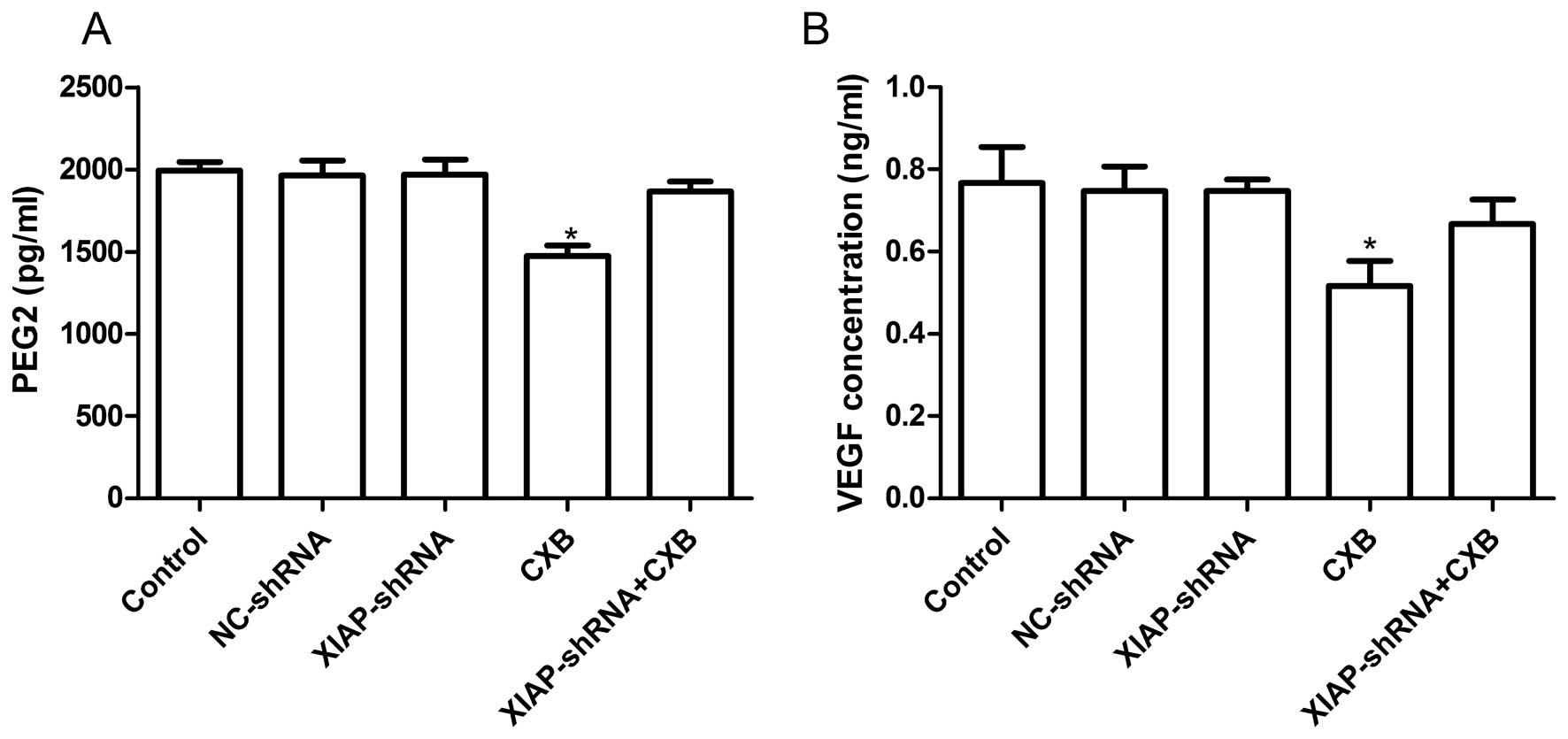
To examine the effects of XIAP-shRNA and CXB alone
and in combination on PGE2 production in A549 cells, ELISA was
performed. As shown in Fig. 5A, CXB
inhibited significant PGE2 production (P<0.05); however, CXB in
combination with XIAP-shRNA or XIAP-shRNA alone did not inhibit
PGE2 production. We also determined the VEGF level by ELISA. As
shown in Fig. 5B, ELISA analysis
revealed that VEGF excretion in the supernatant from the CXB group
was significantly decreased compared to the other groups
(P<0.05). CXB in combination with XIAP-shRNA or XIAP-shRNA alone
did not inhibit VEGF protein expression.
Effects of CXB and XIAP-shRNA alone or
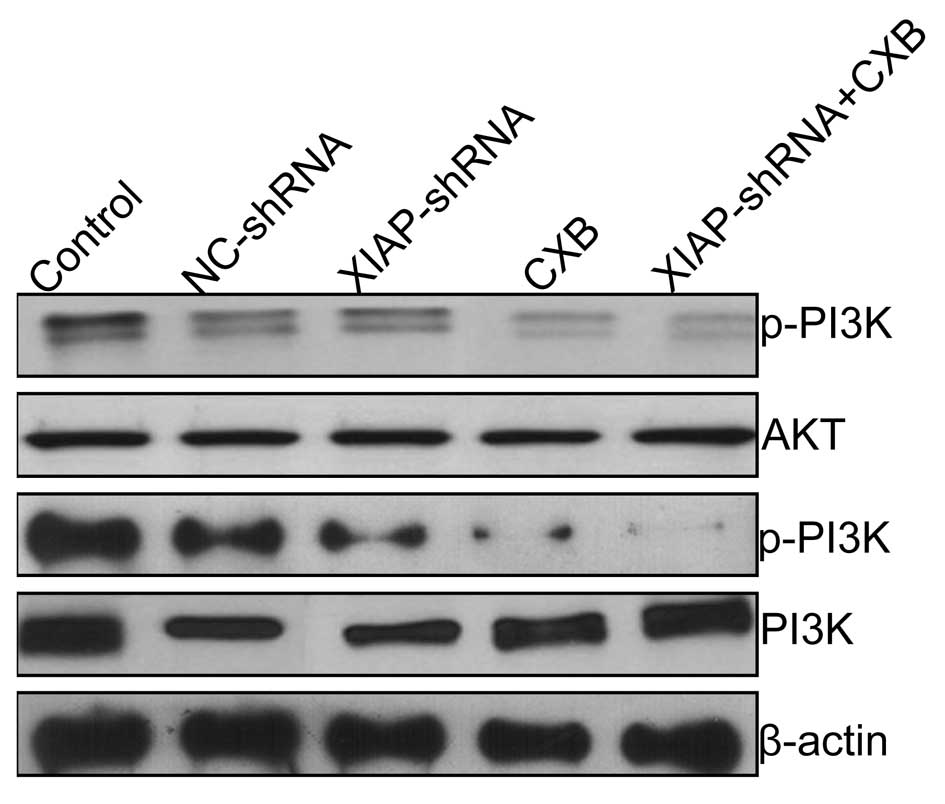
the combination on the PI3K/Akt pathway
It has been demonstrated that celecoxib (CXB) is
involved in Akt phosphorylation inhibition and regulates cell
apoptosis (18). We aimed to
ascertain whether the mediation of this signaling pathway could be
enhanced by CXB in combination with XIAP-shRNA. Measurement of the
phosphorylation/activation pattern of PI3K and Akt was performed by
western blotting. Our results showed that CXB and XIAP-shRNA alone
or the combination resulted in a marked reduction in phosphorylated
PI3K and Akt relative to the control group and NC-shRNA group,
without altering the total protein levels of PI3K or Akt in each
group (Fig. 6). Compared to the CXB
and XIAP-shRNA alone treatment groups, the combination group
exhibited obviously decreased phosphorylation of PI3K and Akt
protein expression, demonstrating a synergistic effect.
Effects of CXB and XIAP-shRNA alone or
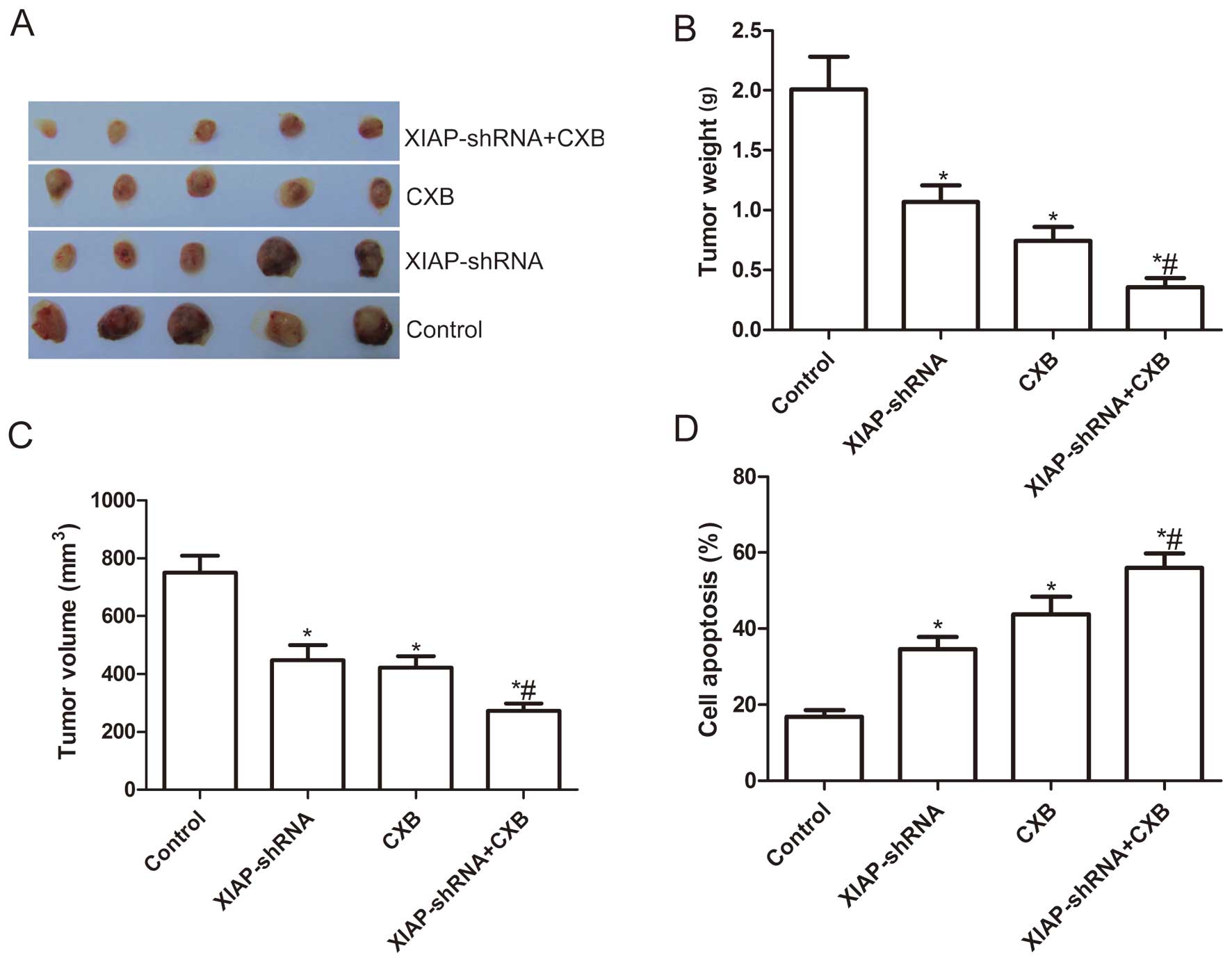
the combination on tumor growth in vivo
Finally, we assessed the in vivo therapeutic
efficacy of XIAP-shRNA and CXB alone or the combination on female
BALB mice bearing A549 cell tumors. In the XIAP-shRNA treatment
group, the tumor size and weight were reduced to 63.61 and 53.23%;
in the CXB treated group, the tumor size and weight were reduced to
56.13 and 37.08%; and in the combination group, tumor size and
weight were reduced to 36.12 and 17.79%. These results suggest that
the most prominent tumor decrease occurred in the combination
group, demonstrating the synergistic effect in vivo
(P<0.05, Fig. 7A–C).
In addition, we also determined the synergistic
effects on tumor tissue cell apoptosis in vivo by TUNEL. The
results showed that XIAP-shRNA and CXB alone or the combination
significantly induced cell apoptosis compared to the control group
and the NC-shRNA group (P<0.05, Fig.
7D). The combination of XIAP-shRNA and CXB greatly induced
tumor cell apoptosis in vivo compared to the single drug
treatment groups (P<0.05, Fig.
7D).
Discussion
Non-small cell lung cancer is one of the leading
causes of cancer-related death in many countries (27). Despite the fact that advances in
diagnostic and therapeutic methodologies have been exploited to
improve the survival of patients with NSCLC, a satisfactory level
has not been achieved as the majority of patients present at an
advanced unresectable stage and their tumors exhibit an inherent
resistance to chemotherapy and radiotherapy. Therefore, the
development of new treatment strategies with high efficacy and low
toxicity is required for NSCLC (28).
It is well known that combination treatments achieve
a better curative effect when compared to single drug treatments.
Combination treatment mainly depends on the synergism between
different therapeutic agents with different mechanisms of action to
increase efficacy, while maintaining a favorable side effect and
toxicity profile. One example of a drug that is commonly used as a
component of combination therapies is CXB. It is one of the most
widely used chemotherapeutic agents for the treatment of lung
cancer (21). Although CXB induces
substantial antitumor activity and cytotoxicity in various cancer
types (29–31), it also results in the development of
resistance through altered bioavailability or inactivation and
nonspecific cytotoxicity (32).
Several studies have demonstrated that the response rate with the
combination of CXB and gefitinib for advanced NSCLC patients was
similar to that with gefitinib alone (33,34).
Recently, Koch et al found that patients with advanced NSCLC
at stage IIIB–IV who received CXB plus chemotherapy also failed to
demonstrate a survival benefit for CXB (35). In addition, CXB causes various side
effects at the therapeutic dosage including cardiovascular
thrombosis, congestive heart failure, gastrointestinal ulceration,
renal or hepatic injury, and platelet aggregation (22). Therefore, by reducing the dosage of
CXB the cytotoxic effects may be achieved by using combination
therapy.
One of the major factors responsible for
chemoresistance is the evolution of tumor cells towards a phenotype
that is resistant to apoptotic cell killing. Cells resist apoptosis
by a key molecular mechanism, the overexpression of inhibitor of
apoptosis protein (IAP) family of proteins. A key member of this
family is XIAP. It has been showed that XIAP over-expression is
correlated with resistance to apoptosis through stimulation of both
the intrinsic (mitochondrial-directed) and extrinsic (death
receptor-directed) pathways (36,37).
Several studies have shown that downregulation of XIAP with siRNA
restores chemosensitivity in various tumor cell lines (9–14). Pan
et al showed that the combination of XIAP-shRNA and TRAIL
resulted in significant reduction in XIAP expression and potent
antitumor activity both in HCC cells and in an animal tumor model
(38). Therefore, in this study, we
selected XIAP-shRNA in combination with a low dose of CXB for
NSCLC. Our results showed that XIAP-shRNA combined with a low dose
of CXB inhibited NSCLC tumor growth in vitro and in an
animal model.
Although the combination therapy improves the
response rate and time to progression compared with sequential
monotherapy, it causes increased toxicity. Consequently, in cases
of advanced cancer, emerging guidelines recommend sequential
monotherapy to improve quality of life. An alternative approach
that could overcome nonspecific toxicity while retaining
therapeutic efficacy, involves the combination of chemotherapy with
targeted therapy by RNAi technology. Recent research shows that
combined therapy with an shRNA targeting gene in combination with
an anticancer drug could decrease toxicity and chemoresistance, and
achieve better antitumor activity (39). For example, Ghosh et al
showed that sequence-dependent combination treatment using low-dose
doxorubicin and an anti-survivin agent was efficacious against both
breast and pancreatic cancer (39).
Du et al showed that treatment with a lentivirus carrying
the COX-2 gene in combination with tamoxifen in breast cancer cells
significantly suppressed tumor growth in vitro and in
vivo (40). Xue et al
found that the combination of sur-shRNA and emodin was effective
for the treatment of ovarian cancer compared to sur-shRNA and
emodin alone treatment (41).
Consistent with these results, our results demonstrated that
XIAP-shRNA in combination with CXB treatment in NSCLC significantly
suppressed the proliferation, migration and invasion, and induced
cell apoptosis in vitro, and inhibited tumor growth in a
mouse model.
The PI3K/Akt pathway plays an important role in lung
cancer pathogenesis, the upregulation of which is associated with a
more aggressive clinical phenotype and worse clinical outcome for
endocrine-treated patients (42–44).
It has been showed that CXB inhibits the expression of
phosphorylated-AKT and regulates cell apoptosis (18). In the present study, our results
showed that CXB and XIAP-shRNA alone or the combination resulted in
a marked reduction in phosphorylated PI3K and Akt relative to the
control group and NC-shRNA group, without altering the total
protein levels of PI3K or Akt in each group. XIAP-shRNA in
combination with CXB obviously decreased phosphorylated PI3K and
Akt protein expression compared to the CXB and XIAP-shRNA alone
treatment groups, demonstrating a synergistic effect. In addition,
our study showed that CXB inhibited PGE2 production and VEGF
expression, which was consistent with a previous study (42). However, CXB in combination with
XIAP-shRNA or XIAP-shRNA alone did not inhibit PGE2 production and
VEGF expression, which suggests that CXB in combination with
XIAP-shRNA did not have a synergistic effect on PGE2 production and
VEGF expression in A549 cells. These findings imply that XIAP-shRNA
in combination with CXB has a synergistic effect on the inhibition
of tumor growth of NSCLC in vitro and in vivo, at
least in part, through affecting the PI3K/AKT signaling
pathway.
In conclusion, our in vitro studies
demonstrated that when plasmid XIAP-shRNA and CXB are used in
combination, they provide an effective therapeutic approach for
NSCLC. This combination significantly suppressed the proliferation,
migration and invasion, and induced cell apoptosis when compared to
treatment with XIAP-shRNA and CXB alone. Additionally, further
studies with in vivo mouse models also confirmed that this
combination suppressed the tumor growth of NSCLC. These findings
suggest that the combination of XIAP-shRNA and CXB is a promising
drug candidate for the treatment of NSCLC.
Acknowledgements
This resea rch was suppor ted by the Science and
Technology Research and Innovation Team Fund of Jilin Province
(JL2013038).
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Reungwetwattana T, Weroha SJ and Molina
JR: Oncogenic pathways, molecularly targeted therapies, and
highlighted clinical trials in non-small-cell lung cancer (NSCLC).
Clin Lung Cancer. 13:252–266. 2012. View Article : Google Scholar
|
|
3
|
Zhang H, Li Z and Wang K: Combining
sorafenib with celecoxib synergistically inhibits tumor growth of
non-small cell lung cancer cells in vitro and in vivo. Oncol Rep.
31:1954–1960. 2014.PubMed/NCBI
|
|
4
|
Hunter AM, LaCasse EC and Korneluk RG: The
inhibitors of apoptosis (IAPs) as cancer targets. Apoptosis.
12:1543–1568. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Danson S, Dean E, Dive C and Ranson M:
IAPs as a target for anticancer therapy. Curr Cancer Drug Targets.
7:785–794. 2007. View Article : Google Scholar
|
|
6
|
Devi GR: XIAP as target for therapeutic
apoptosis in prostate cancer. Drug News Perspect. 17:127–134. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Stennicke HR, Ryan CA and Salvesen GS:
Reprieval from execution: the molecular basis of caspase
inhibition. Trends Biochem Sci. 27:94–101. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Holcik M, Gibson H and Korneluk RG: XIAP:
apoptotic brake and promising therapeutic target. Apoptosis.
6:253–261. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ma JJ, Chen BL and Xin XY: XIAP gene
downregulation by small interfering RNA inhibits proliferation,
induces apoptosis, and reverses the cisplatin resistance of ovarian
carcinoma. Eur J Obstet Gynecol Reprod Biol. 146:222–226. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yamaguchi Y, Shiraki K, Fuke H, et al:
Targeting of X-linked inhibitor of apoptosis protein or survivin by
short interfering RNAs sensitize hepatoma cells to TNF-related
apoptosis-inducing ligand- and chemotherapeutic agent-induced cell
death. Oncol Rep. 14:1311–1316. 2005.PubMed/NCBI
|
|
11
|
Jiang C, Yi XP, Shen H and Li YX:
Targeting X-linked inhibitor of apoptosis protein inhibits
pancreatic cancer cell growth through p-Akt depletion. World J
Gastroenterol. 18:2956–2965. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kwatra SG: Targeting X-linked inhibitor of
apoptosis protein for melanoma therapy: the need for more
homogeneous samples and the importance of cell lines. J Invest
Dermatol. 131:7972011. View Article : Google Scholar
|
|
13
|
Hiscutt EL, Hill DS, Martin S, et al:
Targeting X-linked inhibitor of apoptosis protein to increase the
efficacy of endoplasmic reticulum stress-induced apoptosis for
melanoma therapy. J Invest Dermatol. 130:2250–2258. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mastrangelo E, Cossu F, Milani M, et al:
Targeting the X-linked inhibitor of apoptosis protein through
4-substituted azabicyclo[5.3.0]alkane smac mimetics. Structure,
activity, and recognition principles. J Mol Biol. 384:673–689.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Harlin H, Reffey SB, Duckett CS, Lindsten
T and Thompson CB: Characterization of XIAP-deficient mice. Mol
Cell Biol. 21:3604–3608. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang ZL, Fan ZQ, Jiang HD and Qu JM:
Selective Cox-2 inhibitor celecoxib induces epithelial-mesenchymal
transition in human lung cancer cells via activating MEK-ERK
signaling. Carcinogenesis. 34:638–646. 2013. View Article : Google Scholar
|
|
17
|
Dhawan D, Jeffreys AB, Zheng R, Stewart JC
and Knapp DW: Cyclooxygenase-2 dependent and independent antitumor
effects induced by celecoxib in urinary bladder cancer cells. Mol
Cancer Ther. 7:897–904. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kulp SK, Yang YT, Hung CC, et al:
3-phosphoinositide-dependent-protein kinase-1/Akt signaling
represents a major cyclooxygenase-2-independent target for
celecoxib in prostate cancer cells. Cancer Res. 64:1444–1451. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen ST, Thomas S, Gaffney KJ, Louie SG,
Petasis NA and Schönthal AH: Cytotoxic effects of celecoxib on Raji
lymphoma cells correlate with aggravated endoplasmic reticulum
stress but not with inhibition of cyclooxygenase-2. Leuk Res.
34:250–253. 2010. View Article : Google Scholar
|
|
20
|
Jendrossek V, Handrick R and Belka C:
Celecoxib activates a novel mitochondrial apoptosis signaling
pathway. FASEB J. 17:1547–1549. 2003.PubMed/NCBI
|
|
21
|
Liu X, Yue P, Zhou Z, Khuri FR and Sun SY:
Death receptor regulation and celecoxib-induced apoptosis in human
lung cancer cells. J Natl Cancer Inst. 96:1769–1780. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Menter DG, Schilsky RL and DuBois RN:
Cyclooxygenase-2 and cancer treatment: understanding the risk
should be worth the reward. Clin Cancer Res. 16:1384–1390. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jeon YW and Suh YJ: Synergistic apoptotic
effect of celecoxib and luteolin on breast cancer cells. Oncol Rep.
29:819–825. 2013.
|
|
24
|
Cervello M, Bachvarov D, Lampiasi N, et
al: Novel combination of sorafenib and celecoxib provides
synergistic anti-proliferative and proapoptotic effects in human
liver cancer cells. PLoS One. 8:e655692013. View Article : Google Scholar
|
|
25
|
Morisaki T, Umebayashi M, Kiyota A, et al:
Combining celecoxib with sorafenib synergistically inhibits
hepatocellular carcinoma cells in vitro. Anticancer Res.
33:1387–1395. 2013.PubMed/NCBI
|
|
26
|
Huang KH, Kuo KL, Chen SC, et al:
Down-regulation of glucose regulated protein (GRP) 78 potentiates
cytotoxic effect of celecoxib in human urothelial carcinoma cells.
PLoS One. 7:e336152012. View Article : Google Scholar
|
|
27
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J and
Thun MJ: Cancer statistics, 2009. CA Cancer J Clin. 59:225–249.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mendez M, Custodio A and Provencio M: New
molecular targeted therapies for advanced non-small-cell lung
cancer. J Thorac Dis. 3:30–56. 2011.
|
|
29
|
Dhawan D, Craig BA, Cheng L, et al:
Effects of short-term celecoxib treatment in patients with invasive
transitional cell carcinoma of the urinary bladder. Mol Cancer
Ther. 9:1371–1377. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu X, Yue P, Schönthal AH, Khuri FR and
Sun SY: Cellular FLICE-inhibitory protein down-regulation
contributes to celecoxib-induced apoptosis in human lung cancer
cells. Cancer Res. 66:11115–11119. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Huang S and Sinicrope FA:
Celecoxib-induced apoptosis is enhanced by ABT-737 and by
inhibition of autophagy in human colorectal cancer cells.
Autophagy. 6:256–269. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Davies NM, McLachlan AJ, Day RO and
Williams KM: Clinical pharmacokinetics and pharmacodynamics of
celecoxib: a selective cyclooxygenase-2 inhibitor. Clin
Pharmacokinet. 38:225–242. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gadgeel SM, Ruckdeschel JC, Heath EI,
Heilbrun LK, Venkatramanamoorthy R and Wozniak A: Phase II study of
gefitinib, an epidermal growth factor receptor tyrosine kinase
inhibitor (EGFR-TKI), and celecoxib, a cyclooxygenase-2 (COX-2)
inhibitor, in patients with platinum refractory non-small cell lung
cancer (NSCLC). J Thorac Oncol. 2:299–305. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Agarwala A, Fisher W, Bruetman D, et al:
Gefitinib plus celecoxib in chemotherapy-naive patients with stage
IIIB/IV non-small cell lung cancer: a phase II study from the
Hoosier Oncology Group. J Thorac Oncol. 3:374–379. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Koch A, Bergman B, Holmberg E, et al:
Effect of celecoxib on survival in patients with advanced non-small
cell lung cancer: a double blind randomised clinical phase III
trial (CYCLUS study) by the Swedish Lung Cancer Study Group. Eur J
Cancer. 47:1546–1555. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Holcik M, Yeh C, Korneluk RG and Chow T:
Translational upregulation of X-linked inhibitor of apoptosis
(XIAP) increases resistance to radiation induced cell death.
Oncogene. 19:4174–4177. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wilkinson JC, Cepero E, Boise LH and
Duckett CS: Upstream regulatory role for XIAP in receptor-mediated
apoptosis. Mol Cell Biol. 24:7003–7014. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pan Q, Liu B, Liu J, Cai R, Liu X and Qian
C: Synergistic antitumor activity of XIAP-shRNA and TRAIL expressed
by oncolytic adenoviruses in experimental HCC. Acta Oncol.
47:135–144. 2008. View Article : Google Scholar
|
|
39
|
Ghosh SK, Yigit MV, Uchida M, et al:
Sequence-dependent combination therapy with doxorubicin and a
survivin-specific small interfering RNA nanodrug demonstrates
efficacy in models of adenocarcinoma. Int J Cancer. 134:1758–1766.
2014. View Article : Google Scholar
|
|
40
|
Du Y, Shi A, Han B, et al: COX-2 silencing
enhances tamoxifen antitumor activity in breast cancer in vivo and
in vitro. Int J Oncol. 44:1385–1393. 2014.PubMed/NCBI
|
|
41
|
Xue H, Chen Y, Cai X, et al: The combined
effect of survivin-targeted shRNA and emodin on the proliferation
and invasion of ovarian cancer cells. Anticancer Drugs. 24:937–944.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang Q, Meng X, Zheng G, et al: Antitumor
activity of celecoxib, a selective cyclooxygenase-2 inhibitor, in
medullary thyroid carcinoma. Mol Med Rep. 9:768–772. 2014.
|
|
43
|
Vivanco I and Sawyers CL: The
phosphatidylinositol 3-kinase AKT pathway in human cancer. Nat Rev
Cancer. 2:489–501. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
44
|
Perez-Tenorio G and Stal O; Southeast
Sweden Breast Cancer Group. Activation of AKT/PKB in breast cancer
predicts a worse outcome among endocrine treated patients. Br J
Cancer. 86:540–545. 2002. View Article : Google Scholar : PubMed/NCBI
|