Introduction
Hepatocellular carcinoma (HCC) is a major health
issue and has one of the highest mortality rates in the world
(1). Unfortunately, most cases of
HCC are often diagnosed at an advanced stage and are not suitable
for curative treatments such as resection, transplantation
(2,3), radiofrequency ablation, transarterial
chemoembolization (TACE), or targeting drugs such as sorafenib
(4,5). It is known that tumorigenesis is a
multi-stage complex process involving multiple genes. Activation of
resistant genes or the mutation of sensitive genes in the
development of cancer may lead to the failure of chemotherapeutic
agents. Therefore, the study of anticancer drugs is currently a hot
research topic.
Salinomycin (Salin) has been widely used in animal
husbandry for many years worldwide. Salin is a polyether antibiotic
used to kill gram-positive bacteria including mycobacteria and
parasites such as Plasmodium falciparum. In addition, Salin,
as an ionophore with strict selectivity for alkali ions, exhibits a
wide range of biological activities, including inhibition of
adipogenesis and anti-allergic activity (6,7).
Recent studies indicate that Salin has antitumor effects, with
attenuation of proliferation, autophagy and cell death/apoptosis in
human cancer cells or cancer stem cells (8–11).
However, the effects of Salin on the migratory and invasive
properties of HCC cells, and the underlying molecular mechanisms
remain obscure.
In the present study, we demonstrated that the
anti-invasive and anti-migratory effects of Salin are mediated by
downregulation of MMP9 through the JNK/JunD pathway leading to
inhibition of HCC cell invasion and metastasis.
Materials and methods
Cell lines and culture conditions
The human HCC cell lines with highly invasive
capacities (HCCLM3 and MHCC-97H) were obtained from the Chinese
Academy of Sciences Committee Type Culture Collection cell bank.
The cells were grown in Dulbecco’s modified Eagle’s medium (DMEM;
Life Technologies, Ann Arbor, MI, USA) supplemented with 10% fetal
bovine serum (FBS; Life Technologies) and both penicillin and
streptomycin (100 mg/ml each) at 37°C in a humidified atmosphere of
5% CO2.
Cell migration and invasion assays
Cells (5×104) were seeded in the upper
chamber of Transwell plates with 8-μm pores (Costar, Cambridge, MA,
USA). The lower chambers of the Transwell plates were filled with
500 μl medium containing 10% FBS as a chemoattractant. The plates
were incubated at 37°C for 12 h. Cell invasion assays were
performed using the same method. The Transwell chambers were
covered with 50 μl 1:2 Matrigel and phosphate-buffered Saline
mixture, and the cells were cultivated for 24 h. Cells that
migrated or invaded to the lower surface were stained with Giemsa
solution and quantified by counting five randomly selected
microscopic fields at ×200 magnification.
Angiogenesis assay
The supernatant was collected from the HCCLM3 cells
cultured in a serum-free medium with or without Salin treatment.
The Matrigel angiogenesis assay was performed as previously
described (12,13). In brief, BD Matrigel (BD Biosciences
San Jose, CA, USA) matrix was plated in 96-well flat-bottom cell
culture cluster plates. After incubation for 30 min, 10,000 HUVEC
cells/well and 50 μl of the HCCLM3 supernatant with or without
Salin treatment were placed on the Matrigel. The plate was
incubated at 37°C for 16–18 h. Following incubation, the wells were
photographed, and the results were quantified by measuring the
length of the tube-like structures using Nikon NIS-Elements
computer software.
Enzyme-linked immunosorbent assays
(ELISA)
To measure the concentrations of MMP2 and MMP9
secreted from the cultured tumor cell lines, the supernatants were
assessed using ELISA. Cells (5×104/well) in 24-well
plates were incubated at 37°C in a 5% CO2 atmosphere in
DMEM containing 10% FBS. After 24 h, the cells were washed and
incubated for 24 h in serum-free medium without/with Salin (5–20
μmol/l). The culture-conditioned medium was collected, centrifuged,
and the concentrations of MMP2 and MMP9 were determined by
quantitative ELISA (R&D Systems, Minneapolis, MN, USA),
according to the manufacturer’s instructions.
MMP9 promoter and enhancer and luciferase
reporter gene constructs
To clone the putative promoter and/or enhancer
region of the MMP9 gene, a PCR-based method was used, and specific
primers were designed from the 5′-end of the known MMP9 promoter
sequence from a previous study (14). The amplified DNA fragment of 2302 bp
was cloned into the pGL3-Basic vector (Promega, Madison, WI, USA)
to construct pGL3-MMP9-WT containing the potential enhancer
element. AP-1 site-mutated MMP9 (pGL3-MMP9-mAP1-1, pGL3-MMP9-mAP1-2
and pGL3-MMP9-mAP1-1+2), NF-κB site-mutated MMP9 and Sp-1
site-mutated MMP9 (pGL3-MMP9-mSp1) luciferase promoters were used
in the transient transfection assays as previously described
(15,16). These vectors contain the luciferase
gene driven by the SV40 promoter. The composition of the constructs
was confirmed by restriction endonuclease digestion and DNA
sequencing.
Transfection and luciferase reporter
assays
Cells (5×104/well) in 24-well plates were
incubated at 37°C in a 5% CO2 atmosphere in serum-free
DMEM. After 24 h, the plasmids were transfected into cells using
Lipofectamine-2000 reagent according to the manufacturer’s
instructions (Invitrogen, Carlsbad, CA, USA). After transfection,
the cells were cultured in FBS without or with Salin (5–20 μmol/l)
for 24 h. The cell lysate was used to detect luciferase activity
according to the manufacturer’s protocol (Promega). Luciferase
activity was normalized to the β-galactosidase activity in the cell
lysates and expressed as the average of three independent
experiments.
Western blotting
Total cellular proteins were extracted using RIPA
buffer and quantified by the BCA method. The sample proteins were
separated by electrophoresis on SDS-PAGE and transferred onto a
PVDF membrane. After blocking, the membranes were incubated
overnight at 4°C with various primary antibodies and α-tubulin at
1:500–1:1000 dilution (Santa Cruz Biotechnology, Santa Cruz, CA,
USA). The secondary HRP-conjugated antibodies were diluted 1:2000.
The immunocomplexes were detected using the ECL system (Beyotime
Biotechnology, China). A Li-COR Odyssey scanner (LICOR) was used to
detect the antigens on the blots.
Statistical analysis
All statistical analyses were performed using SPSS
17.0 software. Data are reported as mean ± SD, and mean values were
compared using the Student’s t-test and ANOVA. Values of p<0.05
and p<0.01 were considered statistically significant.
Results
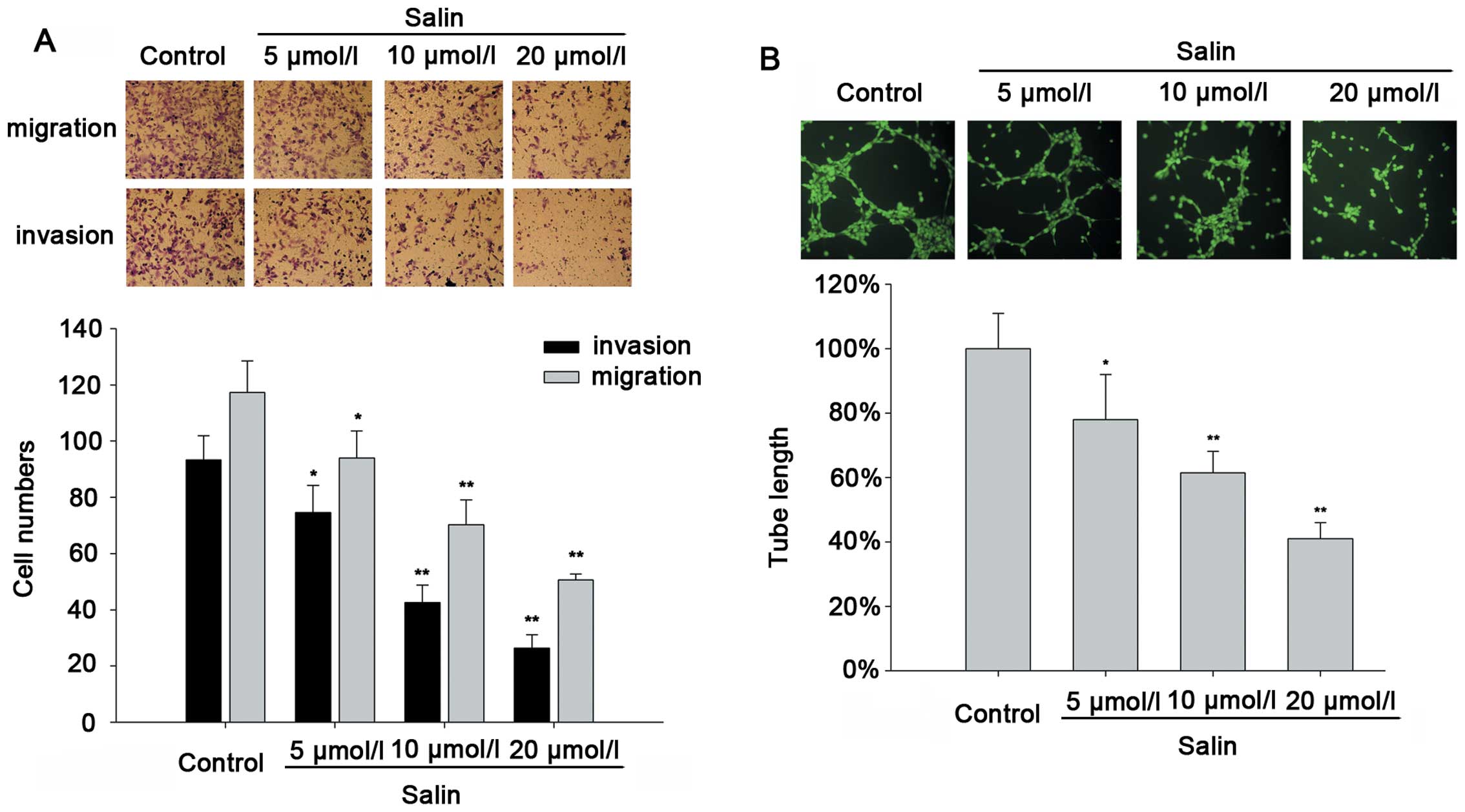
Salinomycin inhibits HCC cell migration
and invasion
It is known that Salin has anticancer activity, and
previous studies have shown that it can induce cell apoptosis and
suppress cancer cell proliferation. We determined the effects of
Salin on the migration and invasion of HCC cells using the
Transwell assay and tube formation assay. Salin (5–20 μmol/l)
significantly inhibited HCCLM3 cell migration and invasion in a
concentration-dependent manner (Fig.
1A). In addition, significantly fewer integrated capillary-like
structures were found in the HCCLM3 cells treated with Salin (5–20
μmol/l), indicating that Salin affected the release of
pro-angiogenic proteins in the HCCLM3 cells (Fig. 1B). Similar results were confirmed in
the other invasive HCC cell line, MHCC-97H (data not shown),
suggesting that the anti-invasive or anti-migratory effect of Salin
in the HCC cell lines is universal.
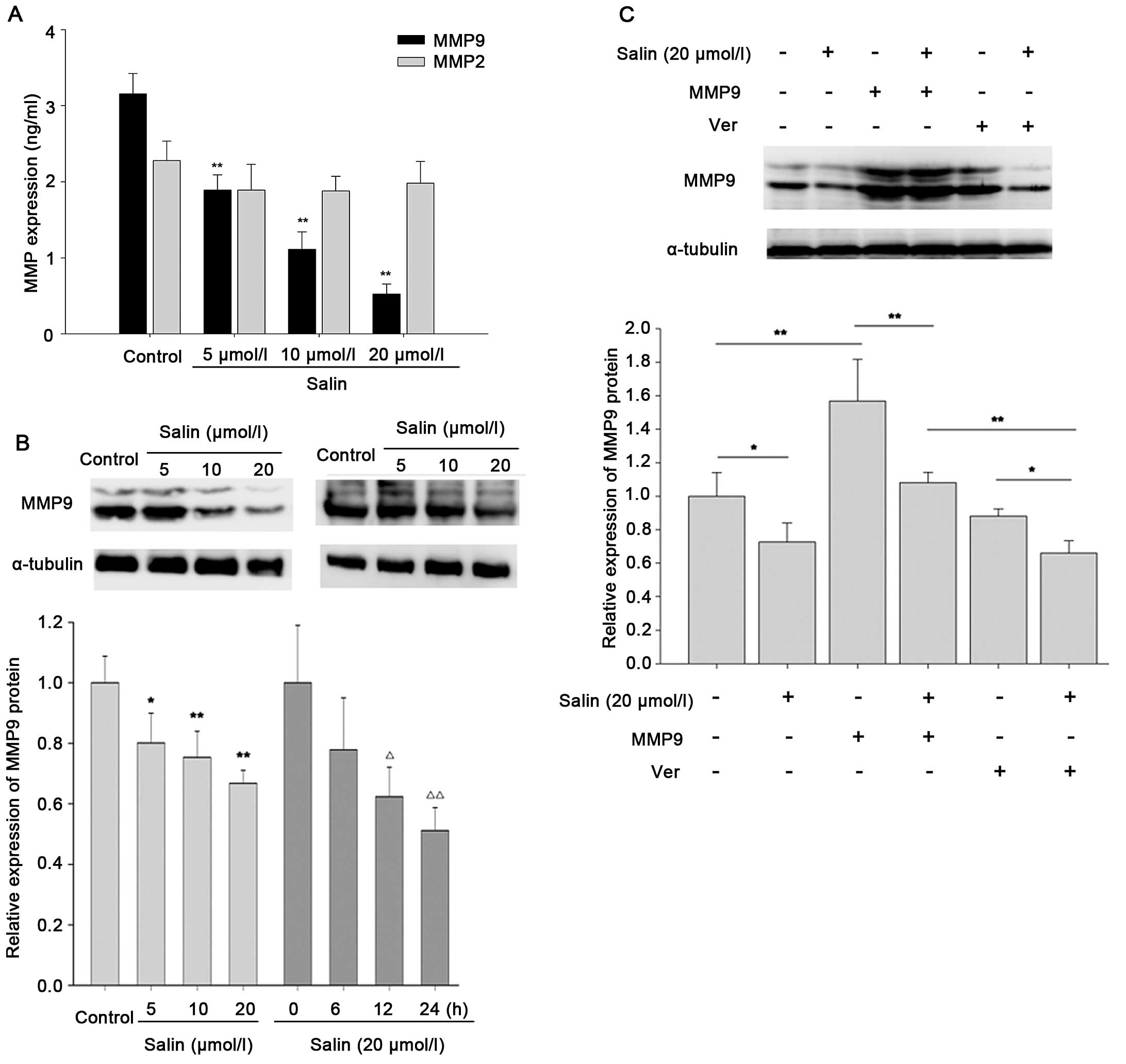
Suppression of cancer cell invasion and
metastasis by Salin is associated with the downregulation of
MMP9
To elucidate the anti-invasive and anti-migratory
mechanisms of Salin, we examined MMP9 and MMP2 expression. HCCLM3
cells were treated with Salin at concentrations of 5–20 μmol/l. The
results showed that Salin at 5 μmol/l or higher concentrations
attenuated MMP9 expression in the HCCLM3 cells, but did not
significantly affect the expression of MMP2 (Fig. 2A). In addition, Salin inhibited MMP9
expression at the protein level in a concentration-dependent and
time-dependent manner (Fig. 2B). To
determine the importance of MMP9 in the anti-invasive effect of
Salin in HCCLM3 cells, we established stable overexpression of MMP9
in the HCCLM3 transfectants by infecting the cells with a
lentivirus encoding MMP9. The results showed that overexpression of
MMP9 was significantly blocked by 10–39% in the Salin-treated group
compared with the group without Salin treatment (p<0.01;
Fig. 2C). The results also showed
that Salin decreased MMP9 expression by 14–36% compared with the
random group (Ver transfectants) (p<0.05; Fig. 2C). These results suggest that MMP9
may play a role in the anti-invasive and anti-migratory effects of
Salin.
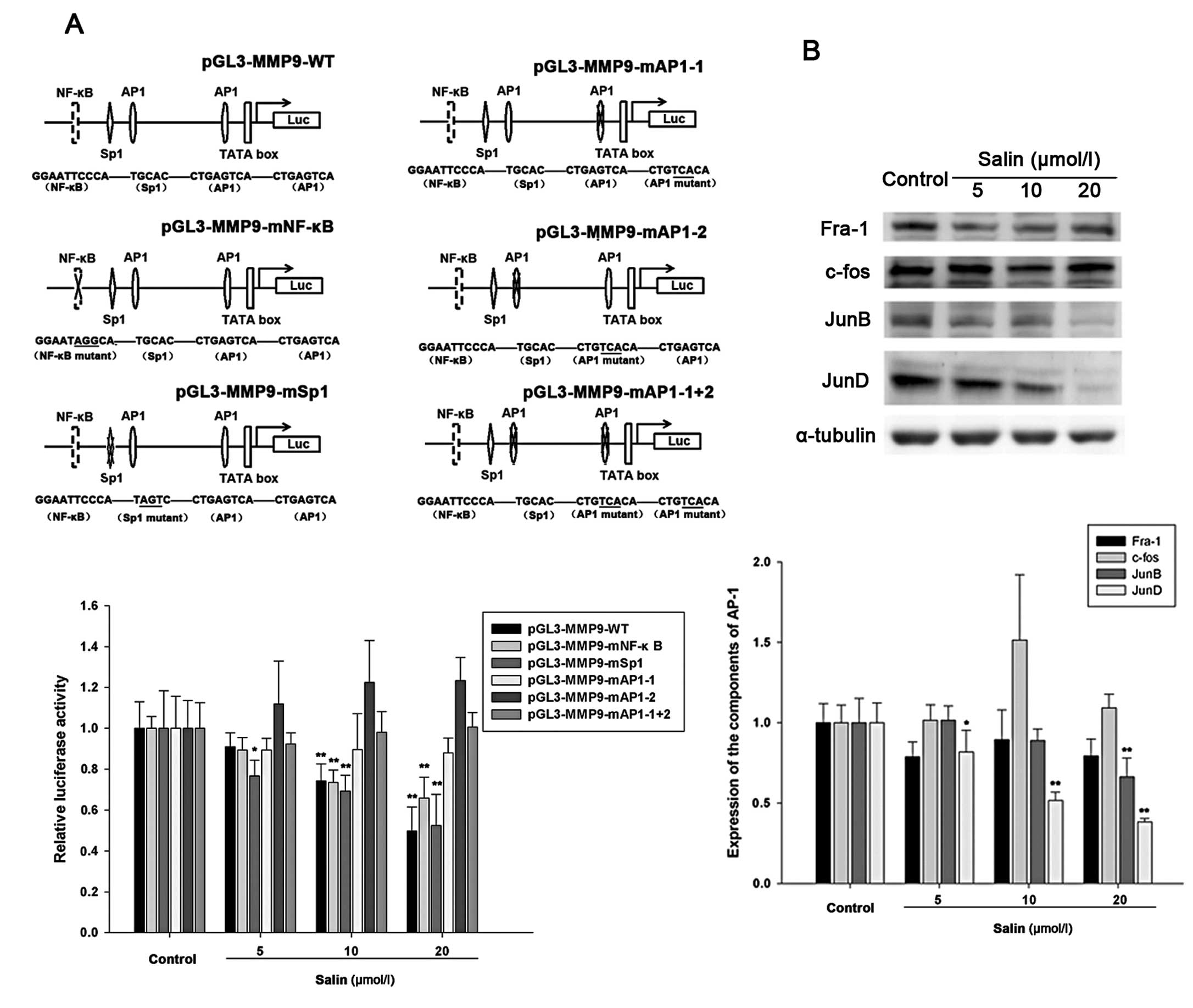
The AP-1 site within the MMP9 enhancer is
essential for Salin-regulated MMP9 activity
The MMP9 promoter contains two AP-1 sites (located
at −79 bp and −533 bp), a Sp1 site (located at −560 bp) and an
NF-κB site (located at −600 bp). To determine whether regulation of
MMP9 is related to these cis-acting regulatory elements for
transcription factors, several constructs with deletions or
mutations were used and have been described in Materials and
methods. Experimental cells were transfected with reporter vectors
that included the tandem repeat of AP-1, NF-κB or Sp-1 binding
sites. Noteworthy, luciferase activity in the cells with the AP-1
construct was significantly reduced by treatment with Salin at
10–20 μmol/l, whereas luciferase activity containing the NF-κB or
Sp-1 binding site construct showed no statistically significant
changes in the cells treated with Salin (Fig. 3A). These results showed that both
AP-1 sites in the MMP9 promoter were essential for Salin-regulated
MMP9 enhancer region activity.
JNK/JunD signaling pathway is responsible
for MMP9 downregulation by Salin
The AP-1 complex is composed of several protein
components, including c-Fos, JunD, JunB and Fra-1. We aimed to
determine which members play a major role in the downregulation of
MMP9 by Salin. The results demonstrated that only JunD was
significantly downregulated by Salin in the HCCLM3 cells (Fig. 3B). JunB showed a slightly increasing
trend, particularly following treatment with 20 μmol/l Salin,
whereas Fra-1 and c-Fos were not significantly affected by Salin at
concentrations ranging from 5 to 20 μmol/l (Fig. 3B).
To evaluate the role of JunD in Salin-regulated MMP9
activity, we investigated JunD and MMP9 expression at different
times and at different Salin concentrations. The data showed that
Salin simultaneously reduced JunD and MMP9 protein expression
(Fig. 4A). It was also shown that
Salin affected protein expression at 10 μmol/l or higher
concentrations, and this was concentration-dependent (Fig. 4A). In addition, as shown in Fig. 4B, the target proteins were inhibited
by Salin in a time-dependent manner. The dynamic effects of JunD
and MMP9 expression occurred after 12 h.
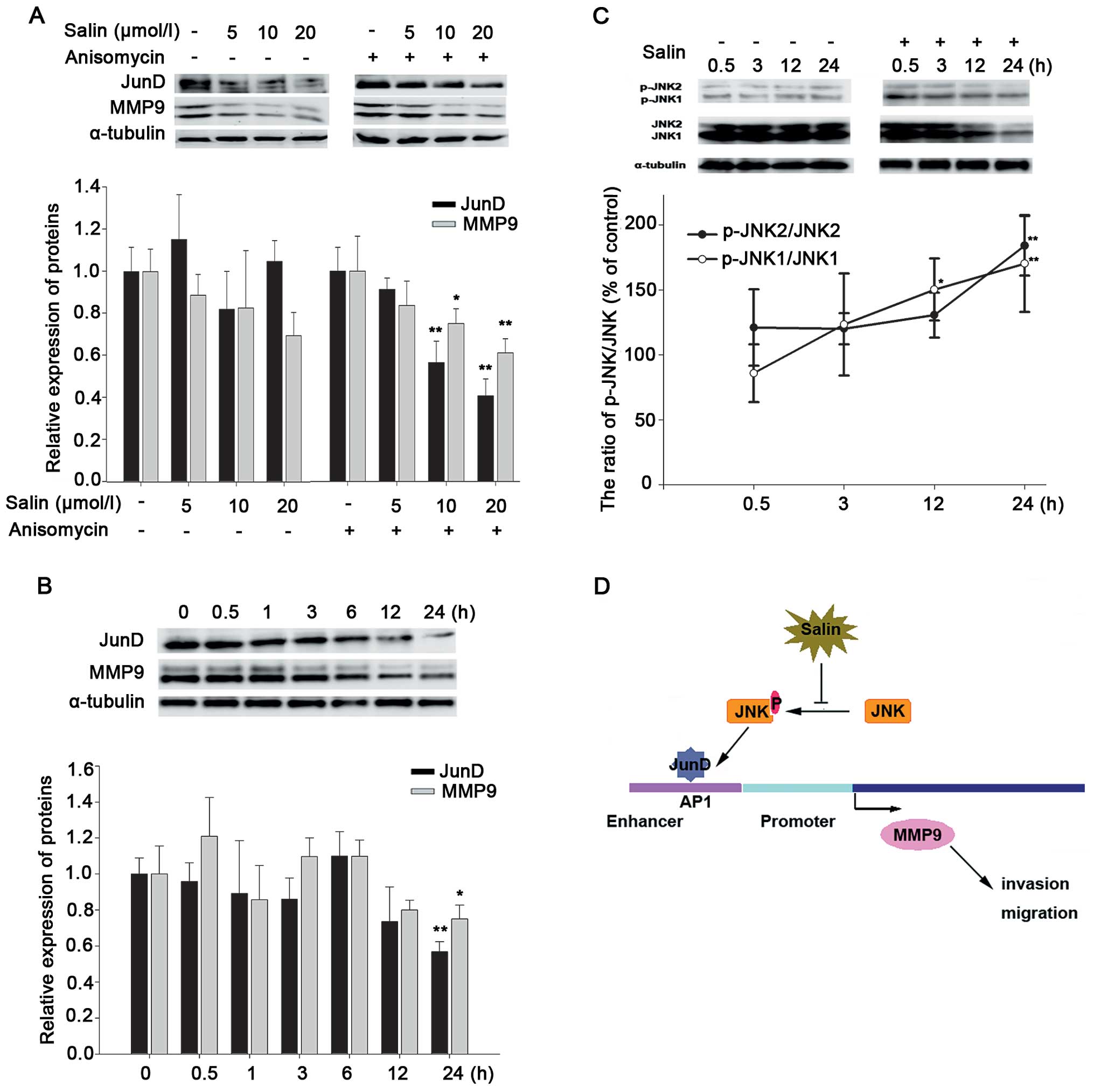 | Figure 4JNK/JunD signaling pathway is involved
in MMP9 downregulation by Salin. (A) Salin-regulated JunD and MMP9
expression. Western blotting showed that Salin inhibited JunD and
MMP9 expression induced by anisomycin, a JNK activator. (B) Western
blotting revealed that JunD and MMP9 expression was inhibited by
Salin at the indicated times (0, 0.5, 1, 3, 6, 12 and 24 h) during
incubation. (C) The effects of Salin on JNK activation at the
indicated time periods (0.5, 3, 12 and 24 h). After treatment
without or with 20 μmol/l Salin, JNK1, JNK2, p-JNK1 and p-JNK2 were
examined by western blotting with JNK and phosphor-JNK specific
antibodies. The ratios of p-JNK1/JNK1 and p-JNK2/JNK2 showed that
JNK phosphorylation was regulated by Salin. (D) The hypothetical
mechanism by which Salin causes MMP9 inhibition and related
signaling events in the presence of its anti-invasive and
anti-migratory effects. *p<0.05 vs. the control,
**p<0.01 vs. the control. Data are representative of
at least three independent experiments. |
To further confirm the effects of Salin on these
signaling cascades, anisomycin, an activator of c-Jun N-terminal
kinase (JNK), was added to the HCCLM3 cells with or without Salin.
The results showed that Salin blocked anisomycin-induced JunD
expression (Fig. 4A). Antibodies
against the phosphorylated forms of JNK were then used to determine
the changes in the phosphorylation level following Salin
intervention. The data showed that the phosphorylated forms of JNK1
and JNK2, p-JNK1 and p-JNK2, were significantly impeded by Salin in
a time-dependent manner (Fig. 4C).
These results showed that JNK phosphorylation and JunD may be
involved in the Salin-regulated MMP9 signaling pathway in HCCLM3
cells and may mediate HCC cell biological characteristics.
Discussion
Salinomycin (Salin) is crucially involved in the
regulation of cell proliferation, growth and survival, in addition
to anti-tumorigenesis. However, the role of Salin in cancer
invasion and metastasis is not well understood. In the prsent
study, we confirmed that Salin inhibited HCC cell migration and
invasion in vitro through the downregulation of MMP9 rather
than MMP2. In addition, our results indicated that Salin regulated
MMP9 at the protein level in a concentration-dependent and
time-dependent manner.
It is well known that degradation of the
extracellular matrix is the angiogenic switch in tumorigenesis. In
particular, MMP9, its enhancer and promoter can bind to the
transcription factors AP-1, Sp1 and NF-κB, and can be affected by a
variety of signals (16). Several
natural products or chemicals play a role in antitumor activity by
interfering with MMP9 gene expression. For example, hesperidin
obstructs the activity of MMP9 by inhibiting NF-κB (17), and genistein suppresses MMP9
transcription by inhibiting AP-1 and NF-κB activity (18). In the present study, we showed, for
the first time, that Salin acts through AP-1 to inhibit MMP9
expression. Previous evidence indicated that the regulation of JunD
expression diverges from the well-characterized growth
factor-inducible pattern of the c-Jun early response genes and AP-1
autoregulation (19,20). Furthermore, we found that the
onco-suppressive effects of Salin may partially mediate MMP9
expression via the JNK/JunD pathway, causing the suppression of
cancer cell invasion and migration (Fig. 4D). These results suggest that Salin
inhibits cancer-cell invasion by decreasing JNK/JunD signaling and
AP-1 activation to prevent MMP9 expression.
In conclusion, we discovered novel pathways
associated with the ability of Salin to suppress tumorigenesis. The
onco-suppressive effects of Salin may be partially mediated via
JNK/JunD-regulated MMP9 expression. These findings confirm the
possibility that Salin may be a potential anticancer drug targeting
MMP9.
Acknowledgements
This study was funded by the Shanghai Municipal
Health Bureau Key Disciplines Grant (no. ZK2012A05), the Shanghai
Municipal Health Bureau (no. 20134100), the Natural Science
Foundation of the Science and Technology Commission of Shanghai
Municipality (no. 14ZR1431600).
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Belghiti J and Fuks D: Liver resection and
transplantation in hepatocellular carcinoma. Liver Cancer. 1:71–82.
2012. View Article : Google Scholar
|
|
3
|
Fan ST: Hepatocellular carcinoma -
resection or transplant? Nat Rev Gastroenterol Hepatol. 9:732–737.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Raza A and Sood GK: Hepatocellular
carcinoma review: current treatment, and evidence-based medicine.
World J Gastroenterol. 20:4115–4127. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Feng K and Ma KS: Value of radiofrequency
ablation in the treatment of hepatocellular carcinoma. World J
Gastroenterol. 20:5987–5998. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Szkudlarek-Mikho M, Saunders RA, Yap SF,
Ngeow YF and Chin KV: Salinomycin, a polyether ionophoric
antibiotic, inhibits adipogenesis. Biochem Biophys Res Commun.
428:487–493. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
García-Domenech R, Zanni R,
Galvez-Llompart M and de Julián-Ortiz JV: Modeling anti-allergic
natural compounds by molecular topology. Comb Chem High Throughput
Screen. 16:628–635. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang F, He L, Dai WQ, Xu YP, Wu D, Lin CL,
Wu SM, Cheng P, Zhang Y, Shen M, Wang CF, Lu J, Zhou YQ, Xu XF, Xu
L and Guo CY: Salinomycin inhibits proliferation and induces
apoptosis of human hepatocellular carcinoma cells in vitro and in
vivo. PLoS One. 7:e506382012. View Article : Google Scholar
|
|
9
|
Wang F, Dai W, Wang Y, Shen M, Chen K,
Cheng P, Zhang Y, Wang C, Li J, Zheng Y, Lu J, Yang J, Zhu R, Zhang
H, Zhou Y, Xu L and Guo C: The synergistic in vitro and in vivo
antitumor effect of combination therapy with salinomycin and
5-fluorouracil against hepatocellular carcinoma. PLoS One.
9:e974142014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Al Dhaheri Y, Attoub S, Arafat K, Abuqamar
S, Eid A, Al Faresi N and Iratni R: Salinomycin induces apoptosis
and senescence in breast cancer: upregulation of p21,
downregulation of survivin and histone H3 and H4 hyperacetylation.
Biochim Biophys Acta. 1830.3121–3135. 2013.
|
|
11
|
Yue W, Hamaï A, Tonelli G, Bauvy C,
Nicolas V, Tharinger H, Codogno P and Mehrpour M: Inhibition of the
autophagic flux by salinomycin in breast cancer
stem-like/progenitor cells interferes with their maintenance.
Autophagy. 9:714–729. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yesildal F, Aydin F, Deveci S, Tekin S,
Aydin I, Mammadov R, Fermanli O, Avcu F, Acikel C and Ozgurtas T:
Aspartame induces angiogenesis in vitro and in vivo models. Hum Exp
Toxicol. pii: 0960327114537535. Jun 12–2014.(Epub ahead of print).
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shi GM, Ke AW, Zhou J, Wang XY, Xu Y, Ding
ZB, Devbhandari RP, Huang XY, Qiu SJ, Shi YH, Dai Z, Yang XR, Yang
GH and Fan J: CD151 modulates expression of matrix
metalloproteinase 9 and promotes neoangiogenesis and progression of
hepatocellular carcinoma. Hepatology. 52:183–196. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Raney BJ, Cline MS, Rosenbloom KR, Dreszer
TR, Learned K, Barber GP, Meyer LR, Sloan CA, Malladi VS, Roskin
KM, Suh BB, Hinrichs AS, Clawson H, Zweig AS, Kirkup V, Fujita PA,
Rhead B, Smith KE, Pohl A, Kuhn RM, Karolchik D, Haussler D and
Kent WJ: ENCODE whole-genome data in the UCSC genome browser.
Nucleic Acids Res. 39(Database issue): D871–D875. 2011. View Article : Google Scholar :
|
|
15
|
Chuang TW, Lee YC and Kim CH: Hepatitis B
viral HBx induces matrix metalloproteinase-9 gene expression
through activation of ERK and PI-3K/AKT pathways: involvement of
invasive potential. FASEB J. 18:1123–1125. 2004.
|
|
16
|
Hong S, Park KK, Magae J, Ando K, Lee TS,
Kwon TK, Kwak JY, Kim CH and Chang YC: Ascochlorin inhibits matrix
metalloproteinase-9 expression by suppressing activator
protein-1-mediated gene expression through the ERK1/2 signaling
pathway: inhibitory effects of ascochlorin on the invasion of renal
carcinoma cells. J Biol Chem. 280:25202–25209. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yeh MH, Kao ST, Hung CM, Liu CJ, Lee KH
and Yeh CC: Hesperidin inhibited acetaldehyde-induced matrix
metalloproteinase-9 gene expression in human hepatocellular
carcinoma cells. Toxicol Lett. 184:204–210. 2009. View Article : Google Scholar
|
|
18
|
Wang SD, Chen BC, Kao ST, Liu CJ and Yeh
CC: Genistein inhibits tumor invasion by suppressing multiple
signal transduction pathways in human hepatocellular carcinoma
cells. BMC Complement Altern Med. 14:262014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hernandez JM, Floyd DH, Weilbaecher KN,
Green PL and Boris-Lawrie K: Multiple facets of junD gene
expression are atypical among AP-1 family members. Oncogene.
27:4757–4767. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen HW, Lee JY, Huang JY, Wang CC, Chen
WJ, Su SF, Huang CW, Ho CC, Chen JJ, Tsai MF, Yu SL and Yang PC:
Curcumin inhibits lung cancer cell invasion and metastasis through
the tumor suppressor HLJ1. Cancer Res. 68:7428–7438. 2008.
View Article : Google Scholar : PubMed/NCBI
|