Introduction
Pancreatic cancer, one of the most lethal human
gastrointestinal malignancies, is the fourth leading cause of
cancer mortality and almost 1/10,000 individuals are diagnosed
annually in the United States (1).
Pancreatic cancer is characterized by late diagnosis, strong local
invasion, early metastasis, high lethal rate, poor prognosis, low
long-term survival and <5% of five-year survival rate (2), and is consequently known as ‘the king
of cancer’.
In China, the incidence of pancreatic cancer ranks
7th among all tumors while it is ranked 6th with regard to
mortality rate in all cancer types (3). Improvement of living standards and
changes of the diet structure, have led to an upward tendency of
the incidence of pancreatic cancer in recent years, particularly in
young men (4).
In clinic, 90–95% of the exocrine pancreatic
carcinoma is pancreatic ductal adenocarcinoma cancer (5). Studies have shown that the median
survival time of pancreatic cancer without surgical resection
following diagnosis is 3–6 months, which may be extended to 23
months following surgical resection. However, only 10–15% of
diagnosed patients can be completely resected, even if the 5-year
survival rate of these patients is only 10% (5,6). In
addition, almost all of the patients with pancreatic cancer succumb
to the disease within seven years after surgery.
Current methods of treatment for pancreatic cancer
include surgery, chemotherapy, radiotherapy and biological therapy,
with surgical excision remaining as the only effective and
treatable means (7–9). Moreover, low pancreatic resection,
high postoperative recurrence and metastasis, and common resistance
to chemotherapeutic drugs are the common courses of death in
pancreatic cancer patients. Therefore, exploiting novel chemicals
is the key to improving the prognosis of pancreatic cancer.
At present, temozolomide, one of the imidazole
tetrazines, has already been applied in the clinic and exhibits
improved curative effect on leukemia, lymphoma and solid tumors.
ZGDHu-1
(N,N′-di-(m-methylphenyi)-3,6-dimethyl-1,4-dihydro-1,2,4,5-tetrazine-1,4-dicarboamide),
which was used in this study is a new tetrazine derivative and was
initially designed and produced by W.-X. Hu (Pharmaceutical College
of Zhejiang University of Technology, China) (10,11).
Moreover, it was previously found that ZGDHu-1 has antitumor
activities in vitro and in vivo and inhibits the
proliferation of myeloid leukemia (12) and lung cancer (13), inducing cell apoptosis and
differentiation. However, little is known concerning the underlying
effect and mechanism of ZGDHu-1 on human pancreatic cancer
cells.
In this study, following treatment with this
tetrazine derivative, we found that ZGDHu-1 inhibited the
proliferative effect and enhanced apoptosis in PANC-1 cells, and
blocked its cell cycle at G2/M phase. Furthermore, the
mitochondrial pathway and activation of caspase-3 are important in
ZGDHu-1-induced apoptosis, and the nuclear factor inhibitor IκB was
also involved in this process. CyclinB1, cdc2 and Chk1 are G2/M
regulatory molecules, whose expression levels were altered
following the treatment of ZGDHu-1.
Materials and methods
Cell culture and reagents
The human PANC-1 pancreatic cancer cell line was
obtained from the Cell Bank of the Chinese Academy of Sciences
(Shanghai, China). The cells were cultured in DMEM medium
containing 10% FBS, 100 U/ml penicillin and 100 μg/ml streptomycin
in a humidified atmosphere of 5% CO2 at 37°C. Dulbecco’s
modified Eagle’s medium (DMEM), fetal bovine serum (FBS) and 0.25%
Trypsin-EDTA were purchased from Dingguo Biotechnology Co., Ltd.
(Gibco, Grand Island, NY, USA). Dimethylsulfoxide (DMSO),
3-(4,5-dimethylthiazol-2–yl)-2,5-diphenyltetrazolium bromide (MTT)
and Hoechst-33258 apoptosis kit (no. B1155) were purchased from
Sigma Chemical Co. (St. Louis, MO, USA). The DNA prep kit (no.
6607055) was purchased from Beckman Coulter, Inc. (Brea, CA, USA).
The Annexin V/Dead Cell Apoptosis kit (no. V13241) was purchased
from MultiSciences Biotech Co., Ltd. (Invitrogen, Carlsbad, CA,
USA), and the ECL Western Blotting Substrate kit (no. P1425) was
purchased from MultiSciences Biotech Co., Ltd. (Hangzhou, China).
The RIPA lysis buffer (no. WB-0071) and BCA Protein Quantitation
kit (no. BCA02) were purchased from DingGuo Biotechnology Co., Ltd.
(Beijing, China), and PageRuler™ Prestained Protein Marker (no.
26616) was purchased from Thermo Scientific (Rockford, IL, USA).
Primary antibodies against Caspase-3 (no. 9665), cleaved-Caspase-3
(no. 9661), PARP (no. 9532), Bax (no. 2774), NF-κB (no. 3034), IκB
(no. 9242), Chk1 (no. 2345), cyclinB1 (no. 4135), cdc2 (no. 9116)
and α-tubulin (no. 3873) were purchased from Cell Signaling
Technology (Danvers, MA, USA), and Bcl-2(sc-7382) was purchased
from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). Primary
antibodies against β-actin and secondary antibodies were purchased
from MultiSciences Biotech Co., Ltd. Drug solutions were freshly
prepared on the day of testing.
Preparation of ZGDHu-1
ZGDHu-1 was kindly provided by the Pharmaceutical
Engineering Research Institute, College of Pharmaceutical Science,
Zhejiang University of Technology. ZGDHu-1 was dissolved in DMSO
(maximum concentration, 10 mg/ml) and stored at −20°C for use.
Prior to the in vitro experiment, DMEM was used to dilute
the stock solution (10 mg/ml) to the final working
concentration.
Cell viability assay
The MTT assay was used to measure the cell viability
of PANC-1 cells. Briefly, PANC-1 cells were seeded in 96-well
plates at a density of 8×103 cells per well and
incubated overnight at 37°C in 10% FBS medium. The cells were then
treated with increasing concentrations (0, 0.01, 0.05, 0.1, 0.2,
0.5 and 1 μg/ml of ZGDHu-1 or DMEM medium (control). After
incubation for 24, 48 and 72 h at 37°C, 20 μl of MTT solution (5
mg/ml) was added to each well, and the cells were incubated for 4 h
at 37°C, and 150 μl of DMSO was added to dissolve the crystal. The
optical density (OD) value at 490 nm was determined using a
microplate reader (Rayto, USA). Cell viability was calculated as:
Cell viability (%) =
[(ODsample−ODblank)/(ODcontrol−OD
blank)] ×100.
Morphological analysis
To evaluate the apoptotic activity of PANC-1 cells,
we used Hoechst-33258 and Wright’s staining to observe the changes
of the cell nucleus. Cover glasses were initially placed in 6-well
plates. The PANC-1 cells (5×105 cells in 2 ml) were
subsequently seeded in 6-well plates overnight and exposed to
increasing concentrations of ZGDHu-1 or DMEM medium (control) for
24 and 48 h. The cells for Hoechst-33258 were fixed with fixative
(methanol:glacial acetic acid 3:1) for 5 min at 4°C and incubated
with Hoechst-33258 (5 μg/ml) for 15 min after washing with PBS. The
cells were then observed under a fluorescence microscope (Nikon
Y-THS, Japan). However, the cells for Wright’s staining were
stained with Wright A and B for 10 min and observed under a light
microscope (Olympus CHA, Japan).
Detection of cell apoptosis
The Annexin V/Dead Cell Apoptosis kit was used to
detect PANC-1 cell apoptosis by flow cytometry. Firstly, PANC-1
cells (5×105 cells in 2 ml) were seeded in 6-well plates
overnight and exposed to increasing concentrations of ZGDHu-1 or
DMEM medium (control) for 24 h. The cells were harvested with
trypsin, collected by centrifugation (192 × g for 5 min), and
washed twice with cold PBS. Cells at 5×105 cells/ml were
resuspended in 1X Annexin V binding buffer at a final concentration
of 1×106 cells/ml. The cells were stained with 5 μl of
Annexin V-FITC and 1 μl of PI (100 μg/ml) for 15 min at room
temperature in the dark. After incubation, 400 μl of 1X Annexin V
binding buffer were added to an adequate cell dilution and detected
by flow cytometry (FC500, Beckman Coulter, Inc.) using 488 nm
excitation.
Flow cytometry cell cycle analysis
PANC-1 cells were treated as described above.
Briefly, after harvesting, centrifugation (192 × g for 5 min) and
washing, the pellet was incubated with 50 μl of DNA PREP LPR
(containing RNase) for 1 min and then treated with DNA PREP™ Stain
(containing PI) in the dark for at least 5 min. The samples were
analyzed by flow cytometry (FC500, Beckman Coulter, Inc.) and at
least 10,000 cells were counted.
Western blot analysis
To identify the potential molecular mechanism of
ZGDHu-1 on PANC-1 cells, expression levels of relative proteins
were measured by western blotting. PANC-1 cells were seeded in the
75 cm2 dishes at a density of 2.5×106
cells/dish. After being cultured overnight, PANC-1 cells were
treated with different concentrations of ZGDHu-1 for 24 and 48 h,
respectively. PANC-1 cells were then collected and lysed using RIPA
lysis buffer and the protein concentration was determined using a
BCA protein quantification kit following the manufacturer’s
instructions. For each sample, an equal amount of protein (50 μg)
was separated by SDS-PAGE (10–12%) and transferred onto a PVDF
membrane at the constant current of 200 mA for 60 min. The membrane
was blocked with 10% non-fat dried milk in Tris-buffered saline
with Tween-20 (TBST) for 2 h and then incubated with primary
antibodies (1:1,000) overnight at 4°C individually. After washing
with TBST three times, the membranes were hybridized with secondary
antibody-conjugated horseradish peroxidase (goat anti-mouse
1:10,000, goat anti-rabbit 1:10,000) at room temperature for 2 h.
The ECL Western blotting substrate kit was added on the membranes
for reaction and the fluorescence of the protein bands was exposed
to X-ray film. All the protein levels were normalized by β-actin or
α-tubulin.
Statistical analysis
Results are presented as the means ± standard
deviation. The significance of differences between two groups was
determined using the t-test. All the results were obtained from
three separate experiments. Differences were considered to be
statistically significant at values of p<0.05.
Results
ZGDHu-1 suppresses the proliferation of
PANC-1 cells
The structure of ZGDHu-1, a new tetrazine
derivative, is shown in Fig. 1A. We
firstly evaluated the cytotoxic efficacy of ZGDHu-1 on the PANC-1
pancreatic cancer cell line by using MTT assay. Following treatment
with various concentrations (0.01, 0.05, 0.1, 0.2, 0.5, 1 or 5
μg/ml) of ZGDHu-1 for 24, 48 and 72 h, the results demonstrated
that ZGDHu-1 inhibited the proliferation of PANC-1 cells in a time-
and dose-dependent manner (Fig.
1B–E). Moreover, the half maximal inhibitory concentration
(IC50) of 24, 48 and 72 h for PANC-1 cells was 2.95,
0.15 and 0.12 μg/ml of ZGDHu-1 (Fig.
1F). Therefore, concentrations of 0.1, 0.2 and 0.5 μg/ml of
ZGDHu-1 on the cells were used for the subsequent experiments.
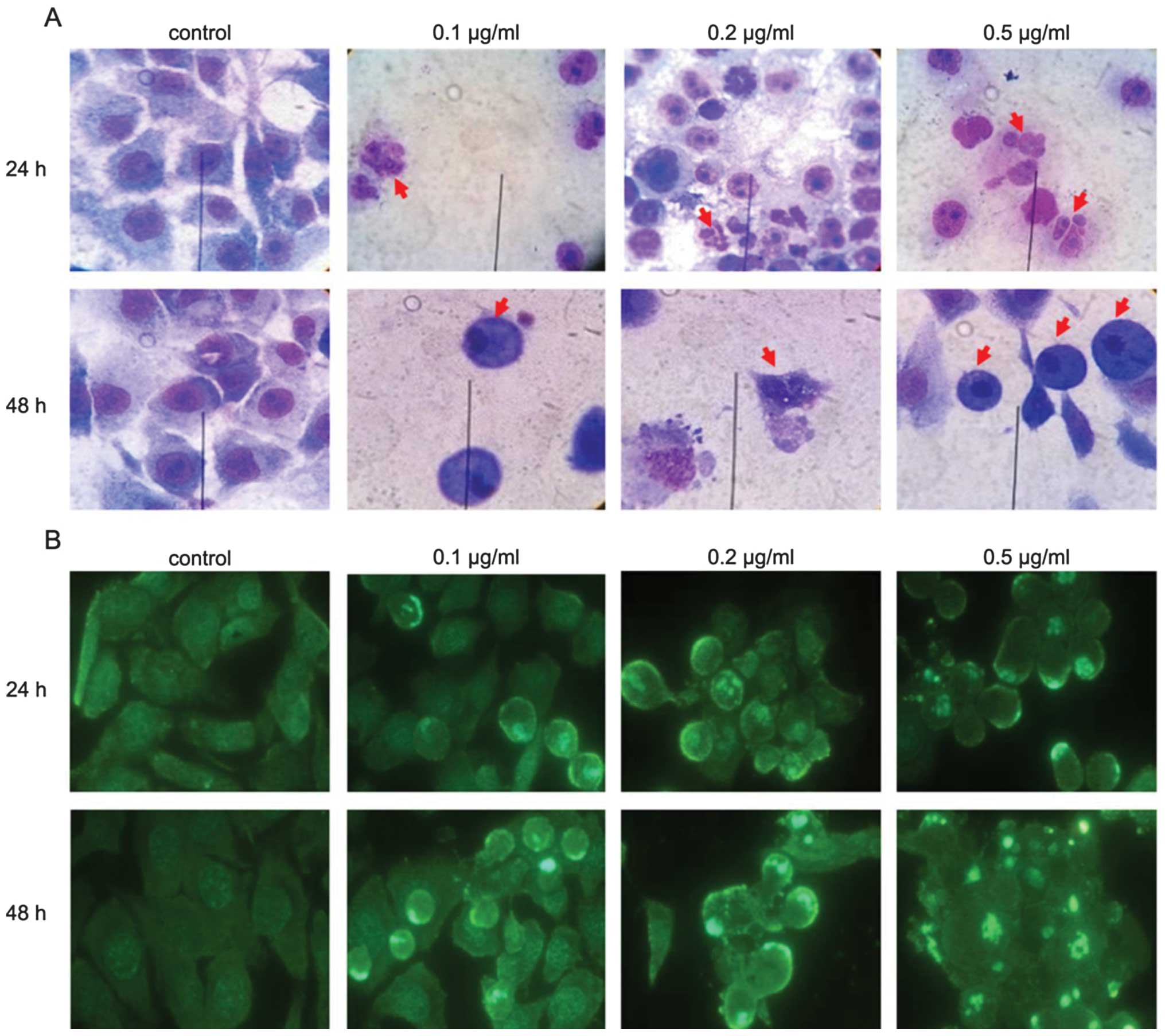
ZGDHu-1 induces apoptosis on PANC-1
cells
To confirm whether the ZGDHu-1-induced inhibition of
PANC-1 cell proliferation was caused by apoptosis, PANC-1 cells
treated with ZGDHu-1 were observed under a microscope. After the
treatment of ZGDHu-1, the morphologic features of apoptosis were
significantly changed, such as cell shrinkage, nuclear chromatin
condensation and fragmentation as well as the formation of
apoptotic bodies, particularly evident in higher concentrations of
ZGDHu-1 (Fig. 2A and B).
Additionally, when using the Annexin V/Dead Cell Apoptosis kit to
detect the early apoptotic effect of ZGDHu-1 on PANC-1 cells, we
found that the apoptotic effect of ZGDHu-1 on PANC-1 cells occurred
in a dose-dependent manner. In addition, the early apoptotic rates
of PANC-1 cells were 3.6, 12.6, 17.1 and 18.5%, while the total
apoptotic rates were 4.5, 14.2, 18.6 and 20.8% at concentrations of
0, 0.1, 0.2 and 0.5 μg/ml of ZGDHu-1 (Fig. 3). Thus, these findings indicated
that ZGDHu-1 can induce apoptosis of PANC-1 cells and the apoptotic
rates of PANC-1 cells were elevated with the increasing
concentration of ZGDHu-1 treatment.
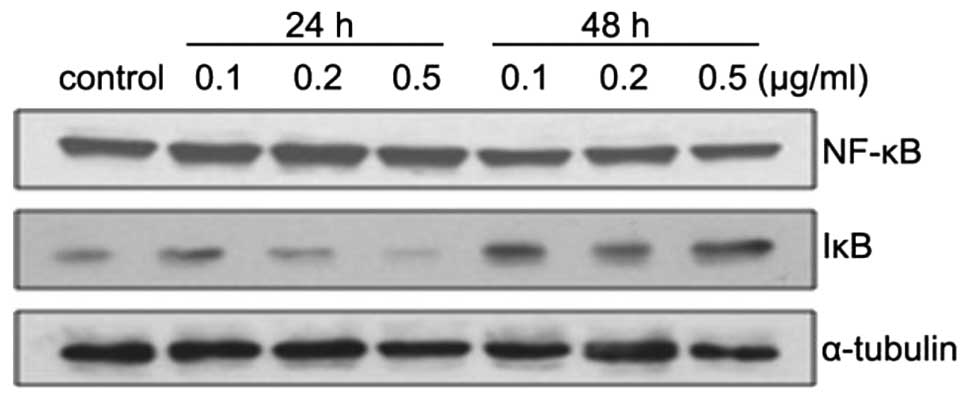
ZGDHu-1 upregulates the expression level
of IκB
NF-κB is a vital nuclear transcription factor that
regulates many genes and participates in cell proliferation,
apoptosis, invasion and differentiation (14). In this study, we observed that the
expression of NF-κB was not significantly changed while its
inhibitor IκB was strongly increased with the addition of ZGDHu-1
in a dose-dependent manner especially after treatment with ZGDHu-1
for 48 h (Fig. 4).
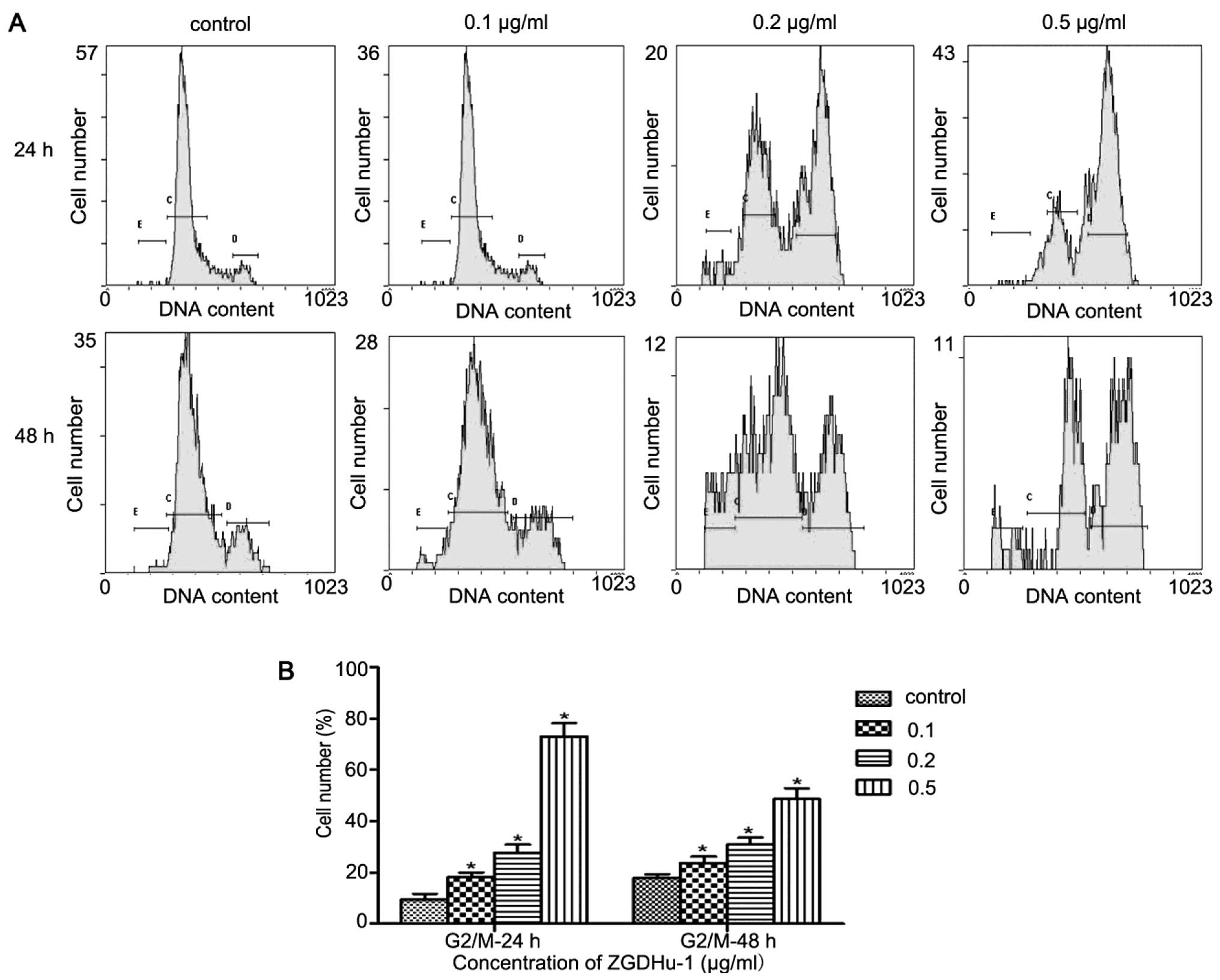
ZGDHu-1 induces cell cycle arrest at G2/M
phase and modulates cell cycle-related protein levels in PANC-1
cells
To determine whether cell-cycle changes are involved
in ZGDHu-1-induced cell apoptosis, we detected cell-cycle phase
distribution by flow cytometry. Following the treatment of PANC-1
cells with different concentrations of ZGDHu-1 (0, 0.1, 0.2 and 0.5
μg/ml) for 24 and 48 h, we found that the PANC-1 cells decreased at
G0/G1 phase and increased at G2/M phase with the increasing
concentrations of ZGDHu-1 (Fig. 5).
The percentages of cells at G2/M phase were 9.6, 18.25, 27.7 and
73.01% at 24 h and 17.9, 23.7, 30.9 and 48.62% at 48 h,
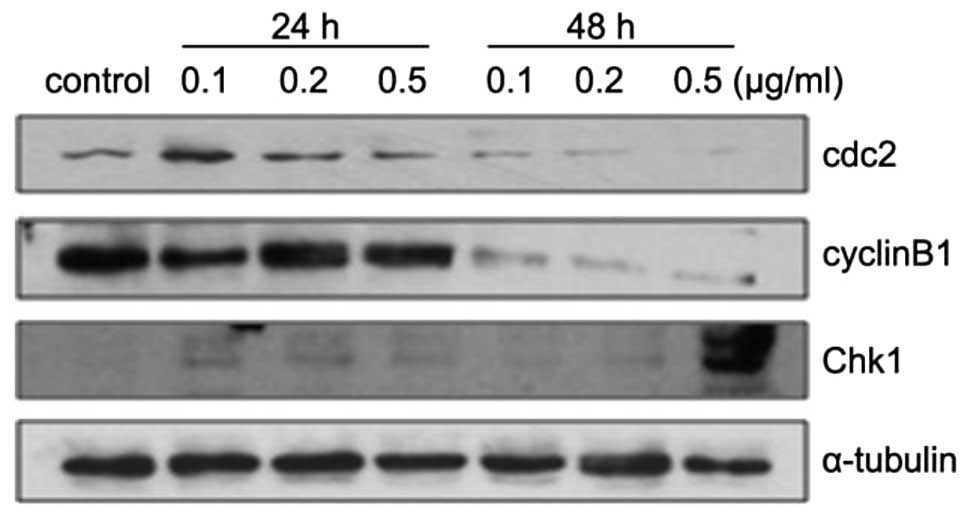
respectively. Furthermore, to investigate the molecular mechanism
for ZGDHu-1-induced G2/M arrest in PANC-1 cells, we detected the
expression levels of cell cycle-related proteins such as Chk1,
cyclinB1 and cdc2. The results showed that the protein levels of
cyclinB1 and cdc2 were downregulated, while the expression of Chk1
was increased following treatment with ZGDHu-1, especially when
treated at 48 h (Fig. 6).
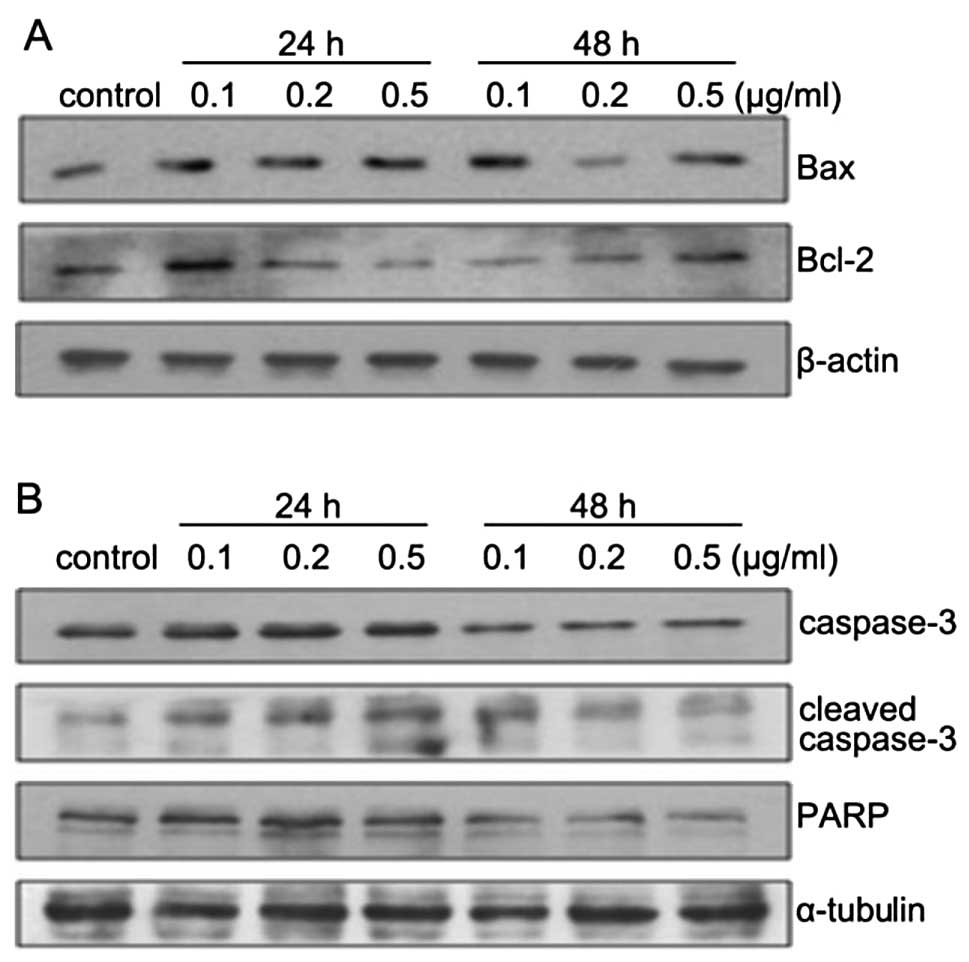
ZGDHu-1-induced PANC-1 cell apoptosis via
mitochondrial apoptosis pathway and the activation of
caspase-3
The mitochondrial pathway is important in the
apoptotic process. To examine whether ZGDHu-1 induced apoptosis of
PANC-1 cells through this pathway, we analyzed the expression of
Bcl-2 and Bax protein after treatment with ZGDHu-1 at
concentrations of 0, 0.1, 0.2 and 0.5 μg/ml for 24 and 48 h. The
result indicated that the expression of Bax was increased in a
dose-dependent manner, whereas Bcl-2 expression was decreased
(Fig. 7A). In addition, we found
that the expression of pro-caspase-3, PARP (the substrate of
caspase-3) was downregulated while cleaved-caspase-3 was
significantly upregulated after treatment with ZGDHu-1 (Fig. 7B). These results suggest that
ZDGHu-1-induced cell apoptosis occurred through a mitochondrial
pathway and the activation of caspase-3.
Discussion
Pancreatic cancer is an aggressive malignant disease
due to the lack of early diagnosis and treatment options (15). Moreover, pancreatic cancer is one of
the most intrinsic chemical resistance tumors. Common intrinsic or
acquired chemical resistance become the main causes of failure for
pancreatic cancer therapy (16).
Identification of a new drug that is highly sensitive and has fewer
side effects may lead to effective treatments for pancreatic
cancer.
ZGDHu-1 has been proven to repress the proliferation
of myeloid leukemia (12) and lung
cancer (13). Pro-apoptotic effects
were demonstrated on those carcinoma cells and were independent of
its anti-proliferative action. In this study, we found that ZGDHu-1
inhibited the proliferation of PANC-1 cells in a time- and
dose-dependent manner with the treatment of ZGDHu-1 for 24, 48 and
72 h. The apoptotic effects of ZGDHu-1 on PANC-1 cells were
enhanced based on the cytomorphology observation and Annexin V/PI
staining. Overall, our study has demonstrated that ZGDHu-1
effectively suppressed cell viability and induced apoptosis of
PANC-1 cells.
The molecular mechanism of ZGDHu-1 regarding which
pathway to induce apoptosis in cancer cells was widely studied. For
example, it was demonstrated that the NF-κB pathway may play a key
role in ZGDHu-1-induced apoptosis in myeloid leukemia (17), while the results of another study
indicated that ZGDHu-1 inhibited lung cancer cells by upregulating
the expression levels of pro-apoptotic proteins, such as Bax and
p53 (13). Although ZGDHu-1 may
induce cell-cycle arrest in cancer cells, the antitumor effects of
ZGDHu-1 on pancreatic cancer as well as the associated molecular
mechanisms have yet to be elucidated. We have shown that ZGDHu-1
can induce PANC-1 cell apoptosis and G2/M phase arrest.
NF-κB is composed of two subunits and is normally
sequestered in the cytoplasm by its inhibitor proteins, IκB
(18). When the cells are exposed
to NF-κB-activating signals, it can induce the phosphorylation of
IκB, targeting them for rapid degradation through a
ubiquitin-proteosome pathway, releasing NF-κB to the nucleus, where
it binds to specific sequences in the promoter regions of genes
(19). Our results have shown that
ZGDHu-1 mainly inhibited the growth of PANC-1 cells by upregulating
the expression level of IκB, whereas the expression of NF-κB did
not change significantly following treatment of ZGDHu-1 because of
its higher expression in the cytoplasm.
The cell cycle is an intricate process to ensure the
replication of cells that can be divided into four stages: G1
phase, S phase (DNA synthesis), G2 phase, and M phase (Mitosis). In
this study, we found that ZGDHu-1 arrested the cell cycle of PANC-1
cells at G2/M phase by using flow cytometry. Previous findings show
that the cell-cycle progression depends on highly ordered events
controlled by cyclins and cyclin-dependent kinases (Cdks). For
example, the expression of p21 induced by p53 exerts an important
role in the G1/S checkpoint by inactivating Cdk2 in response to DNA
damage. However, the transition from G2 to Mitosis principally
depends on the activity of the cyclin B1/cdc2 (Cdk1) complex
(20). Chk1, a serine/threonine
kinase, has been proven an important checkpoint kinase in the cycle
arrest through phosphorylating cdc25C on Ser-216 (21). Our study has demonstrated that the
protein levels of cdc2 and cyclinB1 were obviously decreased in a
dose- and time-dependent manner, which blocked the cell cycle
entering into mitosis and induce the apoptosis of PANC-1 cells.
There are two main apoptotic pathways that have been
extensively investigated: the intrinsic (mitochondrial) and the
extrinsic (death receptor) pathways (22). The intrinsic pathway is activated by
a myriad of stress signals, such as DNA damage, leading to the
mitochondrial outer membrane permeabilization and the release of
apoptotic proteins, such as cytochrome c and second
mitochondrial-derived activator of caspases from mitochondria into
cytoplasm. The intrinsic pathway, leads to the release of
cytochrome c, which is suppressed by Bcl-2 and promoted by
Bad (23,24). However, the extrinsic pathway is
activated by the binding of death ligands to the cell surface
receptors (TNF receptor superfamily), leading to a complex signal
transduction into the cytoplasm and activating the initiator
caspase-8 and the subsequent executioner caspases.
Caspase-3, the executioner caspase, is the
downstream activator of extrinsic and intrinsic apoptotic pathways
(25). Furthermore, it is well
known that antitumor agents can target caspases and their
substrates such as PARP-1 in apoptotic pathways in cancer cells,
while the PARP-1-mediated pathway is a major mechanism for DNA
repair in cancer cells (26).
According to the results of western blotting, we found that the
expression of Bax was increased, whereas Bcl-2 was decreased in
PANC-1 cells treated with ZGDHu-1 compared with the control group
in the present study. In addition, for the expression levels of
pro-caspase-3, PARP was downregulated while the cleaved-caspase-3
was greatly upregulated with the increasing concentrations of
ZGDHu-1. Our results suggest that ZDGHu-1 induced cell apoptosis
through the mitochondrial pathway.
In conclusion, the present study has demonstrated
that ZGDHu-1 elaborates the cytotoxic effects against PANC-1 cells
by inducing apoptosis and arresting the cell cycle at G2/M phase.
The upregulation of pro-apoptotic proteins and activation of
caspase-3 are the key factors that contribute to the inhibitory
effects of ZGDHu-1. In the future, more and more advancing studies
about the effect of ZGDHu-1 in pancreatic cancer will be confirmed,
which may be useful with the development of ZGDHu-1 as a new drug
for cancer treatment.
Acknowledgements
This study was supported by key platform funded
projects from the Zhejiang Province Health Bureau (no. 2013ZDA005)
and a fund from the Zhejiang Province Health Bureau (no.
2012KYA015).
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics for Hispanics/Latinos, 2012. CA Cancer J Clin.
62:283–298. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen WQ, Liang D, Zhang SW, Zheng RS and
He YT: Pancreatic cancer incidence and mortality patterns in China,
2009. Asian Pac J Cancer Prev. 14:7321–7324. 2013. View Article : Google Scholar
|
|
4
|
Ma C, Jiang YX, Liu SZ, et al: Trend and
prediction on the incidence of pancreatic cancer in China. Zhonghua
Liu Xing Bing Xue Za Zhi. 34:160–163. 2013.(In Chinese). PubMed/NCBI
|
|
5
|
Hidalgo M: Pancreatic cancer. N Engl J
Med. 362:1605–1617. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kamangar F, Dores GM and Anderson WF:
Patterns of cancer incidence, mortality, and prevalence across five
continents: defining priorities to reduce cancer disparities in
different geographic regions of the world. J Clin Oncol.
24:2137–2150. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Neoptolemos JP, Dunn JA, Stocken DD, et
al: Adjuvant chemoradiotherapy and chemotherapy in resectable
pancreatic cancer: a randomised controlled trial. Lancet.
358:1576–1585. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Van Laethem JL, Hammel P, Mornex F, et al:
Adjuvant gemcitabine alone versus gemcitabine-based
chemoradiotherapy after curative resection for pancreatic cancer: a
randomized EORTC-40013-22012/FFCD-9203/GERCOR phase II study. J
Clin Oncol. 28:4450–4456. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Moore MJ, Goldstein D, Hamm J, et al:
Erlotinib plus gemcitabine compared with gemcitabine alone in
patients with advanced pancreatic cancer: a phase III trial of the
National Cancer Institute of Canada Clinical Trials Group. J Clin
Oncol. 25:1960–1966. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rao GW and Hu WX: Synthesis, X-ray
crystallographic analysis, and antitumor activity of
1-acyl-3,6-disubstituted phenyl-1,4-dihydro-1,2,4,5-tetrazines.
Bioorg Med Chem Lett. 15:3174–3176. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rao GW and Hu WX: Synthesis, structure
analysis, and antitumor activity of
3,6-disubstituted-1,4-dihydro-1,2,4,5-tetrazine derivatives. Bioorg
Med Chem Lett. 16:3702–3705. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhou YL, Lü YP, Hu WX, et al:
ZGDHu-1-inducing apoptosis of SHI-1 leukemia cells and its
molecular mechanism. Zhongguo Shi Yan Xue Ye Xue Za Zhi.
15:483–489. 2007.(In Chinese). PubMed/NCBI
|
|
13
|
Zhou YL, Hu WX, Lü YP, et al: Effect of
ZGDHu-1 on proliferation and apoptosis of A549 cells in vitro and
antitumor activity in vivo. Yao Xue Xue Bao. 42:26–34. 2007.(In
Chinese). PubMed/NCBI
|
|
14
|
Tchoghandjian A, Jennewein C, Eckhardt I,
Momma S, Figarella-Branger D and Fulda S: Smac mimetic promotes
glioblastoma cancer stem-like cell differentiation by activating
NF-κB. Cell Death Differ. 21:735–747. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Loos M, Kleeff J, Friess H and Büchler MW:
Surgical treatment of pancreatic cancer. Ann NY Acad Sci.
1138:169–180. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Neoptolemos JP, Stocken DD, Bassi C, et
al: Adjuvant chemotherapy with fluorouracil plus folinic acid vs
gemcitabine following pancreatic cancer resection: a randomized
controlled trial. JAMA. 304:1073–1081. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang YX, Zhou YL, Xu F and Qiu LN: Effect
of ZGDHu-1 on proliferation of K562 and K562/Adr and the expression
of NF-κB protein. Bull Med Res. 40:50–54. 2011.(In Chinese).
|
|
18
|
Abraham E: NF-kappaB activation. Crit Care
Med. 28(Suppl 4): N100–N104. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hayden MS and Ghosh S: Shared principles
in NF-kappaB signaling. Cell. 132:344–362. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nyberg KA, Michelson RJ, Putnam CW and
Weinert TA: Toward maintaining the genome: DNA damage and
replication checkpoints. Annu Rev Genet. 36:617–656. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bartek J and Lukas J: Chk1 and Chk2
kinases in checkpoint control and cancer. Cancer Cell. 3:421–429.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Arlt A, Muerkoster SS and Schafer H:
Targeting apoptosis pathways in pancreatic cancer. Cancer Lett.
332:346–358. 2013. View Article : Google Scholar
|
|
23
|
Bai L and Wang S: Targeting apoptosis
pathways for new cancer therapeutics. Annu Rev Med. 65:139–155.
2014. View Article : Google Scholar
|
|
24
|
Shamas-Din A, Kale J, Leber B and Andrews
DW: Mechanisms of action of Bcl-2 family proteins. Cold Spring Harb
Perspect Biol. 5:a0087142013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wen X, Lin ZQ, Liu B and Wei YQ:
Caspase-mediated programmed cell death pathways as potential
therapeutic targets in cancer. Cell Prolif. 45:217–224. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Langelier MF, Planck JL, Roy S and Pascal
JM: Structural basis for DNA damage-dependent
poly(ADP-ribosyl)ation by human PARP-1. Science. 336:728–732. 2012.
View Article : Google Scholar : PubMed/NCBI
|