Introduction
Methotrexate (MTX) is the most widely known
antifolate successfully used in oncology for a long time. MTX is
particularly effective in the treatment of acute lymphoblastic
leukemia, non-Hodgkin lymphoma, breast carcinoma, lung carcinoma,
osteosarcoma, choriocarcinoma, and some neuroectodermal tumors
(1). Although MTX has been included
in therapeutic protocols for more than 60 years, its dosage as well
as administration schedules are still being optimized.
The most important effect of MTX is based on the
inhibition of dihydrofolate reductase (DHFR), which blocks the
reduction of folic acid and, consequently, folic acid metabolism.
When the concentration of MTX exceeds the binding capacity of DHFR,
all available molecules of tetrahydrofolate (THF) are gradually
depleted in the cell, and the synthesis of purine and pyrimidine
precursors, which are necessary for synthesis of nucleic acids, is
reduced (2).
Although MTX is included in many standard
therapeutic regimens, its substantial toxicity limits its wider
use, particularly in pediatric oncology. The cytotoxic effects of
high-dose MTX (HD-MTX) on normal somatic cells could be reduced by
the administration of an antidote, with the most frequent being
leucovorin (LV). Another possibility of an MTX schedule is the
repeated administration of low-dose MTX (LD-MTX) without LV
(3,4).
Nevertheless, there is a fear in clinical practice
that HD-MTX chemotherapy can induce drug resistance, resulting in a
reduced treatment effect (5). The
primary and the most frequent mechanism of resistance to MTX is
caused by defects in reduced folate carrier (RCF)-mediated
transport, which are caused by mutations in the RCF gene or
by the downregulation of its expression (6). Other well-described mechanisms of MTX
resistance include the overexpression of DHFR or thymidylate
synthase (TYMS) or mutations in genes encoding these enzymes,
decreasing their affinity for antifolates. Another important aspect
in resistance to MTX is defective polyglutamylation, which
substantially reduces the cytotoxicity of MTX. Reductions in MTX
polyglutamylation usually result from the decreased expression of
folylpolyglutamate synthetase (FPGS) or from inactivating mutations
in the FPGS gene, as well as from the increased expression
of folylpolyglutamate hydrolase (FPGH) (7).
Our study focused on a detailed analysis of MTX
effects in cell lines derived from two types of pediatric solid
tumors, medulloblastoma and osteosarcoma, which were chosen on the
basis of their different histogenetic origin and because MTX is
typically included in therapeutic protocols for both. The main aim
of this study was to analyze the effects of treatment with MTX at
concentrations comparable to the MTX plasma levels in patients
treated with high-dose or low-dose MTX. Furthermore, an extremely
important part of the treatment with high-dose MTX in clinical
practice is the administration of LV as an antidote to reduce MTX
toxicity in normal cells. Thus, the combined application of MTX and
LV was also included in our experiments. An analysis of the
expression of genes involved in the mechanisms of resistance to MTX
was the final component of our study; the results helped us to
elucidate the mechanisms of the various responses to MTX among the
examined cell lines.
Materials and methods
Cell lines
Two reference cell lines and two cell lines derived
in our laboratory were used in this study. Daoy (ATCC HTB-186™)
medulloblastoma and Saos-2 (ATCC HTB-85™) osteosarcoma cell lines
were purchased from the American Type Culture Collection (Manassas,
VA, USA). MBL-02 is an in-house cell line derived previously from a
biopsy sample obtained from a 7-year-old girl suffering from
desmoplastic medulloblastoma (8).
The OSA-08 cell line was newly derived from a biopsy sample
obtained from an 11-year-old boy surgically treated for
conventional osteosarcoma. The Research Ethics Committee of the
School of Medicine (Masaryk University, Brno, Czech Republic)
approved the study protocol, and a written statement of informed
consent was obtained from each patient or his/her legal
guardian.
Cell culture
Cells were grown in Dulbecco’s modified Eagle’s
medium (DMEM) supplemented with 10% (Daoy and Saos-2) or 20%
(MBL-02 and OSA-08) fetal bovine serum, 100 IU/ml penicillin, 100
mg/ml streptomycin, and 2 mM glutamine. In addition, the medium for
the Daoy cells also contained 1% nonessential amino acids (all cell
culture reagents were purchased from PAA, Linz, Austria).
Experiments with leucovorin (LV) application were performed in
folate-free DMEM (both reagents were purchased from Sigma-Aldrich,
St. Louis, MO, USA). Cell culture was performed under standard
conditions at 37°C in a humidified atmosphere containing 5%
CO2.
Chemicals
MTX (Sigma) was prepared as a stock solution at a
concentration of 20 mM in 1 M NaOH (Sigma). This stock solution was
diluted in DMEM or folate-free DMEM to obtain the final
concentrations used in the experiments. For determination of the
IC50 value, 7 different concentrations of MTX ranging
from 1×10−4 to 1×102 μM were tested.
For all other experiments, concentrations of 0.1, 1, 10 and 40
μM MTX were used; these concentrations are in the range of
MTX plasma levels reached in patients suffering from cancer. The
maximum used concentration of MTX, i.e., 40 μM, is
comparable with the peak of the MTX plasma concentration achieved
during HD-MTX treatment of pediatric solid tumors (4). LV was dissolved in deionized water to
prepare a 1 mM stock solution. LV at final concentrations of 10 and
100 nM was prepared in folate-free DMEM.
MTT assay
To evaluate cell proliferation, an MTT assay to
detect the activity of mitochondrial dehydrogenases in living cells
was used; 96-well plates were seeded with 1×104
cells/well in 200 μl of culture medium, and the cells were
allowed to adhere overnight. The medium was then removed and a new
medium containing the selected concentrations of MTX described
above or control MTX-free medium was added. The plates were
incubated under standard conditions, and LV at the chosen
concentrations was added after 42 h. To evaluate changes in cell
proliferation, medium with reagents was removed and replaced by 200
μl of DMEM containing
3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT)
at 0.5 mg/ml. The plates were then incubated at 37°C for 2.5 h.
Subsequently, the medium was carefully removed, and the formazan
crystals were dissolved in 200 μl of DMSO. The absorbance at
570 nm with a reference absorbance at 620 nm was measured using the
Sunrise Absorbance Reader (Tecan, Männedorf, Switzerland).
RT-PCR
Differences in the expression of MTX
resistance-related genes in the cell lines under standard
conditions were evaluated using RT-PCR. Total RNA was extracted
using the GenElute™ Mammalian Total RNA Miniprep kit (Sigma), and
its concentration and integrity were determined
spectrophotometrically. For all samples, equal amounts of RNA
(i.e., 25 ng of RNA per 1 μl of total reaction volume) were
reverse transcribed into cDNA using M-MLV (Top-Bio, Prague, Czech
Republic) and oligo dT (Qiagen, Hilden, Germany) priming. PCR was
carried out in 25 μl reactions containing 12.5 μl of
PPP master mix, 0.5 μl of PCR enhancer (both from Top-Bio),
0.5 μM of each primer and 5 μl of diluted cDNA. The
primers used for RFC1, DHFR, TYMS,
FPGS, FPGH and HSP90AB1 are described in
Table I. A total of 10 μl of
the PCR product was loaded onto a 2% agarose gel stained with
Midori Green (Nippon Genetics, Dueren, Germany) and examined after
electrophoresis. The optical density was stained and quantified
using ImageJ software (9). The data
were normalized to HSP90AB1 expression.
 | Table ISequences of the primers used for
RT-PCR. |
Table I
Sequences of the primers used for
RT-PCR.
| Gene | Primer
sequences | Product (bp) |
|---|
| RFC1 | F:
5′-GCGGGCTTCGTGAAGATC-3′ | 330 |
| R:
5′-CTGGAACTGCTTGCGGAC-3′ | |
| DHFR | F:
5′-CAGAACATGGGCATCGGCAAGAACG-3′ | 328 |
| R:
5′-AAACAGAACTGCCACCAACTATCCA-3′ | |
| TYMS | F:
5′-CGGGAGACATGGGCCTCGGT-3′ | 353 |
| R:
5′-GCATCCAGCCCAACCCCTAA-3′ | |
| FPGS | F:
5′-CACTGGGACGAAGGGGAA-3′ | 322 |
| R:
5′-GTCATAAGCCCCGCCAAT-3′ | |
| FPGH | F:
5′-AAAGTACTTGGAGTCTGCAGGTGC-3′ | 327 |
| R:
5′-TGCAATTGACCTCCAGTGAAGTTCA-3′ | |
| HSP90AB1 | F:
5′-CGCATGAAGGAGACACAGAA-3′ | 169 |
| R:
5′-TCCCATCAAATTCCTTGAGC-3′ | |
Flow cytometry
To evaluate changes in the cell cycle,
1.2×105 cells were seeded in 25 cm2 Petri
dishes and allowed to attach overnight. The cells were then treated
with MTX for 3 or 6 days. Both the detached and adherent cells were
harvested together, fixed with 70% ethanol and stained with
Vindelov’s solution [0.01 M Tris, 10 μg/ml RNase, 50
μg/ml PI and 1 mM NaCl (all from Sigma)] at 37°C for 30
min.
To quantify the rate of apoptosis, 1×106
cells were seeded in 75 cm2 Petri dishes and allowed to
attach overnight. The cells were treated with MTX for 1 or 3 days.
Both the detached and adherent cells were harvested together, fixed
with 3% paraformaldehyde (Sigma) at room temperature for 30 min,
permeabilized in 0.2% Triton X-100 (Sigma) for 1 min, and incubated
with 2% BSA (PAA) for 10 min to block nonspecific antibody binding.
The cells were then treated with a rabbit polyclonal anti-cleaved
caspase-3 (Asp-175) primary antibody (dilution 1:250, cat. no.
9661; Cell Signaling Technology, Beverly, MA, USA) at 37°C for 60
min. After washing with PBS twice, goat anti-rabbit IgG conjugated
with Alexa Fluor® 488 (dilution 1:300, cat. no. A-11008;
Life Technologies, Carlsbad, CA, USA) was applied at 37°C for 45
min.
The BD FACSVerse™ flow cytometer with BD FACSuite
software (Beckton Dickinson, San Jose, CA, USA) was employed to
analyze both the cell cycle and frequency of caspase-3-positive
cells at the intervals specified above. Ten thousand events per
sample were evaluated in all experiments.
Results
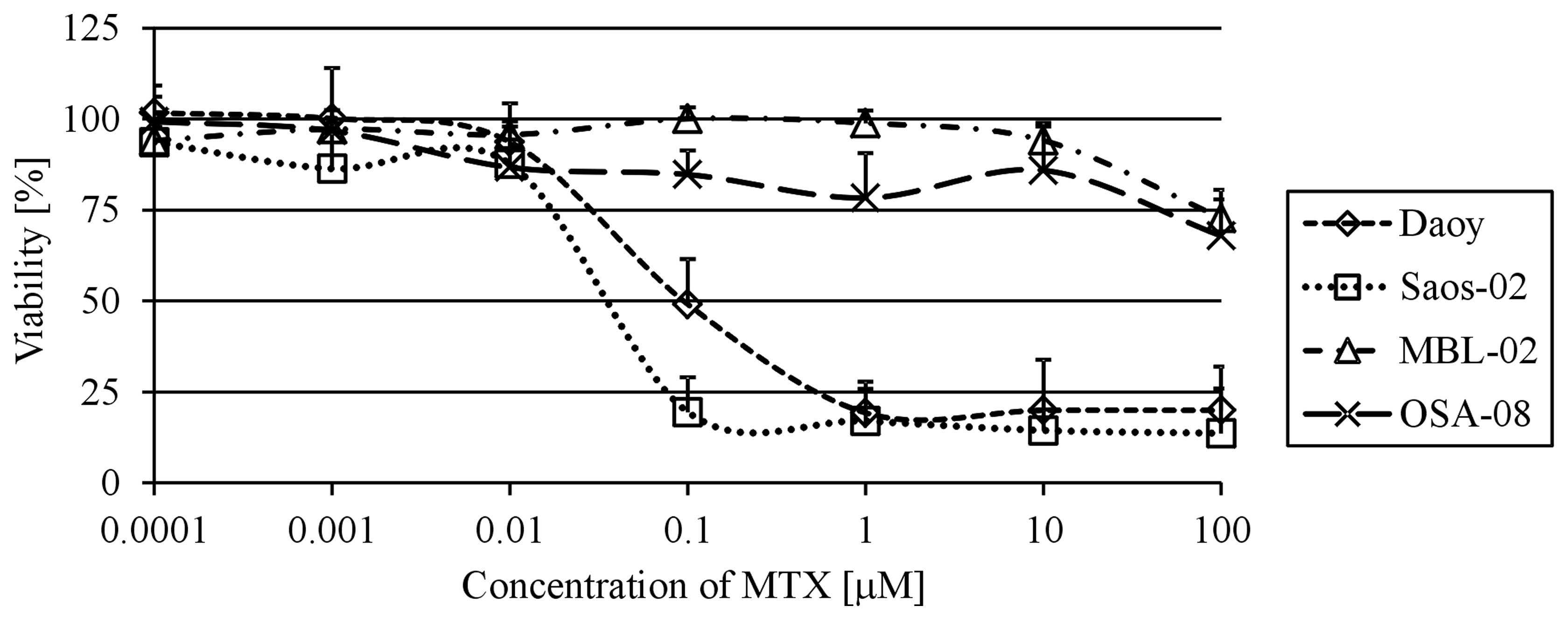
Determination of MTX IC50
To confirm that the Daoy and Saos-2 reference cell
lines are useful models for the examination of the MTX effects on
medulloblastoma and osteosarcoma cells, the IC50 values
were first determined. Using the MTT assay, we analyzed cell
viability at day 6 of MTX treatment in a range of MTX
concentrations from 1×10−4 to 1×102
μM. Both of these cell lines showed a very similar
IC50 value: 9.5×10−2 μM for Daoy cells
and 3.5×10−2 μM for Saos-2 cells (Fig. 1). In contrast, neither the MBL-02
medulloblastoma nor the OSA-08 osteosarcoma patient-derived cell
lines reached the IC50 value within the concentrations
of MTX used. The highest concentration of MTX used for experiments
with the reference cell lines, i.e., 100 μM, only led to 27
and 32% decreases in viability when compared with the untreated
MBL-02 and OSA-08 cells, respectively.
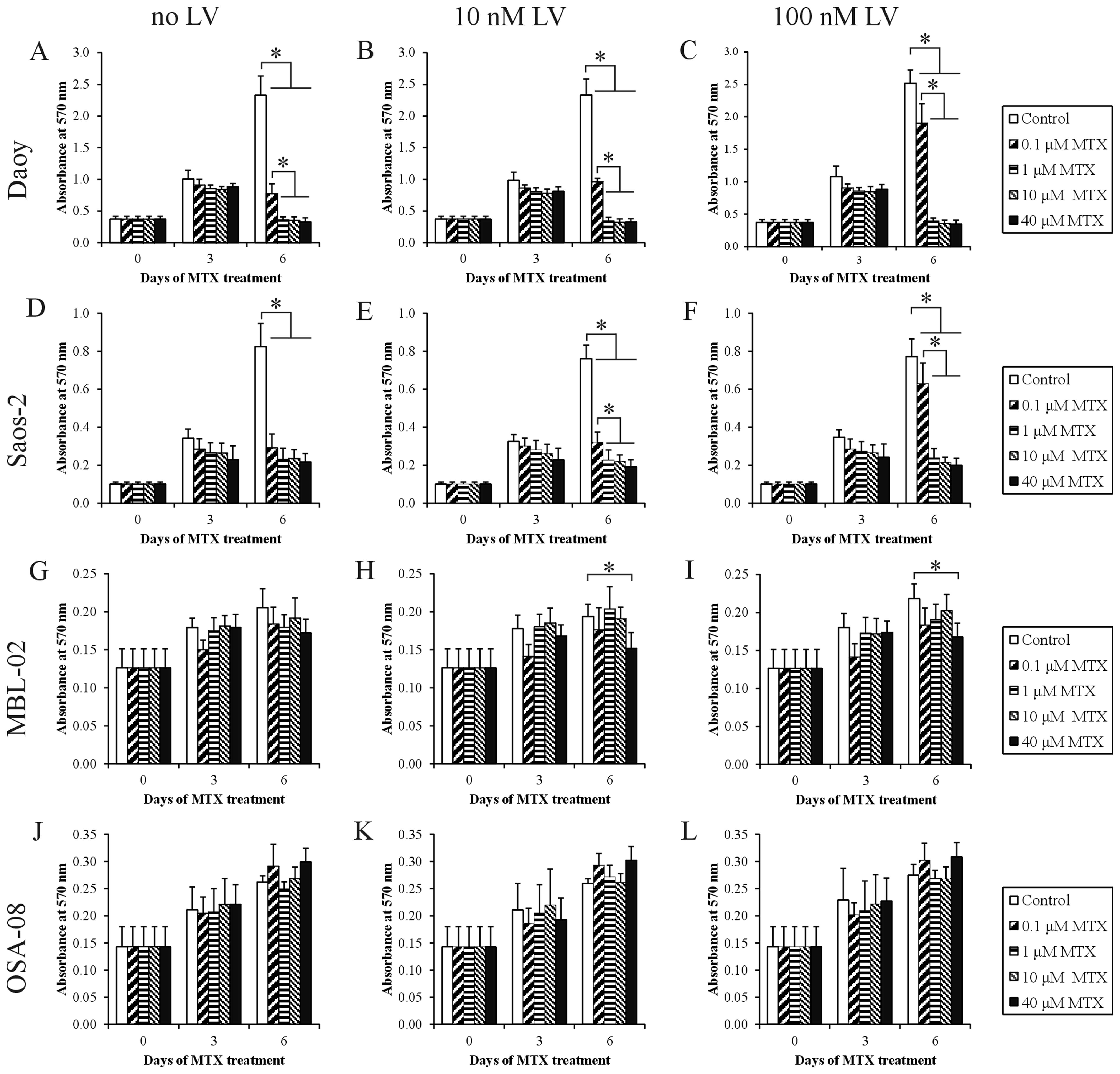
Effect of MTX and ‛leucovorin rescue’
treatment on cell proliferation
To analyze the effects of MTX on cell proliferation,
concentrations corresponding to MTX plasma levels were used. Daoy
(Fig. 2A) and Saos-2 (Fig. 2D) cell lines showed evident
cytostatic effects at day 6 of treatment with MTX at all the chosen
concentrations. For Saos-2 cells, no statistically significant
differences were observed among all the different MTX treatments.
It was also apparent that treatment with 0.1 μM MTX
decreased the proliferation of Daoy cells to a significantly lesser
extent than the other MTX concentrations. Both the MBL-02 and
OSA-08 patient-derived cell lines did not show any marked decrease
in the number of viable cells within the concentration interval
from 0.1 to 40 μM. Nevertheless, the MBL-02 medulloblastoma
cell line (Fig. 2G) appeared to be
more sensitive than the OSA-08 osteosarcoma cell line in terms of
cell viability (Fig. 2J).
To determine whether the application of LV
influences the observed cytostatic effects of MTX, we added LV at
two different concentrations, 10 and 100 nM, to the cultivation
medium at 42 h after treatment with MTX. The application of 10 nM
LV resulted in a slight but statistically significant increase in
the proliferation activity of Daoy (Fig. 2B) and Saos-2 (Fig. 2E) cells pretreated with 0.1
μM MTX. In contrast, the use of an elevated concentration of
LV, i.e., 100 nM, caused a statistically significant increase in
proliferation activity and an inhibition of MTX action in both cell
lines pretreated with 0.1 μM MTX (Fig. 2C and F). The cytostatic effects of
higher concentrations of MTX were not affected by LV in these cell
lines, and the MTX-pretreated in-house cell lines did not respond
to the application of LV (Fig. 2H, I, K
and L).
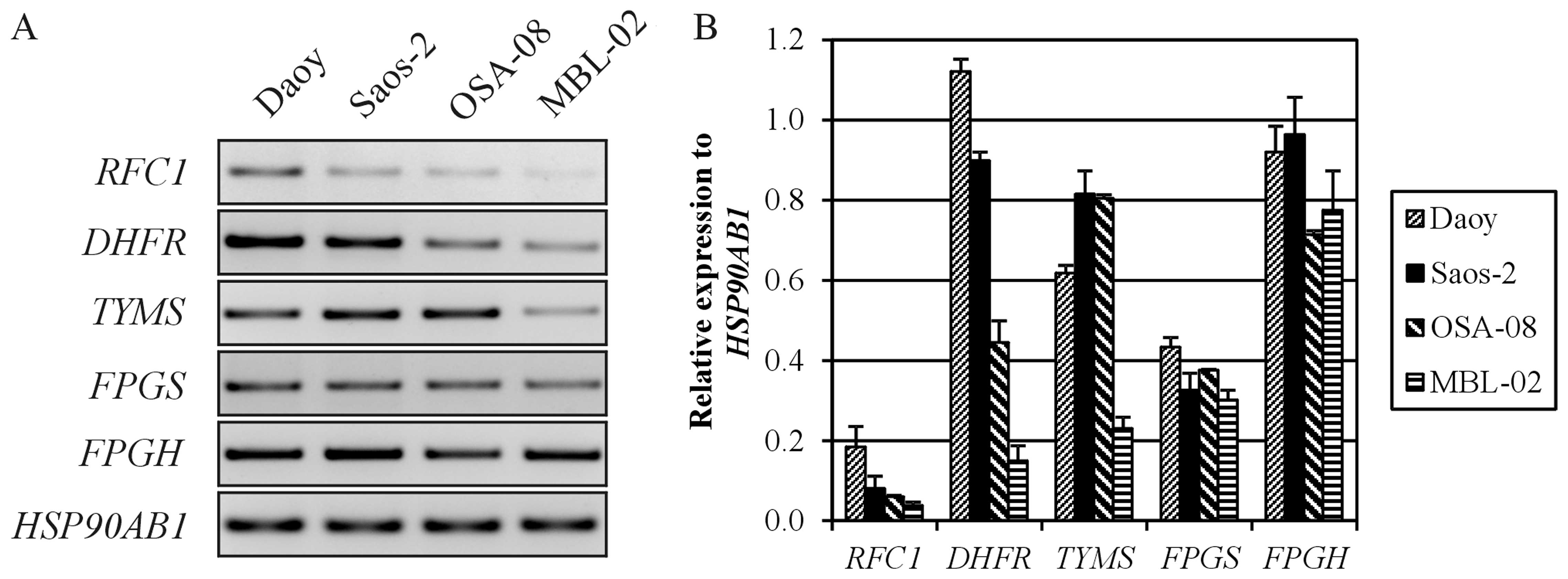
Expression of MTX resistance-related
genes
To understand the strong differences in MTX toxicity
between our in-house cell lines (MBL-02 and OSA-08) and the
reference cell lines obtained from ATCC (Daoy and Saos-2), we
examined the expression of genes involved in the resistance of
tumor cells to MTX (Fig. 3). For
this RT-PCR expression analysis, we chose genes encoding the main
membrane transporter of MTX, i.e., RFC, two key enzyme targets for
MTX, i.e., DHFR and TYMS, and two enzymes catalyzing the
glutamylation of MTX, i.e., FPGS and FPGH. The Daoy medulloblastoma
cells showed higher relative expression of the RFC1,
DHFR and TYMS genes, whereas the expression of these
genes was very weak in the MBL-02 medulloblastoma cells. In
contrast, both osteosarcoma cell lines displayed similar expression
levels of MTX resistance-related genes, with the exception of
DHFR, the expression level of which was also decreased in
the OSA-08 cells.
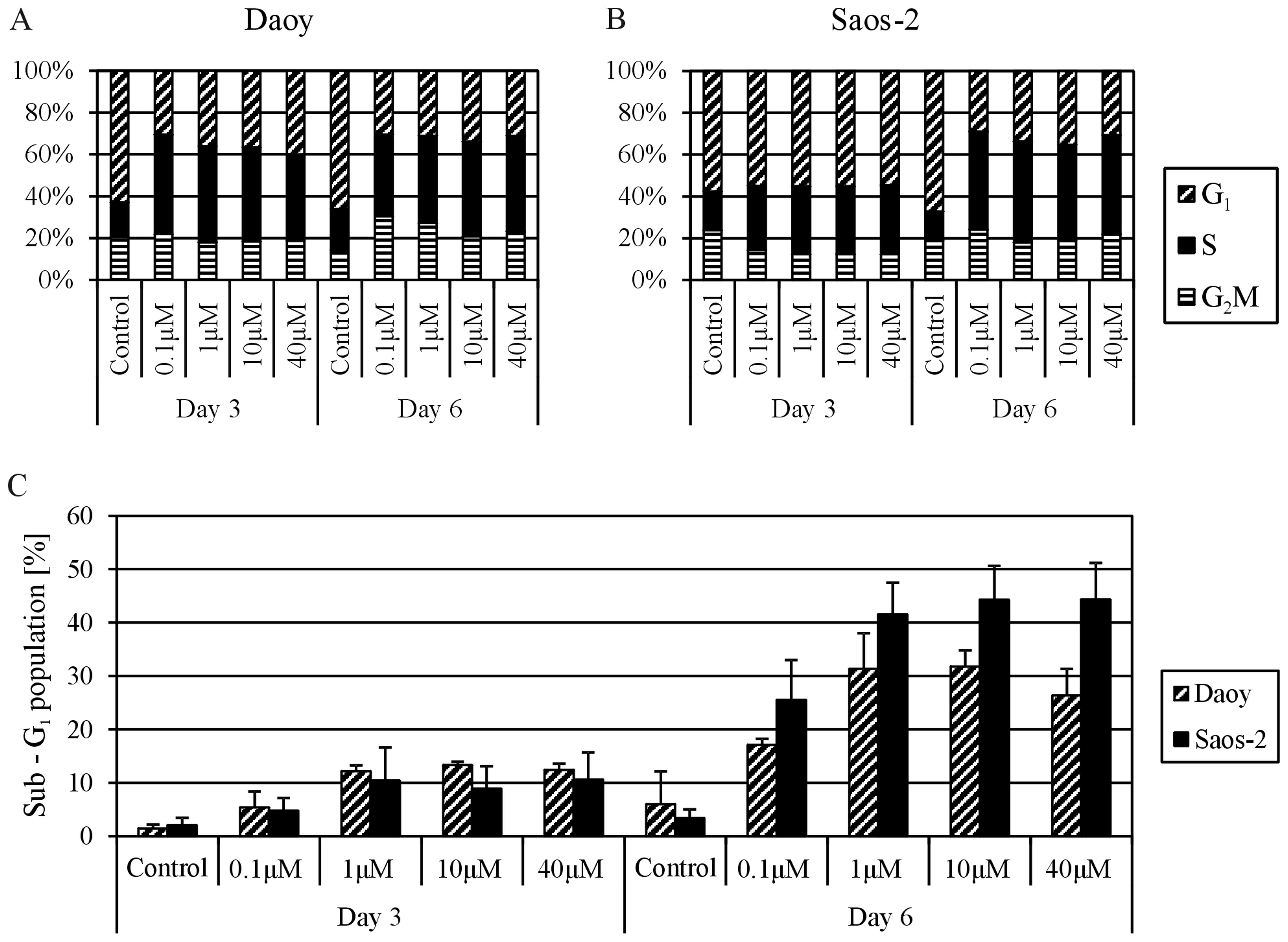
Effect of MTX on the cell cycle and cell
death
Based on the results of previous experiments, both
MTX-responding cell lines, i.e., the Daoy and Saos-2 lines, were
chosen for cell cycle analysis. The cells were treated with
different concentrations of MTX, and the proportions of cells in
sub-G1, G1, S and G2/M phases were
determined using flow cytometry at day 3 and 6 of treatment with
MTX. All MTX concentrations had the same effect on the distribution
of the cell cycle phases; an increase in cells in the S phase was
accompanied by a decrease in cells in the G1 phase in
the treated cell lines compared with the untreated controls.
Importantly, this phenomenon was noted sooner in the Daoy cells, at
day 3 of MTX treatment (Fig. 4A),
but was partially delayed in the Saos-2 cells (Fig. 4B). The analysis of the
sub-G1 population revealed cytotoxic effects of MTX on
both cell lines (Fig. 4C). The
population of cells with reduced DNA content markedly increased at
day 6 of treatment. Furthermore, this trend was more apparent in
the Saos-2 cells; the sub-G1 population was >40% in
the Saos-2 cells and ~30% in the Daoy cells. The cytotoxic effect
of MTX at concentrations ranging from 1 to 40 μM was nearly
the same in both cell lines.
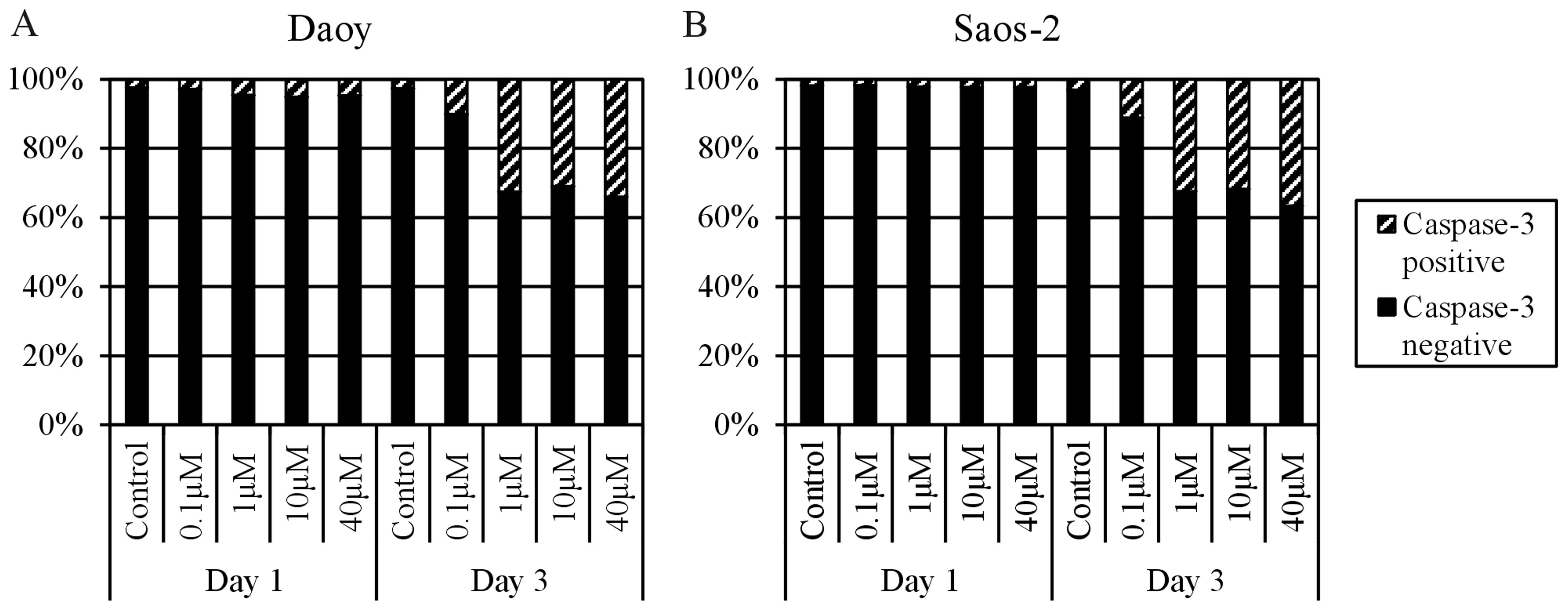
To prove whether the increase in the
sub-G1 proportion after MTX treatment is caused by
apoptosis induction, the MTX-treated cell populations were labeled
with an anti-active caspase-3 antibody at day 1 and 3 of MTX
application. Both cell lines showed a >30% increase in
caspase-3-positive cells at day 3 of treatment with 1, 10 or 40
μM MTX. In contrast, treatment with 0.1 μM MTX led to
a 7–8% increase in caspase-3-positive cells in comparison to the
control cells (Fig. 5).
Discussion
At present, the standard protocol for cancer
treatment with MTX includes the application of HD-MTX, defined as
>1 g/m2 of body surface, in combination with
leucovorin, which enables reaching high plasma concentrations with
enhanced anticancer and cytotoxic effects (10).‛Leucovorin rescue’ is administered in
a specific time schedule after treatment with MTX, usually from 24
to 42 h, to protect noncancerous proliferating cells from the side
effects of MTX. Nevertheless, the toxicity of MTX may induce
myelosuppression, mucositis, nephrotoxicity, hepatotoxicity, and,
in severe cases, multiorgan failure (11). Although MTX has long been an
integral part of many therapeutic regimens, a definite agreement in
regards to MTX dosage and timetables and/or LV treatment is still
lacking (12).
The main aim of this study was to analyze the
effects of MTX on cell lines derived from two types of pediatric
solid tumors, medulloblastoma and osteosarcoma, and to determine
how these cell lines respond to doses of MTX that correspond to
concentrations in a patient’s plasma during administration in
clinical practice.
Daoy medulloblastoma and Saos-2 osteosarcoma cell
lines were chosen as reference cell lines for this study and were
compared to two other cell lines that were derived in our
laboratory from these tumors. The Saos-2 osteosarcoma cells were
apparently more sensitive to treatment with MTX than the Daoy
cells, as revealed by determination of the IC50 value
(Fig. 1); however, the
IC50 value obtained for both of these cell lines was
within a similar range of concentrations, i.e., 10−8 M
MTX. These results are in accordance with those obtained by other
research groups (5,13). The negative effect of MTX on cell
proliferation was clearly evident at day 6 of treatment (Fig. 1) and importantly was in the same
concentration range, from 1 to 40 μM, for both of these cell
lines. In contrast, only a slight cytotoxic effect of 0.1 μM
MTX was able to be reverted by LV (Fig.
2). This finding can be explained by an incomplete inhibition
of DHFR by a concentration lower than 1 μM of MTX, as
described by Assaraf et al (14) on the basis of computational
simulation.
Both of our in-house cell lines, i.e., MBL-02
medulloblastoma and OSA-08 osteosarcoma cell lines, appeared to be
strongly resistant to MTX; 100 μM MTX did not induce a 50%
inhibitory effect in these cells. One of the possible explanations
for this difference is the low proliferation rate of these tumor
cells in comparison with the reference cell lines (15). No observable effect of LV in these
cell lines could be explained by the same mechanisms since
treatment with MTX is targeted to quickly proliferating tumor
cells.
Other possible specific mechanisms of resistance
include impaired transmembrane uptake, alterations in the
expression or activity of target enzymes, or impaired intracellular
polyglutamylation as a determining process of drug efficacy
(6). The RFC1 gene, which
encodes the transmembrane solute carrier and is considered to be a
main MTX transporting pathway to the cytoplasm (16), was only expressed weakly in our
in-house cell lines (Fig. 3).
Consequently, the low levels of RCF may have caused a decrease in
MTX intracellular availability. Conversely, high levels of
DHFR expression were detected in both the Daoy and Saos-2
cells compared with these levels in the in-house cell lines
(Fig. 3). On the one hand,
increased levels of DHFR have been commonly observed in cells
exhibiting an MTX-resistant phenotype (17). On the other hand, this key enzyme
involved in the de novo synthesis of purine and pyrimidine
precursors plays a critical role in cell growth and proliferation,
and its high expression in rapidly proliferating cells is thus
expected (2,18). Although the expression levels of
TYMS in both osteosarcoma cell lines were identical, marked
differences in the expression of this gene were detected between
the medulloblastoma cell lines, with higher levels of TYMS
found in Daoy cells with higher proliferation activity (Fig. 3). The product of the TYMS
gene catalyzes dUMP conversion into dTMP and thus provides the sole
source of deoxythymidylate for DNA biosynthesis (6). In fact, ectopic TYMS expression has
been shown to promote cell proliferation in vitro, and the
high expression of TYMS in tumor tissue is also associated with
poor clinical outcome in some types of cancers (19). Another mechanism of resistance to
MTX affects the ratio of FPGS/FPGH since polyglutamylated MTX has a
substantially longer half-life than monoglutamated MTX (20). Nevertheless, all four cell lines
showed similar expression levels of both FPGS and
FPGH (Fig. 3); thus, the
differences in MTX effects on cell proliferation were not caused by
changes in MTX polyglutamylation.
The flow cytometric analysis of the MTX-sensitive
cell lines, i.e., Daoy and Saos-2 cells, clearly confirmed the two
main effects of MTX on tumor cells that are responsible for its
ability to restrict cell proliferation. First, the cell cycle was
arrested in S-phase due to the depletion of nucleotide precursors;
our results showed apparent MTX-induced cell cycle arrest in
S-phase (Fig. 4). Similar findings
were previously described for cell lines derived from
adrenocortical carcinoma (21),
glioblastoma (22) and lung
carcinoma (23). Notably, we did
not observe any significant differences in the effects of the MTX
concentrations ranging from 0.1 to 40 μM on the distribution
of cell cycle phases (Fig. 4).
Secondly, the induction of cell death detected as the
sub-G1 fraction following treatment with MTX was also
involved in proliferation failure (Fig.
4). Furthermore, we noted a marked difference between treatment
with 0.1 μM MTX and the treatments with other concentrations
(Fig. 4), and these results were in
accordance with those achieved by the detection of activated
caspase-3-positive cells (Fig. 5).
Caspase-3-dependent/p53-independent apoptosis induced by MTX was
previously described in MCF-7 breast carcinoma cells (24). The apoptosis induced in the Saos-2
and Daoy cells was also p53-independent since Saos-2 cells do not
express p53 (25) and the C242F
mutation in the TP53 gene, which disables the
transactivation function of the p53 protein, was proven in Daoy
cells (26,27),
To summarize, our results showed that treatment with
MTX significantly decreased proliferation activity, inhibited the
cell cycle at S-phase and induced apoptosis in the Daoy and Saos-2
reference cell lines. Such effects apparently belong to the
DHFR-mediated mechanism of MTX action and are based on the
depletion of purine and pyrimidine precursors necessary for the
biosynthesis of nucleic acids. Importantly, we noted no difference
in these effects after treatment with various doses of MTX ranging
from 1 to 40 μM. These findings suggest the possibility of
achieving the same outcome with the application of low-dose MTX,
which is an extremely important result, particularly for clinical
practice, and may explain the lack of clinical advantage of HD-MTX
in children with advanced lymphoblastic lymphomas vs. low doses
(28). Moreover, the combined
application of MTX together with LV did not produce any detectable
effect, with exception of a partial reduction in MTX toxicity after
the use of the lowest concentration of MTX, i.e., 0.1
μM.
Acknowledgments
The present study was supported by the grant IGA
MZCR NT14327-3.
Abbreviations:
|
DHFR
|
dihydrofolate reductase
|
|
FPGH
|
folylpolyglutamate hydrolase
|
|
FPGS
|
folylpolyglutamate synthetase
|
|
LV
|
leucovorin
|
|
MTX
|
methotrexate
|
|
RCF
|
reduced folate carrier
|
|
THF
|
tetrahydrofolate
|
|
TYMS
|
thymidylate synthase
|
References
|
1
|
Takimoto CH: New Antifolates: Pharmacology
and Clinical Applications. Oncologist. 1:68–81. 1996.PubMed/NCBI
|
|
2
|
Neradil J, Pavlasova G and Veselska R: New
mechanisms for an old drug; DHFR- and non-DHFR-mediated effects of
methotrexate in cancer cells. Klin Onkol. 25:2S87–2S92.
2012.PubMed/NCBI
|
|
3
|
Sterba J, Valík D, Bajciová V, Kadlecová
V, Gregorová V and Mendelová D: High-dose methotrexate and/or
leucovorin rescue for the treatment of children with lymphoblastic
malignancies: do we really know why, when and how? Neoplasma.
52:456–463. 2005.PubMed/NCBI
|
|
4
|
Sterba J, Dusek L, Demlova R and Valik D:
Pretreatment plasma folate modulates the pharmacodynamic effect of
high-dose methotrexate in children with acute lymphoblastic
leukemia and non-Hodgkin lymphoma: ‘folate overrescue’ concept
revisited. Clin Chem. 52:692–700. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang JJ and Li GJ: Relationship between
RFC gene expression and intracellular drug concentration in
methotrexate-resistant osteosarcoma cells. Genet Mol Res.
13:5313–5321. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fotoohi AK and Albertioni F: Mechanisms of
antifolate resistance and methotrexate efficacy in leukemia cells.
Leuk Lymphoma. 49:410–426. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Assaraf YG: Molecular basis of antifolate
resistance. Cancer Metastasis Rev. 26:153–181. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Veselska R, Neradil J, Nekulova M,
Dobrucka L, Vojtesek B, Sterba J and Zitterbart K: Intracellular
distribution of the ΔNp73 protein isoform in medulloblastoma cells:
a study with newly generated rabbit polyclonal antibodies. Histol
Histopathol. 28:913–924. 2013.PubMed/NCBI
|
|
9
|
Schneider CA, Rasband WS and Eliceiri KW:
NIH Image to ImageJ: 25 years of image analysis. Nat Methods.
9:671–675. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Holmboe L, Andersen AM, Mørkrid L, Slørdal
L and Hall KS: High dose methotrexate chemotherapy:
pharmacokinetics, folate and toxicity in osteosarcoma patients. Br
J Clin Pharmacol. 73:106–114. 2012. View Article : Google Scholar :
|
|
11
|
Rahiem Ahmed YAA and Hasan Y: Prevention
and management of high dose methotrexate toxicity. J Cancer Sci
Ther. 5:106–112. 2013.
|
|
12
|
Cohen IJ1 and Wolff JE: How long can
folinic acid rescue be delayed after high-dose methotrexate without
toxicity? Pediatr Blood Cancer. 61:7–10. 2014. View Article : Google Scholar
|
|
13
|
Najim N, Podmore ID, McGown A and Estlin
EJ: Methionine restriction reduces the chemosensitivity of central
nervous system tumour cell lines. Anticancer Res. 29:3103–3108.
2009.PubMed/NCBI
|
|
14
|
Assaraf YG, Ifergan I, Kadry WN and Pinter
RY: Computer modelling of antifolate inhibition of folate
metabolism using hybrid functional petri nets. J Theor Biol.
240:637–647. 2006. View Article : Google Scholar
|
|
15
|
Bastian L, Einsiedel HG, Henze G, Seeger K
and Shalapour S: The sequence of application of methotrexate and
histone deacetylase inhibitors determines either a synergistic or
an antagonistic response in childhood acute lymphoblastic leukemia
cells. Leukemia. 25:359–361. 2011. View Article : Google Scholar
|
|
16
|
Huang Y: Pharmacogenetics/genomics of
membrane transporters in cancer chemotherapy. Cancer Metastasis
Rev. 26:183–201. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yoon SA, Choi JR, Kim JO, Shin JY, Zhang X
and Kang JH: Influence of reduced folate carrier and dihydrofolate
reductase genes on methotrexate-induced cytotoxicity. Cancer Res
Treat. 42:163–171. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nazki FH, Sameer AS and Ganaie BA: Folate:
metabolism, genes, polymorphisms and the associated diseases. Gene.
533:11–20. 2014. View Article : Google Scholar
|
|
19
|
Furuta E, Okuda H, Kobayashi A and Watabe
K: Metabolic genes in cancer: their roles in tumor progression and
clinical implications. Biochim Biophys Acta. 1805:141–152.
2010.PubMed/NCBI
|
|
20
|
Rots MG, Willey JC, Jansen G, van Zantwijk
CH, Noordhuis P, DeMuth JP, Kuiper E, Veerman AJ, Pieters R and
Peters GJ: mRNA expression levels of methotrexate
resistance-related proteins in childhood leukemia as determined by
a standardized competitive template-based RT-PCR method. Leukemia.
14:2166–2175. 2000. View Article : Google Scholar
|
|
21
|
Nilubol N, Zhang L, Shen M, Zhang YQ, He
M, Austin CP and Kebebew E: Four clinically utilized drugs were
identified and validated for treatment of adrenocortical cancer
using quantitative high-throughput screening. J Transl Med.
10:1982012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Capelôa T, Caramelo F, Fontes-Ribeiro C,
Gomes C and Silva AP: Role of methamphetamine on glioblastoma
cytotoxicity induced by Doxorubicin and methotrexate. Neurotox Res.
26:216–227. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yan KH, Lee LM, Hsieh MC, Yan MD, Yao CJ,
Chang PY, Chen TL, Chang HY, Cheng AL, Lai GM and Chuang SE:
Aspirin antagonizes the cytotoxic effect of methotrexate in lung
cancer cells. Oncol Rep. 30:1497–1505. 2013.PubMed/NCBI
|
|
24
|
Hattangadi DK, DeMasters GA, Walker TD,
Jones KR, Di X, Newsham IF and Gewirtz DA: Influence of p53 and
caspase 3 activity on cell death and senescence in response to
methotrexate in the breast tumor cell. Biochem Pharmacol.
68:1699–1708. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hellwinkel OJ, Müller J, Pollmann A and
Kabisch H: Osteosarcoma cell lines display variable individual
reactions on wildtype p53 and Rb tumour-suppressor transgenes. J
Gene Med. 7:407–419. 2005. View Article : Google Scholar
|
|
26
|
Jordan JJ, Inga A, Conway K, Edmiston S,
Carey LA, Wu L and Resnick MA: Altered-function p53 missense
mutations identified in breast cancers can have subtle effects on
transactivation. Mol Cancer Res. 8:701–716. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Künkele A, De Preter K, Heukamp L, Thor T,
Pajtler KW, Hartmann W, Mittelbronn M, Grotzer MA, Deubzer HE,
Speleman F, Schramm A, Eggert A and Schulte JH: Pharmacological
activation of the p53 pathway by nutlin-3 exerts anti-tumoral
effects in medulloblastomas. Neuro Oncol. 14:859–869. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Termuhlen AM, Smith LM, Perkins SL, Lones
M, Finlay JL, Weinstein H, Gross TG and Abromowitch M: Disseminated
lymphoblastic lymphoma in children and adolescents: results of the
COG A5971 trial: a report from the Children’s Oncology Group. Br J
Haematol. 162:792–801. 2013. View Article : Google Scholar : PubMed/NCBI
|