Introduction
Cutaneous melanoma remains the most aggressive type
of cancer due to its invasiveness and propensity to metastasize
(1). Malignant melanoma
preferentially metastasizes to the lymph nodes, lungs and liver
(2). While surgery and radiation
therapy play a role in the palliation of the symptoms from local
tumor growth, systemic therapy is the primary mode of treatment for
metastatic melanoma, involving either single-agents or a
combination of chemotherapy, immunotherapy or biochemical
therapies. Combinations of these therapies may improve the response
rate but do not extend survival and are associated with greater
hematologic and multi-organ toxicity (3,4). As
the incidence of melanoma continues to rise, significant unmet
medical needs remain for novel, effective and safe therapies for
the treatment of malignant melanoma.
The increased metastatic potential of melanoma has
been associated with altered expression patterns of cell adhesion
receptors including integrins (5,6).
During melanoma progression from the benign melanocytic nervus to
the metastatic melanoma, the melanoma lesion undergoes
histopathologically distinct stages from the radial growth phase
lesions to the vertical spreading into the adjacent papillary
dermis-forming ‘vertical growth phase’ lesions (7,8). The
tumor cells then enter the vasculature of the lymphatic system to
invade and colonize distant target organs. Findings of previous
studies suggest that integrin expression controls melanoma
tumorigenicity by modulating cell migration (5,9),
facilitating cell invasion and angiogenesis (10), while promoting tumor cell survival
(11–13). Among the integrins associated with
melanoma progression, the αvβ3 integrin, although not normally
expressed on epidermal melanocytes or in most of the benign
melanocytic nevi, has been found to be greatly increased as
epidermal melanoma progresses to the vertical growth and invasive
phases and is expressed in metastatic melanoma lesions (7).
Integrin αvβ3, a heterodimeric cell-surface adhesion
receptor, specifically recognizes the arginine-glycine-aspartic
(RGD) tripeptide sequence in a variety of extracellular matrix
proteins, including vitronectin, osteopontin, fibrinogen,
fibronectin, thrombospondin, von Willebrand factor and cryptic
collagens (14,15). Notably, the integrin αvβ3 has been
demonstrated to mediate osteoclastic bone resorption and
endothelial neovascularization or angiogenesis. Significant
upregulation of αvβ3 integrin expression is seen in tumoral
endothelial cells, as well as in some tumor cells (9). Overexpression of αvβ3 by gene transfer
in melanoma cell lines derived from the radial growth phase changed
the properties of the cells to those of the vertical growth phase
(16). Blocking antibodies to β3
integrin were reported to inhibit the migration, proliferation and
metastasis of melanoma, as well as other tumor cells (17). Ligand engagement of αvβ3 integrin
has been shown to induce melanoma cell growth by inhibiting
apoptosis (11). Additionally,
blocking of this integrin in human melanoma HT168M1 cells using an
anti-β3 integrin monoclonal antibody resulted in the inhibition of
lung colonization in an experimental metastasis assay (17). In that study, the anti-β3 integrin
antibody recognized integrin αIIbβ3 and αvβ3 expressed in the human
melanoma cell line, suggesting that the two integrins may
participate in promoting tumor metastasis (17). Several RGD-disintegrins have also
been shown to inhibit melanoma cell proliferation (18,19).
MK-0429 (L-000845704) is an orally active, potent
and selective inhibitor of the integrin αvβ3 (20). As this inhibitor was originally
developed for the treatment of osteoporosis, robust preclinical
evidence demonstrated that MK-0429 potently inhibits binding of the
ligand to the purified human integrin αvβ3 (IC50=0.08
nM) (20), and inhibits
osteoclastic bone resorption (IC50=12.2±4.5 nM) in
vitro. MK-0429 significantly reduces bone turnover and
increases bone mass in ovariectomized rats and monkeys (20). In a randomized phase II trial,
postmenopausal women with osteoporosis receiving MK-0429 at either
100 or 400 mg once daily, or 200 mg twice daily for 12 months
showed a significant increase in bone mineral density at the spine
and hip compared to those on placebo (21). In a small clinical study, men with
hormone refractory prostate cancer and metastatic bone disease were
administered MK-0429 at 200 and 1,600 mg orally, twice daily for
four weeks (22). MK-0429
significantly decreased the bone resorption marker urinary
N-telopeptide in these patients. In both clinical studies, MK-0429
was well tolerated and without serious adverse events.
In the present study, to gain better insight into
the function of integrin αvβ3 in melanoma progression and
metastasis, we first evaluated the efficacy of MK-0429 compared to
cyclophosphamide in the prevention of metastatic melanoma
progression using a model of murine B16F10 melanoma metastasis to
the lungs. MK-0429 was then evaluated in a second experiment that
employed B16F10 luciferase-expressing cells to examine the de
novo progression of metastasis, wherein the
treatment-associated effects on tumor progression in target tissues
were evaluated with bioluminescent imaging in vivo and ex
vivo.
Materials and methods
Synthesis of MK-0429
This compound is also known as L-000845704
(3(S)-(6-methoxypyridin-3-yl)-3-[2-oxo-3-[3-(5,6,7,8-tetrahydro-[1,8]-naphthyridin-2-yl)
propyl] imidazolidin-1-yl] propionic acid). The synthesis and
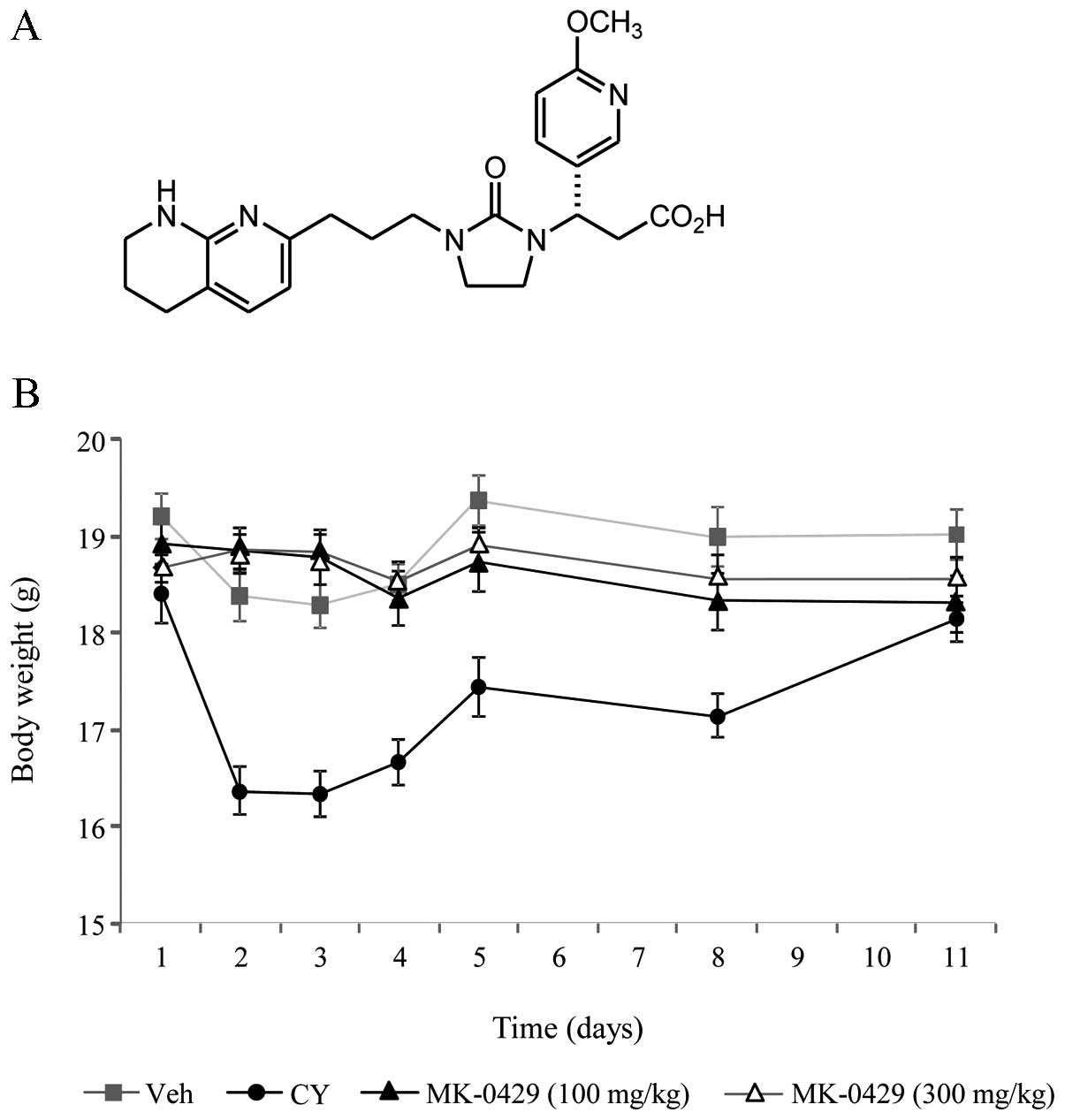
structure of MK-0429 (Fig. 1a) were
as previously described (20).
Integrin-mediated binding assays
Human embryonic kidney 293 (HeK293) cells were
stably co-transfected with human integrin subunits αv and β3 to
establish the HeK293-αvβ3 cell line as previously described
(23). Purification of these
integrins and assay conditions were performed as previously
described (20). Briefly, the
affinity of 3H-MK-0429 for various integrins was
determined by binding assays using the purified receptors.
Cell adhesion assays
HeK293 cells overexpressing human integrins αvβ3,
αvβ5, αIIbβ3 or α5β1 were established as previously described
(23,24). The cells (25×103
cells/well) were added to microtiter wells that were coated with
vitronectin (αvβ3 and αvβ5), fibrinogen (αIIbβ3) or fibronectin
(α5β1) and were allowed to attach for 2 h at 37°c in a humidified
incubator in the absence or presence of increasing concentrations
of MK-0429. The non-attached cells were gently washed away. The
attached cells were quantified by colorimetric detection of
hexosaminidase enzymatic activity (25) in a Vmax micro-plate reader
(Molecular Devices, Menlo Park, CA, USA). The number of attached
cells was quantified using a standard curve for each cell line
assayed and expressed as a mean value of triplicate samples.
B16F10 melanoma cell lines
The B16F10 murine melanoma cell line, was obtained
from the American Type Culture Collection (ATCC; Manassas, VA,
USA). The cells were routinely cultured in high glucose Dulbecco’s
modified eagle’s medium (DMEM) supplemented with 10% fetal bovine
serum (FBS) and 1% penicillin/streptomycin (Life Technologies,
Carlsbad, CA, USA) at 37°C in a humidified 5% CO2
incubator. Stable expression of luciferase was established in the
B16F10 melanoma cells through Geneticin (G418) selection. Briefly,
B16F10 cells were transfected with pcDNA3.1 and pGL3 plasmid using
Lipofectamine 2000 (Life Technologies, Carlsbad, CA, USA) according
to the manufacturer’s instructions. The cells were then selected in
1,250 μg/ml G418 (Life Technologies) for two weeks and
individual colonies were isolated, expanded and maintained in G418.
Luciferase expression in these clones was confirmed by flow
cytometry and bioluminescent imaging.
Reverse transcription and TaqMan
The total RNA was extracted using an RNeasy kit
(Qiagen, Valencia, CA, USA) according to the manufacturer’s
instructions. Reverse transcription and real-time PCR were
performed as previously described (26). Primer/probe pairs for real-time PCR
were as follows (Life Technologies): ITGAV forward,
5′-CGGGTCCCGAGGGAAGT-3′ and reverse, 5′-GGGTCGTGTTCGCTTTGG-3′), and
fluorogenic probe, 5′-TCGAGCCCAGCACGTCCTCCA-3′; ITGB3 forward,
5′-GATGCTTACGGGAAAATCCG-3′ and reverse,
5′-TTGAAGGACAGTGACAGCTCTCC-3′, and fluorogenic probe,
5′-CTAAAGTGGAGCTGGAAGTACGTGACCTGC-3′); and ITGB5 forward,
5′-GGTTTCGGGTCTTTTGTTGACA-3′ and reverse,
5′-GGAATAACTTGTAACCAATACACGGA-3′, and fluorogenic probe,
5′-TCTCTCCTTTCTCCTACACGGCACCGA-3′.
Development of lung metastasis in the
murine B16F10 melanoma model
The in-life portion of the present study was
conducted at the Piedmont Research Center (Morrisville, NC, USA).
The Piedmont Research Center complies with the recommendations of
the Guide for Care and Use of Laboratory Animals and is accredited
by AAALAC International, which assures compliance with accepted
standards for the care and use of laboratory animals. In total, 55
6-week-old B6D2F1 hybrid female mice received 1.5×105
B16F10 melanoma cells by intravenous (i.v.) tail vein injection.
The animals received either saline, MK-0429 at 100 or 300 mg/kg,
p.o., twice daily (b.i.d.) or cyclophosphamide, 300 mg/kg, i.p.,
once daily (q.d.), 1 day post-tumor injection, n=10–15/group. The
body weight was recorded daily to determine whether treatment
affected the health of the animals. Necropsy was performed when the
number of lung colonies reached 100 metastases/lung counted from a
separate set of control mice, ~2 weeks after study initiation
(27,28). Lungs were dissected with minimal
bronchus attached. Melanoma colonies on the surface of all lung
regions were counted.
Histological analysis of the mouse lungs was
performed. The mouse lungs were fixed in 10% formalin. After being
embedded, the samples were sectioned beginning at ~1 mm into the
tissue along the frontal plane. The sections were stained with
hematoxylin and eosin (H&E) followed by imaging and tumor area
quantification using ImagePro software.
Non-invasive bioluminescent imaging of de
novo lung metastasis in the murine B16F10 melanoma model
The in-life portion of the present study was
conducted at Merck Research Laboratories (Rahway, NJ, USA). Animal
procedures were in accordance with the national guidelines and were
approved by the Institutional Animal Care and Use Committee (IACUC)
at Merck.
Twenty 8-week-old (nu/nu) female mice, were injected
with 2.5×105 B16F10-luc melanoma cells by i.v. tail vein
injection. The animals received saline or MK-0429, 300 mg/kg, p.o.,
b.i.d., 1 day post-tumor injection, n=10/group. Bioluminescence
imaging was performed using the Xenogen IVIS 200 (Perkin-Elmer,
Waltham, MA, USA) twice/week until the study end and images were
quantified using Living image software. The mice were anesthetized
with 2.5% isoflurane prior to imaging and then a 90 mg/kg dose of
D-luciferin, i.p. (Perkin-Elmer). Exposure time was adjusted to
avoid pixel saturation. Default settings for bioluminescent
scanning were used for scanning at each time-point. A total
bioluminescent flux (photons/second) was calculated for each region
of interest (ROI). Square ROIs were placed around the head, lung
and abdomen of each animal for each scan followed by image analysis
of head, lungs and abdomen, on dorsal and ventral planes.
The animals were sacrificed by carbon dioxide
asphyxiation. Following necropsy, ex vivo bioluminescent
imaging of the lungs was performed by Xenogen IVIS 200. Default
bioluminescent settings of Living Image were used with exposure
times manually adjusted to avoid saturation. ROIs were placed on
the 2D bioluminescent image to encompass the entire lung tissues.
Melanoma colonies on the surface of the lung regions were
counted.
Statistical analysis
Data are presented as mean ± SEM and were analyzed
with GraphPad Prism 6 software (San Diego, CA, USA). Study
endpoints were tested for Gaussian distribution. Statistical
analysis was performed by the unpaired Student’s t-test or the
one-way ANOVA followed by the Tukey’s multiple comparison test. The
histological quantification of the tumor area was analyzed using
StatView, followed by the Fisher’s PLSD test. P<0.05 was
considered to indicate a statistically significant result.
Results
Potency and safety profile of MK-0429 and
integrin expression profile of B16F10 melanoma
The structure of MK-0429 (Fig. 1a) was previously described (20). MK-0429 binds with high affinity to
the purified human αvβ3 integrin. The equilibrium dissociation
constants (Kds) of 3H-MK-0429 in binding to the purified
human, murine and rat αvβ3 integrin are 0.33±0.04, 0.56±0.07 and
1.23±0.11 nM, respectively. This inhibitor blocks the adhesion of
HeK293-αvβ3 cells to vitronectin with an IC50 of
0.58±0.30 nM. MK-0429 is ~100-fold less potent in blocking the
adhesion of HeK293 overexpressing the closely related αvβ5 integrin
to vitro-nectin, and >1,000-fold less active in blocking
adhesion functions mediated by integrins αIIbβ3 or α5β1 to
fibrinogen or fibronectin, respectively.
The mRNA expression levels of integrin subunits were
determined for the highly metastatic B16F10 cell line. Integrin αv
was the predominant subunit, demonstrating a mRNA expression
~8-fold greater than that of the β5 subunit. The β3 subunit was
detectable at the cycle threshold values near 40 (data not shown),
consistent with previous reports from the FACS analysis (29). Having established detectable
expression of the subunits of the vitronectin receptors in the
melanoma cell line, we then investigated MK-0429 as a potential
therapeutic for the treatment of melanoma.
Effects of MK-0429 on body weight of mice
injected with melanoma cells
MK-0429 has been demonstrated to be well tolerated
and efficacious in preclinical and clinical studies of osteoporosis
(21,22). In the present study, we evaluated
its effect on body weight compared to cyclophosphamide in mice
employing a B16F10 murine melanoma model in the prevention mode.
Animals received tail-vein injection of B16F10 melanoma cells
followed by treatment with vehicle (Veh), MK-0429 (at 100 and 300
mg/kg, p.o., b.i.d.) or cyclophosphamide (CY; 300 mg/kg, i.p.,
q.d.) one day after cell inoculation. To validate the utility of
the model, metastatic lung nodule development was monitored in a
separate cohort, with ~100 metastatic lung colonies developing
within two weeks of B16F10 cell inoculation and this time period
was defined as the operative study duration (data not shown). Veh-
and MK-0429-treated animals showed no significant weight loss over
the study duration (Fig. 1B). By
contrast, the CY-treated animals experienced a rapid loss of weight
in the first four days of the study, losing ~9–11% of their total
body weight. This was followed by a return towards the baseline
weight levels by the end of the study (Fig. 1B).
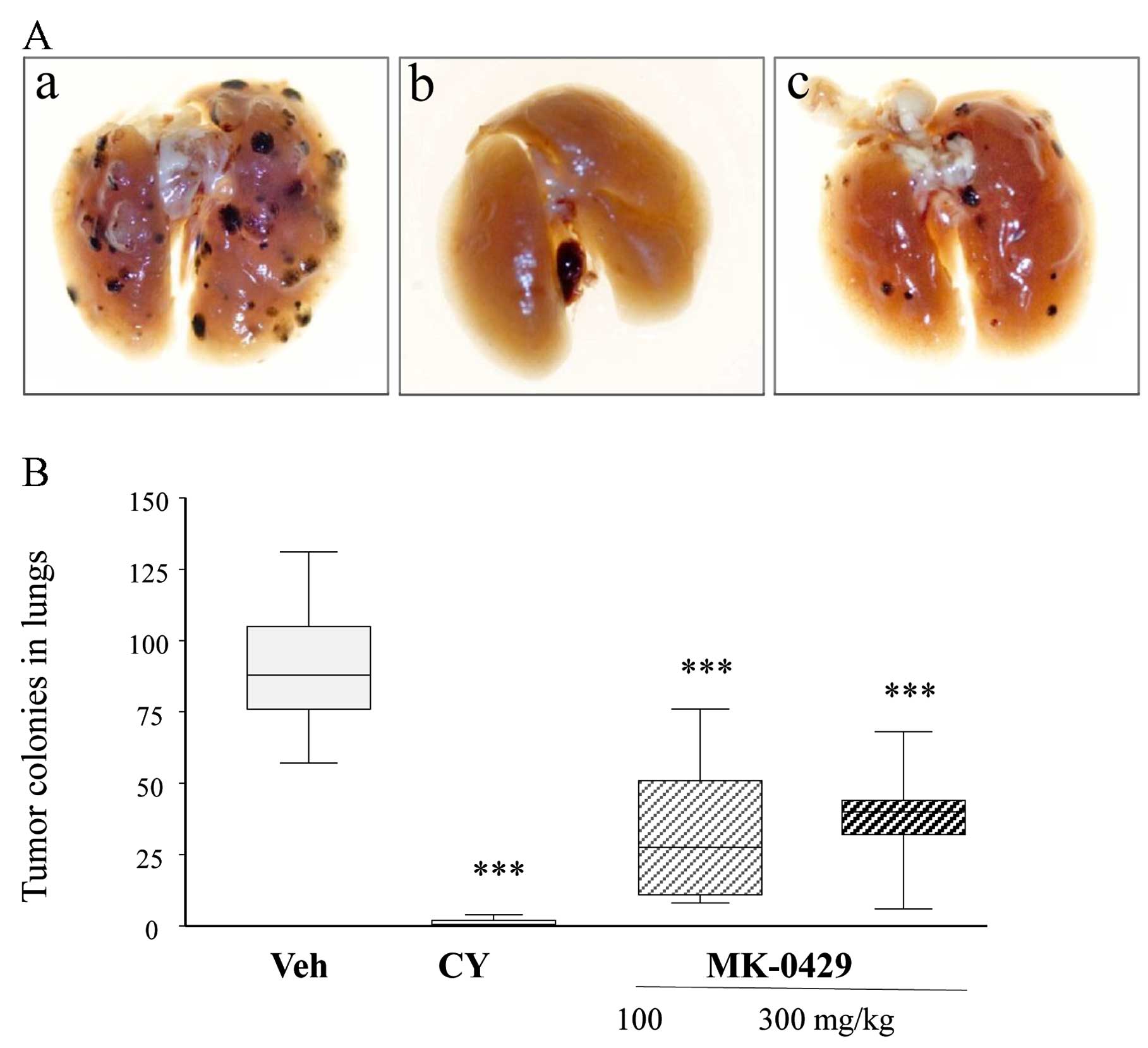
MK-0429 reduces metastatic tumor colony
formation and area in the lungs
The extent of lung metastasis and the effect of drug
treatment were assessed by examining the lung surfaces for
observable melanoma tumor colony formation. Following termination
of the study, lung surfaces were visually inspected and the total
number of ex vivo melanoma colonies on the lung surface was
counted. The gross appearance of representative ex vivo lung
sets revealed clearly visible metastatic melanoma colonies (dark
spots) in Veh-treated animals and notably fewer observable colonies
following CY or MK-0429 treatment (Fig.
2A). CY and MK-0429 effectively reduced the number of tumor
colonies on the entire lung surface (Fig. 2B). The mean number of the metastatic
melanoma nodules was 90±5 in the Veh-treated controls.
Cyclophosphamide treatment eradicated almost all the tumor
colonies, 99% as compared to that in the Veh control (p<0.001).
MK-0429 treatment also decreased the total tumor colony number to
33±6 and 39±9, at 100 and 300 mg/kg, respectively, (p<0.001 vs.
Veh) thus reducing tumor burden by 64 and 57%.
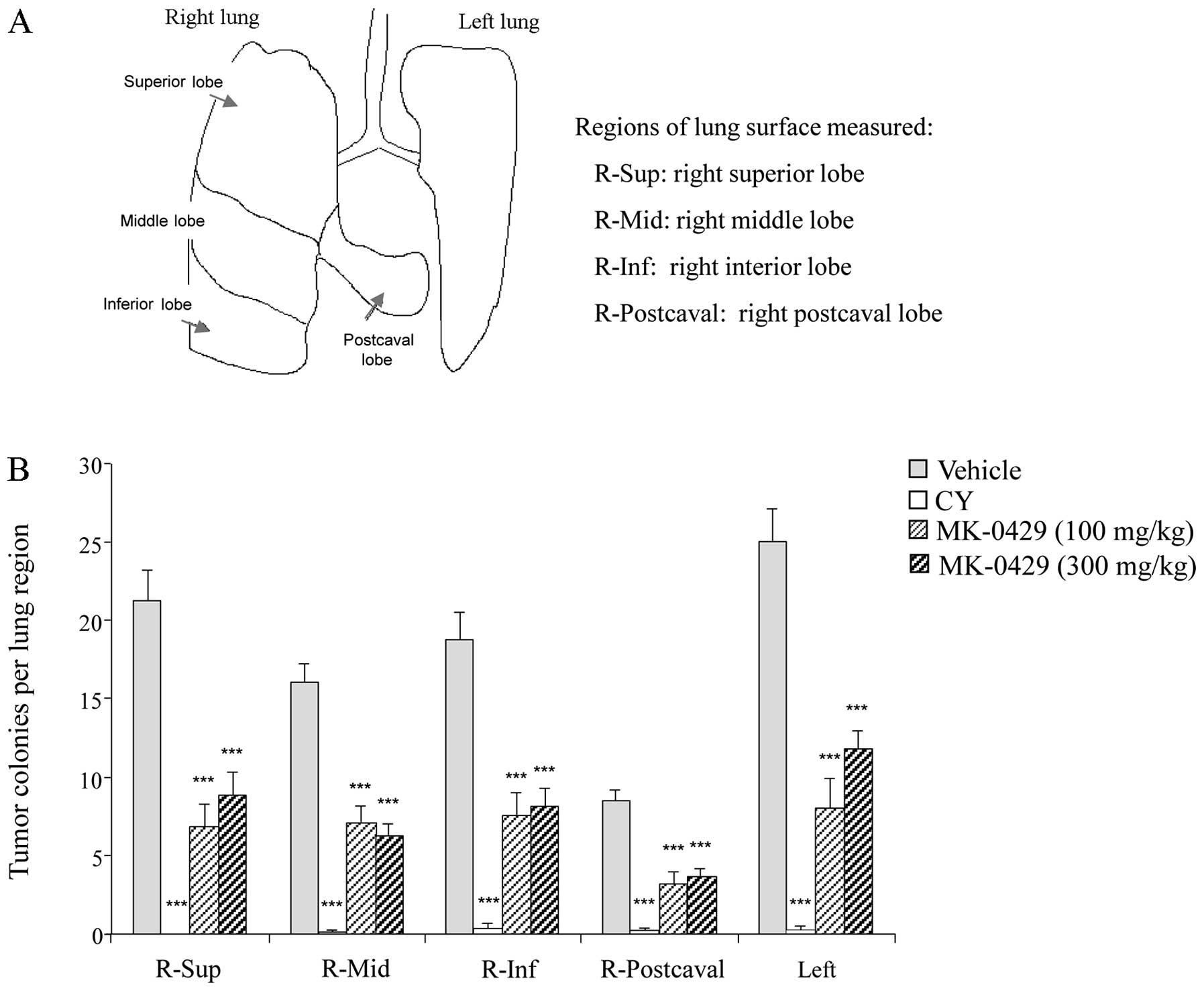
The lung was subdivided into five separate
compartments to determine whether particular surface areas were
preferentially affected by metastasis or drug treatment (Fig. 3A). Tumor colonies were manually
counted within each lung region. In the Veh-treated animals, the
scope of tumor colonies across the lung surfaces averaged 9–25
tumor colonies/individual lung surface (Fig. 3B). The largest lung surface, the
left lung, contained the highest number of melanoma colonies
(25±8), whereas the smallest lung surface, the post caval lobe,
showed the least tumor colony formation (9±1). Metastatic melanoma
burden was significantly reduced within each lung compartment with
drug treatment. Consistent with the results from the entire lung
surface, CY treatment markedly reduced tumor formation in each of
the five lung regions (p<0.001 vs. Veh). MK-0429 at the two
doses significantly reduced melanoma colonies in all the lung
regions by 53–68% indicating no regional effect of treatment
(p<0.001 vs. Veh) (Fig. 3B).
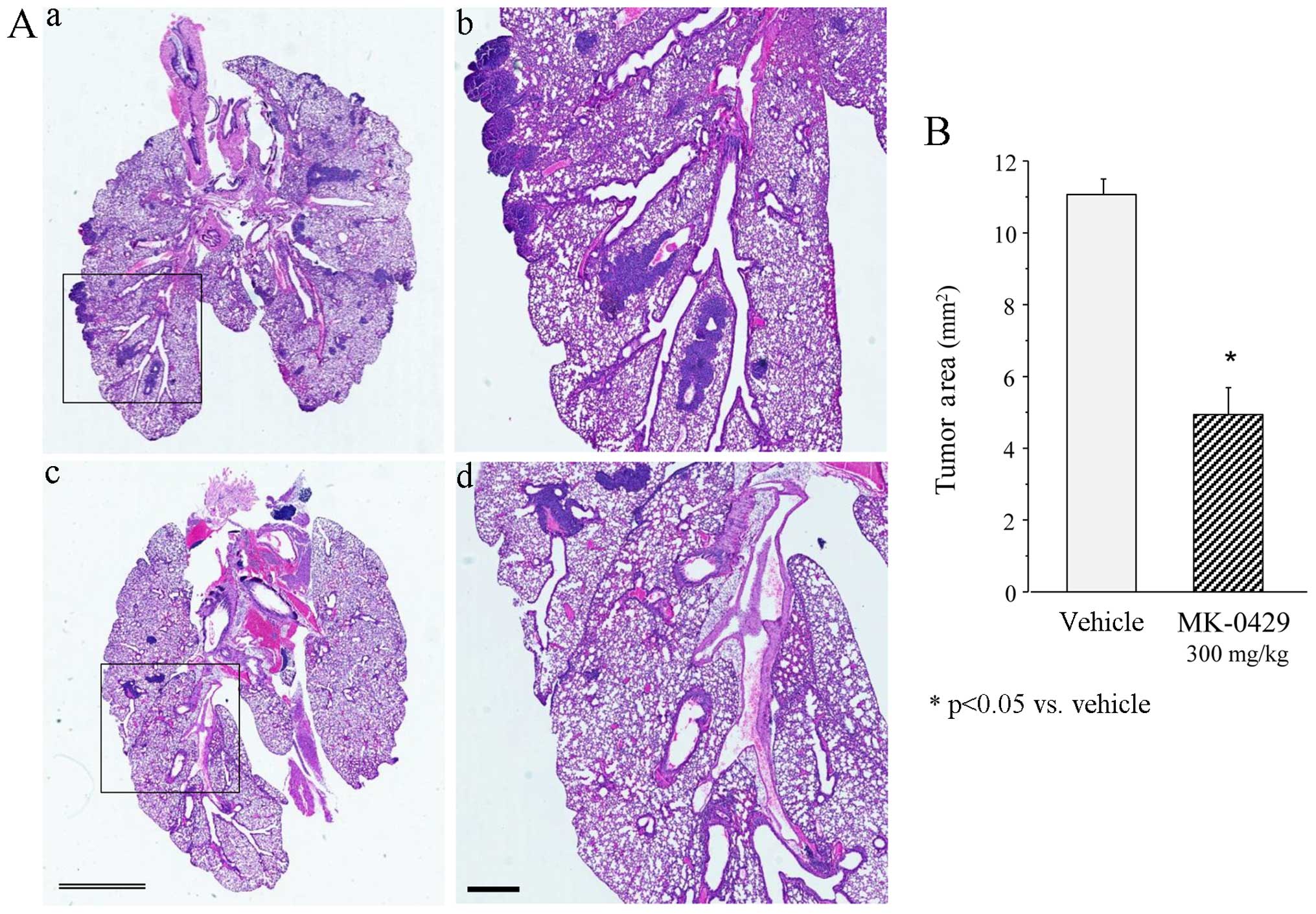
The presence of metastatic lesions in the lungs was
confirmed by histological analysis of the sections from the Veh-
and MK-0429 (300 mg/kg)-treated animals. Fig. 4Aa and b shows a typical
H&E-stained section of Veh-treated lungs containing melanoma
colonies. However, fewer colonies were present in the
MK-0429-treated lungs (Fig. 4Ac and
d). Veh-treated animals showed a total tumor area of 11.1±0.4
mm2. The total number and area of the melanoma colonies
was significantly reduced following treatment with MK-0429 to
5.0±0.7 mm2 (p<0.01 vs. Veh) (Fig. 4B). Treatment with MK-0429 decreased
the tumor area by 60% compared to that in the Veh controls,
consistent with the treatment-associated reduction of the number of
melanoma colonies.
MK-0429 reduces the de novo progression
of lung metastases
The second experiment focused on the de novo
progression patterns of metastatic spread and the resulting effect
of drug treatment on melanoma metastasis in mice by utilizing
non-invasive imaging and B16F10 melanoma cells stably transfected
with luciferase. In vitro bioluminescent imaging confirmed
the expression levels of luciferase for B16F10 clones selected for
in vivo studies. Additionally, real-time PCR demonstrated
integrin subunit mRNA levels consistent with the parental cell line
(data not shown). Athymic mice were injected with
2.5×105 B16F10-luc cells. One-day post injection, the
animals were orally administered either Veh or MK-0429, 300 mg/kg,
b.i.d. for two weeks. The animals were imaged for bioluminescence
on day 0, 3, 8, 11 and 15 to determine the time required to develop
measurable lung metastases and the extent of metastatic spread
within other organs. Tumor progression was monitored and quantified
by image analysis in the head, lungs and abdomen.
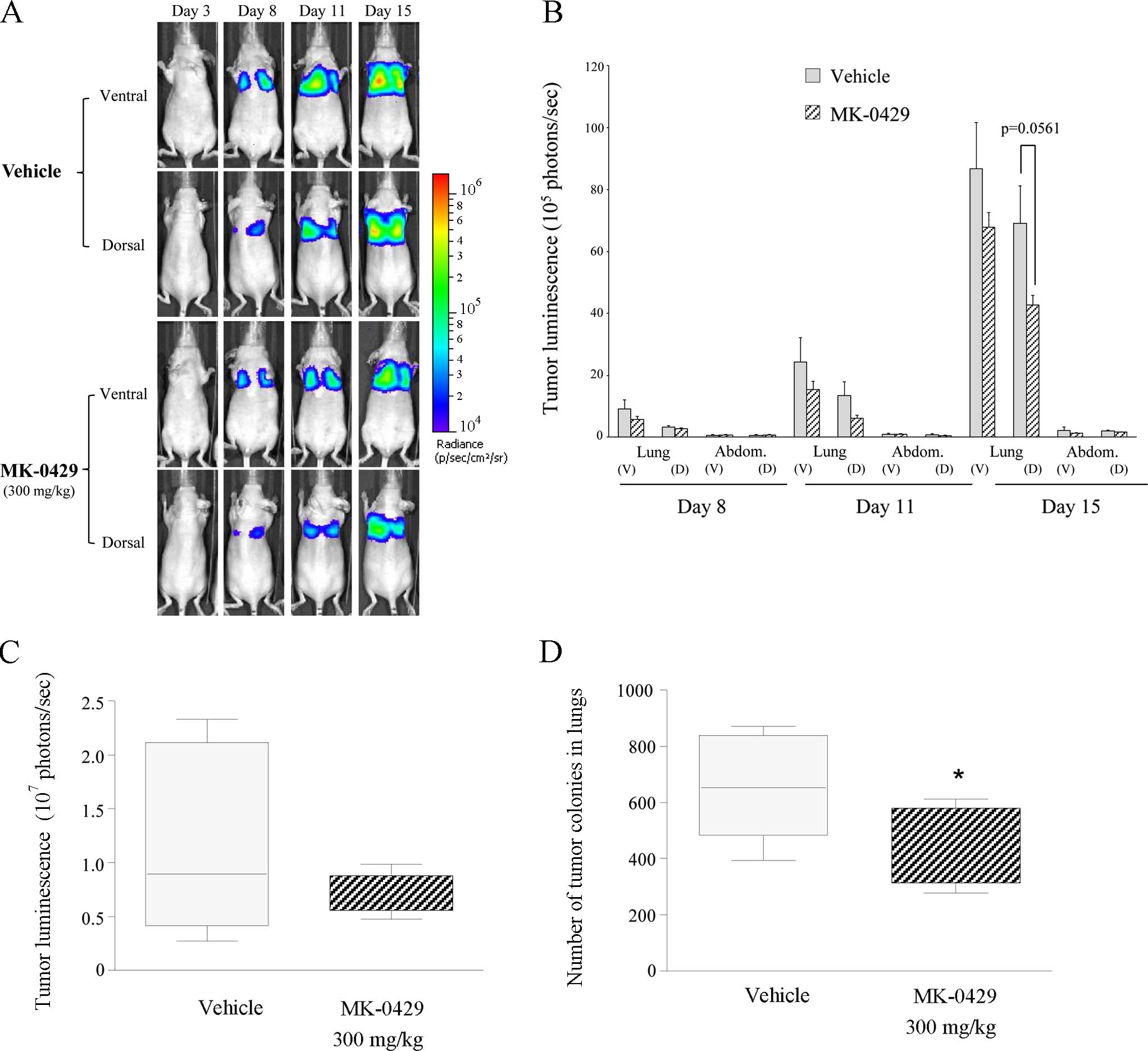
Fig. 5A shows a
progressive increase in tumor signal from day 3 through day 15 in
the study animals. Representative bioluminescent images show that
tumor progression was negligible in the abdomen and head throughout
the study time course. However, melanoma burden gradually increased
in the lungs in the dorsal and ventral planes in the Veh-treated
animals (Fig. 5A). Tumor burden
advanced less aggressively in MK-0429-treated animals.
Quantification of tumor burden demonstrated a time-dependent
increase in lung metastases in the Veh controls, whereas the
abdomen and head showed little detectable signal above the
background in the Veh- and drug-treated animals (Fig. 5B). MK-0429 300 mg/kg on day 8, 11
and 15, reduced melanoma burden in the ventral lung by 38, 37 and
21%, respectively. Similarly, MK-0429 reduced tumor metastases in
the dorsal lung by 15, 54 and 38% (p=0.0561 vs. Veh), respectively,
as compared to the Veh controls (Fig.
5B). Lung metastasis was confirmed by ex vivo
bioluminescent analysis. Veh-treated lungs demonstrated a tumor
burden of 118.1±27.1×105 photons/sec. MK-0429 decreased
metastasis in the lungs by ~40%, reducing melanoma burden to
69.9±5.9×105 photons/sec. This difference was not
statistically significant (Fig.
5c). However, the number of visible metastatic colonies counted
on the ex vivo lung surface was significantly reduced by 32%
by MK-0429 treatment twice daily for two weeks (p<0.05 vs. Veh),
consistent with the percentage reduction of tumor burden observed
by bioluminescent imaging (Fig.
5D). Thus, evaluation of melanoma burden at study termination
by in vivo and ex vivo bioluminescent imaging, as
well as by manual melanoma colony count, indicated an ~30–40%
reduction in lung metastasis following treatment with MK-0429 300
mg/kg administered twice daily as compared to the Veh-treated
controls.
Discussion
During melanoma progression from the benign
melanocytic nevus to the metastatic melanoma, the melanoma lesion
undergoes several clinically and histopathologically distinct
stages (7,8). Initially, the radial growth phase
lesions are limited to the epidermis and are essentially benign.
Then, the cells begin to spread vertically into the adjacent
papillary dermis, while they continue to invade the adjacent
reticular dermis, subcutaneous fat and eventually enter the
lymphatics and vascular circulation. Degradation of the basement
membranes and vascularized structures around malignant melanoma are
also crucial to local invasion and hematogenous metastases of
melanoma (8). Expression of αvβ3
integrin, previously found to be involved in the regulation of cell
growth, motility and invasion (30,31),
coincides with progression of the invasive phase of melanocytes to
the vertical growth phase of metastatic melanoma (32,33).
Furthermore, αvβ3 integrin is involved in angiogenesis as well as
in several processes of melanoma metastasis by promoting cell
proliferation, attachment, transendothelial migration and invasion
through an interaction with MMP-2 to support cell intravasation,
extravasation and target organ colonization (34,35).
Collectively, the above mentioned data raise the possibility that
blocking the functions of αvβ3 integrin prevents the early stages
of melanoma metastasis.
The focus of this preclinical study was to evaluate
the role of this adhesion receptor αvβ3 in melanoma in target organ
colonization. MK-0429, a known selective inhibitor of human αvβ3
integrin (20), also blocks
adhesion of the HEK293-αvβ5 cells, albeit at ~100-fold lower
efficacy compared to that of αvβ3 integrin (23). The selectivity profile of MK-0429
was further demonstrated by its lack of potency in inhibition in a
panel of adhesion assays, including the attachment of HEK293 cells
overexpressing the αIIbβ3 or α5β1 integrin to fibrinogen or
fibronectin, respectively (25).
Although the expression levels of αvβ5 integrin appear to be higher
than that of αvβ3 integrin in B16F10 cells, the selectivity profile
of MK-0429 supported the predominant role of αvβ3 integrin in the
colonization and growth of this murine melanoma in the lungs.
While high expectations for the mechanism of αvβ3
integrin in mediating tumor angiogenesis, growth and invasion have
derived from the significant body of positive preclinical and early
clinical findings (36), blocking
antibody or small molecular weight inhibitors led to mostly
negative results from various phase II and III trials in
pancreatic, prostate, head and neck cancers, glioblastoma and
melanoma (9,37–40).
Etaracizumab is a monoclonal antibody against αvβ3 that was
evaluated at 8 mg/kg once weekly, i.v., and administered for two
cycles of a 3-week infusion in 112 patients with stage IV
metastatic melanoma in the presence and absence of dacarbazine for
a year (41). The median survival
rate was not different in patients treated with etaracizumab alone
or in combination with dacarbazine, suggesting etaracizumab
treatment was unlikely to result in clinically meaningful
improvement over dacarbazine alone (41). Cilengitide (EMD 121974), a dual
inhibitor of αvβ3 and αvβ5 integrins, was administered
intravenously twice weekly in a small number of patients (n=12–14)
with stage IV or unresectable stage III metastatic melanoma to
assess the clinical efficacy of cilengitide in the progression-free
survival rate at eight weeks (38).
Cilengitide demonstrated minimal clinical efficacy as a
single-agent therapy for advanced melanoma (38). Despite the implications of the role
of αvβ3 integrin involvement in the early stages of melanoma
metastasis, the aim of these previous studies was clearly geared
towards testing efficacy of αvβ3 inhibitors in patients in later
stages of advanced melanoma.
Other pitfalls of the early trials may explain the
lack of efficacy of anti-αvβ3 integrin in the melanoma population.
Targeting of αvβ3 in tumor tissues is complex due to the
dose-dependent opposing effects of the inhibitor: low doses of
RGD-peptides have been reported to stimulate VEGF-blood vessel
growth and tumor angiogenesis (42), in contrast with inhibition at higher
doses (36). Alternative adhesion
receptors, including αvβ5 or α5β1 integrins, may also compensate
for tumor angiogenesis (36).
Metabolic imaging and tissue analysis suggested that etaracizumab
or cilengitide reached the target tissues. However, little is known
regarding the effect of these drugs on tumor vasculature or
invasiveness in patients. Moreover, etaracizumab and cilengitide
have relatively short half-lives of approximately a few days and
2–4 h, respectively. Thus, unfavorable pharmacokinetics and
non-continuous dosing regimens of these drugs may partly explain
the negative results. Intravenous administration of these drugs,
together with the infrequency of dosing intervals, may be
suboptimal to achieve appropriate anti-angiogenic or anti-invasion
pressure, which may require persistent long-term therapy for
effectiveness.
An issue that is to be considered is whether the
mechanism of blocking αvβ3 integrin should be re-evaluated with an
orally active agent with a proven excellent safety profile in the
clinic, particularly in the context of early intervention of the
disease course, such as preventing systemic spread. MK-0429 may be
suitable for this approach. Although safety and efficacy of MK-0429
were clearly demonstrated in animal models (20) and in postmenopausal osteoporotic
women for two years (21), the
safety profile of this compound has also been recently tested at
the maximally tolerated oral dose of 1,600 mg twice daily for four
weeks in men with hormone refractory prostate cancer and metastatic
bone disease (22). Markedly,
MK-0429 was well tolerated at this high dose.
At present, there are three classes of therapies
approved for melanoma, the alkylating agents such as dacarbazine,
the targeted therapies such as the selective inhibitor of BRAF
V600E, and immunotherapeutic agents including anti-CTLA-4 antibody
and more recently anti-PD-1 antibody (43). Dacarbazine remains the gold standard
in chemotherapy, while combining dacarbazine with new
pharmacological or immunotherapeutic agents is currently under
evaluation in order to achieve better clinical responses in
patients with advanced melanoma. The development of targeted- and
immuno-therapies has changed the paradigm for treatment of patients
with advanced melanoma, although not all patients respond to these
therapies. The lack of predictive biomarkers of response is
probably the major weakness of these novel therapies. The recent
development of MK-0429-based imaging markers may be useful to
monitor melanoma responses to therapies (25). Furthermore, for future therapy of
metastatic melanoma, the challenge is to expand and combine the
current therapies to prevent systemic metastasis at the earlier
stage of the disease. In the present study, we have provided new
insight into the application of a novel small molecule integrin
inhibitor with an excellent safety profile as a therapeutic agent
for the early prevention of metastatic melanoma. Combination of the
αvβ3 integrin inhibitor with the current standard of care may
improve pharmacologic management and clinical outcomes for patients
with metastatic melanoma.
Acknowledgments
We would like to thank Ponney Palanisamy, Rachel
Graves and Ya Zhuo for their excellent technical support. M.P.,
A.G., B.B. and L.T.D. are active or retired employees of Merck,
which funded the study, and have financial interests. The present
study was funded by Merck & Co., Whitehouse Station, NJ,
USA.
References
|
1
|
Balch CM, Soong SJ, Gershenwald JE,
Thompson JF, Reintgen DS, Cascinelli N, Urist M, McMasters KM, Ross
MI, Kirkwood JM, et al: Prognostic factors analysis of 17,600
melanoma patients: Validation of the American Joint Committee on
Cancer melanoma staging system. J Clin Oncol. 19:3622–3634.
2001.PubMed/NCBI
|
|
2
|
Cummins DL, Cummins JM, Pantle H,
Silverman MA, Leonard AL and Chanmugam A: Cutaneous malignant
melanoma. Mayo Clin Proc. 81:500–507. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Culos KA and Cuellar S: Novel targets in
the treatment of advanced melanoma: New first-line treatment
options. Ann Pharmacother. 47:519–526. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ma C and Armstrong AW: Severe adverse
events from the treatment of advanced melanoma: A systematic review
of severe side effects associated with ipilimumab, vemurafenib,
interferon alfa-2b, dacarbazine and interleukin-2. J Dermatolog
Treat. 25:401–408. 2014. View Article : Google Scholar
|
|
5
|
Kuphal S, Bauer R and Bosserhoff AK:
Integrin signaling in malignant melanoma. Cancer Metastasis Rev.
24:195–222. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Marshall JF, Rutherford DC, Happerfield L,
Hanby A, McCartney AC, Newton-Bishop J and Hart IR: Comparative
analysis of integrins in vitro and in vivo in uveal and cutaneous
melanomas. Br J Cancer. 77:522–529. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nip J and Brodt P: The role of the
integrin vitronectin receptor, alpha v beta 3 in melanoma
metastasis. Cancer Metastasis Rev. 14:241–252. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Johnson JP: Cell adhesion molecules in the
development and progression of malignant melanoma. Cancer
Metastasis Rev. 18:345–357. 1999. View Article : Google Scholar
|
|
9
|
Desgrosellier JS and Cheresh DA: Integrins
in cancer: Biological implications and therapeutic opportunities.
Nat Rev Cancer. 10:9–22. 2010. View
Article : Google Scholar
|
|
10
|
Jin H and Varner J: Integrins: Roles in
cancer development and as treatment targets. Br J Cancer.
90:561–565. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Petitclerc E, Strömblad S, von Schalscha
TL, Mitjans F, Piulats J, Montgomery AM, Cheresh DA and Brooks PC:
Integrin αvβ3 promotes M21 melanoma growth in
human skin by regulating tumor cell survival. Cancer Res.
59:2724–2730. 1999.PubMed/NCBI
|
|
12
|
Smith SD, Enge M, Bao W, Thullberg M,
Costa TD, Olofsson H, Gashi B, Selivanova G and Strömblad S:
Protein kinase Cα (PKCα) regulates p53 localization and melanoma
cell survival downstream of integrin αv in three-dimensional
collagen and in vivo. J Biol Chem. 287:29336–29347. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bao W and Strömblad S: Integrin
alphav-mediated inactivation of p53 controls a MEK1-dependent
melanoma cell survival pathway in three-dimensional collagen. J
Cell Biol. 167:745–756. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Horton MA: The alpha v beta 3 integrin
‘vitronectin receptor’. Int J Biochem Cell Biol. 29:721–725. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Eble: Integrin-Ligand Interaction. R.G.
Landes Company; Georgetown: 1997
|
|
16
|
Hsu MY, Shih DT, Meier FE, Van Belle P,
Hsu JY, Elder DE, Buck CA and Herlyn M: Adenoviral gene transfer of
beta3 integrin subunit induces conversion from radial to vertical
growth phase in primary human melanoma. Am J Pathol. 153:1435–1442.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Trikha M, Zhou Z, Timar J, Raso E, Kennel
M, Emmell E and Nakada MT: Multiple roles for platelet GPIIb/IIIa
and alphavbeta3 integrins in tumor growth, angiogenesis, and
metastasis. Cancer Res. 62:2824–2833. 2002.PubMed/NCBI
|
|
18
|
Oliva IB, Coelho RM, Barcellos GG,
Saldanha-Gama R, Wermelinger LS, Marcinkiewicz C, Benedeta Zingali
R and Barja-Fidalgo C: Effect of RGD-disintegrins on melanoma cell
growth and metastasis: Involvement of the actin cytoskeleton, FAK
and c-Fos. Toxicon. 50:1053–1063. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kang IC, Kim DS, Jang Y and Chung KH:
Suppressive mechanism of salmosin, a novel disintegrin in B16
melanoma cell metastasis. Biochem Biophys Res Commun. 275:169–173.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hutchinson JH, Halczenko W, Brashear KM,
Breslin MJ, Coleman PJ, Duong T, Fernandez-Metzler C, Gentile MA,
Fisher JE, Hartman GD, et al: Nonpeptide alphavbeta3 antagonists.
8. In vitro and in vivo evaluation of a potent alphavbeta3
antagonist for the prevention and treatment of osteoporosis. J Med
Chem. 46:4790–4798. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Murphy MG, Cerchio K, Stoch SA,
Gottesdiener K, Wu M and Recker R; L-000845704 Study Group: Effect
of L-000845704, an alphaVbeta3 integrin antagonist, on markers of
bone turnover and bone mineral density in postmenopausal
osteoporotic women. J Clin Endocrinol Metab. 90:2022–2028. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rosenthal MA, Davidson P, Rolland F,
Campone M, Xue L, Han TH, Mehta A, Berd Y, He W and Lombardi A:
Evaluation of the safety, pharmacokinetics and treatment effects of
an αvβ3 integrin inhibitor on bone turnover
and disease activity in men with hormone-refractory prostate cancer
and bone metastases. Asia Pac J Clin Oncol. 6:42–48. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Simon KO, Nutt EM, Abraham DG, Rodan GA
and Duong LT: The alphavbeta3 integrin regulates
alpha5beta1-mediated cell migration toward fibronectin. J Biol
Chem. 272:29380–29389. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Abraham DG, Nutt EM, Bednar RA, Bednar B,
Gould RJ and Duong LT: Arginine-glycine-aspartic acid mimics can
identify a transitional activation state of recombinant alphaIIb
beta3 in human embryonic kidney 293 cells. Mol Pharmacol.
52:227–236. 1997.PubMed/NCBI
|
|
25
|
Kossodo S, Pickarski M, Lin SA, Gleason A,
Gaspar R, Buono C, Ho G, Blusztajn A, Cuneo G, Zhang J, et al: Dual
in vivo quantification of integrin-targeted and protease-activated
agents in cancer using fluorescence molecular tomography (FMT). Mol
Imaging Biol. 12:488–499. 2010. View Article : Google Scholar
|
|
26
|
Pickarski M, Hayami T, Zhuo Y and Duong T:
Molecular changes in articular cartilage and subchondral bone in
the rat anterior cruciate ligament transection and meniscectomized
models of osteoarthritis. BMC Musculoskelet Disord. 12:1972011.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fidler IJ: Biological behavior of
malignant melanoma cells correlated to their survival in vivo.
Cancer Res. 35:218–224. 1975.PubMed/NCBI
|
|
28
|
Fidler IJ and Kripke ML: Metastasis
results from preexisting variant cells within a malignant tumor.
Science. 197:893–895. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cowden Dahl KD, Robertson SE, Weaver VM
and Simon MC: Hypoxia-inducible factor regulates alphavbeta3
integrin cell surface expression. Mol Biol Cell. 16:1901–1912.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Barczyk M, Carracedo S and Gullberg D:
Integrins. Cell Tissue Res. 339:269–280. 2010. View Article : Google Scholar
|
|
31
|
Giancotti FG and Ruoslahti E: Integrin
signaling. Science. 285:1028–1032. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Van Belle PA, Elenitsas R, Satyamoorthy K,
Wolfe JT, Guerry D IV, Schuchter L, Van Belle TJ, Albelda S, Tahin
P, Herlyn M, et al: Progression-related expression of beta3
integrin in melanomas and nevi. Hum Pathol. 30:562–567. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Albelda SM, Mette SA, Elder DE, Stewart R,
Damjanovich L, Herlyn M and Buck CA: Integrin distribution in
malignant melanoma: Association of the beta 3 subunit with tumor
progression. Cancer Res. 50:6757–6764. 1990.PubMed/NCBI
|
|
34
|
Brooks PC: Role of integrins in
angiogenesis. Eur J Cancer. 32A:2423–2429. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Brooks PC, Strömblad S, Sanders LC, von
Schalscha TL, Aimes RT, Stetler-Stevenson WG, Quigley JP and
Cheresh DA: Localization of matrix metalloproteinase MMP-2 to the
surface of invasive cells by interaction with integrin alpha v beta
3. Cell. 85:683–693. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Danen EHJ: Integrin signaling as a cancer
drug target. ISRN Cell Biol. 2013:1351642013.http://dx.doi.org/10.1155/2013/135164.
Accessed: Jan 10, 2015. View Article : Google Scholar
|
|
37
|
Reardon DA, Fink KL, Mikkelsen T,
Cloughesy TF, O’Neill A, Plotkin S, Glantz M, Ravin P, Raizer JJ,
Rich KM, et al: Randomized phase II study of cilengitide, an
integrin-targeting arginine-glycine-aspartic acid peptide, in
recurrent glioblastoma multiforme. J Clin Oncol. 26:5610–5617.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kim KB, Prieto V, Joseph RW, Diwan AH,
Gallick GE, Papadopoulos NE, Bedikian AY, Camacho LH, Hwu P, Ng CS,
et al: A randomized phase II study of cilengitide (EMD 121974) in
patients with metastatic melanoma. Melanoma Res. 22:294–301. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gutheil JC, Campbell TN, Pierce PR,
Watkins JD, Huse WD, Bodkin DJ and Cheresh DA: Targeted
antiangiogenic therapy for cancer using Vitaxin: A humanized
monoclonal antibody to the integrin alphavbeta3. Clin Cancer Res.
6:3056–3061. 2000.PubMed/NCBI
|
|
40
|
Mcneel DG, Eickhoff J, Lee FT, King DM,
Alberti D, Thomas JP, Friedl A, Kolesar J, Marnocha R, Volkman J,
et al: Phase I trial of a monoclonal antibody specific for
alphavbeta3 integrin (MEDI-522) in patients with advanced
malignancies, including an assessment of effect on tumor perfusion.
Clin Cancer Res. 11:7851–7860. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hersey P, Sosman J, O’Day S, Richards J,
Bedikian A, Gonzalez R, Sharfman W, Weber R, Logan T, Buzoianu M,
et al: Etaracizumab Melanoma Study Group: A randomized phase 2
study of etaracizumab, a monoclonal antibody against integrin
αvβ3, ± dacarbazine in patients with stage IV
metastatic melanoma. Cancer. 116:1526–1534. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Reynolds AR, Hart IR, Watson AR, Welti JC,
Silva RG, Robinson SD, Da Violante G, Gourlaouen M, Salih M, Jones
MC, et al: Stimulation of tumor growth and angiogenesis by low
concentrations of RGD-mimetic integrin inhibitors. Nat Med.
15:392–400. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Girotti MR, Saturno G, Lorigan P and
Marais R: No longer an untreatable disease: How targeted and
immunotherapies have changed the management of melanoma patients.
Mol Oncol. 8:1140–1158. 2014. View Article : Google Scholar : PubMed/NCBI
|