Introduction
Cervical cancer remains the second leading
malignancy for women worldwide (1,2),
although the incidence of this type of cancer is on the decrease in
developed countries due to early diagnosis. Nevertheless, cervical
cancer remains a serious health issue for women in developing
countries, such as China, where diagnostic programs are not well
established. Moreover, local recurrence remains challenging for
those patients with cervical cancers (particularly at the advanced
stage) (3), even though such
therapies as surgery, chemotherapy and/or radiotherapy have been
utilized.
Persistent cervical infection with high-risk human
papillomavirus (HPV), particularly with HPV type 16 or 18,
contributes to the development of cervical cancer (4–6). The
early oncoproteins of HPVs E5, E6 and E7 are known to contribute to
tumor progression, such as the proliferation, migration and
invasion of cervical cancer cells (7). Accumulating evidence has identified
molecular mechanisms involved in cervical cancer. Molecules such as
vascular endothelial growth factor-C (VEGF-C) (8), Src homology-2 domain containing
protein, tyrosine phosphatase-2 (SHP-2) (9), or CD147 isoform-4 (CD147-4) (10) are known to promote the
proliferation, migration or invasion of cervical cancers by
activating focal adhesion kinases (8), through inhibition of interferon-β
production (9), or with an
upregulated expression of the cancerous inhibitors PP2A (CIP2A),
polo-like kinase (PLK) and cyclin D1 but a downregulated p27
expression (10). By contrast,
tumor-suppressive molecules, such as Beclin1 (11) and histone deacetylase (HDAC) 10
(12) have been confirmed to
inhibit the invasion and migration of cervical cancer cells by
decreasing the expression of VEGF and matrix metalloproteinase
(MMP)-9 proteins (11), or through
the inhibition of MMP-2 and -9 expression (12).
A type of non-coding RNA with ~22 nucleotides, known
as microRNAs (miRNAs) (13), has
been found to be important in the development and progression of
cervical cancers (14–16). Dysregulated miRNAs, such as miR-135a
(17), miR-10a (18), or miR-205 (19) promote cell growth, migration and
invasion in human cervical cancer cells, by regulating β-catenin
(17), by targeting CHL1 (18), or by downregulating CYR61 and CTGF
(19). Some tumor-suppressive
microRNAs, such as miR-218 (20),
miR-372 (21), or miR-214 (22) are also deregulated in cervical
cancers and contribute to cancer progression. In addition, the
oncogenic miR-21 has been widely recognized to play a role in
non-small cell lung cancers (NSCLCs) (23,24),
as well as colorectal (25),
ovarian (26), breast (27) and esophageal cancers (28). Accumulating evidence shows that the
promoting role of miR-21 in NSCLCs or in colorectal cancers occurs
through the modulation of the phosphatase and the tensin homolog
(PTEN) signaling pathway (24,25).
Moreover, miR-21 has been demonstrated to be deregulated in
cervical cancers, with a marked association with the worsening
clinical diagnosis of cervical cancers (29).
In the present study, we examined the expression of
miR-21 and PTEN in cervical cancer specimens, and investigated the
regulation of miR-21 and PTEN on the proliferation and migration of
the cervical cancer CaSki and HeLa cells. We also determined the
regulation of miR-21 on the PTEN expression. The present study
demonstrated the regulation of miR-21 on the progression of the
cervical cancer cells.
Materials and methods
Human tissue specimens
The 36 invasive cervical cancer and 21 normal human
cervical tissue specimens were collected using surgical resection
prior to radiotherapy or chemotherapy. The tissue specimens were
stored at −80°C prior to utilization. Utilization of the cervical
cancer specimens and normal cervical tissues was approved by the
Hospital Internal Review Board (IRB) in our hospital.
Cell culture and recombinant plasmid
transfection
The CaSki and HeLa cervical cancer cell lines were
purchased from the American Type Culture Collection (Manassas, VA,
USA) and grown, respectively, in RPMI-1640 medium (Sigma-Aldrich,
St. Louis, MO, USA) or Eagle’s minimum Essential medium (EMEM;
Invitrogen, Carlsbad, CA, USA) supplemented with 10% fetal bovine
serum (FBS; Gibco, Rockville, MD, USA). The two cell types were
incubated at 37°C, with 5% CO2. miR-21 mimics, miR-21
inhibitor or miRNA control (Qiagen, Valencia, CA, USA) were
utilized to manipulate the miR-21 level. Then, 25 or 50 nM miR-21
mimics, miR-21 inhibitor or miRNA control were transfected with
Lipofectamine 2000 (Invitrogen) into the CaSki or HeLa cells.
To overexpress PTEN in the CaSki cells, we
constructed a recombinant plasmid, PTEN-pcDNA3.1 (+) by cloning the
PTEN (GenBank accession no. NG_007466.1) coding sequence into a
eukaryotic expression vector, pcDNA3.1 (+) (Invitrogen). To
overexpress the PTEN in the CaSki cells, the cells were transfected
with the PTEN-pcDNA3.1 (+) or CAT-pcDNA3.1 (+) plasmid. The
post-transfected cells were then cultured for 24 h to determine
PTEN expression at the mRNA and protein levels. For the
proliferation assay, the PTEN-overexpressed cells [CaSki PTEN (+)]
or the CAT-overexpressed cells (CaSki control) were cultured for
24, 48 or 72 h. For the colony-forming assay, the CaSki PTEN (+)
cells or the CaSki control cells were cultured for 48 h.
RNA extraction and reverse
transcriptase-quantitative PCR (RT-qPCR)
Cellular mRNA was extracted from cervical cancer
specimens or from cell samples using TRIzol reagent (Thermo Fisher
Scientific, Waltham, MA, USA). cDNA from each sample was
synthesized with the Superscript First-Strand Synthesis System for
the RT-PCR kit (Gibco-BRL, Grand Island, NY, USA) with a random
Uni-12 primer. The PTEN mRNA was quantified by RT-qPCR performed
using a TaqMan Assay based on real-time detection in a LightCycler
2.0 (Roche Diagnostics GmbH, Mannheim, Germany). The PTEN mRNA
level in each sample was normalized to β-actin. miRNA extraction
was performed using the mirVana miRNA Isolation kit (Ambion,
Austin, TX, USA). Quantification of the miR-21 level in the
cervical cancer specimens and cell samples was conducted using the
mirVana RT-qPCR miRNA Detection kit (Ambion), with the U6 small
nuclear RNA used as the internal control. The ΔΔCt method was used
for relative quantification (30),
and the PTEN mRNA or the miR-21 level was expressed as a relative
value to the control group.
CCK-8 assay, cell colony-forming assay
and proliferation
The Cell proliferation was examined using the CCK-8
assay. Briefly, CaSki or HeLa cells post-transfected with miR-21
mimics, miR-21 inhibitor or miR-21 control were incubated in CCK-8
(Dojindo, Kumamoto, Japan). Absorbance at 450 nm of the treated
cells was detected following incubation at 37°C, with 5%
CO2, for 24, 36 or 48 h. For the cell colony-forming
assay, 300 CaSki or HeLa cells were incubated in 12-well plates at
37°C, 5% CO2, and were then transfected with or without
50 nM miR-21 mimics, miR-21 inhibitor or miRNA control. Following
48-h post-incubation, the cells were stained with crystal violet
(0.005%) for 20 min and the colony numbers were recorded using
Image J software.
Cell migration and invasion assay
The ability of cell migration was examined by the
wound-healing assay. The cells were seeded in 12-well plates and
cultivated to a confluence of 85%. The 85% confluent HeLa cells
were then transfected with miR-21 mimics, miR-21 inhibitor or
miR-21 control and were scratched with Cell Scrapers (Corning Inc.,
Corning, NY, USA), 6 h post-transfection. Cell growth was observed
at 0, 24 and 48 h. The HeLa cells that migrated across the baseline
were counted under an optical microscope. Cell invasion was
examined by the Matrigel-coated Transwell assay. Briefly, the cells
were seeded at a density of 1×105 cells in serum-free
medium on the upper chamber with the a non-coated membrane (8
μm pore size; Millipore, Zug, Switzerland). The lower
chamber contained medium with 20% FBS as a chemoattractant. The
cells in the upper chamber were discarded using cotton wool after
24 h and the migratory cells in the lower chamber were counted
under an optical microscope. The experiments were repeated in
triplicate.
Statistical analysis
Data are presented as the mean ± SEM. Statistical
analysis was performed using the SPSS 18.0 software (IBM SPSS,
Armonk, NY, USA). Correlations between the miR-21 and PTEN mRNA
level in cervical cancer specimens were analyzed using the
Spearman’s rank correlation. PTEN expression at the mRNA or the
protein levels, miR-21 expression, the colony-forming and the
migratory cells between the two groups were analyzed using the
Student’s t-test. The cell proliferation difference among miR-21
mimics, miR-21 inhibitor and miRNA control groups was analyzed by
the two-way ANOVA test. P≤0.05 was considered to indicate a
statistically significant difference.
Results
miR-21 is overexpressed in invasive
cervical cancer specimens, in association with a reduced PTEN
expression
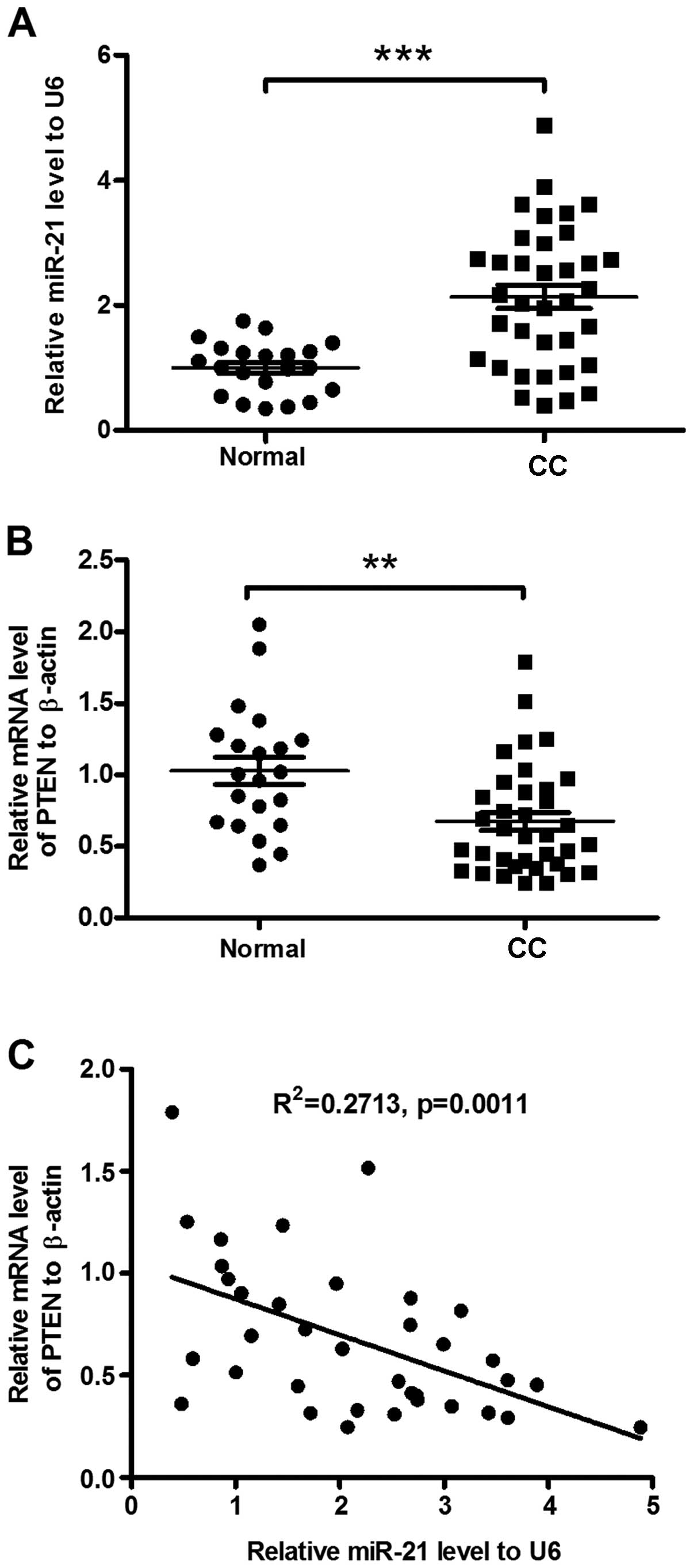
The miR-21 level in the cervical cancer specimens
was examined using RT-qPCR, compared to the normal cervical
tissues. Thirty-six invasive cervical cancer patients with an
average diagnosis age of 52 years were included in the study. Of
the 36 subjects, 28 were HPV-positive and 8 were HPV-negative. The
mean value of miR-21 was 2.14±0.19 in the 36 samples from patients
with cervical cancer and 1.00±0.09 in the healthy controls
(P<0.001, Fig. 1A). No
significant difference was identified in the miR-21 level between
the HPV-positive and -negative samples. Thus, miR-21 was confirmed
to be significantly upregulated in the invasive cervical cancer
specimens. It has been indicated that the tumor-suppressive
phosphatase and tensin homolog (PTEN) (31) is downregulated in cervical cancers
(29,32,33).
To investigate the association of the upregulated miR-21 level with
the downregulated PTEN in cervical cancers, we examined the
expression of PTEN mRNA. As shown in Fig. 1B, there was a significant reduction
of PTEN mRNA in the invasive cervical cancer specimens (P<0.01).
Downregulation of PTEN correlated with the miR-21 upregulation in
the specimens (R2=0.2713, P=0.0011, Fig. 1C). Thus, miR-21 overexpression was
confirmed in the invasive cervical cancer specimens, in association
with the PTEN downregulation.
Manipulation of the miR-21 level
regulates the proliferation of cervical cancer cells
To identify the regulation of miR-21 on the
proliferation of the cervical cancer cells, we manipulated the
miR-21 level in the CaSki and HeLa cells by transfecting the cells
with miR-21 mimics, miR-21 inhibitor or miRNA control. There was a
significant increase or reduction in the miR-21 level in the CaSki
or HeLa cells post-transfected with miR-21 mimics or miR-21
inhibitor (P<0.01 or P<0.0001 for the miR-21 inhibitor or
miR-21 mimics in the CaSki or HeLa cells, Fig. 2A). Proliferation of the CaSki and
HeLa cells following transfection with miR-21 mimics, miR-21
inhibitor or miRNA control was assessed using the CCK-8 assay. In
the CaSki or HeLa cells, the miR-21 mimics promoted cell
proliferation instead of miRNA control (P<0.01 and P<0.05 for
the CaSki and HeLa cells, respectively, Fig. 2B), whereas the miR-21 inhibitor
transfection inhibited the proliferation of the CaSki and HeLa
cells (both P<0.05, Fig. 2B).
There was also a time dependence in the regulation on the
proliferation of the CaSki (Fig.
2C) or HeLa cells (Fig.
2D).
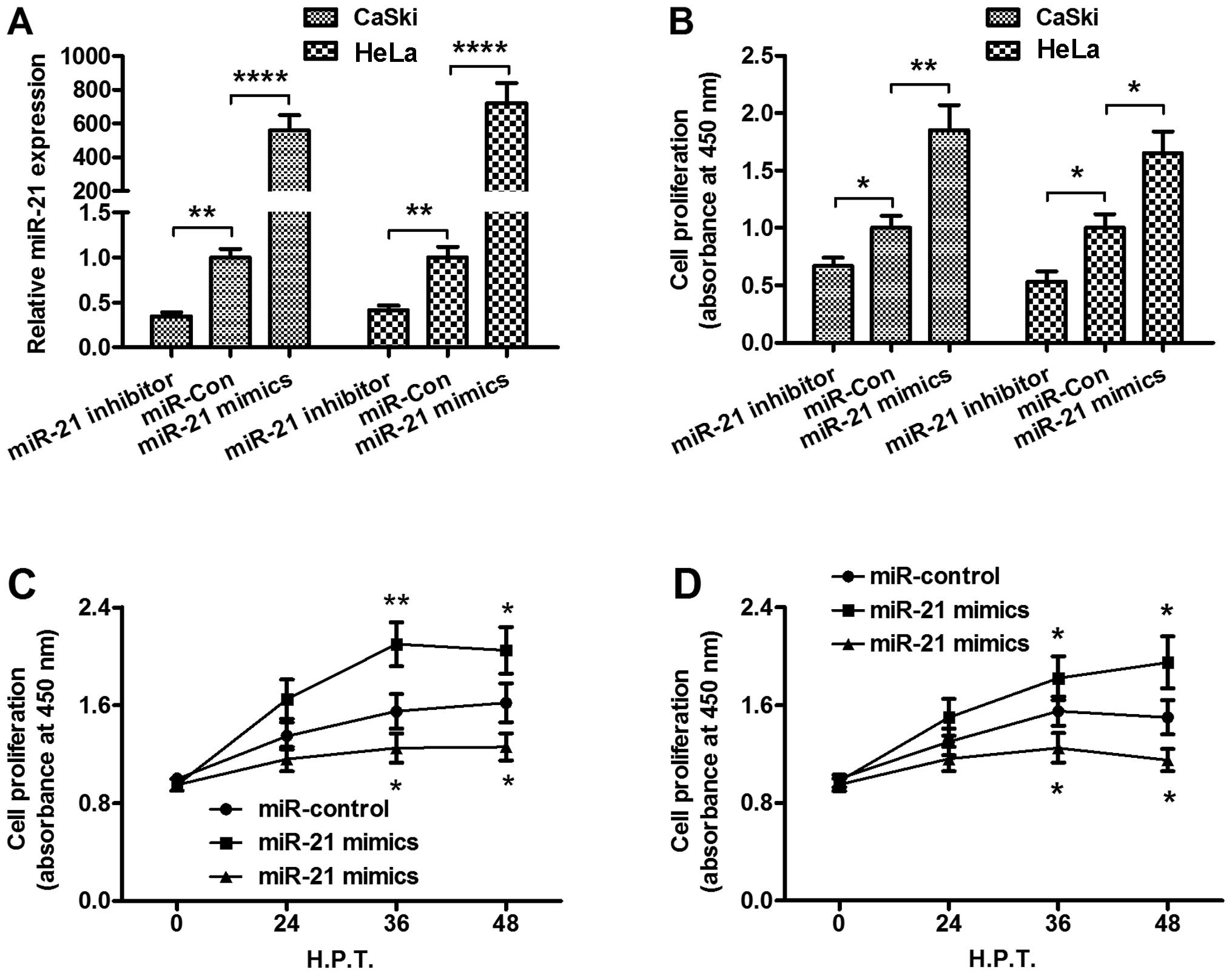 | Figure 2miR-21 manipulations with miR-21
mimics or miR-21 inhibitor promote or inhibit the proliferation of
CaSki or HeLa cells in vitro. (A) The up- or downregulation
of miR-21 by transfection with miR-21 mimics or with miR-21
inhibitor in the CaSki and HeLa cells. The miR-21 level was
examined using RT-qPCR, 24 h following transfection with 25 nM
miR-control, miR-21 mimics or miR-21 inhibitor. (B) CCK-8 assay for
the relative proliferation of the CaSki or HeLa cells, 24 h
following transfection with 25 nM miR-control, miR-21 mimics or
miR-21 inhibitor. Growth curve of the (C) CaSki or (D) HeLa cells,
following transfection with 25 nM miR-control, miR-21 mimics or
miR-21 inhibitor, examined by the CCK-8 assay. Each value was
averaged for three independent results. *P<0.05,
**P<0.01 and ****P<0.0001,
statistically significant differences. CCK-8, Cell Counting
Kit-8. |
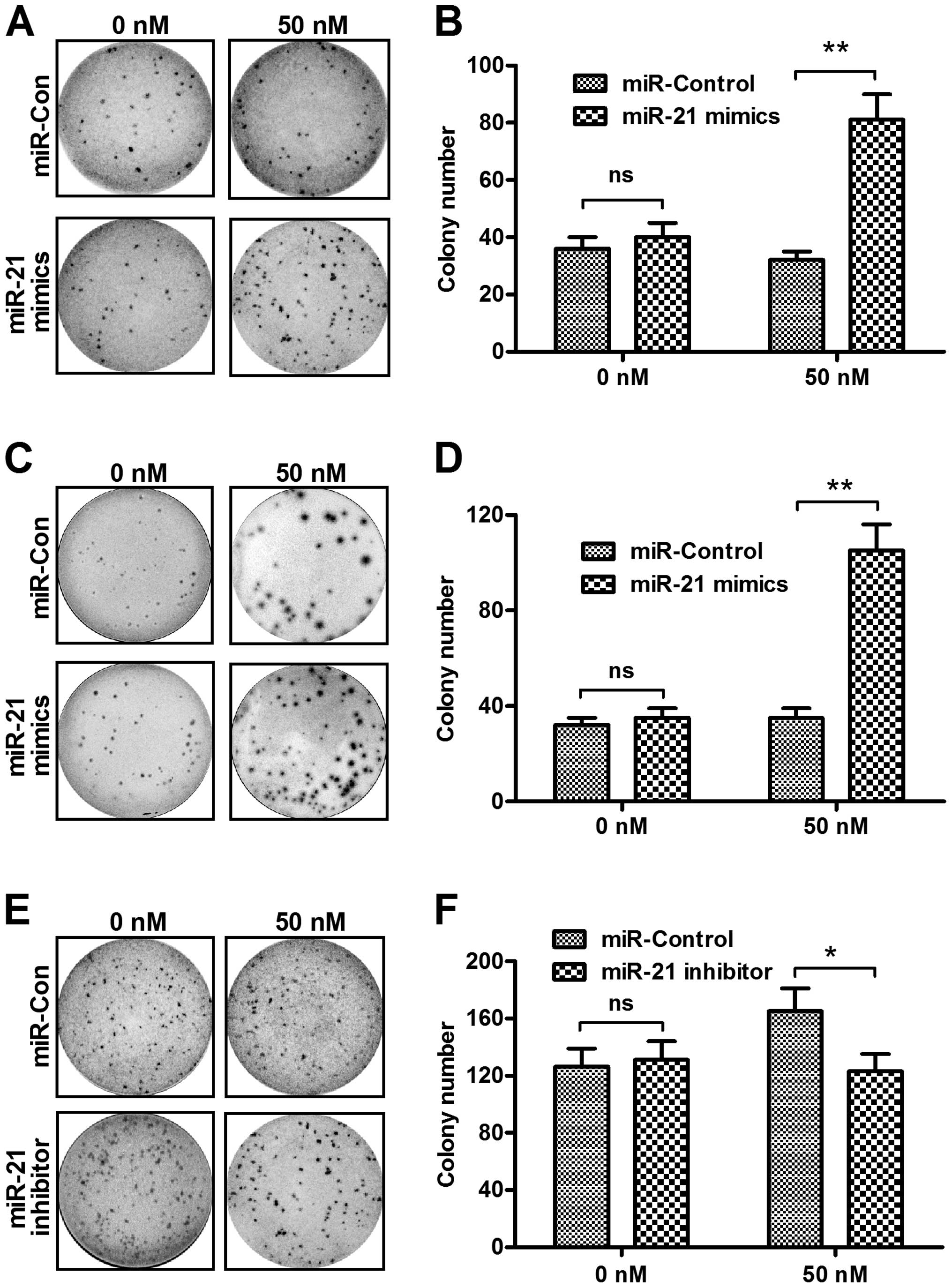
To confirm the proliferation regulation by miR-21
manipulation, we performed a colony formation assay of the CaSki
and HeLa cells transfected with miR-21 mimics, miR-21 inhibitor or
miRNA control. Fig. 3A shows that,
the CaSki cells formed more colonies following transfection with 50
nM miR-21 mimics as compared to the control miRNA (Fig. 3B, P<0.01). The promotion of
colony formation by the miR-21 mimics transfection was reconfirmed
in HeLa cells and the transfection with the miR-21 mimics promoted
more colonies in HeLa cells than the transfection with control
miRNA (Fig. 3C and D, P<0.01).
On the other hand, the miR-21 inhibitor suppressed colony formation
in the CaSki cells with a transfection concentration of 50 nM
(Fig. 3E and F, P<0.05). These
findings demonstrated that upregulated miR-21 enhanced the
proliferative ability and colony formation of cervical cancer
cells.
Manipulation of miR-21 level regulates
the migration and invasion of cervical cancer cells
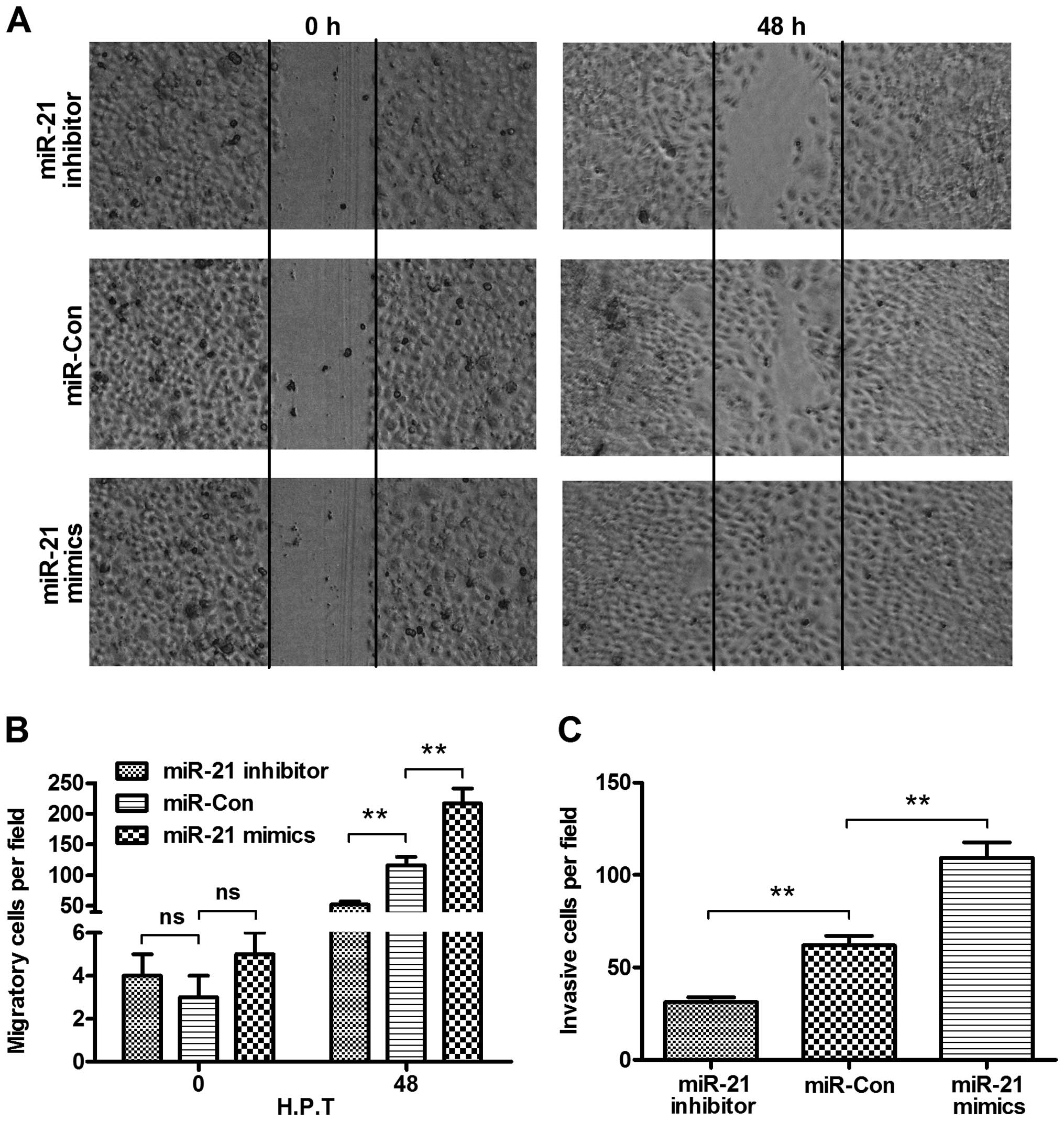
To identify the oncogenic promotion of miR-21, the
migration and invasion of the cervical cancer cells were evaluated
using the wound-healing and Transwell assays. Firstly, the
difference in the migration of HeLa cells post-transfected with 50
nM miR-21 mimics, miR-21 inhibitor or control miRNA using a
wound-healing assay was determined. Fig. 4A shows that there were more HeLa
cells migrating across the baseline in the miR-21 mimics
transfection group as compared to that in the control miRNA
transfection group (P<0.01, Fig.
4B), whereas the miR-21 inhibitor reduced HeLa cell migration
as compared to that in the control group (P<0.01, Fig. 4B). In addition, we investigated cell
invasion using the Transwell assay. Fig. 4C shows that the invaded cells in the
miR-21 mimics transfection group were significantly more than those
in the control miRNA group (P<0.01), whereas the invaded cells
in the miR-21 inhibitor transfection group were significantly less
than those in the control miRNA group (P<0.01). Collectively,
miR-21 promoted the migration and invasion of cervical cancer
cells.
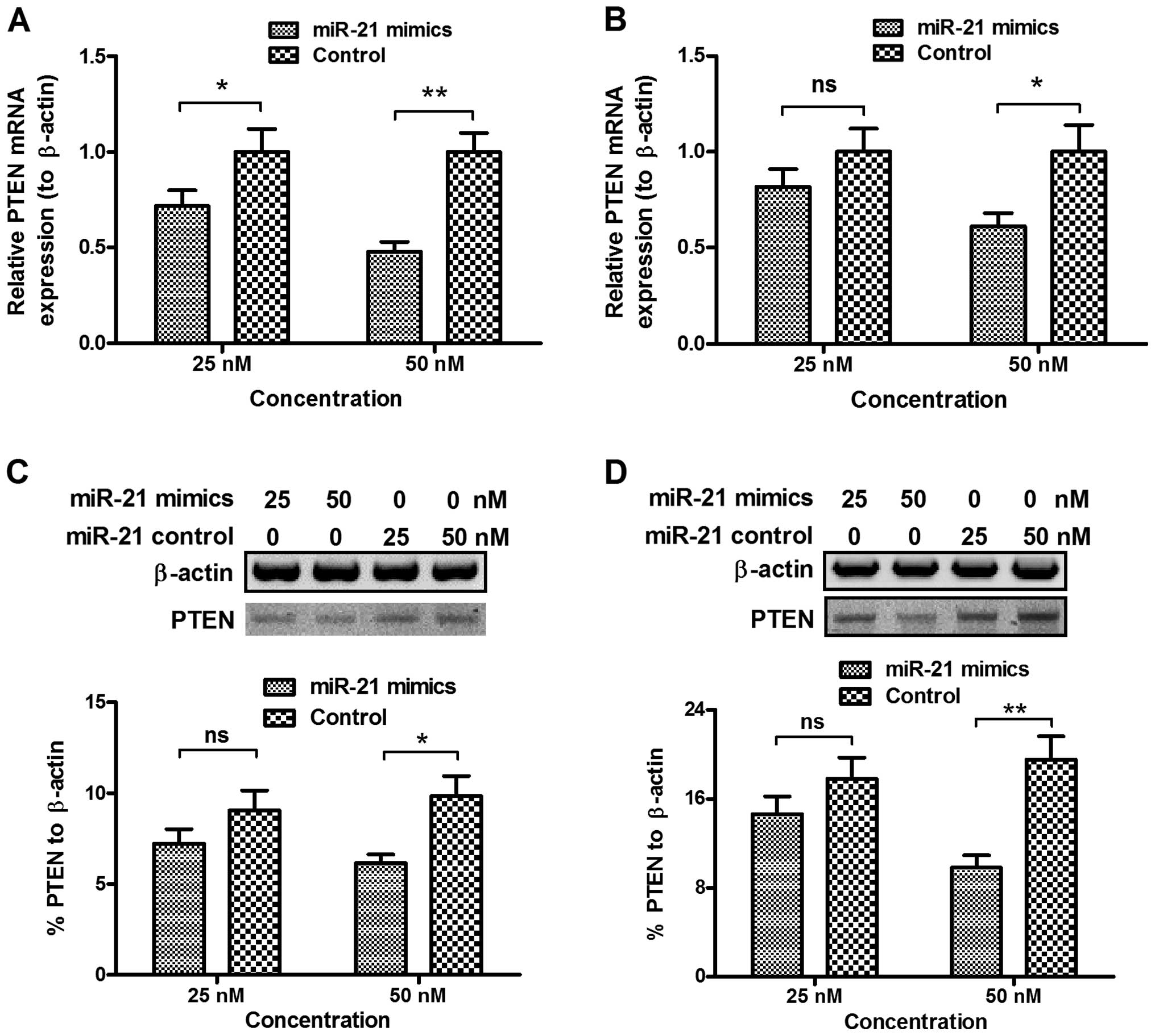
miR-21 mimics transfection significantly
downregulates the PTEN level in cervical cancer cells
To identify the mechanism involved in the promotion
of miR-21 to the proliferation of the CaSki or HeLa cells, and to
investigate the regulation of PTEN expression by miR-21 in the
cervical cancer cells, we examined PTEN expression at the mRNA and
protein levels in the CaSki and HeLa cells following transfection
with miR-21 mimics or miRNA control. PTEN was downregulated at the
mRNA level in the CaSki cells by transfection with 25 or 50 nM
miR-21 mimics (P<0.05 for 25 nM or P<0.01 for 50 nM, Fig. 5A). This downregulation in the PTEN
mRNA was confirmed in the HeLa cells, where transfection with 50 nM
miR-21 mimics significantly downregulated PTEN mRNA in the HeLa
cells (P<0.05, Fig. 5A). By
contrast, there was no regulation of the PTEN mRNA level by the
control miRNA transfection (Fig. 5A and
B). To reconfirm the PTEN downregulation by miR-21, we analyzed
PTEN expression at the protein level which was examined in cells
following transfection with miR-21 mimics or miRNA control.
Fig. 5C shows that transfection
with 50 nM miR-21 mimics, instead of the 50 nM control miRNA
reduced PTEN expression at the protein level (P<0.05).
Furthermore, the PTEN reduction by transfection with 50 nM miR-21
mimics was observed in the HeLa cells (P<0.01, Fig. 5D). Therefore, miR-21 mimics
transfection significantly downregulated the PTEN level in the
cervical cancer cells.
PTEN overexpression inhibits cervical
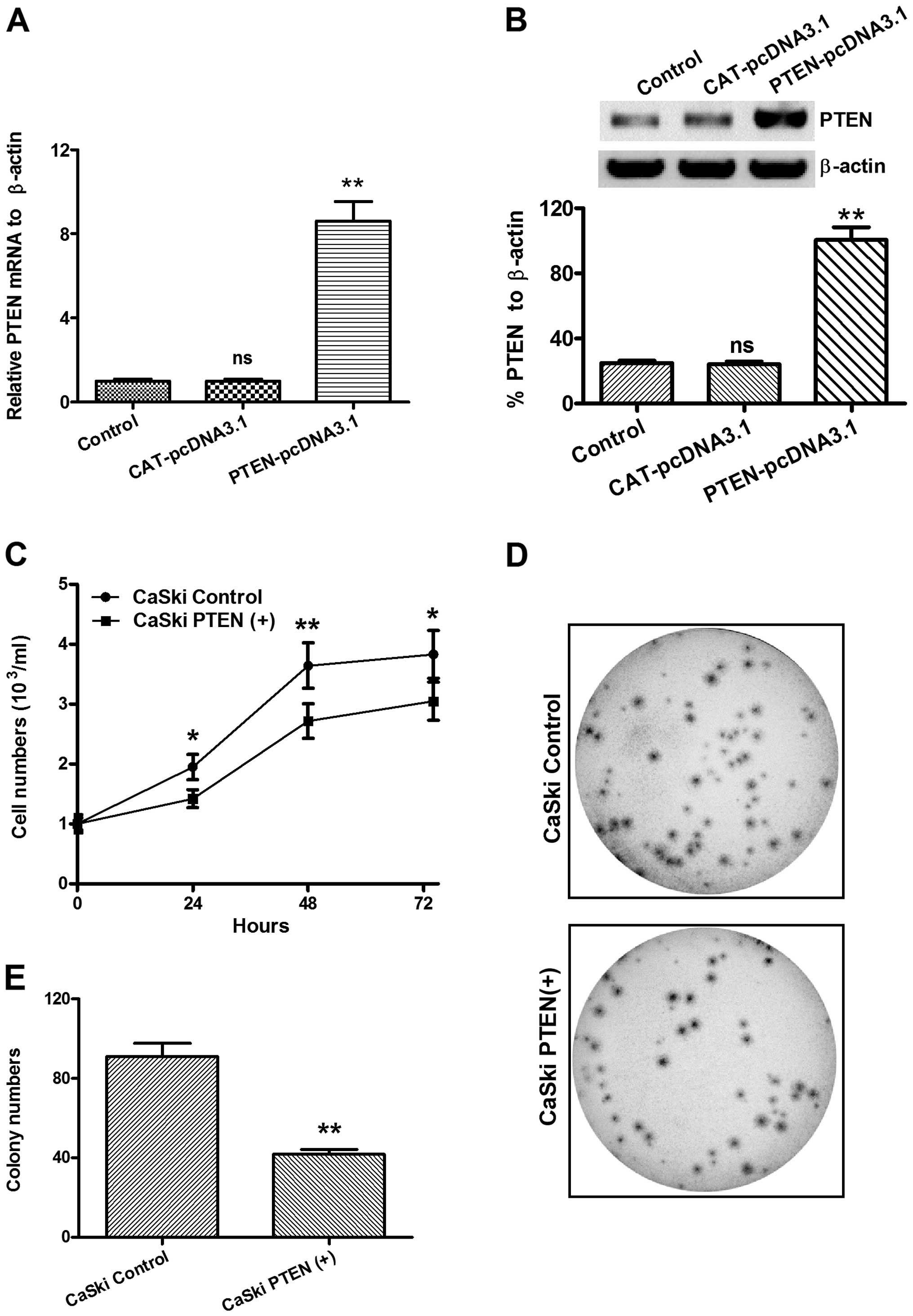
cancer cell proliferation
To determine whether miR-21-promoted cervical cancer
proliferation was mediated by the downregulation of PTEN, we
overexpressed PTEN with a eukaryotic expression vector,
pcDNA3.1(+), in the CaSki cells. Then, we examined the
proliferation of CaSki cells with PTEN over-expressed [CaSki
PTEN(+)] or CaSki cells transfected with CAT-pcDNA3.1(+) (CaSki
control) using the CCK-8 assay. Figs.
6A and B shows that there was a significantly overexpressed
PTEN at the mRNA (Fig. 6A) and
protein (Fig. 6B) levels in the
CaSki PTEN(+) cells, compared to the CaSki control cells.
Furthermore, the CCK-8 assay indicated that, from 24 to 72 h
following incubation, the CaSki PTEN(+) cells grew to a
significantly lower level than the CaSki control cells (Fig. 6C, P<0.05 for 24 or 72 h and
P<0.01 for 48 h). As shown in Fig.
6D and E, the colony-forming assay confirmed the lower
proliferation of CaSki PTEN(+) cells, compared to the CaSki control
cells (P<0.01). These results indicated that the overexpression
of PTEN inhibited the proliferation of cervical cancer cells.
Discussion
Increased miR-21 has been identified in various
types of cancer (34–38), such as colorectal (25), hepatocellular (35) and breast cancers (27,36,37).
It has been reported that miR-21 expression increased with
worsening clinical diagnosis in cervical cancers (29). The cytoplasmic expression of the
programmed cell death 4 (PDCD4) protein, which is a known target of
miR-21 (39), was also
significantly lower in women with invasive cervical carcinoma (ICC)
(29). However, there was no
significant correlation between miR-21 overexpression and PDCD4
downregulation. Thus, there may be other target molecules for
miR-21 in invasive cervical cancers. The PTEN gene is mutated or
abrogated in a wide range of human cancers (40–42).
The tumor suppressive PTEN inhibits cell migration and invasion by
directly dephosphorylating two key tyrosine-phosphorylated
proteins, thereby antagonizing the interactions of integrins with
the extracellular matrix and integrin-triggered signaling pathways
(43,44). The dephosphorylating role of PTEN is
also necessary in a lipid signal transduction pathway (45). Therefore, PTEN performs as a unique
tumor suppressor through the inhibition of lipid phosphatase and
protein tyrosine phosphatase activities, thus negatively regulating
cell proliferation and invasion. The tumor suppressive PTEN gene
has also been reported to mutate or to decrease in expression in
cervical cancers (32,46). However, its role in the
proliferation or invasion of cervical cancers needs to be
determined in detail.
In the present study, we found that miR-21 was
overexpressed, whereas PTEN was downregulated in the invasive
cervical cancer specimens, and that miR-21 overexpression was
associated with the downregulated PTEN in such cervical cancer
specimens. To recognize the oncogenic role of miR-21 in the
cervical cancer cells, we manipulated miR-21, with miR-21 mimics or
miR-21 inhibitor in the CaSki and HeLa cervical cancer cell lines,
and examined the regulation of miR-21 on cell proliferation and
migration. Results in the present study confirm that the
proliferation of CaSki or HeLa cells was promoted by transfection
with miR-21 mimics, whereas prolife ration was inhibited by
transfection with the miR-21 inhibitor, in a time-dependent manner.
Furthermore, the migration and invasion of the cervical cancer
cells was evaluated by the wound-healing and Transwell assays,
following miR-21 manipulation. The results show that there were
more HeLa cells migrating across the baseline in the miR-21 mimics
transfection group, in comparison to the control miRNA
transfection, whereas the miR-21 inhibitor reduced HeLa cell
migration, compared to that in the control group. In addition, the
Transwell assay indicated that the invaded cells were more in the
miR-21 mimics transfection group than in the control miRNA group.
Collectively, miR-21 promoted the prolife ration, migration and
invasion of the cervical cancer cells.
Given the regulation by miR-21 on PTEN expression in
various types of cancer (24,25,47),
PTEN expression at the mRNA and protein levels was also examined in
the CaSki or HeLa cells following transfection with miR-21 mimics
or miRNA control. The results show that PTEN was down-regulated at
the mRNA and protein levels in the CaSki or HeLa cells by miR-21
mimics transfection, in comparison to the control miRNA
transfection. Therefore, miR-21 mimics transfection significantly
downregulated the PTEN level in the cervical cancer cells.
Moreover, to identify the association of miR-21-promoted cervical
cancer, proliferation and invasion were mediated by the
downregulation of PTEN. We then overexpressed PTEN in the CaSki
cells and investigated the regulation of CaSki cell proliferation
by PTEN overexpression. The results show that the manipulated
upregulation of the PTEN expression at the mRNA and protein levels
in the CaSki cells significantly reduced cell proliferation.
In summary, the present study has shown the
upregulation of miR-21 in invasive cervical cancers and confirms
the promotion of miR-21 to the proliferation, migration and
invasion in the CaSki or HeLa cervical cancer cells by
down-regulating the tumor-suppressive PTEN expression. To the best
of our knowledge, this is the first study to confirm that the
miR-21/PTEN pathway promotes cervical cancers, suggesting that the
miR-21/PTEN pathway may be an effective target for cervical cancer
treatment.
References
|
1
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Movva S, Rodriguez L, Arias-Pulido H and
Verschraegen C: Novel chemotherapy approaches for cervical cancer.
Cancer. 115:3166–3180. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sittidilokratna K, Cheewakriangkrai C,
Khunamornpong S and Siriaunkgul S: Recurrence patterns after
radical hysterectomy in stage IB1-IIA cervical. Asian Pac J Cancer
Prev. 11:499–502. 2010.PubMed/NCBI
|
|
4
|
zur Hausen H: Papillomavirus infections-a
major cause of human cancers. Biochim Biophys Acta. 1288:F55–F78.
1996.PubMed/NCBI
|
|
5
|
de Sanjose S, Quint WG, Alemany L, Geraets
DT, Klaustermeier JE, Lloveras B, Tous S, Felix A, Bravo LE, Shin
HR, et al: Retrospective international survey and HPV time trends
study group: Human papillomavirus genotype attribution in invasive
cervical cancer: a retrospective cross-sectional worldwide study.
Lancet Oncol. 11:1048–1056. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schiffman M, Castle PE, Jeronimo J,
Rodriguez AC and Wacholder S: Human papillomavirus and cervical
cancer. Lancet. 370:890–907. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liao S, Deng D, Zhang W, Hu X, Wang W,
Wang H, Lu Y, Wang S, Meng L and Ma D: Human papillomavirus 16/18
E5 promotes cervical cancer cell proliferation, migration and
invasion in vitro and accelerates tumor growth in vivo. Oncol Rep.
29:95–102. 2013.
|
|
8
|
Chen H, Suo K, Cheng Y, Zheng B and Xu L:
Vascular endothelial growth factor C enhances cervical cancer
migration and invasion via activation of focal adhesion kinase.
Gynecol Endocrinol. 29:20–24. 2013. View Article : Google Scholar
|
|
9
|
Meng F, Zhao X and Zhang S: SHP-2
phosphatase promotes cervical cancer cell proliferation through
inhibiting interferon-β production. J Obstet Gynaecol Res.
39:272–279. 2013. View Article : Google Scholar
|
|
10
|
Wu Y, Zhou X and Zheng PS: Involvement of
CD147 isoform 4 in the proliferation of SiHa cells: A possible
molecular mechanism of cervical cancer. Oncol Rep. 26:717–724.
2011.PubMed/NCBI
|
|
11
|
Sun Y, Liu JH, Sui YX, Jin L, Yang Y, Lin
SM and Shi H: Beclin1 overexpression inhibitis proliferation,
invasion and migration of CaSki cervical cancer cells. Asian Pac J
Cancer Prev. 12:1269–1273. 2011.PubMed/NCBI
|
|
12
|
Song C, Zhu S, Wu C and Kang J: Histone
deacetylase (HDAC) 10 suppresses cervical cancer metastasis through
inhibition of matrix metalloproteinase (MMP) 2 and 9 expression. J
Biol Chem. 288:28021–28033. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Siomi H and Siomi MC: Posttranscriptional
regulation of microRNA biogenesis in animals. Mol Cell. 38:323–332.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee JW, Choi CH, Choi JJ, Park YA, Kim SJ,
Hwang SY, Kim WY, Kim TJ, Lee JH, Kim BG, et al: Altered MicroRNA
expression in cervical carcinomas. Clin Cancer Res. 14:2535–2542.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lui WO, Pourmand N, Patterson BK and Fire
A: Patterns of known and novel small RNAs in human cervical cancer.
Cancer Res. 67:6031–6043. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pereira PM, Marques JP, Soares AR, Carreto
L and Santos MA: MicroRNA expression variability in human cervical
tissues. PLoS One. 5:e117802010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Leung CO, Deng W, Ye TM, Ngan HY, Tsao SW,
Cheung AN, Pang RT and Yeung WS: miR-135a leads to cervical cancer
cell transformation through regulation of β-catenin via a SIAH1-
dependent ubiquitin proteosomal pathway. Carcinogenesis.
35:1931–1940. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Long MJ, Wu FX, Li P, Liu M, Li X and Tang
H: MicroRNA-10a targets CHL1 and promotes cell growth, migration
and invasion in human cervical cancer cells. Cancer Lett.
324:186–196. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xie H, Zhao Y, Caramuta S, Larsson C and
Lui WO: miR-205 expression promotes cell proliferation and
migration of human cervical cancer cells. PLoS One. 7:e469902012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yu J, Wang Y, Dong R, Huang X, Ding S and
Qiu H: Circulating microRNA-218 was reduced in cervical cancer and
correlated with tumor invasion. J Cancer Res Clin Oncol.
138:671–674. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tian RQ, Wang XH, Hou LJ, Jia WH, Yang Q,
Li YX, Liu M, Li X and Tang H: MicroRNA-372 is down-regulated and
targets cyclin-dependent kinase 2 (CDK2) and cyclin A1 in human
cervical cancer, which may contribute to tumorigenesis. J Biol
Chem. 286:25556–25563. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Peng RQ, Wan HY, Li HF, Liu M, Li X and
Tang H: MicroRNA-214 suppresses growth and invasiveness of cervical
cancer cells by targeting UDP-N-acetyl-α-D-galactosamine:
polypeptide N-ace tylgalactosaminyltransferase 7. J Biol Chem.
287:14301–14309. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang Y, Li J, Tong L, Zhang J, Zhai A, Xu
K, Wei L and Chu M: The prognostic value of miR-21 and miR-155 in
non-small-cell lung cancer: A meta-analysis. Jpn J Clin Oncol.
43:813–820. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu ZL, Wang H, Liu J and Wang ZX:
MicroRNA-21 (miR-21) expression promotes growth, metastasis, and
chemo- or radiore-sistance in non-small cell lung cancer cells by
targeting PTEN. Mol Cell Biochem. 372:35–45. 2013. View Article : Google Scholar
|
|
25
|
Xiong B, Cheng Y, Ma L and Zhang C: miR-21
regulates biological behavior through the PTEN/PI-3 K/Akt signaling
pathway in human colorectal cancer cells. Int J Oncol. 42:219–228.
2013.
|
|
26
|
Liu S, Fang Y, Shen H, Xu W and Li H:
Berberine sensitizes ovarian cancer cells to cisplatin through
miR-21/PDCD4 axis. Acta Biochim Biophys Sin (Shanghai). 45:756–762.
2013. View Article : Google Scholar
|
|
27
|
Teng Y, Manavalan TT, Hu C, Medjakovic S,
Jungbauer A and Klinge CM: Endocrine disruptors fludioxonil and
fenhexamid stimulate miR-21 expression in breast cancer cells.
Toxicol Sci. 131:71–83. 2013. View Article : Google Scholar :
|
|
28
|
Alder H, Taccioli C, Chen H, Jiang Y,
Smalley KJ, Fadda P, Ozer HG, Huebner K, Farber JL, Croce CM, et
al: Dysregulation of miR-31 and miR-21 induced by zinc deficiency
promotes esophageal cancer. Carcinogenesis. 33:1736–1744. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Deftereos G, Corrie SR, Feng Q, Morihara
J, Stern J, Hawes SE and Kiviat NB: Expression of mir-21 and
mir-143 in cervical specimens ranging from histologically normal
through to invasive cervical cancer. PLoS One. 6:e284232011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
31
|
Chu EC and Tarnawski AS: PTEN regulatory
functions in tumor suppression and cell biology. Med Sci Monit.
10:RA235–RA241. 2004.PubMed/NCBI
|
|
32
|
Vázquez-Ulloa E, Lizano M, Avilés-Salas A,
Alfaro-Moreno E and Contreras-Paredes A: Abnormal distribution of
hDlg and PTEN in premalignant lesions and invasive cervical cancer.
Gynecol Oncol. 122:663–668. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hsieh SM, Maguire DJ, Lintell NA, McCabe M
and Griffiths LR: PTEN and NDUFB8 aberrations in cervical cancer
tissue. Adv Exp Med Biol. 599:31–36. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Meng F, Henson R, Wehbe-Janek H, Ghoshal
K, Jacob ST and Patel T: MicroRNA-21 regulates expression of the
PTEN tumor suppressor gene in human hepatocellular cancer.
Gastroenterology. 133:647–658. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhu S, Si ML, Wu H and Mo YY: MicroRNA-21
targets the tumor suppressor gene tropomyosin 1 (TPM1). J Biol
Chem. 282:14328–14336. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Iorio MV, Ferracin M, Liu CG, Veronese A,
Spizzo R, Sabbioni S, Magri E, Pedriali M, Fabbri M, Campiglio M,
et al: MicroRNA gene expression deregulation in human breast
cancer. Cancer Res. 65:7065–7070. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sempere LF, Christensen M, Silahtaroglu A,
Bak M, Heath CV, Schwartz G, Wells W, Kauppinen S and Cole CN:
Altered MicroRNA expression confined to specific epithelial cell
subpopulations in breast cancer. Cancer Res. 67:11612–11620. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Si ML, Zhu S, Wu H, Lu Z, Wu F and Mo YY:
miR-21-mediated tumor growth. Oncogene. 26:2799–2803. 2007.
View Article : Google Scholar
|
|
39
|
Roldo C, Missiaglia E, Hagan JP, Falconi
M, Capelli P, Bersani S, Calin GA, Volinia S, Liu CG, Scarpa A, et
al: MicroRNA expression abnormalities in pancreatic endocrine and
acinar tumors are associated with distinctive pathologic features
and clinical behavior. J Clin Oncol. 24:4677–4684. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li J, Yen C, Liaw D, Podsypanina K, Bose
S, Wang SI, Puc J, Miliaresis C, Rodgers L, McCombie R, et al:
PTEN, a putative protein tyrosine phosphatase gene mutated in human
brain, breast, and prostate cancer. Science. 275:1943–1947. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sakai A, Thieblemont C, Wellmann A, Jaffe
ES and Raffeld M: PTEN gene alterations in lymphoid neoplasms.
Blood. 92:3410–3415. 1998.PubMed/NCBI
|
|
42
|
Tashiro H, Blazes MS, Wu R, Cho KR, Bose
S, Wang SI, Li J, Parsons R and Ellenson LH: Mutations in PTEN are
frequent in endometrial carcinoma but rare in other common
gynecological malignancies. Cancer Res. 57:3935–3940.
1997.PubMed/NCBI
|
|
43
|
Li DM and Sun H: TEP1, encoded by a
candidate tumor suppressor locus, is a novel protein tyrosine
phosphatase regulated by transforming growth factor beta. Cancer
Res. 57:2124–2129. 1997.PubMed/NCBI
|
|
44
|
Li L, Ernsting BR, Wishart MJ, Lohse DL
and Dixon JE: A family of putative tumor suppressors is
structurally and functionally conserved in humans and yeast. J Biol
Chem. 272:29403–29406. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Maehama T and Dixon JE: The tumor
suppressor, PTEN/MMAC1, dephosphorylates the lipid second
messenger, phosphatidylinositol 3,4,5-trisphosphate. J Biol Chem.
273:13375–13378. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Harima Y, Sawada S, Nagata K, Sougawa M,
Ostapenko V and Ohnishi T: Mutation of the PTEN gene in advanced
cervical cancer correlated with tumor progression and poor outcome
after radiotherapy. Int J Oncol. 18:493–497. 2001.PubMed/NCBI
|
|
47
|
Odar K, Boštjančič E, Gale N, Glavač D and
Zidar N: Differential expression of microRNAs miR-21, miR-31,
miR-203, miR-125a-5p and miR-125b and proteins PTEN and p63 in
verrucous carcinoma of the head and neck. Histopathology.
61:257–265. 2012. View Article : Google Scholar : PubMed/NCBI
|