Introduction
Renal cell carcinoma (RCC) is the most common type
of kidney cancer. RCC accounts for an estimated 3–5% of all
malignant tumours, and this figure increases annually (1). Nearly 50% of patients suffer relapse
and metastasis after curative surgical resection (2). RCC is highly resistant to radiotherapy
and other formal chemotherapies (3). More recently, immunotherapy with
cytokines, such as interleukin-2, interferon-α and interferon-γ,
has become the preferred treatment for later disease phases;
however, the response rate is less than 20% (4). Therefore, there is a need for an
alternative systemic treatment for RCC that overcomes resistance to
current anticancer drugs.
Resistance to multiple categories of chemotherapy
agents, termed multidrug resistance (MDR), is a complex hurdle in
cancer therapy (5). The mechanisms
behind MDR are still under active investigation, yet members of the
ATP binding cassette (ABC) transporter superfamily, which often act
as cellular efflux pumps for a wide range of chemotherapeutic
compounds are implicated (6). Over
49 MDR genes have been described, among which the ABCB1 gene,
encoding the efflux transporter P-glycoprotein (P-gp) or MDR1, is
probably the most well known (7).
The prognostic significance of P-gp as an indicator for failure in
chemotherapy has been demonstrated in a number of clinical studies
(8).
Induction of apoptosis in cancer cells is desirable,
and is a novel cancer therapeutic strategy (9). Apoptosis can be induced through two
major pathways: i) the extrinsic pathway, which is initiated by
specific ligands, such as the Fas-ligand (Fas-L), tumour necrosis
factor (TNF) and the tumour necrosis factor-related
apoptosis-inducing ligand (TRAIL), which activates caspase-8; and
ii) the intrinsic pathway, which is activated by intracellular
signals from the mitochondria to activate caspase-9 (10–12).
Caspase-3, activated by the intrinsic and extrinsic pathways,
induces the cleavage of poly(ADP-ribose) polymerase (PARP), a DNA
repair enzyme, ultimately leading to programmed cell death
(13).
Betulin [lup-20(29)-ene-3β,28diol] (Fig. 1A) is a triterpene compound found in
plants. It is commonly isolated from the bark of various species of
birch tree, including Betula pendula Roth., B. alba
L. and B. pubescens Ehrh. (14), as well as from other plants, such as
Pyrus pyrifolia (15) and
Platycodon grandiflorum (16), by solvent extraction or purified
chromatography. Betulin exhibits numerous biological effects,
including antiviral activities against the human Epstein-Barr virus
(EBV) (17), and antifungal
activities against Epidermophyton floccosum and
Microsporum spp. (18).
Recently, betulin has been shown to exhibit activity against
several cancer cell types, including DLD-1, PC-3, HeLa, SK-HEP-1,
HepG2, A549 and MCF-7 cells, through induction of the mitochondrial
pathway associated with apoptosis (19,20).
However, the cytotoxicity of betulin in RCCs and its mechanisms of
action have not yet been reported.
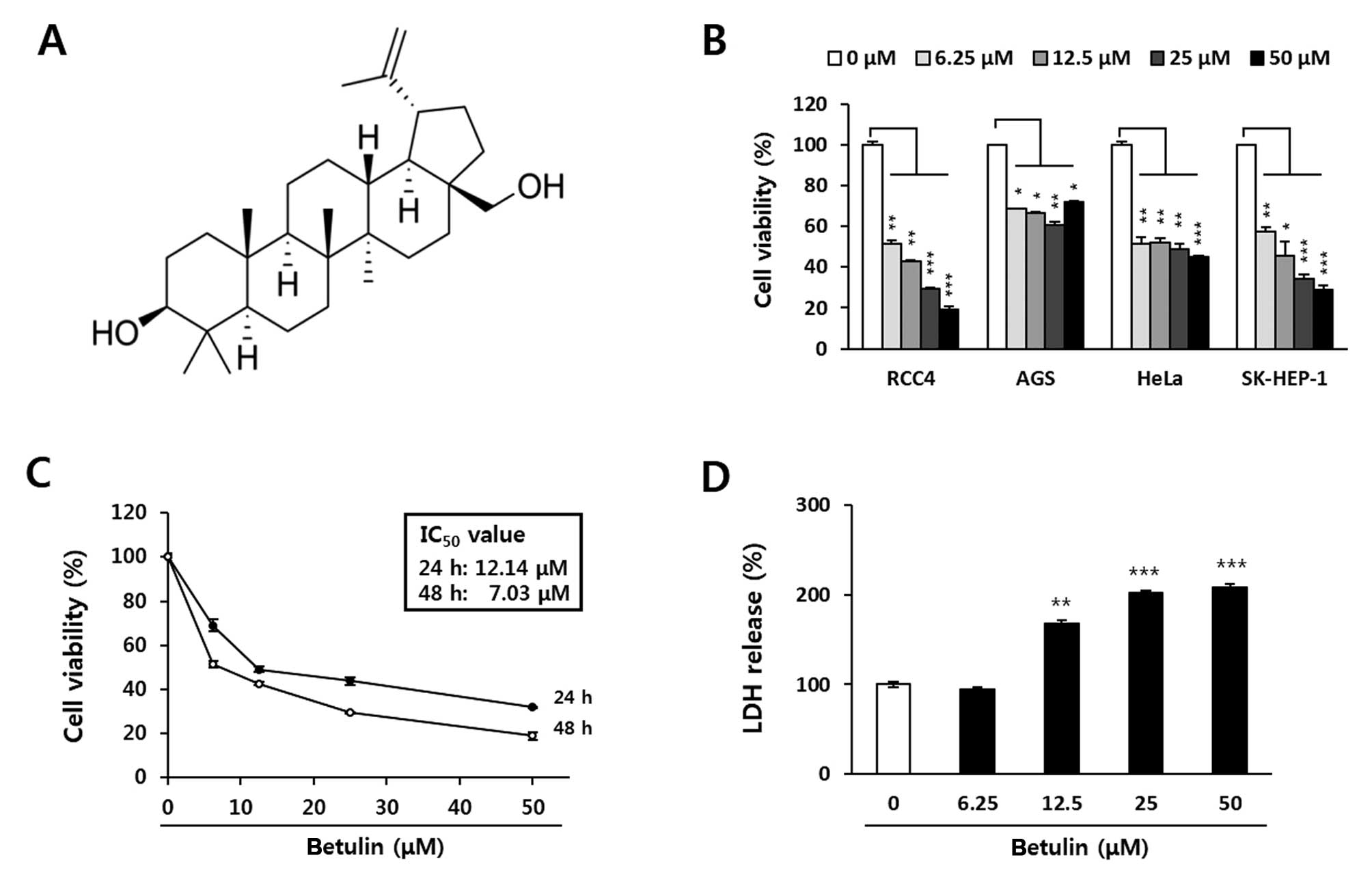 | Figure 1Inhibitory effect of betulin on the
proliferation of human cancer cells. Cell viability was determined
using the MTT assay. (A) Chemical structure of betulin. (B)
Inhibition of viability by betulin in several human cancer cell
lines. Cancer cells were treated with 0, 6.25, 12.5, 25 or 50
μM betulin for 48 h. (C) IC50 values of betulin
in the RCC4 cells. Cells were treated with 0, 1.56, 3.12, 6.25,
12.5, 25 or 50 μM betulin for 24 or 48 h. (D) LDH release in
response to betulin treatment of the RCC4 cells. Cells were treated
with 0, 6.25, 12.5, 25 or 50 μM betulin for 24 h, and the
release of LDH was determined using an LDH detection kit. Data are
presented as the means ± SD of two independent triplicates.
*P<0.05, **P<0.01 and
***P<0.001 indicate statistical significance,
compared to an untreated control. LDH, lactate dehydrogenase. |
The aim of the present study was to investigate
whether betulin inhibits the growth of anticancer drug-resistant
human renal cancer cells, and to define the molecular basis of this
effect. In the present study, we confirmed that betulin exerts its
anticancer effect in human renal cancer cells by modulating key
elements of the apoptotic pathway. We also demonstrated that the
cytotoxicity of betulin towards RCC4 cells is enhanced by
combination with anticancer drugs, through suppression of MDR1
levels.
Materials and methods
Cell culture
Various human cancer cell lines, obtained from the
Korean Cell Line Bank (KCLB; Seoul, Korea) and the American Type
Culture Collection (ATCC; Rockville, MD, USA), were cultured in
Dulbecco’s modified Eagle’s medium (DMEM) and RPMI-1640 (Lonza,
Walkersville, MD, USA) with 10% fetal bovine serum (FBS; HyClone,
Logan, UT, USA). All media contained 100 U/ml penicillin G and 100
μg/ml streptomycin (Gibco, Gaithersburg, MD, USA). Cells
were maintained in an incubator with a humidified atmosphere of 5%
CO2 at 37°C.
Reagents and antibodies
Betulin used in the present study and anticancer
drugs including 5-fluorouracil (5-FU), etoposide and temozolomide,
were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Dimethylsulfoxide (DMSO) and
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT),
hexamethylpararosaniline chloride (crystal violet) and
necrostatin-1 (Nec-1) were also purchased from Sigma-Aldrich.
General caspase inhibitor, Z-VAD-fmk, was purchased from R&D
Systems (Minneapolis, MN, USA). Cytotoxicity detection kit [lactate
dehydrogenase (LDH)] was purchased from Roche Diagnostics
(Mannheim, Germany). RIPA buffer for protein lysis from cells was
obtained from Millipore™ (Billerica, MA, USA). Primary antibodies
against caspase-3, -7, -8 and -9, PARP, NF-κB, phospho-NF-κB,
Bcl-2, Bax, Bid, Bad, PUMA, X-linked inhibitor of apoptosis protein
(XIAP), GAPDH and TBP were purchased from the Cell Signaling
(Danvers, MA, USA). The primary antibodies against active caspase-3
and MDR1 were purchased from Abcam (Cambridge, UK) and GeneTex
(Irvine, CA, USA), respectively.
Cell viability and cytotoxicity
assays
The cell viability assay was determined by a
modified MTT colorimetric assay based on the reduction of
tetrazolium salt as previously described (21). The color intensity was measured as
absorbance at 540 nm with an ELISA microplate reader (Sunrise;
Tecan, Männedorf, Switzerland). To determine cytotoxicity induced
by betulin, LDH released from the cells was evaluated with a
commercial kit according to the manufacturer’s instructions.
Caspase activity assay
The activation of caspases including caspase-3/7, -8
and -9, was respectively measured using a Caspase-Glo 3/7, 8 and 9
assay kit (Promega, Madison, WI, USA) according to the
manufacturer’s instructions. Culture medium was used as a blank
control, and luminescence was measured using an MLX Microtiter
Luminometer (Dynex Technologies, Inc., Chantilly, VA, USA).
Preparation of cytosolic and nuclear
extracts for NF-κB detection
Cytosolic and nuclear fractions were isolated from
the cells using NE-PER nuclear and cytoplasmic extraction reagents
(Thermo Scientific, Rockford, IL, USA) according to the
manufacturer’s instructions. The fractions were stored at −80°C
before use.
Western blot analysis
The cells treated with betulin were cultured for 12
or 24 h and were lysed in RIPA buffer. The lysates were centrifuged
at 14,000 rpm for 20 min at 4°C and its clear supernatant was
collected. Western blot analysis was performed as previously
described (21). The specific
proteins were detected using a SuperSignal West Pico
Chemiluminescent Substrate (Pierce, Rockford, IL, USA) and an
ImageQuant LAS 4000 Mini (GE Healthcare, Piscataway, NJ, USA). Band
intensities were determined using ImageJ software (National
Institute of Health, USA).
Apoptosis array
Human Apoptosis Proteome Profiler™ array (R&D
Systems) was composed of a nitrocellulose membrane duplicate spot
of 35 apoptosis-related proteins. The cells (1×107
cells/ml) were harvested, and 250 μg of total protein was
used for each array. All analyses were performed according to the
manufacturer’s instructions. The membranes were incubated with
horseradish peroxidase-conjugated followed by chemiluminescent
detection reagent.
Statistical analysis
Data are presented as means ± standard deviation
(SD). The statistical significance was analyzed by the two-tailed
Student’s t-test, and a P-value of <0.05 was considered to
indicate a statistically significant result.
Results
Betulin decreases the viability of RCC4
multidrug-resistant human renal carcinoma cells in a time- and
concentration-dependent manner
Four human cancer cell lines [RCC4 (kidney), AGS
(stomach), HeLa (cervical) and SK-Hep-1 (liver)] were treated with
0, 6.25, 12.5, 25 or 50 μM betulin for 48 h, and cell
viability was assessed using the MTT assay. Among the cell lines,
the viability of RCC4 cells was potently inhibited by treatment
with betulin (Fig. 1B). Betulin
inhibited the viability of the RCC4 cells with IC50
values of 23.1 and 14.8 μM after 24 and 48 h, respectively
(Fig. 1C). In addition, LDH release
was observed after a 24-h betulin treatment, and this was
significantly increased at betulin doses exceeding 12.5 μM
(Fig. 1D). Accordingly, 10 and 25
μM betulin were used in further studies to identify the
mechanisms underlying the effect on RCC4 cell viability.
Betulin induces cancer cell death by
activating the caspase-mediated apoptosis pathway in RCC4
cells
To determine whether betulin-induced cytotoxicity is
related to apoptosis, the expression of pro-apoptotic and
anti-apoptotic proteins in RCC4 cells was assessed by western
blotting. The expression levels of proteins associated with
apoptosis were clearly influenced by betulin treatment (Fig. 2A). Pro-caspase-3, -7, -8 and -9 were
cleaved to their active forms after exposure to 10 and 25 μM
betulin for 12 h in RCC4 cells. Betulin also induced truncation of
Bid and cleavage of PARP in a time- and dose-dependent manner. Most
caspases were strongly activated by a 24-h betulin treatment.
Particularly at a 25 μM dose, all caspases were induced
>6-fold, compared to the levels detected in the untreated RCC4
cells. In addition, levels of truncated Bid (t-Bid) and cleaved
PARP increased by 12- and 6-fold, respectively. To confirm that
apoptosis induced by betulin requires caspase activation, a caspase
activity assay was performed in the RCC4 cells (Fig. 2B). Betulin at 10 and 25 μM
doses significantly increased the activities of caspase-3/7, -8 and
-9 at 24 h. Betulin at 10 μM markedly increased the
activities of caspase-3/7, -8 and -9, ~10-, 5- and 7-fold compared
to the non-treated cells, respectively. As shown in Fig. 3C, in the inhibitor study,
pre-incubation with Z-VAD-fmk, a broad-spectrum caspase inhibitor,
for 30 min significantly blocked betulin-induced cell death at 10
μM betulin. Pre-incubation with Nec-1, a necroptosis
inhibitor, blocked betulin-induced cell death; however, the cell
death was not completely blocked at 10 and 25 μM betulin
compared to the effects of Z-VAD-fmk. These data support the
hypothesis that the anticancer effects of betulin in RCC4 cells are
related to apoptotic cell death resulting from activation of
caspases.
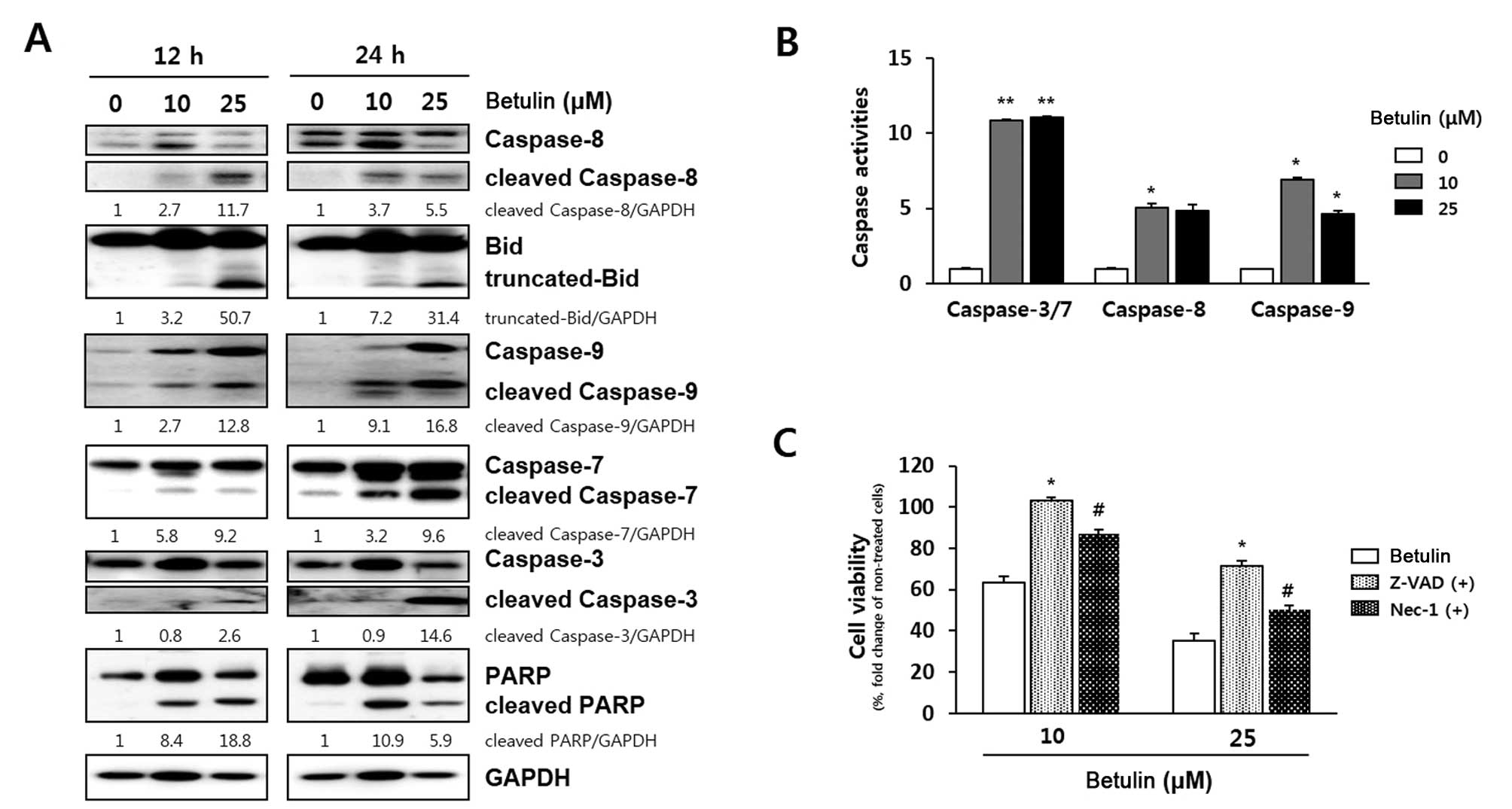 | Figure 2Verification of caspase-dependent
apoptosis induced by betulin in RCC4 cells. (A) Effect of betulin
on caspase protein expression. Cells were exposed to 10 and 25
μM betulin for 12 and 24 h. Band intensity compared to
untreated cells was calculated using ImageJ after neutralization
relative to GAPDH expression. Data represent two independent
experiments. (B) Relative luminescence indicating dose-dependent
activation of caspase-3/7, -8 and -9 induced by a 24-h treatment
with betulin. Data represent two independent triplicates.
*P<0.05 and **P<0.01 indicate
statistical significance, compared to an untreated control. (C)
Assessment of the apoptotic effect of a 24-h betulin treatment
using a caspase inhibitor. Cells were respectively exposed to
Z-VAD-fmk, a general caspase inhibitor and Nec-1, a necroptosis
inhibitor, for 30 min before addition of betulin (10 and 25
μM). After 24 h, cell viability was determined using the MTT
assay. Data are presented as means ± SD. P-values indicate
statistical significance, compared to betulin treatment
(*P<0.05, Z-VAD-fmk; #P<0.05, Nec-1).
Nec-1, necrostatin-1. |
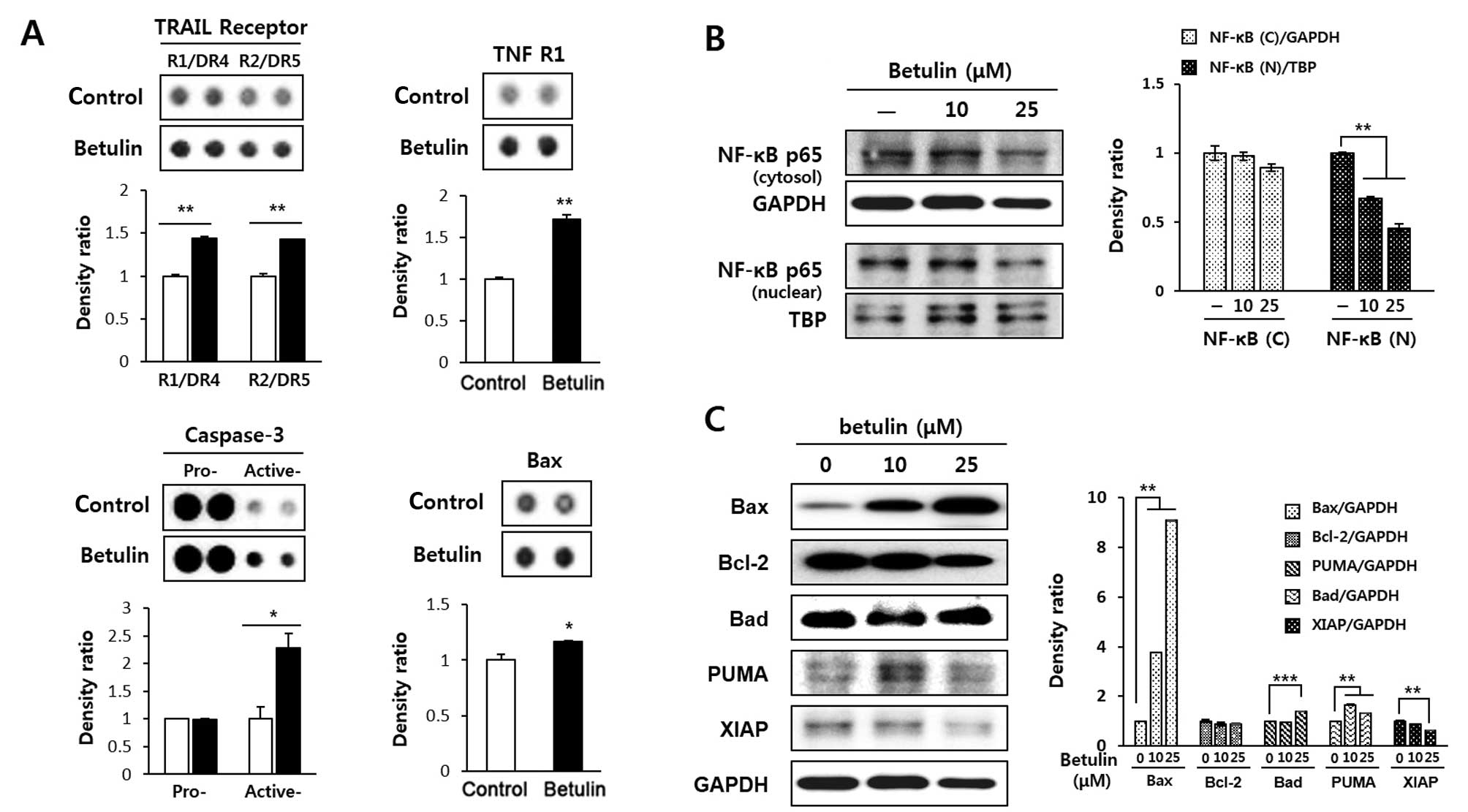 | Figure 3Effect of betulin on apoptosis-related
proteins in RCC4 cells. (A) Analysis of the proteome profiler array
of betulin. Cells were exposed to 10 μM betulin for 24 h and
subjected to a proteome profiler array, according to the
manufacturer’s instructions. The levels of the indicated proteins
(Bax, caspase-3, TRAIL R1/DR4 and R2/DR5, and TNFR1), as shown in
the histograms, were quantified using ImageJ. *P<0.05
and **P<0.01 indicate statistical significance,
compared to an untreated control. (B) Inhibition of betulin induced
nuclear translocation of NF-κB. After a 24-h treatment of RCC4
cells with betulin, the levels of NF-κB (p65) in the cytosol and
nucleus were determined by western blot analysis and quantified
using ImageJ. TBP was used as a nuclear loading control. Data
represent two independent experiments. *P<0.05, and
**P<0.01 indicate statistical significance, compared
to an untreated control. (C) Betulin-induced regulation of Bcl-2
family protein expression. Cells were treated with betulin for 24
h, and the expression levels of Bax, Bcl-2, Bad, PUMA and XIAP were
assessed by western blot analysis. Protein levels were quantified
using ImageJ and normalised relative to GAPDH expression.
**P<0.01 and ***P<0.001 indicate
statistical significance, compared to an untreated control. |
Betulin treatment leads to the expression
of apoptosis-related proteins in RCC4 cells
The betulin-induced expression of apoptosis-related
proteins was calculated using proteome array panels (Fig. 3A). After a 24-h treatment of RCC4
cells with 10 μM betulin, the protein levels related to
apoptosis, such as Bax, active caspase-3, TNF-related
apoptosis-inducing ligand (TRAIL) receptor 1/DR4 and 2/DR5, and
tumour necrosis factor receptor 1 (TNF R1) were upregulated. Bax
and active caspase-3 levels were significantly upregulated, with
active caspase-3 levels increasing >2-fold compared to the
untreated cells. Betulin increased the levels of TRAIL R1/DR4 and
R2/DR5 by 43 and 42%, respectively. Betulin also induced
upregulation of TNFR1 protein expression by 72%. The NF-κB pathway
is closely associated with the expression of TNFR1. Therefore, we
determined the effect of betulin on the nuclear translocation of
p65 in the RCC4 cells. As shown in Fig.
3B, the nuclear translocation of p65 was significantly
suppressed by betulin in a concentration-dependent manner for 24 h.
To confirm that betulin-induced apoptosis is associated with the
mitochondrial-mediated pathway, the levels of Bcl-2 family proteins
including Bcl-2, Bax, Bad and PUMA and XIAP, were estimated by
western blot analysis (Fig. 3C).
The results demonstrated that the betulin increased the expression
of pro-apoptotic proteins. Specifically, a significant increase in
Bax and PUMA expression was observed in response to a 10 μM
dose of betulin for 24 h. Conversely, the expression levels of
anti-apoptotic proteins, such as Bcl-2 and XIAP, were decreased
markedly following treatment with 10 μM betulin.
Betulin enhances the sensitivity of RCC4
cells to anticancer drugs
We hypothesised that betulin sensitises renal cancer
cells to anticancer drugs. To test this, we treated cells with
betulin in combination with anticancer drugs, including 5-FU,
etoposide and temozolomide. 5-FU, etoposide and temozolomide weakly
inhibited the growth of the RCC4 cells after 24 h by 13.54, 24.38
and 2.43%, respectively, compared to the control condition, whereas
the addition of 25 μM betulin resulted in inhibition of cell
growth by up to 56.12% (Fig. 4A).
In addition, by co-treating cells with etoposide in the presence of
betulin, the cytotoxicity of the RCC4 cells was significantly
increased. The cytotoxicity induced by co-treatment with 5
μM etoposide and 5 μM betulin was almost equal to
that of 10 μM betulin alone (Fig. 4B). To confirm the synergistic effect
of betulin and etoposide on cancer cell death, the levels of
cleaved PARP and MDR1 were assessed in RCC4 cells using western
blot analysis. As shown in Fig. 4C,
co-treatment with 10 μM etoposide and 10 μM betulin
increased the level of cleaved PARP >2- and 3-fold, compared to
betulin and etoposide treatment alone, respectively. In the case of
MDR1, elevated expression of MDR1 was observed in the RCC4 cells,
and this was decreased ~2-fold by co-treatment with 10 μM
betulin and 10 μM etoposide. However, treatment with betulin
or etoposide alone did not affect MDR1 expression.
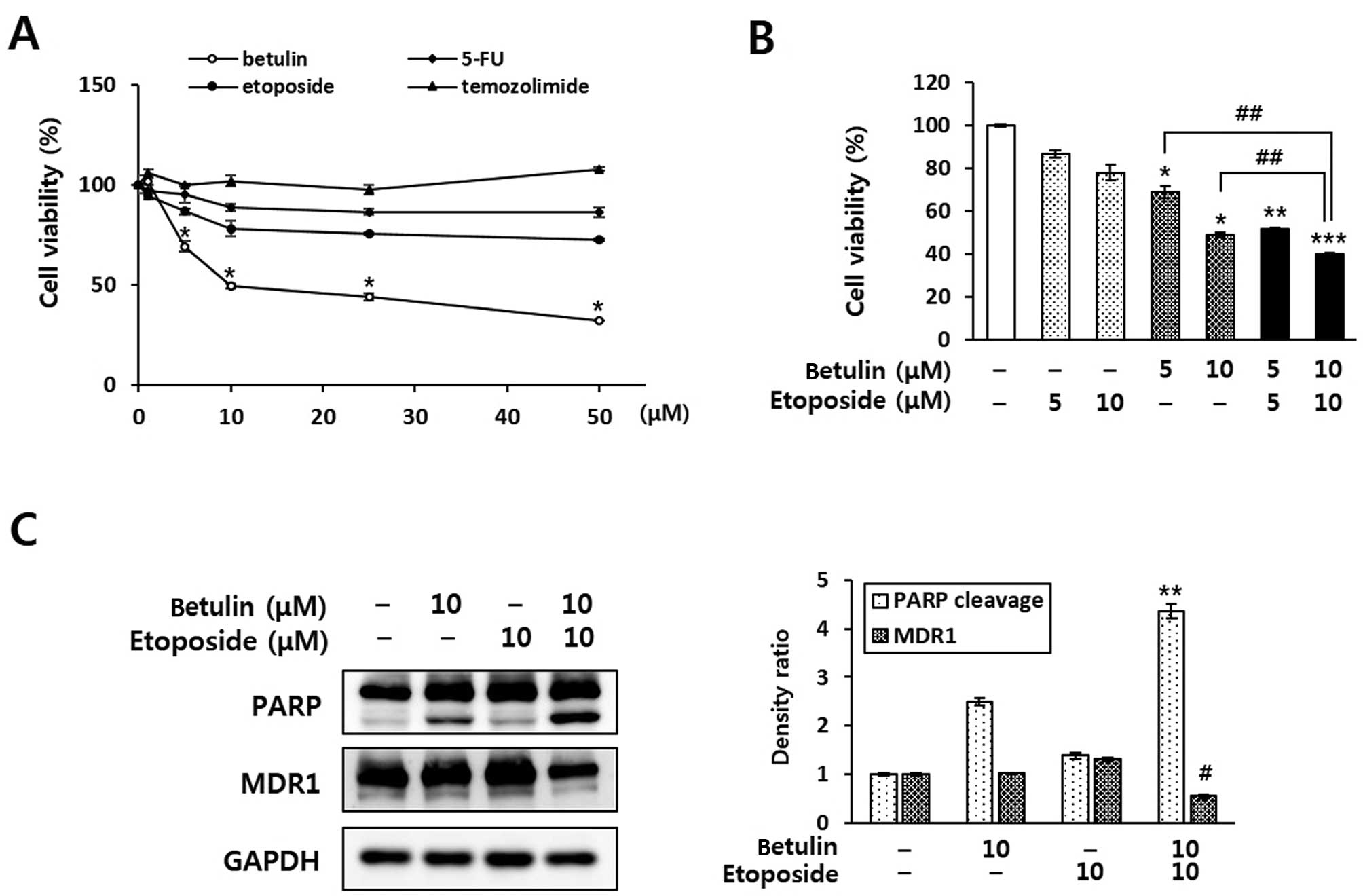 | Figure 4The combined effect of the
co-treatment of betulin and anticancer agents in RCC4 cells. (A)
Enhancement of anticancer agent-induced cell death by betulin in
the RCC4 cells. Cells were treated with various concentrations (1,
5, 10, 25 and 50 μM) of betulin or anticancer drugs (5-FU,
etoposide and temozolimide). After 24 h, cell viability was
determined using an MTT assay. Data are presented as means ± SD.
*P<0.05 indicates statistical significance, compared
to the untreated control. (B) Evaluation of the combined effect of
betulin and etoposide on the cell growth of RCC4 cells. After a
24-h treatment of RCC4 cells with betulin (5 and 10 μM)
and/or etoposide (5 and 10 μM), an MTT assay was performed.
*P<0.05, **P<0.01 and
***P<0.01 indicate statistical significance, compared
to an untreated control. ##P<0.01 indicates
statistical significance, compared to betulin alone. (C)
Enhancement of the anticancer effect by combined treatment with
betulin and etoposide. Cells were treated with betulin (10
μM) and/or etoposide (10 μM) for 24 h, and the
expression levels of PARP and MDR1 were determined by western blot
analysis. Protein levels were normalised relative to GAPDH
expression and quantified using ImageJ. **P<0.01
indicates statistical significance of PARP cleavage, compared to
etoposide treatment alone and #P<0.01 indicates
statistical significance of MDR1 expression compared to etoposide
treatment alone. |
Discussion
The induction of apoptosis in cancer cells by
natural compounds is a key goal for cancer prevention and therapy.
Betulin is a pharmacologically active triterpene that is naturally
occurring in birch bark. It is used in its extracted form or as a
base for chemical modification to compounds such as betulinic acid
(22). Previous studies have
reported that betulin has an anticancer effect in human lung,
cervical and liver cancer cells (23). Betulin has also been shown to
protect hepatoma cells from damage by ethanol-induced liver
stellate or cadmium-induced apoptosis, via a mechanism involving
inhibition of reactive oxygen species (ROS) production or apoptosis
induction (24,25). In HepG2 cells, betulin was found to
induce apoptotic cell death through the intrinsic apoptotic pathway
that includes activation of caspase-9 and -3 (23). Caspase is a key mediator of
apoptosis that is activated via endoplasmic reticulum stress,
extracellular stimuli and mitochondrial damage (26). The activation of caspase-8 is
initiated by the stimulation of cell-surface death receptors, such
as Fas, TNF and TRAIL. This leads to activation of caspase-3 or -7,
or modulation of the mitochondrial pathway via truncation of Bid
(27). Conversely, caspase-9 is
activated by cytochrome c released from the mitochondria,
which in turn, leads to the activation of effector caspases, such
as caspase-3, -6 and -7. Activated caspase-3 induces cleavage of
PARP, a DNA repair enzyme, and ultimately, apoptosis (28). The present study revealed that
betulin treatment inhibited the viability of human renal cancer
cells and increased the activity of caspases, including caspase-3,
-7, -8 and -9. Furthermore, our data indicated that betulin
regulates mitochondrial signalling pathways associated with
apoptosis, including the Bcl-2 family and XIAP pathways, which
suggests that sensitisation of cancer cells by betulin to apoptosis
is caspase-9-dependent. Notably, in the present study, betulin
significantly increased the levels of TRAIL R1/DR4 and R2/DR5, and
TNFR1 in an apoptosis array. TRAIL induces apoptosis in a variety
of transformed or tumour cells, but not in normal cells; however,
many cancer cells are resistant to TRAIL (27). Binding of TRAIL to the TRAIL R1/DR4
and R2/DR5 receptors results in trimerisation to form a
death-inducing signalling complex (DISC). DISC recruits the adapter
protein FADD and caspase-8, leading to activation of intact
caspases and cleavage of PARP (29). Therefore, increased expression of
the TRAIL death receptors could play an important role in the
sensitisation of cancer cells to TRAIL-associated apoptosis
following proteasome inhibition. TNFR1 induces the activation of
NF-κB and plays a role in caspase-8 activation by controlling the
receptor interacting protein-1 (RIP1) (30). NF-κB is an apoptosis regulator, and
its inhibition results in apoptosis in a variety of cell types
found to be initially resistant to TNF-α-induced apoptosis
(31,32). Furthermore, in cancer therapy,
suppression of NF-κB translocation can increase the effectiveness
of cancer therapy (33). Hence, our
data demonstrated that stimulation of TRAIL receptors and TNFR1 by
betulin results in activation of caspase-8 and inhibition of NF-κB
signalling. Therefore, we suggest that betulin treatment leads to
induction of apoptosis through both the intrinsic and extrinsic
pathways in RCC4 cells.
Additionally, we demonstrated that betulin inhibits
cell viability by inducing caspase-mediated apoptosis in RCC4 human
renal carcinoma cells shown to be resistant to anticancer drugs,
such as 5-FU, temozolomide and etoposide. Among them, etoposide is
a topoisomerase inhibitor that disrupts the repair of DNA damage.
DNA-damaging drugs are highly effective anticancer drugs that are
in current clinical use; however, side-effects, including vomiting,
have been reported in several studies (34). RCCs are the most common malignant
tumours found in the adult kidney, and unlike other malignancies,
they are generally resistant to conventional therapies. Therapeutic
resistance in RCCs occurs as a result of genetic changes that
confer tumourigenic potential and survival advantages during
chemotherapy (35,36). Most of the established
resistance-modifying agents (RMA) are excessively toxic at the
required doses. The search for P-gp inhibitors among libraries of
natural products may be more promising, as many natural products
and phytotherapeutics are appreciated for their low incidence and
severity of side-effects and good tolerability (5). Previous studies have revealed that the
first intron of the human MDR1 gene binds to NF-κB complexes
(37). In the present study,
betulin alone or in combination with etoposide, significantly
increased the protein level of cleaved PARP, while etoposide alone
did not affect cleavage of PARP. Furthermore, the combination of
betulin and etoposide significantly suppressed MDR1 expression,
whereas either drug alone did not. Betulin inhibited the
translocation of NF-κB, yet failed to affect the expression of
MDR1. Thus, we suggest that betulin and anticancer drugs
synergistically enhance toxicity in multidrug-resistant human renal
carcinoma cells. Further research is necessary to identify the
sensitising molecular mechanism and in vivo therapeutic
benefits of combination therapy involving betulin and anticancer
drugs.
In conclusion, we demonstrated that betulin induced
apoptosis in RCC4 cells via activation of death-inducing receptors,
including TRAIL receptors and TNFR1, and regulation of
anti-apoptotic or pro-apoptotic proteins. Furthermore, the
combination of betulin and an anticancer agent synergistically
inhibited PARP expression and upregulated MDR1 expression in RCC4
cells. Therefore, we suggest that betulin is a promising candidate
chemopreventive and chemotherapeutic agent for multidrug-resistant
human renal cancer.
Acknowledgments
The present study was supported by a grant (K14050)
awarded to the Korean Institute of Oriental Medicine by the
Ministry of Education, Science and Technology (MEST), Republic of
Korea.
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Morais C, Pat B, Gobe G, Johnson DW and
Healy H: Pyrrolidine dithiocarbamate exerts anti-proliferative and
pro-apoptotic effects in renal cell carcinoma cell lines. Nephrol
Dial Transplant. 21:3377–3388. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hong MH1, Kim HS, Kim C, Ahn JR, Chon HJ,
Shin SJ, Ahn JB, Chung HC and Rha SY: Treatment outcomes of
sunitinib treatment in advanced renal cell carcinoma patients: A
single cancer center experience in Korea. Cancer Res Treat.
41:67–72. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
McDermott DF: Immunotherapy of metastatic
renal cell carcinoma. Cancer. 115(Suppl 10): S2298–S2305. 2009.
View Article : Google Scholar
|
|
5
|
Zhu L, Zhao L, Wang H, Wang Y, Pan D, Yao
J, Li Z, Wu G and Guo Q: Oroxylin A reverses
P-glycoprotein-mediated multidrug resistance of MCF7/ADR cells by
G2/M arrest. Toxicol Lett. 219:107–115. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Benabbou N, Mirshahi P, Cadillon M, Soria
J, Therwath A and Mirshahi M: Hospicells promote upregulation of
the ATP-binding cassette genes by insulin-like growth factor-I via
the JAK2/STAT3 signaling pathway in an ovarian cancer cell line.
Int J Oncol. 43:685–694. 2013.PubMed/NCBI
|
|
7
|
Harris AL and Hochhauser D: Mechanisms of
multidrug resistance in cancer treatment. Acta Oncol. 31:205–213.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kimura Y, Morita SY, Matsuo M and Ueda K:
Mechanism of multidrug recognition by MDR1/ABCB1. Cancer Sci.
98:1303–1310. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kopnin BP: Targets of oncogenes and tumor
suppressors: Key for understanding basic mechanisms of
carcinogenesis. Biochemistry. Biokhimiia. 65:2–27. 2000.PubMed/NCBI
|
|
10
|
Lowe SW and Lin AW: Apoptosis in cancer.
Carcinogenesis. 21:485–495. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Desagher S and Martinou JC: Mitochondria
as the central control point of apoptosis. Trends Cell Biol.
10:369–377. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jin Z and El-Deiry WS: Overview of cell
death signaling pathways. Cancer Biol Ther. 4:139–163. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
D’Amours D, Desnoyers S, D’Silva I and
Poirier GG: Poly(ADP-ribosyl)ation reactions in the regulation of
nuclear functions. Biochem J. 342:249–268. 1999. View Article : Google Scholar
|
|
14
|
Flekhter OB, Medvedeva NI, Tolstikov GA,
Galin FZ, Iunusov MS, Huong NT, Le VT, Svinova OV, Boreko EI, Titov
LP, et al: Synthesis of olean-18(19)-ene derivatives from betulin.
Bioorg Khim. 35:253–259. 2009.In Russian. PubMed/NCBI
|
|
15
|
Yoo JH and Yang KS: Lupane triterpenoids
from Pyrus pyrifolia. Nat Prod Sci. 18:13–15. 2012.
|
|
16
|
Hwang CR, Oh SH, Kim HY, Lee SH, Hwang IG,
Shin YS, Lee JS and Jeong HS: Chemical composition and antioxidant
activity of Deoduk (Codonopsis lanceolate) and Doragi (Platycodon
grandiforum) according to temperature. J Korean Soc Food Sci Nutr.
40:798–803. 2011. View Article : Google Scholar
|
|
17
|
Akihisa T, Takamine Y, Yoshizumi K, Tokuda
H, Kimura Y, Ukiya M, Nakahara T, Yokochi T, Ichiishi E and Nishino
H: Microbial transformations of two lupane-type triterpenes and
anti-tumor-promoting effects of the transformation products. J Nat
Prod. 65:278–282. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Machado KE, Cechinel Filho V, Cruz RC,
Meyre-Silva C and Cruz AB: Antifungal activity of Eugenia
umbelliflora against dermatophytes. Nat Prod Commun. 4:1181–1184.
2009.PubMed/NCBI
|
|
19
|
Pyo JS, Roh SH, Kim DK, Lee JG, Lee YY,
Hong SS, Kwon SW and Park JH: Anti-cancer effect of Betulin on a
human lung cancer cell line: A pharmacoproteomic approach using 2 D
SDS PAGE coupled with nano-HPLC tandem mass spectrometry. Planta
Med. 75:127–131. 2009. View Article : Google Scholar
|
|
20
|
Li Y, He K, Huang Y, Zheng D, Gao C, Cui L
and Jin YH: Betulin induces mitochondrial cytochrome c release
associated apoptosis in human cancer cells. Mol Carcinog.
49:630–640. 2010.PubMed/NCBI
|
|
21
|
Yim NH, Cho WK, Lee JH, Jung YP, Yang HJ
and Ma JY: HRT, herbal formula, induces G2/M cell cycle
arrest and apoptosis via suppressing Akt signaling pathway in human
colon cancer cells. eCAM. 2012:871–893. 2012. View Article : Google Scholar
|
|
22
|
Kvasnica M, Sarek J, Klinotova E, Dzubak P
and Hajduch M: Synthesis of phthalates of betulinic acid and
betulin with cytotoxic activity. Bioorg Med Chem. 13:3447–3454.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li Y, Shen JT, Gao C, Li Q and Jin Y:
Caspase-9 activation, critical for betulin-induced apoptosis of
human hepatoma cells. Chem Res Chin Univ. 26:792–797. 2010.
View Article : Google Scholar
|
|
24
|
Szuster-Ciesielska A, Plewka K, Daniluk J
and Kandefer-Szerszeń M: Betulin and betulinic acid attenuate
ethanol-induced liver stellate cell activation by inhibiting
reactive oxygen species (ROS), cytokine (TNF-α, TGF-β) production
and by influencing intracellular signaling. Toxicology.
280:152–163. 2011. View Article : Google Scholar
|
|
25
|
Oh SH, Choi JE and Lim SC: Protection of
betulin against cadmium-induced apoptosis in hepatoma cells.
Toxicology. 220:1–12. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nuñez G, Benedict MA, Hu Y and Inohara N:
Caspases: The proteases of the apoptotic pathway. Oncogene.
17:3237–3245. 1998. View Article : Google Scholar
|
|
27
|
Wu GS: TRAIL as a target in anti-cancer
therapy. Cancer Lett. 285:1–5. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Green DR: Apoptotic pathways: Paper wraps
stone blunts scissors. Cell. 102:1–4. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Humphreys RC and Halpern W: Trail
receptors: Targets for cancer therapy. Adv Exp Med Biol.
615:127–158. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wajant H: Increasing complexity in TNFR1
signaling. FEBS J. 278:8612011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Beg AA and Baltimore D: An essential role
for NF-kappaB in preventing TNF-alpha-induced cell death. Science.
274:782–784. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Van Antwerp DJ, Martin SJ, Kafri T, Green
DR and Verma IM: Suppression of TNF-alpha-induced apoptosis by
NF-kappaB. Science. 274:787–789. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Van Waes C: Nuclear factor-kappaB in
development, prevention, and therapy of cancer. Clin Cancer Res.
13:1076–1082. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Moriarty TJ, Dupuis S and Autexier C:
Rapid upregulation of telomerase activity in human leukemia HL-60
cells treated with clinical doses of the DNA-damaging drug
etoposide. Leukemia. 16:1112–1120. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Thompson Coon JS, Liu Z, Hoyle M, Rogers
G, Green C, Moxham T, Welch K and Stein K: Sunitinib and
bevacizumab for first-line treatment of metastatic renal cell
carcinoma: A systematic review and indirect comparison of clinical
effectiveness. Br J Cancer. 101:238–243. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Porta C, Paglino C, Imarisio I, Ganini C
and Pedrazzoli P: Immunological effects of multikinase inhibitors
for kidney cancer: A clue for integration with cellular therapies?
J Cancer. 2:333–338. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kaewpiboon C, Srisuttee R, Malilas W, Moon
J, Kaowinn S, Cho IR, Johnston RN, Assavalapsakul W and Chung YH:
Extract of Bryophyllum laetivirens reverses etoposide resistance in
human lung A549 cancer cells by downregulation of NF-κB. Oncol Rep.
31:161–168. 2014.
|














