Introduction
Osteosarcoma is the most common malignant bone
tumor, with high morbidity and lethality in young adults and
adolescents. Although intensive chemotherapy, radiotherapy, as well
as aggressive surgical procedures, have been combined and applied
for the treatment of patients with osteosarcoma, only 60% achieve
5-year survival. What is striking is that ~80% of these patients
have clinically detectable metastatic disease at the time of
diagnosis (1,2). This disappointing current situation
warrants the urgent need to identify the underlying molecular
signaling mechanisms involved in osteosarcoma carcinogenesis and
metastasis.
Visfatin, initially found in visceral fat, also
known as nicotinamide phosphoribosyltransferase (Nampt) and
pre-B-cell colony-enhancing factor (PBEF), plays an important role
in a variety of metabolic and stress responses, such as NAD
biosynthesis, and exhibits proliferative, anti-apoptotic,
pro-inflammatory and pro-angiogenic properties (3). Recently, the effects of visfatin on
carcinogenesis and its use as a chemotherapeutic target have
attracted our attention. Visfatin was found to be overexpressed in
colorectal cancer (4), and its
over-expression predicted a poor prognosis and decreased response
to doxorubicin therapy in breast cancer (5). These data indicate that the expression
and activity of visfatin may become novel therapeutic strategy
targets for cancer patients. However, to date, only a few studies
exist that concern the effect of visfatin on the clinical outcome
of osteosarcoma patients, and even less studies have researched the
potential mechanisms (6).
Epithelial-mesenchymal transition (EMT), whose
function was primarily found in the process of embryogenesis
(7), has been characterized by
promoting downregulation of intercellular cohesion of cancer cells,
elevated rate of cancer cell migration and invasion, and increased
resistance to apoptosis even under chemotherapeutics (8). The most important hallmarks of EMT are
decreased expression of the epithelial molecule E-cadherin and
increased expression of mesenchymal marker N-cadherin (9). Previous studies have demonstrated that
EMT significantly induces cancer cells to disseminate from the
tumor mass leading to distant metastasis.
However, there is still no evidence to date on the
direct effect of visfatin on osteosarcoma cell migration and
invasion. Thus, in view of the above, in the present study, we
aimed to evaluate whether osteosarcoma cell migration and invasion
are influenced by visfatin through promotion of EMT. Finally, the
mechanism of EMT promotion in osteosarcoma cells was also
investigated, thus, offering a potential improvement in
osteosarcoma treatment management.
Materials and methods
Reagents
Recombinant human visfatin and BAY11-7082, a
selective inhibitor of NF-κB, were both purchased from
Sigma-Aldrich (St. Louis, MO, USA). Rabbit monoclonal antibody to
E-cadherin was purchased from Abcam (Cambridge, UK). Rabbit
polyclonal antibodies to N-cadherin, Snail-1 and GAPDH were also
purchased from Abcam. Rabbit anti-mouse antibody for NF-κB (P65)
was purchased from Cell Signaling Technology Inc. (Danvers, MA,
USA). Snail-1 siRNA was purchased from Santa Cruz Biotechnology,
Inc. (Dallas, TX, USA).
Cell culture
The U2OS osteosarcoma cell line was purchased from
the American Type Culture Collection (ATCC; Manassas, VA, USA).
Dulbecco’s modified Eagle’s medium (DMEM; HyClone, Logan, UT, USA)
supplemented with 10% fetal bovine serum (FBS; HyClone) was used to
culture the U2OS cells. The U2OS cells were grown in a 37°C
incubator with 5% CO2. To determine the effect of
visfatin on the expression of EMT markers, the U2OS cells were
maintained in DMEM supplemented with 1% FBS for 12 h and
subsequently cultured in the absence or presence of visfatin or
other reagents for the specified times.
Migration assay
Cell migration was studied using a scratch wound
healing assay. The U2OS cells were cultured in 6-well plates
(1×106/well; Corning, NY, USA) and grown in a confluent
monolayer. Straight scratches of the same width were made in
monolayers of the U2OS cells with a pipette tip. After incubation
with the reagents for a specified time, photo images were captured
to measure the wound healing under a microscope.
Invision assay
Cell invasion was studied using a Matrigel-coated
Transwell assay. Modified Boyden chambers with 8-μm pore
filter inserts were coated with Matrigel (50 μg/well; BD
Biosciences, San Jose, CA, USA). The U2OS cells were cultured in
24-well plates, and the upper chamber contained cells in DMEM plus
1% FBS, while the lower chamber contained DMEM plus 10% FBS. The
cells (1×105 cells/well) were re-suspended in the upper
chamber at 37°C and 5% CO2. After a 24-h incubation, the
cells that had invaded onto the lower surface of the
Matrigel-coated membrane were fixed with methanol for 30 min and
stained with hexamethylpararosaniline, while the cells that
remained on the upper surface were wiped away.
Western blot analysis
After stimulation with visfatin and other reagents,
the U2OS cells were collected and the protein was extracted in
ice-cold RIPA buffer (Beyotime Institute of Biotechnology, Nantong,
Jiangsu, China) with 1 mM phenylmethyl-sulfonyl fluoride (PMSF;
Beyotime Institute of Biotechnology) for 30 min. Concentration of
the extracted protein was measured using the BCA protein assay kit
(Beyotime Institute of Biotechnology). Cell lysates were
electrophoresed on a 10% SDS polyacrylamide gel and transferred
onto nitrocellulose (NC) membranes (Millipore, Merck KGaA,
Darmstadt, Germany). The blots were blocked and incubated with
primary antibodies overnight followed by incubation with the
secondary antibodies for 2 h at room temperature. Finally, the
blots were visualized with an electrochemiluminescence (ECL)
detection system (Millipore).
Real-time quantitative PCR (qPCR)
analysis
Total RNA was isolated and extracted using TRIzol
reagent (Invitrogen) according to the manufacturer’s directions
from the cells that were cultured following the specified
experimental programs. The concentration of total RNA was
quantified by spectrophotometry. The RNA samples were
reverse-transcribed into cDNA using M-MLV (Moloney murine leukemia
virus) reverse transcriptase system (Fermentas, Shenzhen, China)
(10). Total cDNA was amplified and
detected using LightCycler FastStart DNA Master SYBR-Green I
(Takara Biotechnology, Dalian, China). 18s was chosen as the
reference gene, and the primer sequences for real-time PCR analyses
were: E-cadherin forward primer, 5′-ACCAGAATAAAGACCAAGTGACCA-3′ and
reverse primer, 5′-AGCAAGAGCAGCAGAATCAG AAT-3′ (11); N-cadherin forward primer,
5′-CACTGCTCA GGACCCAGAT-3′ and reverse primer, 5′-TAAGCCGAGT
GATGGTCC-3′ (12); Snail forward
primer, 5′-TTGGATAC AGCTGCTTTGAG-3′ and reverse primer, 5′-ATTGCATA
GTTAGTCACACCTC-3′ (11); 18S
forward primer, 5′-CTT AGTTGGTGGAGCGATTTG-3′ and reverse primer,
5′-GCT GAACGCCACTTGTCC-3′ (13).
Statistical analysis
Data in the present study were evaluated with
Predictive Analytic Software (PASW) Statistics 18.0 (SPSS, Inc.,
Chicago, IL, USA). The normally distributed data were analyzed by
one-way ANOVA, while the nonparametric variables were analyzed by
the Mann-Whitney U test. Statistical significance was confirmed at
a probability value (P) <0.05. Each assay was performed in
triplicate.
Results
Effects of visfatin on expression of EMT
markers in the osteosarcoma cells
It is well known that cancer metastasis involves
multiple steps, among which the acquisition of invasiveness through
EMT has been well established and illustrated. Meanwhile, the loss
of the epithelial marker E-cadherin and gain of mesenchymal marker
N-cadherin have been both confirmed as the most important hallmarks
of EMT (9). We thus examined the
effects of visfatin on expression of EMT markers in the
osteosarcoma cells with western blot analysis and real-time PCR.
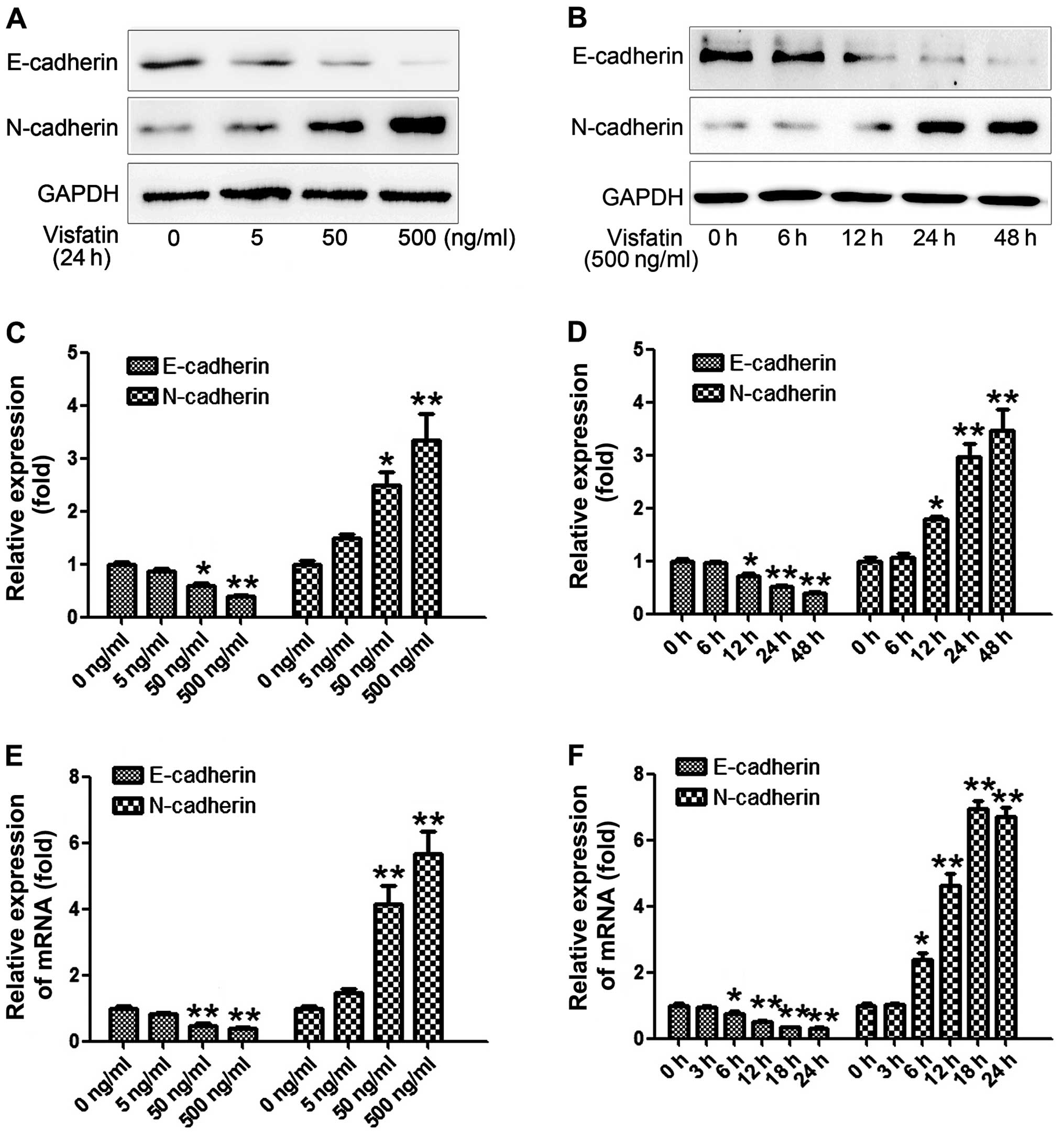
The osteosarcoma cells were cultured with various concentrations of
visfatin (0, 5, 50 and 500 ng/ml) respectively, for 24 h. As shown
in Fig. 1A and C, visfatin
concentration-dependently decreased expression of the epithelial
marker E-cadherin and enhanced the expression of mesenchymal marker
N-cadherin in the osteosarcoma cells. Then, real-time PCR was
performed and the relative expression of the mRNA levels of
E-cadherin and N-cadherin were assessed, and the results
demonstrated the same tendency as the expression levels at the
protein level (Fig. 1E).
Subsequently, visfatin of 500 ng/ml was added to the osteosarcoma
cells and cultured for various times (0, 6, 12, 24 and 48 h). As
shown in Fig. 1B and D, visfatin
time-dependently decreased the expression of E-cadherin and
enhanced expression of N-cadherin in the osteosarcoma cells,
reaching peak activity at 48 h. Finally, we also examined the mRNA
expression of E-cadherin and N-cadherin after various culture times
(0, 3, 6, 12, 18 and 24 h). Similar results as the protein level
were obtained. The results above indicated that visfatin altered
the expression of EMT markers in a concentration- and
time-dependent manner.
Visfatin affects EMT via regulation of
the expression of Snail
It has been revealed that EMT is affected by a
variety of regulatory networks and is mainly triggered by a series
of transcription factors, such as Snail-1, Slug, Twist, Zeb1 and
SIP1. Among these factors, Snail-1 has been the most extensively
studied and has been found to play a pivotal role in the process
(14). Thus, we investigated the
effects of visfatin on the expression of Snail-1 and its subsequent
effects on EMT.
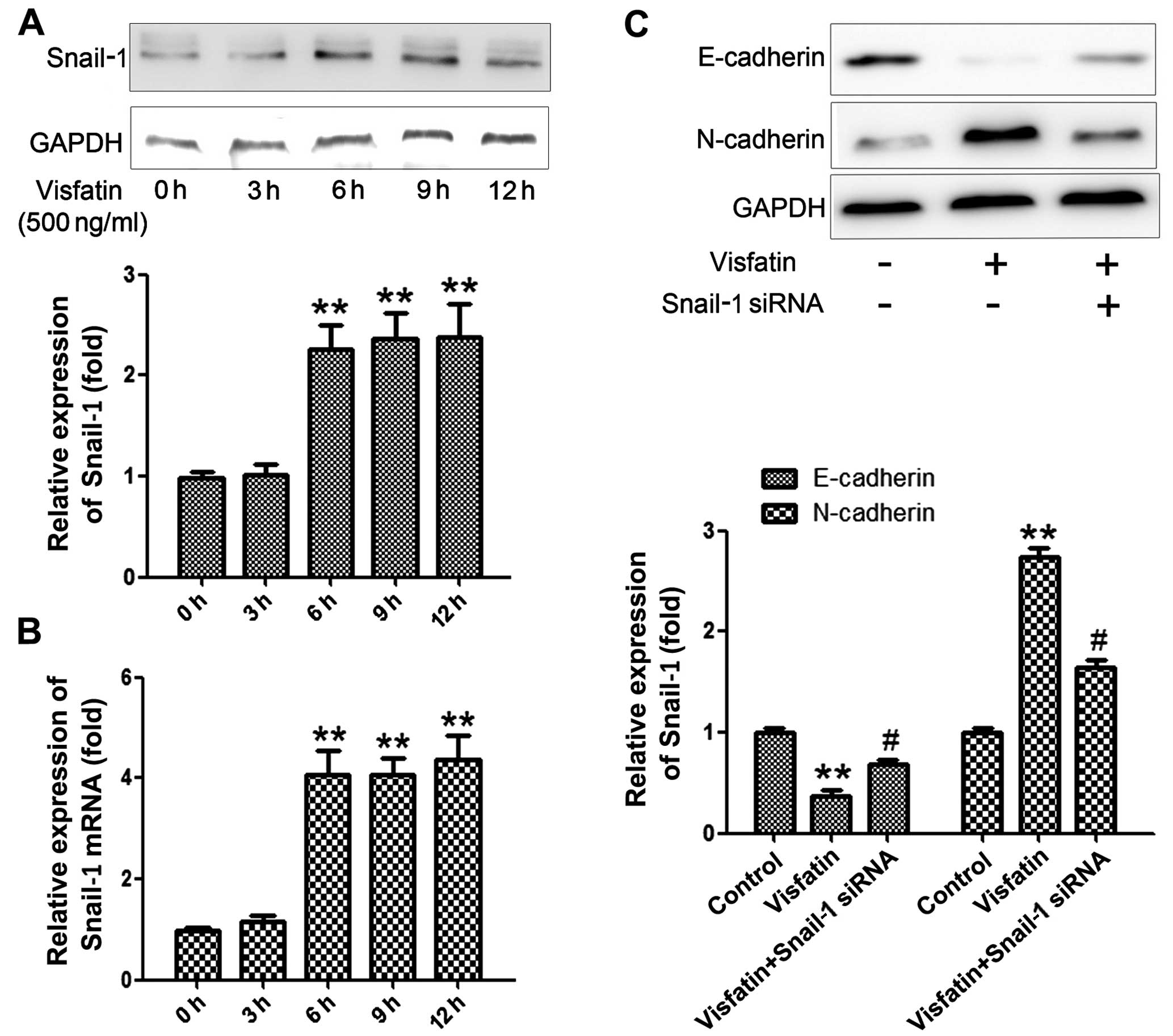
Visfatin of 500 ng/ml was selected to stimulate
osteosarcoma cells for various times (0, 3, 6, 9 and 12 h). As
shown in Fig. 2A, visfatin
significantly promoted the expression of Snail-1 along with the
increase in the stimulation time, reaching a peak at 12 h.
Subsequently, real-time PCR was performed to examine the relative
expression of Snail-1, and the results were similar with those at
the protein level (Fig. 2B).
Subsequently, in order to ascertain the compulsory role of Snail-1
in promoting EMT, the cells were pre-treated with siRNA of Snail-1
for 2 h before the stimulation of visfatin. As shown in Fig. 2C, pretreatment with the siRNA of
Snail-1 significantly abrogated the promotive effect of visfatin on
E-cadherin and the inhibitory effect on N-cadherin. The results
above confirmed that Snail-1 is a key regulator of EMT and visfatin
affects EMT via regulation of the expression of Snail-1.
Visfatin induces NF-κB nuclear
translocation
It has been reported that NF-κB is a key
transcriptional signaling molecule that mediates the expression of
many downstream genes. In previous studies, the expression of NF-κB
and its nuclear translocation were proven to be involved in the
invasion of osteosarcoma cells (15,16).
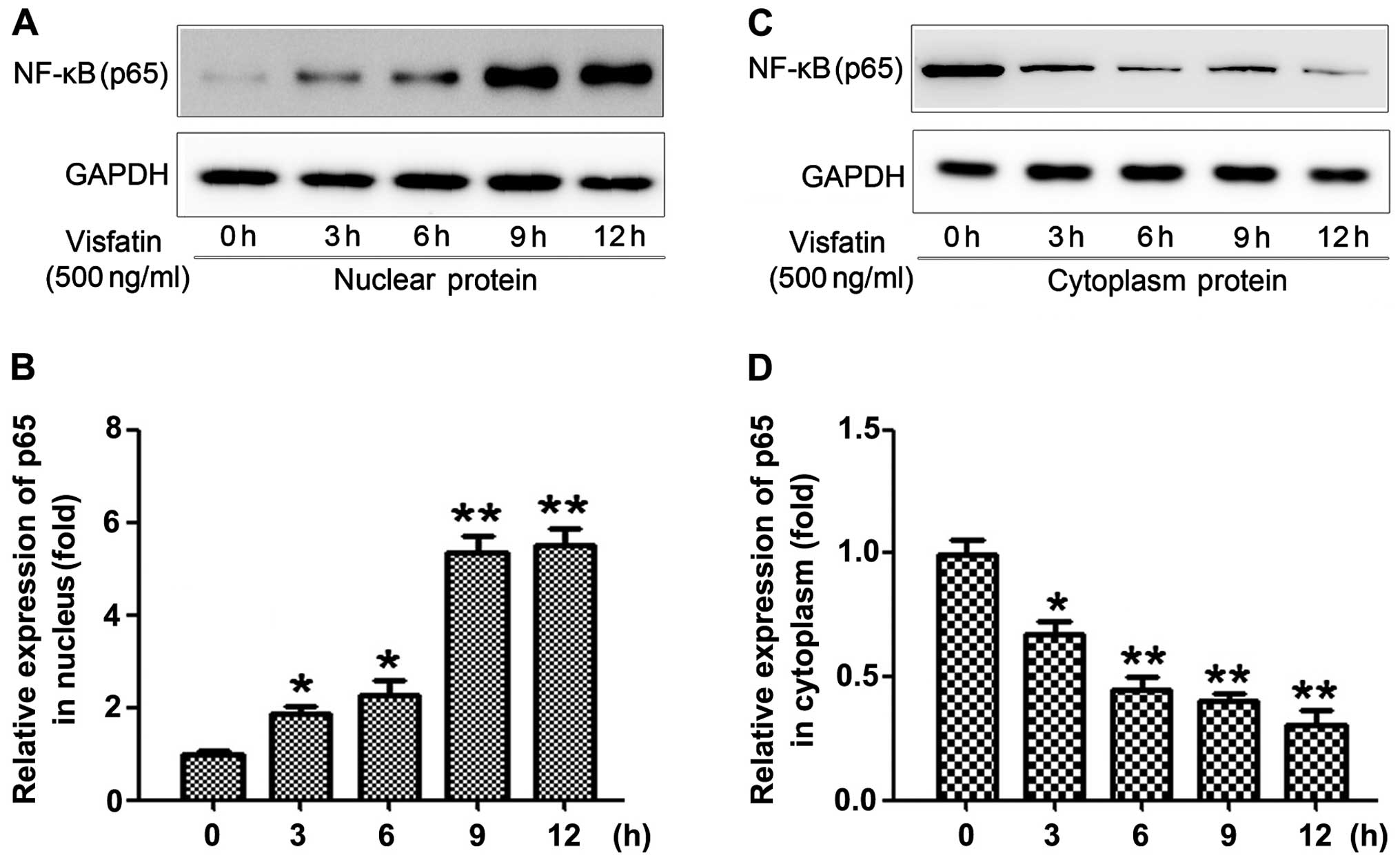
Thus, in the present study, in order to determine the effects of
visfatin on the expression and nuclear translocation of NF-κB, we
respectively extracted and measured the phosphorylation levels of
p65 in the nucleus and in the cytoplasm after various stimulation
times (0, 3, 6, 9 and 12 h). As shown in Fig. 3A and B, the results demonstrated
that visfatin significantly increased the expression of p65 in the
nucleus and simultaneously decrease its expression in the cytoplasm
(Fig. 3C and D). In addition, we
observed that only a 3-h stimulation by visfatin could
significantly transfer p65 from the cytoplasm to the nucleus.
NF-κB mediates visfatin-induced
expression of EMT markers
In order to further determine the molecular
mechanisms underlying visfatin-induced promotion and suppression of
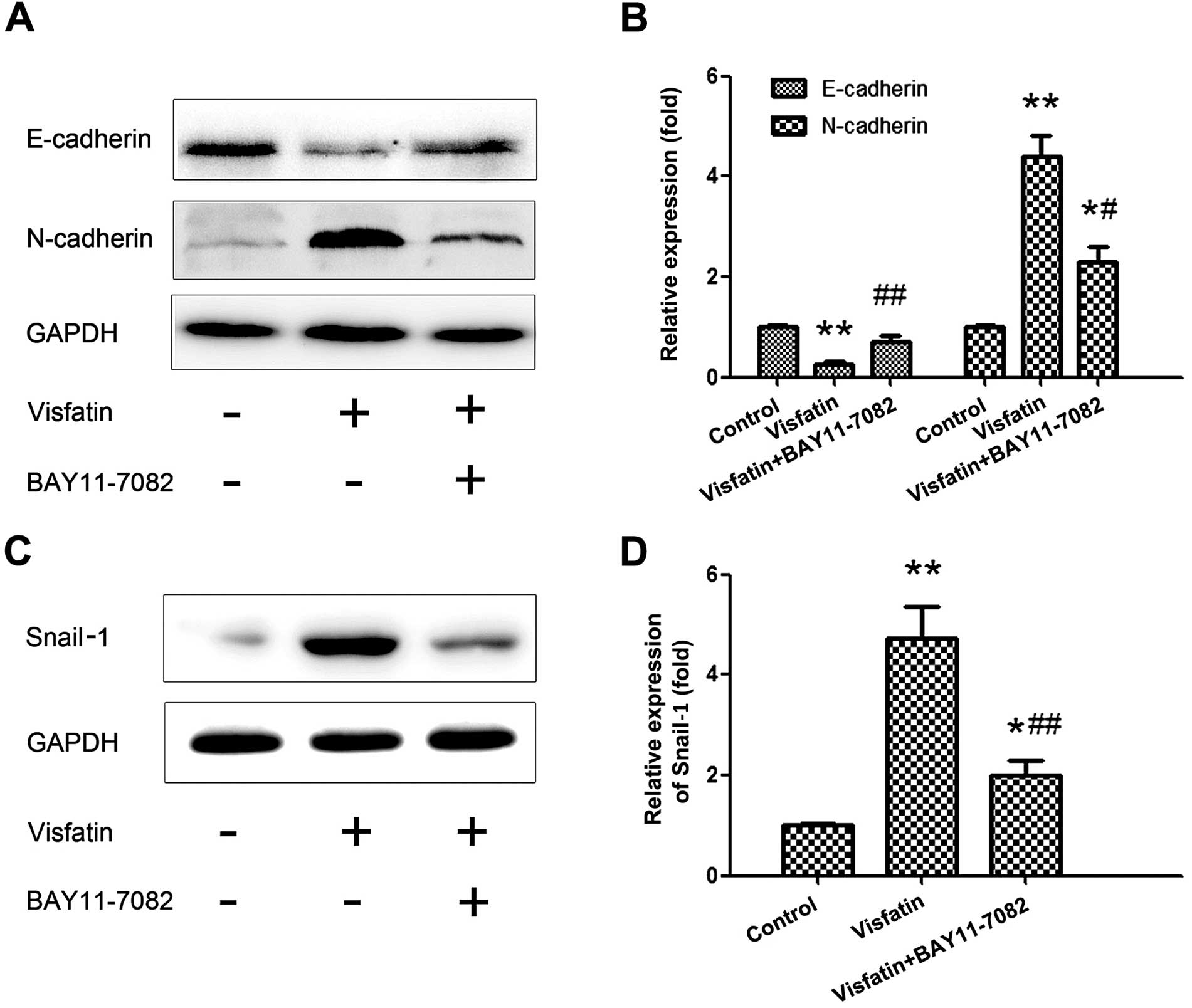
EMT markers, we chose 500 ng/ml of visfatin to stimulate the
osteosarcoma cells for 24 h and pre-treated the cells with
BAY11-7082 (20 μM) for 2 h. As shown in Fig. 4A, pretreatment with the NF-κB
inhibitor BAY11-7082 significantly abrogated the effects of
visfatin on E-cadherin and N-cadherin. Subsequently, the expression
of Snail-1 was examined following the same treatments, and a
similar tendency was obtained. These results indicate that NF-κB is
a key signaling molecule that mediates the expression of Snail-1
and EMT.
Visfatin promotes the migration and
invasion of osteosarcoma cells mainly through the Snail-1 and NF-κB
signaling pathways
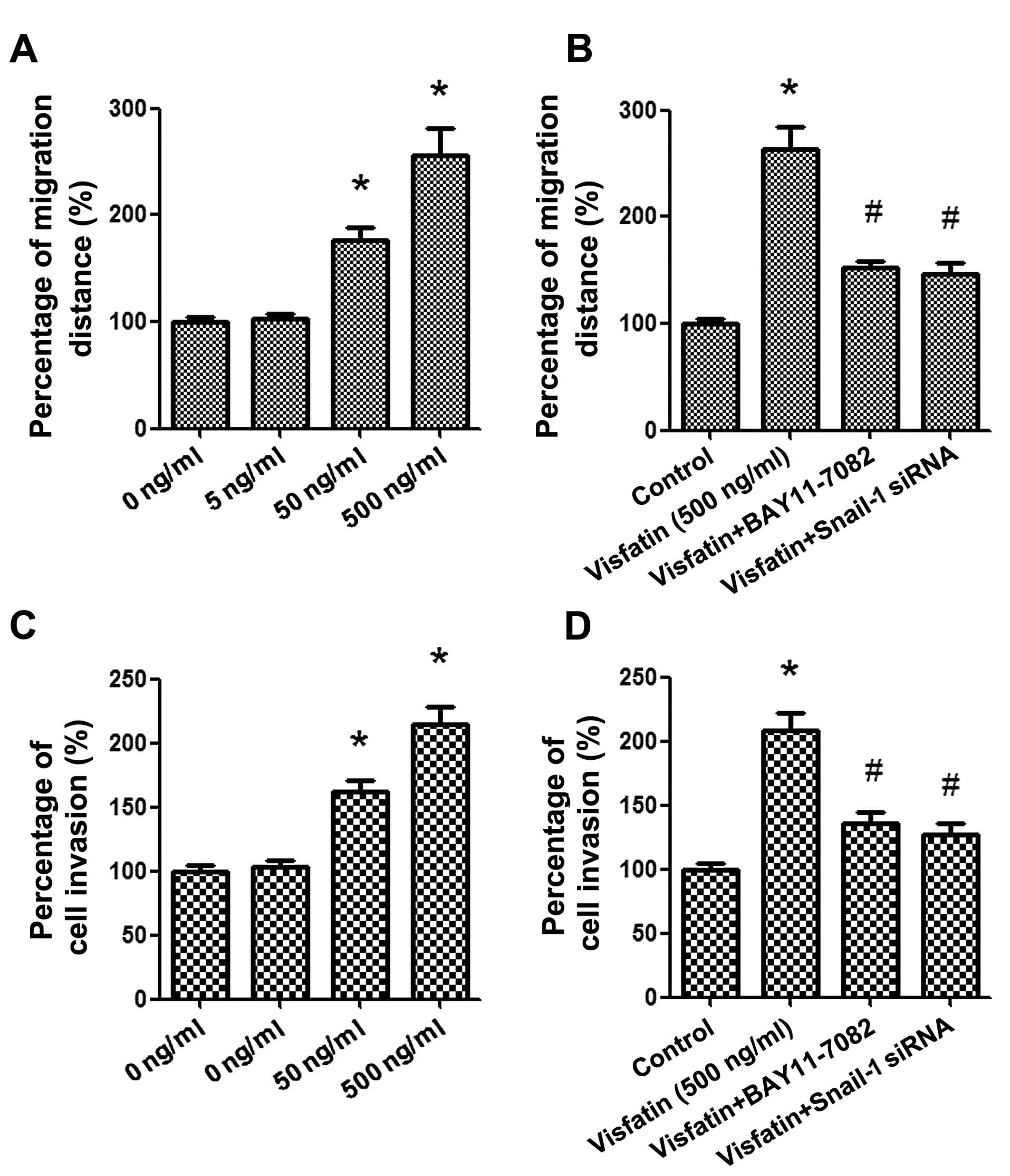
Wound healing and Transwell assays were respectively
applied to evaluate the migration and invasion of osteosar-coma
cells. Osteosarcoma cells were treated with various concentrations
of visfatin (0, 5, 50 and 500 ng/ml) for 24 h. As shown in Fig. 5A, the results of the wound healing
assay demonstrated that healing over the scratch was increased
gradually in a concentration-dependent manner after treatment with
visfatin. Moreover, as shown in Fig.
5C, a Transwell assay exhibited that the number of invading
cells transferred from the upper surfaces to the lower surfaces
were also prominently promoted by visfatin in a similar trend.
Next, in order to further ascertain the signaling pathway
underlying the effects of visfatin on cancer cell migration and
invasion, Snail-1 siRNA and BAY11-7082 (20 μM) were
repectively used to pre-treat the cells for 2 h. As shown in
Fig. 5B and D, blockage of the
Snail-1 and NF-κB signaling pathways markedly suppressed the
migration and invasion of osteosarcoma cells, which suggests that
visfatin promotes the migration and invasion of osteosarcoma cells
mainly through the Snail-1 and NF-κB signaling pathways.
Discussion
Osteosarcoma, arising from primitive transformed
cells of mesenchymal origin, mostly occurs in the long bones of
children and adolescents, and is the most common malignant tumor of
bone (17). The underlying
mechanisms involved in the occurrence and development of
osteosarcoma are so complex that the exact etiology remains unclear
to date. Despite the fact that the mortality rate of osteosarcoma
has declined in the past few years, ~30% of osteosarcoma patients
succumb to lung metastases (18,19).
As shown in a previous studies, lung is the most common and primary
site of metastasis for osteosarcoma (20). Thus, it is urgent to further
investigate the potential mechanisms underlying the pathogenesis of
osteosarcoma and to search for new factors that affect the
migration and invasion of osteosarcoma cells, as well as to explore
novel therapeutic approaches and treatment strategies for
osteosarcoma patients.
Nampt/PBEF/visfatin has been considered to belong to
the dimeric class of type II phosphoribosyltransferases because of
its crystal structure (21,22). This enzyme consists of two different
forms: an intracellular form, named iNampt and an extracellular
form, named eNampt (6). Since
little attention has been paid to visfatin, the biological
functions and the underlying mechanisms of visfatin remain
incompletely understood and its function has been mainly explored
in regards to endothelial cells and vascular disorders (6,23).
However, along with the findings of its roles in carcinogenesis,
recently visfatin has received more attention. As shown in previous
studies, visfatin expression is markedly increased in various types
of tumors, such as glioblastoma (24), malignant astrocytomas (24), breast (4,25) and
prostate cancer (26). In addition,
it has been demonstrated that the expression level of visfatin is
gradually elevated with the progressive stage of gastric (27) and colorectal cancers (28). Thus, visfatin is regarded as a
biomarker for many types of cancer. However, there is still no
evidence showing the specific effects of visfatin on osteosarcoma
to date, and thus, the present study was designed to elucidate the
role of visfatin in osteosarcoma, as well as the underlying
mechanisms.
Tumor-associated EMT is known as one of the most
important contributors promoting osteosarcoma progression and
metastasis (17). According to
investigations, EMT is associated with elevated grades of
incursion, higher tumor recurrence, poorer prognosis and decreased
patient survival rates (29).
During the process of EMT, the conversion of polar-ized epithelial
cells acquiring a mesenchymal phenotype alters cell-cell and
cell-extracellular matrix (ECM) interactions and makes cell
motility more unconstrained through the ECM, which is intensively
associated with tumor metastasis. The most important marks of EMT
are the loss of epithelial marker E-cadherin and the acquisition of
mesenchymal marker N-cadherin. Thus, in order to ascertain the
effect of visfatin on EMT, the expression levels of E-cadherin and
N-cadherin were detected, respectively, at the levels of protein
and mRNA following treatment with visfatin. The data (Fig. 1) indicated a significant role of
visfatin; vitsfatin treatment concentration- and time-dependently
decreased and increased the expression of E-cadherin and
N-cadherin, respectively. The results intensively suggest that
visfatin is an inducer of EMT in osteosarcoma cells.
Although the molecular mechanisms associated with
loss of E-cadherin expression and increase of N-cadherin expression
are still not clear, the Snail-1 superfamily, particularly Snail-1,
have been considered as the master regulators. A number of
experiments have been performed to indicate that Snail-1 is the
transcriptional repressor of E-cadherin and transcriptional inducer
of N-cadherin. It has been reported that Snail-1 is overexpressed
in osteosarcoma (30), and its
overexpression inhibits the action of 1,25-dihydroxyvitamin
D3 [1,25(OH)-D3] which was thought to
suppress the proliferation, migration and invasion of cancers
(31,32), so that Snail-1 exerts its promoting
effect on osteosarcoma progression (30). In the present study, we detected the
protein and mRNA expression of Snail-1 after various times of
stimulation with visfatin, and the results showed that a 6-h
stimulation significantly increased Snail-1 expression. When we
blocked Snail-1 with its siRNA, the effects of visfatin on
E-cadherin and N-cadherin were signifi-cantly reversed, which
indicated that visfatin-induced EMT was mediated by Snail-1.
It has been firmly established that NF-κB exerts a
pivotal role in the regulation of tumor metastasis (33). A previous study proved that the
pulmonary metastasis of osteosar-coma is reduced by inhibition of
NF-κB (33). In a study of Huber
et al (34), it was found
that inhibition of NF-κB prevented EMT, while activation of this
pathway promoted EMT, and inhibition of NF-κB caused a reverse in
EMT, suggesting that NF-κB is essential for EMT. Maier et al
(35) and Chen et al
(36) also found that NF-κB
promoted the migration and invasion of carcinoma cells via its
induction of EMT. In the present study, we aimed to ascertain
whether NF-κB was involved in visfatin-induced EMT and the
metastasis of osteosarcoma. First, the expression of the p65
subunit of NF-κB was detected respectively in the nucleus and the
cytoplasm, and the increase in the nucleus and the decrease in the
cytoplasm indicated the occurrence of NF-κB nuclear translocation
under the stimulation of visfatin. When the NF-κB pathway was
blocked by its inhibitor, the effects of visfatin on the expression
levels of E-cadherin and N-cadherin were entirely reversed, as well
as the expression of Snail-1. This suggests that NF-κB is the
mediator of visfatin-induced EMT.
In conclusion, the results presented in this study
elucidated the visfatin/NF-κB/Snail-1/EMT pathway as a novel
mechanism by which visfatin promotes the aggressive behavior of
osteosarcoma cells. In addition, the data above indicate the
importance of visfatin as a potential therapeutic target. Thus,
inhibition of visfatin may provide a new anti-osteosarcoma
strategy.
Acknowledgments
The present study was supported by the Science
Foundation of Shandong Province (no. ZR2014HP005).
References
|
1
|
Link MP, Goorin AM, Miser AW, Green AA,
Pratt CB, Belasco JB, Pritchard J, Malpas JS, Baker AR, Kirkpatrick
JA, et al: The effect of adjuvant chemotherapy on relapse-free
survival in patients with osteosarcoma of the extremity. N Engl J
Med. 314:1600–1606. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Luu HH, Zhou L, Haydon RC, Deyrup AT,
Montag AG, Huo D, Heck R, Heizmann CW, Peabody TD, Simon MA, et al:
Increased expression of S100A6 is associated with decreased
metastasis and inhibition of cell migration and anchorage
independent growth in human osteosarcoma. Cancer Lett. 229:135–148.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dalamaga M: Nicotinamide
phosphoribosyl-transferase/visfatin: A missing link between
overweight/obesity and postmenopausal breast cancer? Potential
preventive and therapeutic perspectives and challenges. Med
Hypotheses. 79:617–621. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hufton SE, Moerkerk PT, Brandwijk R, de
Bruïne AP, Arends JW and Hoogenboom HR: A profile of differentially
expressed genes in primary colorectal cancer using suppression
subtractive hybridization. FEBS Lett. 463:77–82. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Folgueira MA, Carraro DM, Brentani H,
Patrão DF, Barbosa EM, Netto MM, Caldeira JR, Katayama ML, Soares
FA, Oliveira CT, et al: Gene expression profile associated with
response to doxorubicin-based therapy in breast cancer. Clin Cancer
Res. 11:7434–7443. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bi TQ and Che XM: Nampt/PBEF/visfatin and
cancer. Cancer Biol Ther. 10:119–125. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hay ED: The mesenchymal cell, its role in
the embryo, and the remarkable signaling mechanisms that create it.
Dev Dyn. 233:706–720. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Haslehurst AM, Koti M, Dharsee M, Nuin P,
Evans K, Geraci J, Childs T, Chen J, Li J, Weberpals J, et al: EMT
transcription factors snail and slug directly contribute to
cisplatin resistance in ovarian cancer. BMC Cancer. 12:912012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rosanò L, Cianfrocca R, Spinella F, Di
Castro V, Nicotra MR, Lucidi A, Ferrandina G, Natali PG and Bagnato
A: Acquisition of chemoresistance and EMT phenotype is linked with
activation of the endothelin A receptor pathway in ovarian
carcinoma cells. Clin Cancer Res. 17:2350–2360. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang XC, Chen JQ and Li B: Salvianolic
acid A suppresses CCL-20 expression in TNF-α-treated macrophages
and ApoE-deficient mice. J Cardiovasc Pharmacol. 64:318–325. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bae KM, Parker NN, Dai Y, Vieweg J and
Siemann DW: E-cadherin plasticity in prostate cancer stem cell
invasion. Am J Cancer Res. 1:71–84. 2011.PubMed/NCBI
|
|
12
|
Pon YL, Auersperg N and Wong AS:
Gonadotropins regulate N-cadherin-mediated human ovarian surface
epithelial cell survival at both post-translational and
transcriptional levels through a cyclic AMP/protein kinase A
pathway. J Biol Chem. 280:15438–15448. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li B, Dong Z, Liu H, Xia YF, Liu XM, Luo
BB, Wang WK, Li B, Gao F, Zhang C, et al: Serum amyloid A
stimulates lipoprotein-associated phospholipase A2 expression in
vitro and in vivo. Atherosclerosis. 228:370–379. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Peinado H, Olmeda D and Cano A: Snail, Zeb
and bHLH factors in tumour progression: An alliance against the
epithelial phenotype? Nat Rev Cancer. 7:415–428. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao Z, Wu MS, Zou C, Tang Q, Lu J, Liu D,
Wu Y, Yin J, Xie X, Shen J, et al: Downregulation of MCT1 inhibits
tumor growth, metastasis and enhances chemotherapeutic efficacy in
osteosar-coma through regulation of the NF-κB pathway. Cancer Lett.
342:150–158. 2014. View Article : Google Scholar
|
|
16
|
Li Y, Zhang ZN, Zhao HM, Tong ZC, Yang J,
Wang H and Liang XJ: Matrine inhibits the invasive properties of
human osteosarcoma cells by downregulating the ERK-NF-κB pathway.
Anticancer Drugs. 25:1035–1043. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang G, Yuan J and Li K: EMT transcription
factors: Implication in osteosarcoma. Med Oncol. 30:6972013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
PosthumaDeBoer J, Witlox MA, Kaspers GJ
and van Royen BJ: Molecular alterations as target for therapy in
metastatic osteosarcoma: A review of literature. Clin Exp
Metastasis. 28:493–503. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Krishnan K, Khanna C and Helman LJ: The
biology of metastases in pediatric sarcomas. Cancer J. 11:306–313.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Harting MT and Blakely ML: Management of
osteosarcoma pulmonary metastases. Semin Pediatr Surg. 15:25–29.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Khan JA, Tao X and Tong L: Molecular basis
for the inhibition of human NMPRTase, a novel target for anticancer
agents. Nat Struct Mol Biol. 13:582–588. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang T, Zhang X, Bheda P, Revollo JR, Imai
S and Wolberger C: Structure of Nampt/PBEF/visfatin, a mammalian
NAD+ biosynthetic enzyme. Nat Struct Mol Biol.
13:661–662. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kim SR, Bae SK, Choi KS, Park SY, Jun HO,
Lee JY, Jang HO, Yun I, Yoon KH, Kim YJ, et al: Visfatin promotes
angiogenesis by activation of extracellular signal-regulated kinase
1/2. Biochem Biophys Res Commun. 357:150–156. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Reddy PS, Umesh S, Thota B, Tandon A,
Pandey P, Hegde AS, Balasubramaniam A, Chandramouli BA, Santosh V,
Rao MR, et al: PBEF1/NAmPRTase/Visfatin: A potential malignant
astrocytoma/glioblastoma serum marker with prognostic value. Cancer
Biol Ther. 7:663–668. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Van Beijnum JR, Moerkerk PT, Gerbers AJ,
De Bruïne AP, Arends JW, Hoogenboom HR and Hufton SE: Target
validation for genomics using peptide-specific phage antibodies: A
study of five gene products overexpressed in colorectal cancer. Int
J Cancer. 101:118–127. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Patel ST, Mistry T, Brown JE, Digby JE,
Adya R, Desai KM and Randeva HS: A novel role for the adipokine
visfatin/pre-B cell colony-enhancing factor 1 in prostate
carcinogenesis. Peptides. 31:51–57. 2010. View Article : Google Scholar
|
|
27
|
Nakajima TE, Yamada Y, Hamano T, Furuta K,
Gotoda T, Katai H, Kato K, Hamaguchi T and Shimada Y: Adipocytokine
levels in gastric cancer patients: Resistin and visfatin as
biomarkers of gastric cancer. J Gastroenterol. 44:685–690. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nakajima TE, Yamada Y, Hamano T, Furuta K,
Matsuda T, Fujita S, Kato K, Hamaguchi T and Shimada Y:
Adipocytokines as new promising markers of colorectal tumors:
Adiponectin for colorectal adenoma, and resistin and visfatin for
colorectal cancer. Cancer Sci. 101:1286–1291. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cichon MA and Radisky DC: ROS-induced
epithelial-mesen-chymal transition in mammary epithelial cells is
mediated by NF-κB-dependent activation of Snail. Oncotarget.
5:2827–2838. 2014.PubMed/NCBI
|
|
30
|
Yang H, Zhang Y, Zhou Z, Jiang X and Shen
A: Snail-1 regulates VDR signaling and inhibits
1,25(OH)-D3 action in osteosarcoma. Eur J Pharmacol.
670:341–346. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Larriba MJ, Ordóñez-Morán P, Chicote I,
Martín-Fernández G, Puig I, Muñoz A and Pálmer HG: Vitamin D
receptor deficiency enhances Wnt/β-catenin signaling and tumor
burden in colon cancer. PLoS One. 6:e235242011. View Article : Google Scholar
|
|
32
|
Stubbins RE, Hakeem A and Núñez NP: Using
components of the vitamin D pathway to prevent and treat colon
cancer. Nutr Rev. 70:721–729. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nishimura A, Akeda K, Matsubara T,
Kusuzaki K, Matsumine A, Masuda K, Gemba T, Uchida A and Sudo A:
Transfection of NF-κB decoy oligodeoxynucleotide suppresses
pulmonary metastasis by murine osteosarcoma. Cancer Gene Ther.
18:250–259. 2011. View Article : Google Scholar
|
|
34
|
Huber MA, Azoitei N, Baumann B, Grünert S,
Sommer A, Pehamberger H, Kraut N, Beug H and Wirth T: NF-kappaB is
essential for epithelial-mesenchymal transition and metastasis in a
model of breast cancer progression. J Clin Invest. 114:569–581.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Maier HJ, Schmidt-Strassburger U, Huber
MA, Wiedemann EM, Beug H and Wirth T: NF-kappaB promotes
epithelial-mesenchymal transition, migration and invasion of
pancreatic carcinoma cells. Cancer Lett. 295:214–228. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cheng ZX, Sun B, Wang SJ, Gao Y, Zhang YM,
Zhou HX, Jia G, Wang YW, Kong R, Pan SH, et al: Nuclear
factor-κB-dependent epithelial to mesenchymal transition induced by
HIF-1α activation in pancreatic cancer cells under hypoxic
conditions. PLoS One. 6:e237522011. View Article : Google Scholar
|