Introduction
The immune system functions as a host defensive
mechanism protecting against tumor development. Evidence suggests
that cell autoantigens, which are displayed by antigen-presenting
cells (APC) on MHC class II molecules, stimulate CD4+ T
helper cells in local tumor-draining lymph node (TDLN) (1). Although numerous antigen-specific T
lymphocytes proliferate in draining lymph nodes, advanced-stage
cancer cells are able to invade TDLN from primary lesions. Thus, it
is important to investigate the relationship between lymph node
metastasis and the underlying mechanisms of immune tolerance in
TDLN. However, few studies have examined changes in immune cells in
lymph nodes of human clinical samples.
T lymphocytes are divided into three subsets: naïve
(TN, CD45RA+CCR7+), effector
(TE, CD45RA+ CCR7−) and memory T
cells (CD45RA−). Memory T cells are further divided into
effector memory T cells (TEM,
CD45RA−CCR7−), which are short-lived
tissue-resident memory cells and central memory T cells
(TCM, CD45RA−CCR7+), which are
long-lived memory cells that are found in secondary lymphoid organs
(2,3). After an immune response is induced,
antigen-responsive TCM and TEM persist in the
memory pool and provide systemic immune surveillance in lymphoid
organs. In turn, a previous study revealed that CD8+
TEM in cervical cancer metastatic TDLN were fewer than
in metastasis free-TDLN (4).
Previous studies investigating tumor-infiltrating
effector CD4+ T cells revealed that immune escape plays
an important role in tumor growth and the incidence of cancer.
Notably, a high Th1 to Th2 ratio among tumor-infiltrating
lymphocytes is positively correlated with better prognosis in
various types of carcinoma (5–8). The
suppression of antitumor T cell responses leads to increased
regulatory T cells (Treg) in TIL and TDLN and is correlated with
poor prognosis (9–15). We previously reported that
CD11b+ APCs can induce immune tolerance, and that high
infiltration of CD11b+ APCs in gastric cancer is
associated with poor prognosis (16). Recently, the increased infiltration
of tumor-associated neutrophils (TAN) in primary tumors has been
shown to correlate with poor prognosis in head and neck, and
gastric cancers (17–19).
In mice, several studies have examined the role of
T-cell tolerance and tumor-infiltrating cells in the cancer
microenvironment. However, the cancer progression-dependent changes
of immune cells in TDLN in humans remain unclear. Thus, the aim of
the present study was to investigate the impact of metastasis and
other X factors on the CD4+ T cell subset in human
clinical samples of TDLN to shed light on cellular immune
regulation in tumor microenvironments related to gastric
cancer.
Materials and methods
Clinical samples
TDLN and primary tumors were collected from 28
consecutive patients who underwent surgical resection for gastric
adenocarcinoma at the Osaka City University Graduate School of
Medicine between 2011 and 2013. Pathological stage was determined
according to the Japanese classification of gastric carcinoma: 3rd
edition (20). The present study
was approved by the Medical Ethics Committee of Osaka City
University Hospital and written informed consent was obtained from
each patient before treatment.
We obtained about five D1 regional lymph node
samples per one case according to the Japanese gastric cancer
treatment guidelines 2010 (ver. 3) (21). Half of the lymph nodes were used for
making single cell suspensions, remaining lymph nodes were
determined for possible lymph node metastasis by
immunohistochemistry. Since we thought that necrotic changes skew
immune profiles in TDLN, lymph nodes with macroscopic metastasis
were excluded in the present study. Fresh lymph node samples were
suspended in 5 ml Roswell Park Memorial Institute (RPMI)-1640
complete medium (Wako, Osaka, Japan) supplemented with 10% fetal
bovine serum (FBS; Nichirei Biosciences, Tokyo, Japan) and 2%
penicillin/streptomycin (Wako). The lymph node samples were washed
twice with phosphate-buffered saline (PBS), sliced with a scalpel
into ~2–3 mm3 pieces and then passed through a cell
strainer (Becton-Dickinson, San Diego, CA, USA). The obtained
single-cell suspensions were mixed with 900 µl FBS and 100
µl dimethylsulfoxide (Wako), and were then cryopreserved at
-80°C until needed for analysis. After pathological diagnosis, we
divided TDLN into three groups: control, metastasis-free TDLN
(mfTDLN) and metastatic TDLN (mTDLN). We defined the control as
TDLN from patients with no lymph node metastasis who underwent
surgical resection for gastric cancer. Both mfTDLN and mTDLN were
obtained from the same patient. We applied and compared among one
control, one mfTDLN and one mTDLN per patient. The single-cell
suspensions were separated into CD4-positive and -negative cells
using a Dynal CD4 Positive Isolation kit (Life Technologies,
Carlsbad, CA, USA) according to the manufacturer’s
instructions.
Phenotypic analysis
CD4-positive single-cell suspensions were analyzed
by polychromatic flow cytometry. Briefly, the cells were incubated
with mouse anti-human CD3-APC, CD4-FITC, CD45RA-PECy7, CCR7-PE,
CD25-PECy7 or programmed cell death-1 (PD-1)-PE antibodies (BD
Pharmingen, San Diego, CA, USA). To detect myeloid-derived APCs and
neutrophils, whole single-cell suspensions collected from lymph
nodes were incubated with CD11b-PE, CD11c-APC, HLA-DR-FITC,
programmed cell death ligand 1 (PD-L1)-PE or CD16b-PE antibodies
(BD Pharmingen), and then analyzed by flow cytometry. Flow
cytometric analyses were performed using a BD LSR II flow cytometer
with FACSDiva™ software (both from Becton-Dickinson).
Immunohistochemistry
Primary tumor specimens of the largest size from
each patient were fixed in 10% formalin, embedded in paraffin and
serially sectioned (4-µm thickness) for immunohistochemistry
analysis. After antigen retrieval by autoclaving at 105°C for 10
min, the tissue sections were incubated overnight at 4°C with
rabbit anti-human anti-CD4 monoclonal antibody (ab133616, 1:100
dilution; Abcam, Cambridge, UK) or anti-CD8 monoclonal antibody
(NCL-CD8–4B11, 1:40 dilution; Leica, Newcastle, UK). The signal was
amplified by streptavidin-biotin complex formation and developed
with 3,3′-diaminobenzidine (DAB) followed by counterstaining with
hematoxylin. All cell counts were performed using a BX41 microscope
(Olympus, Tokyo, Japan) at ×200 magnification (×20 objective and
×10 eyepiece). Samples were scored in a blinded manner by two
investigators with respect to clinicopathological features. For
each section, five areas of a representative field of tumor were
assessed.
Intracellular staining and T cell
activation
Intracellular staining for FOXP3 (Ab-PE) was
performed using a FOXP3 buffer set (BD Pharmingen) according to the
manufacturer’s instructions. To assess the TE subset,
isolated CD4+ T cells were activated using a Leukocyte
Activation Cocktail with GolgiPlug™ (BD Pharmingen) according to
the manufacturer’s instructions. Briefly, lymph node lymphocytes
were stimulated with the Leukocyte Activation Cocktail mixture
containing a phorbol ester (phorbol 12-myristate 13-acetate, PMA),
calcium ionophore (Ionomycin), and protein transport inhibitor (BD
GolgiPlug™, containing brefeldin A) in 24-well plates at a
concentration of 1×106 cells/ml for 5 h at 37°C with 5%
CO2 in RPMI-1640 complete medium. To stain the cell
surface, the activated cells were mixed with mouse anti-human
FITC-conjugated anti-CD4 antibody, and incubated for 30 min on ice.
To detect intracellular cyto-kines, a Cytofix/Cytoperm
intracellular cytokine staining kit (BD Pharmingen) was used
according to the manufacturer’s instructions. Briefly, cells were
re-suspended in 100 µl Cytofix/Cytoperm™ solution for 20 min
at 4°C for fixation and permeabilization, and were then washed
twice in 1X Perm/Wash™ solution, pelleted by centrifugation at 400
× g for 5 min at 4°C, and separated from the supernatant. After
fixation and permeabilization, cells were incubated with
appropriate antibodies for 30 min on ice.
Quantitative real-time (RT)-PCR
Total RNA was prepared using an RNeasy Micro kit
(Qiagen, Chatsworth, CA, USA) according to the manufacturer’s
instructions. cDNA was synthesized from 100 ng mRNA by standard
RT-PCR amplification with ReverTra Ace qPCR RT Master mix (Toyobo,
Osaka, Japan). TaqMan PCRs for interferon (IFN)-γ (Hs00989291_m1),
interleukin (IL)-12A (IL-12p35, Hs01073447_m1), IL-8
(Hs00174103_m1), matrix metalloproteinase 2 (MMP2) (Hs01548727_m1)
and matrix metalloproteinase 9 (MMP9) (Hs00234579_m1) were
performed using an ABI Prism Detection System (Applied Biosystems).
For amplification, 2.5 µl cDNA was incubated with 12.5
µl 2X TaqMan Master mix (8% glycerol, 1X TaqMan buffer, 200
µmol/l dATP, 200 µmol/l dGTP, 200 µmol/l dCTP,
400 µmol/l dUTP, 0.05 U/µl AmpErase uracil
N-glycosylase, 5 mmol/l MgCl2 and 0.01 U/µl
AmpliTaq Gold; Perkin-Elmer, Wellesley, MA, USA), 2.5 µl
TaqMan probe and sense and reverse primers at a final concentration
of 10 pmol/µl. The reaction mixture was adjusted to a final
volume of 25 µl with RNase-free distilled water. For
quantification of the human housekeeping gene
glyceraldehyde-3-phosphate dehydrogenase, a Pre-Developed Assay kit
(Perkin-Elmer) was used. The amplification was performed and
analyzed using an ABI Prism 7700 Sequence Detector. To rule out
contamination from buffers and tubes, a negative control with water
in place of the cDNA template was included on every plate.
Statistical analysis
The association between lymph node metastasis and
various clinicopathological variables was examined, and the
significance of different prognostic markers was evaluated using
binomial logistic regression analysis. The level of significance
was set at P<0.05. Statistical analyses were performed using
SPSS 13.0 statistical software (SPSS, Inc., Chicago, IL, USA).
Results
Impact of metastasis on differentiation
of CD4+ T cells in lymph nodes
Fresh lymph node samples were collected from 28
gastric adenocarcinoma patients, which consisted of 15 patients
with pathological nodal metastasis and 13 patients without
metastasis. No significant differences between patient
characteristics, including gender, age, histological type, tumor
size and lymphovascular invasion were detected based on lymph node
metastasis status (Table I). Flow
cytometric analysis revealed that the purity of isolated
CD4+ T cells was >98% (data not shown).
 | Table ICorrelation between lymph node
metastasis and clinicopathological characteristics in gastric
cancer. |
Table I
Correlation between lymph node
metastasis and clinicopathological characteristics in gastric
cancer.
| Variables | N | Lymph node status
| P-value | HR | 95% CI |
|---|
| Control (n=13) | Metastatic LN
(n=15) |
|---|
| Gender | | | | | | |
| Male | 20 | 9 | 11 | 0.453 | 2.065 | 0.298–14.261 |
| Female | 8 | 4 | 4 | | | |
| Age (years) | | 68 | 67 | 0.789 | 0.990 | 0.918–1.066 |
| Histological
type | | | | | | |
| Intestinal | 12 | 4 | 8 | 0.226 | 2.571 | 0.562–13.207 |
| Diffuse | 16 | 9 | 7 | | | |
| Tumor size
(mm) | | 49.5 | 62.2 | 0.257 | 0.985 | 0.955–1.010 |
| T | | | | | | |
| 1–2 | 21 | 13 | 8 | 0.705 | 0.750 | 0.169–3.325 |
| 3–4 | 7 | 0 | 7 | | | |
| V | | | | | | |
| (−) | 23 | 11 | 12 | 0.824 | 0.8 | 0.091–5.776 |
| (+) | 5 | 2 | 3 | | | |
| Ly | | | | | | |
| (−) | 11 | 6 | 5 | 0.488 | 0.583 | 0.121–2.687 |
| (+) | 17 | 7 | 10 | | | |
| Stage | | | | | | |
| 1 | 9 | 7 | 2 | 0.007 | 7.512 | 1.727–32.623 |
| 2 | 10 | 6 | 4 | | | |
| 3 | 7 | 0 | 7 | | | |
| 4 | 2 | 0 | 2 | | | |
To examine the inhibitory effect of metastasis on
CD4+ T cell differentiation in TDLN, we investigated the
proportion of four subsets of CD4+ T cells:
TE, TN, TEM and TCM,
and calculated the percentage of each group among total
CD4+ T cells isolated from the patients lymph nodes
based on flow cytometric analysis. As shown in Fig. 1A and B, the percentage of
TE cells in mTDLN was significantly lower than that in
mfTDLN (P=0.021). TE cells in mfTDLN were also increased
compared with the normal control samples, whereas the number of
TEM cells in mTDLN was significantly lower than in the
control samples (P=0.010) and mfTDLN (P=0.041). No significant
differences in the proportions of TN and TCM
cells were detected. Despite the lack of CCR7, the lymph node
homing receptor, in TDLN, TEM were the predominant
CD4+ T cell subset in TDLN.
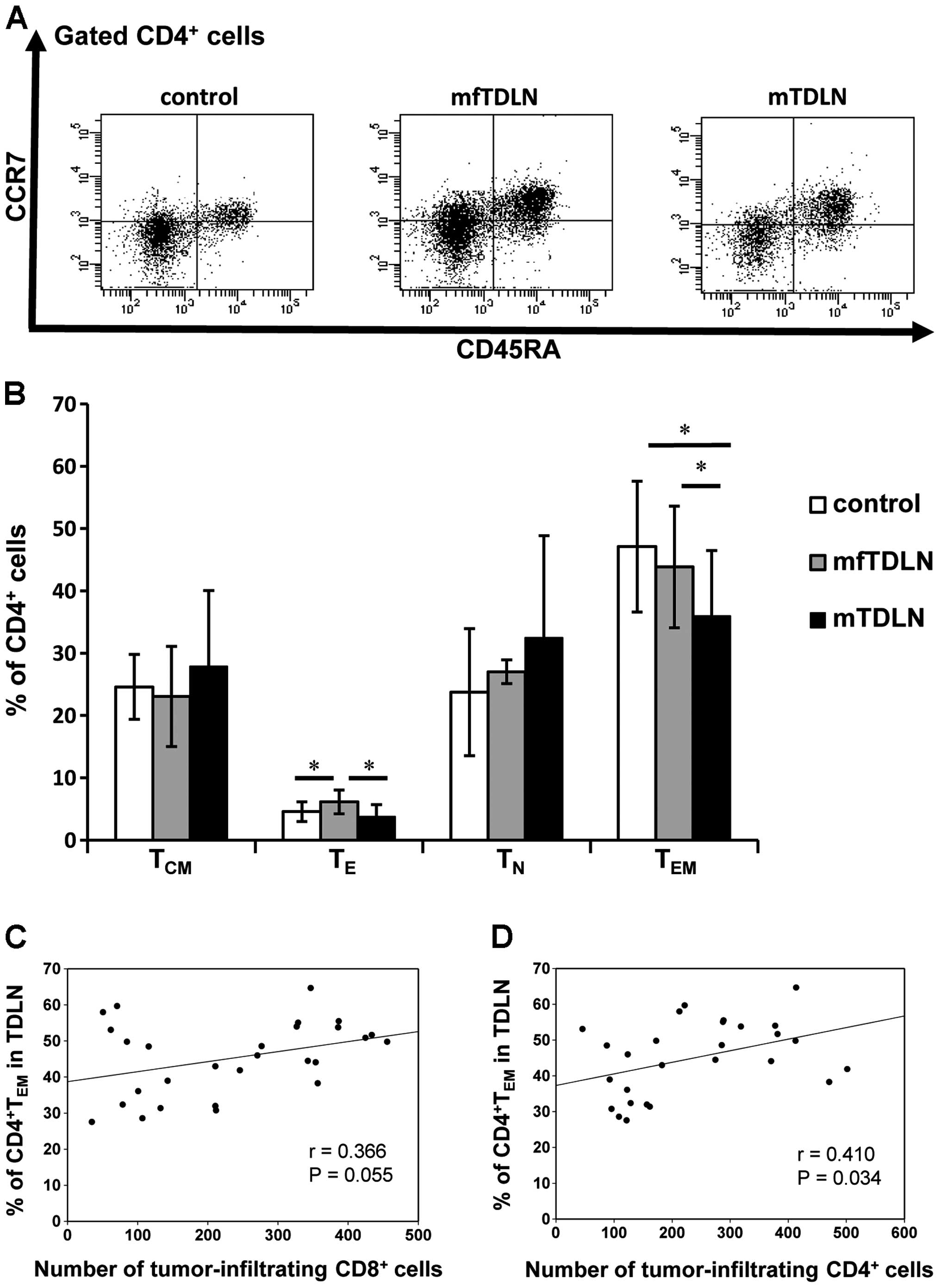 | Figure 1Flow cytometric analysis of
CD4+ T cell subsets in TDLN. (A) Cells in TDLN were
mechanically dissociated into single-cell suspensions and
CD4+ T cells were then obtained using a CD4+
T cell isolation kit in combination with flow cytometry. Results
were gated on CD4+ cells, and representative dot plots
of control TDLN, mfTDLN and mTDLN are shown. CD4+ cells
were subdivided into four populations:
CD45RA−CCR7− cells (TEM),
CD45RA-CCR7+ cells (TCM),
CD45RA+CCR7− cells (TE) and
CD45RA+CCR7+ cells (TN). (B)
Proportion of each T cell subset among total CD4+ T
cells was compared between control TDLN (white bars), mfTDLN (gray
bars) and mTDLN (black bars). Histograms show mean values ± SD from
analyses performed for 13 control TDLN, 15 mfTDLN and 15 mTDLN.
Comparisons between groups were performed using the t-test. The
number of TE cells in mfTDLN was significantly higher
than in control (P=0.025) and mTDLN (P=0.021). TEM in
mTDLN was significantly lower than in control and mfTDLN. No
significant differences in TN or TCM were
detected. (C and D) Spearman’s correlation coefficients were used
to determine the relationship between the proportion of
CD4+ TEM cells in control and mTDLN and the
number of tumor-infiltrating (C) CD4+ or (D)
CD8+ T cells. Tumor-infiltrating CD4+ T cells
had a significantly positive correlation with CD4+
TEM in TDLN (r=0.410, P=0.034). Tumor-infiltrating
CD8+ T cells were correlated with CD4+
TEM in TDLN, yet the association was not significant
(r=0.366, P=0.055). Asterisk (*) shows a significant difference
(P<0.05). TDLN, tumor-draining lymph node; mfTDLN,
metastatic-free TDLN; mTDLN, metastatic TDLN; TEM,
effector memory T cells; TE, effector T cells. |
We next examined the correlation between the number
of tumor-infiltrating CD4+ and CD8+ T cells
in primary tumors and CD4+ T cell subset in TDLN. As
shown in Fig. 1C and D, a positive
relationship was observed for CD4+ and TEM
cells (r=0.410, P=0.034), suggesting that TEM cells in
TDLN could be considered tumor-related CD4+ T cells.
Impact of metastasis on effector T cell
subsets in lymph nodes
To investigate the effects of metastasis on the
function of effector T cell subsets, we examined differences in the
proportions of TE CD4+ cells among normal
control lymph nodes, mfTDLN and mTDLN by intracellular cytokine
detection using multicolor flow cytometric analysis. The examined
cell populations were defined as follows: Th1, INF-γ-positive and
IL-4-negative; Th2, INF-γ-negative and IL-4-positive; Th17,
IL-17-positive; and Treg, CD25-positive and FOXP3-positive. As
shown in Fig. 2A, the Th1/Th2 ratio
significantly decreased in both mfTDLN (P=0.015) and mTDLN
(P=0.001) compared with normal lymph nodes. Although no significant
differences in the Th1/Th2 ratio were found between mfTDLN and
mTDLN, mTDLN and mfTDLN (Fig. 2A).
The proportion of Th17 cells did not markedly differ among the
three lymph node groups (Fig. 2B),
whereas Treg cells were increased in mTDLN compared with mfTDLN
(Fig. 2C). In addition, the number
of PD-1+CD4 T cells appeared to be increased in mTDLN
compared to mfTDLN (P=0.07) and normal lymph nodes (P=0.09),
although the difference was not significant (Fig. 2D).
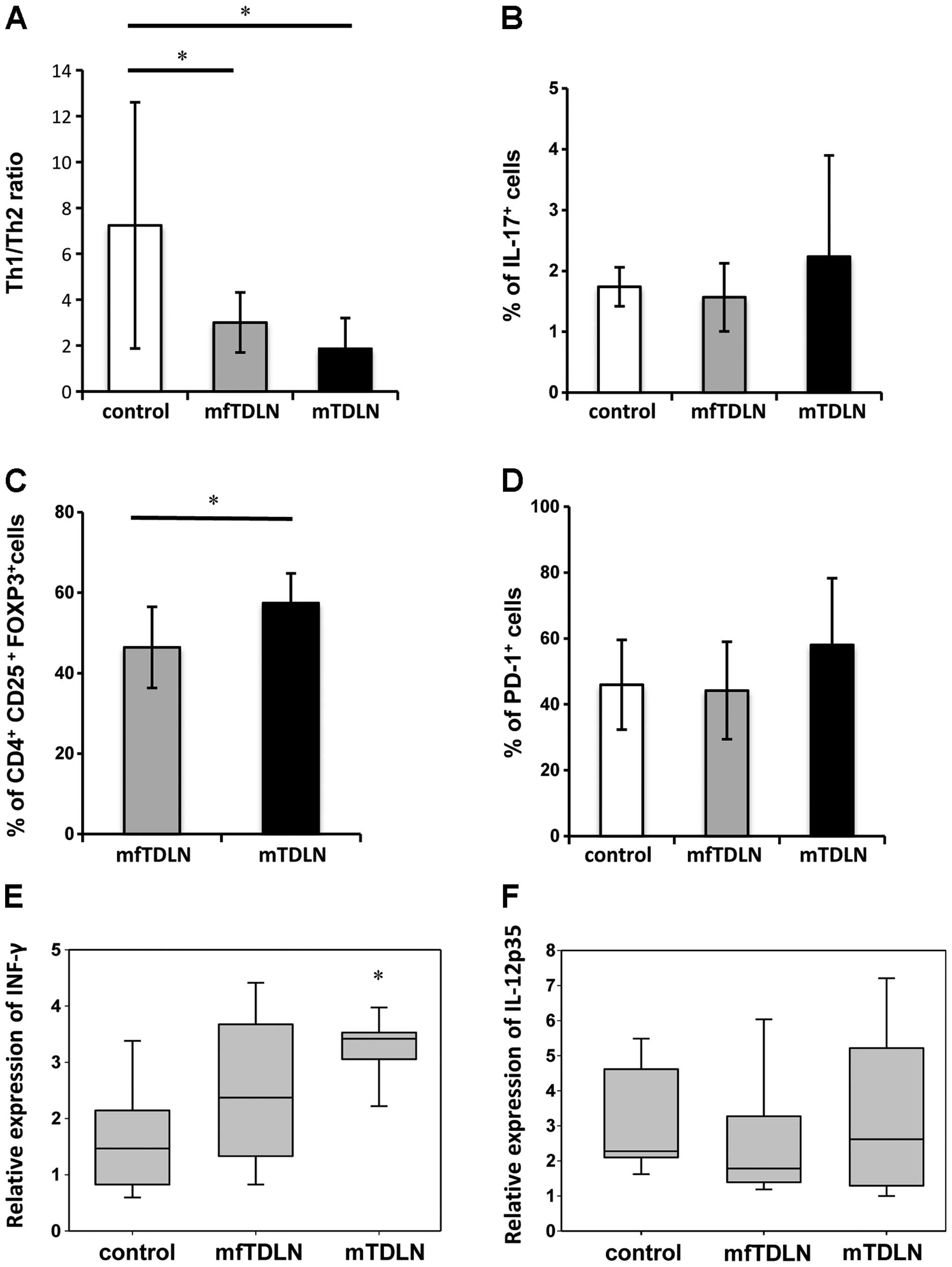 | Figure 2CD4+ effector T cell
subsets and T cell exhaustion in TDLN. Isolated CD4+ T
cells were analyzed by intracellular cytokine staining using
multicolor flow cytometry. (A) Th1 and Th2 cells were detected by
intracellular INF-γ and IL-4 staining, respectively. The Th1/Th2
ratio was assessed by analyzing the proportion of INF-γ+
and IL-4+ cells among total CD4+ cells.
Significant differences in the Th1/Th2 ratio were detected between
mfTDLN and mTDLN, and between the control lymph nodes and mfTDLN.
(B) Th17 cells were detected by intracellular IL-17 staining, and
the proportion of IL-17+ among total CD4+
cells is shown. No significant differences in the proportion of
Th17 cells were detected among the three groups. (C) Treg cells
were detected based on the surface expression of CD4+
CD25+ and intracellular FOXP3 staining, and the
proportion of FOXP3+ cells among total
CD4+CD25+ cells is shown. A significant
difference between mfTDLN and mTDLN was detected. (D) To assess the
exhaustion of CD4+ T cells, PD-1 expression on
CD4+ T cells was analyzed by flow cytometry. There is no
significant differences in PD-1+CD4+ cell
infiltration among three groups. Relative mRNA expression of (E)
IFN-γ and (F) IL-12p35 was compared among control TDLN, mfTDLN and
mTDLN by qRT-PCR. Significantly higher IFN-γ expression was
observed in control TDLN compared to mfTDLN (P=0.012). An asterisk
(*) indicates a significant difference (P<0.05). TDLN,
tumor-draining lymph node; mfTDLN, metastatic-free TDLN; mTDLN,
metastatic TDLN; PD-1, programmed cell death-1. |
To examine the influence of the tumor
microenvironment on the levels of the different T cell subsets, the
mRNA expression of IFN-γ and IL-12 was measured in lymph nodes.
Although IL-12 expression was not upregulated in mTDLNs, IFN-γ was
significantly elevated (P=0.012, Fig.
2E and F).
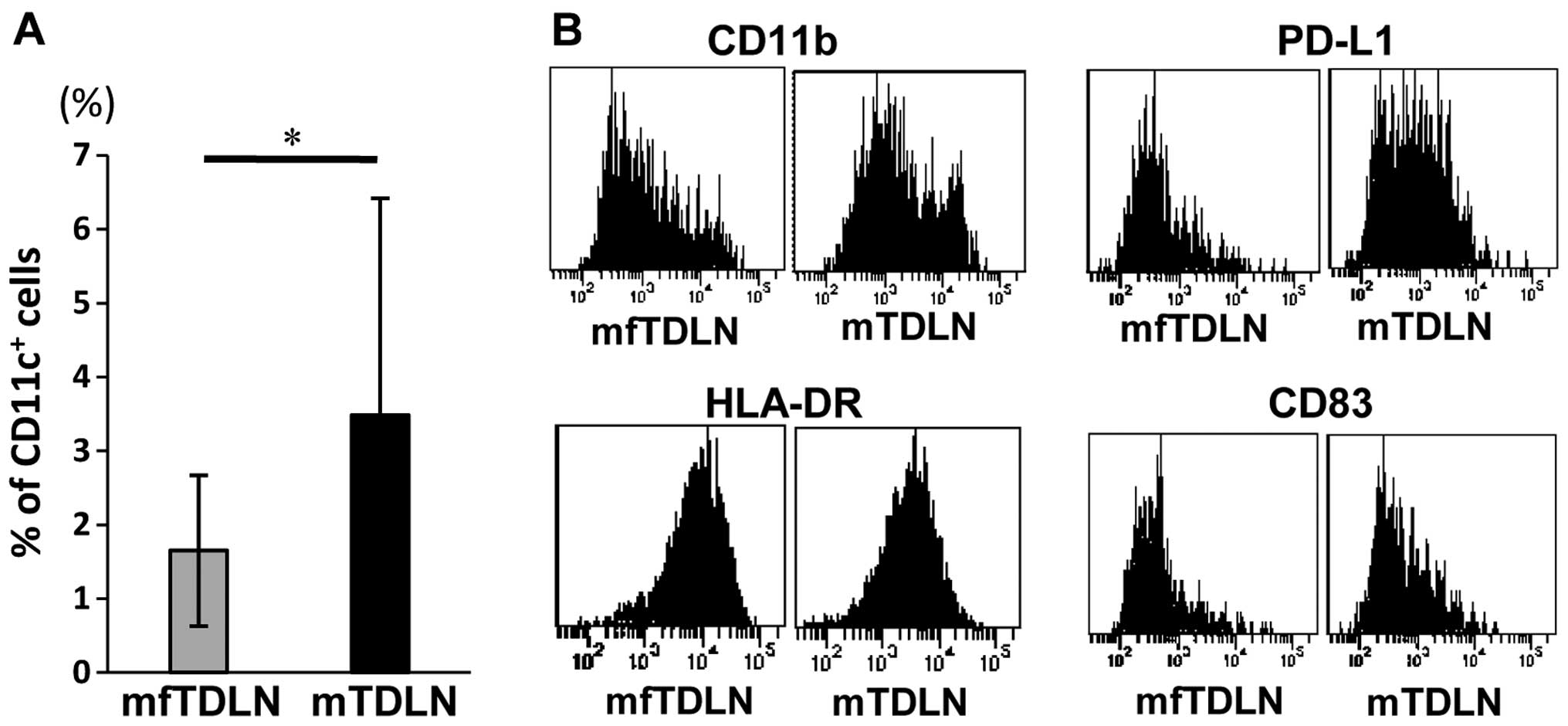
Alteration of APC phenotype in TDLN
To determine the factors that influence
TE in TDLN, the phenotype of CD11c+ cells
isolated from lymph nodes was examined (Fig. 3). The proportion of
CD11c+ cells in mTDLN was significantly higher than in
mfTDLN (Fig. 3A). We have
previously reported that gastric cancer cells augment CD11b and
PD-L2 expression on CD11c+ cells and attenuate HLA-DR
expression by the production of immunosuppressive cytokines
secreted by gastric cancer cells (16). In the present study, the expression
of CD11b within CD11c+ cells in mTDLN was higher than
that in mfTDLN (Fig. 3B). PD-L1
expression on CD11c+ cells in mTDLN was also higher than
that in mfTDLN (Fig. 3B). However,
no significant differences in the expression of MHC-class II
(HLA-DR) or CD83 were detected (Fig.
3B).
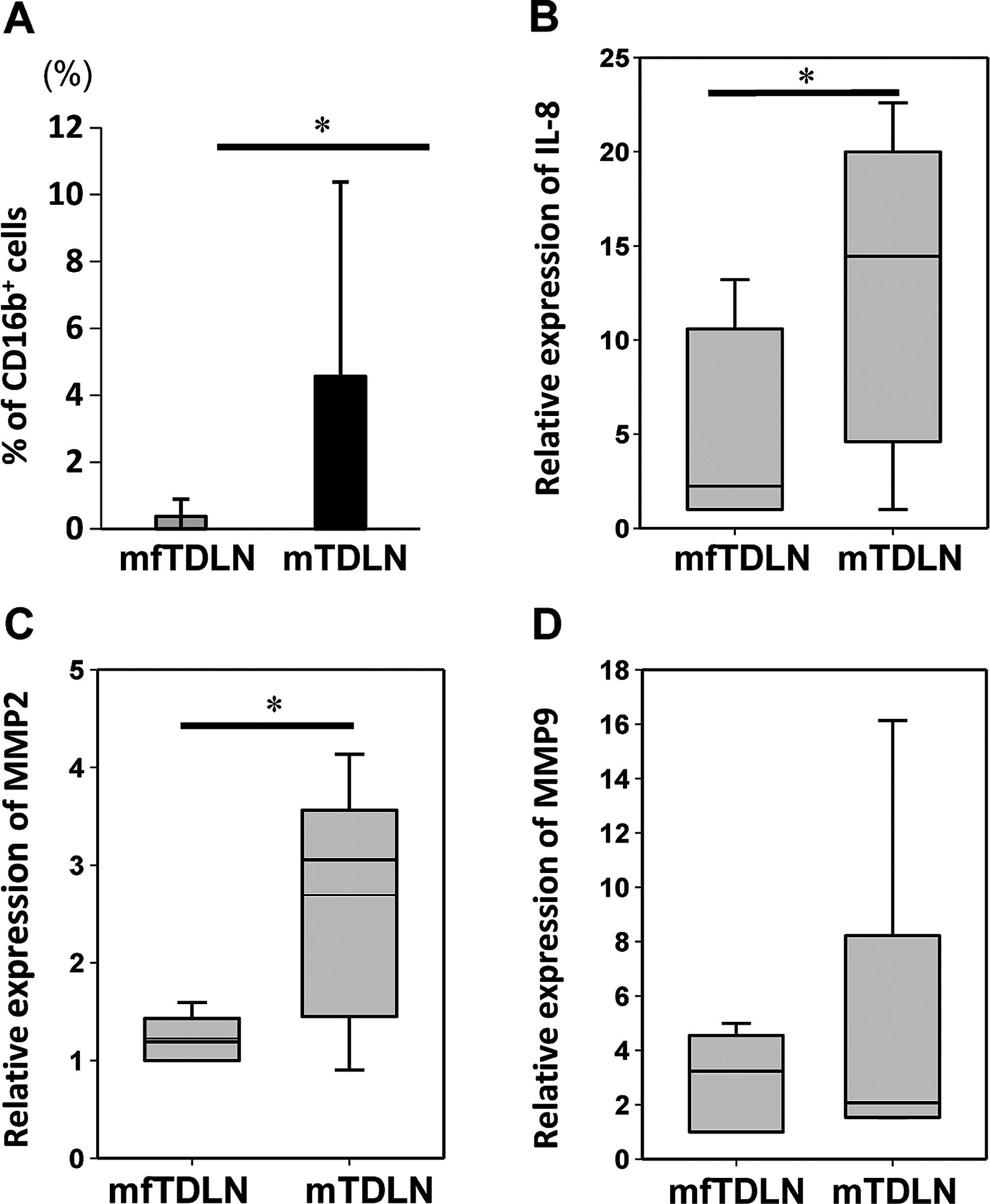
Neutrophil-related cytokines and proteins
in TDLN
To examine whether cytokines produced by neutrophils
were associated with the decrease of TE in TDLN, we
determined and compared the proportion of neutrophils
(CD16b+) in mTDLN and mfTDLN (Fig. 4A). The flow cytometric analysis
revealed that neutrophil infiltration was significantly higher in
mTDLN compared to mfTDLN (P=0.016). Based on this finding, we next
measured the mRNA expression levels of neutrophil-related
chemokines in cells isolated from mTDLN and mfTDLN by qRT-PCR.
Increased expression of IL-8 and/or its receptors is a
characteristic of cancer and endothelial cells, infiltrating
neutrophils and tumor-associated macrophages, suggesting that IL-8
functions as a significant regulatory factor within the tumor
microenvironment (22,23). In the present analysis, IL-8, which
induces neutrophils to migrate out of the bloodstream, was
expressed at significantly higher levels in mTDLNs compared to
mfTDLNs (P=0.013; Fig. 4B).
Tumor-associated neutrophils reportedly contribute to tumor
angiogenesis and invasion by the production of MMP2 and MMP9
(23,24), which subsequently break down
extracellular matrix proteins, such as collagens and release the
cryptic information. We examined mRNA expression of MMP2 and MMP9,
which are required for the initial recruitment of neutrophils into
tissues, and found that MMP2 but not MMP9, showed significantly
higher expression in mTDLN than in mfTDLN (P=0.013, Fig. 4C and D).
Discussion
In the present study, we demonstrated that the
populations of effector and effector memory CD4+ T cells
are decreased in metastatic lymph nodes, whereas Tregs,
CD11b+ APCs and neutrophils are increased. To the best
of our knowledge, this is the first study to elucidate the changes
that occur in the populations of immune cells in TDNL from human
clinical samples. Considering the important roles that
CD4+ T cells play in antitumor immune responses, these
findings will help define the cellular events that occur within the
tumor microenvironment in regional lymph nodes of gastric
cancer.
Tumor-infiltrating CD4+ TEM
cells capable of homing to the tumor microenvironment are
responsible for long-term protection against tumors (4,25).
Several investigators have shown that infiltrating
CD45RO+ memory T cells are decreased in primary tumors
compared with normal tissue, and that the high tumor infiltration
of such cells correlates with favorable prognosis (7,26,27).
In the present study, we observed that TEM are decreased
in mTDLNs. As shown in a previous study, PD-L1 expressed on cancer
cells has negative regulatory effects on CD4+ memory T
cells (28). Thus, it is possible
that CD4+ TEM in lymph nodes may generate
less cytokines, such as IL-7 and IL-15 by surrounding cancer cells,
resulting in a decrease of TE (29). Although it was previously reported
that TCM cells correlate with antitumor functions
(30), in the resent study, no
significantly differences in the proportion of TCM were
observed among the three types of lymph node tissue. TCM
do not produce any prototypic cytokines of effector cells
immediately after stimulation (31,32),
suggesting that TCM cells likely undergo a secondary
response. However, since large differences in the proportion of
TCM were observed between patients in the present study,
future studies examining a greater number of patients may be
required to better elucidate the contribution of TCM
towards long-term antitumor responses.
TEM produce signals that are involved in
the generation of TE cells. In the present study, we
found that the population of TE is increased in mfTDLN
compared with control TDLN, suggesting the existence of an
intratumoral adaptive immune response. Alternatively, it is
possible that subsets of Tregs or Th2 cells that regulate antitumor
immune responses are augmented in mfTDLN. CD4+
TE cells can change their cytokine secretion profiles by
genetic reprogramming (33,34). Our present data showed that the
Th1/Th2 ratio is decreased in mTDLN, this finding is consistent
with the decrease in the TEM population in mTDLN.
Although our results also demonstrated that the population of Treg
cells is increased in mTDLN, we did not find direct evidence for
the link between increasing numbers of Tregs and metastasis.
However, our finding that IFN-γ expression is upregulated in mTDLN
may be related with the protumoral functions of IFN-γ, including
the differentiation of Tregs and suppression of cytotoxic T
lymphocytes (35,36).
The underlying cellular mechanism of local immune
regulation in regional lymph nodes remains to be determined. We
speculate that naïve CD4+ T cells which attach to
regulatory dendritic cells (DC) fail to receive further stimulatory
signals. These DCs, in turn, may be affected by immunosuppressive
cytokines, such as transforming growth factor (TGF)-β and IL-10
produced by cancer cells. We previously reported that
tumor-infiltrating CD11b+ DCs affect the tumor
microenvi-ronment and prognosis (16), and that a positive correlation
exists between the number of tumor-infiltrating CD11b+
APC and Treg cells. In the present study, an increased population
of CD11c+ APCs, which augment expression of the negative
stimulatory markers CD11b and PD-L1 was observed in mTDLN from the
gastric cancer patients.
Moreover, in the present study, DCs in mTDLN did not
show downregulated expression of MHC class II (HLA-DR) and did not
upregulate CD83 expression, suggesting that they had a semi-mature
phenotype. These findings, taken together with our present results,
suggest that gastric cancer cells promotes the differentiation of
APC into tolerogenic semi-mature CD11b+
PD-L1+ APC, which induce the proliferation of Tregs,
resulting in decreased populations of TE and
TEM.
Tumor-infiltrating neutrophils consist of
antitumoral (N1) and pro-tumoral (N2) types, which have contrasting
roles in cancer immunity (37).
Neutrophils also have the ability to inhibit the antitumoral
function of DC, and neutrophil-derived ectosomes, cathelicidins and
elastase suppress DC activation and maturation (38). Our present results indicate that
N2-type neutrophils induced by gastric cancer cells also promote
the formation of immunosuppressive DC and correlate with the skewed
CD4+ T cell and CD4 TE subsets in the
advanced stages of disease. In addition, we showed that IL-8 and
MMP2 expression is increased in mTDLNs. Although both cancer and
immune cells are able to produce IL-8 and MMP2, we could not
determine which cell type predominantly produced IL-8 and MMP2 in
TDLN. It has been reported that neutrophils activated by IL-8
inhibits T cell receptor expression and cytokine secretion
depending on the H2O2 concentration (39).
Our data suggest that MMP2 from cancer cells or
neutrophils contributes to the destruction of lymphatic vessels in
lymph nodes and promotes lymph node metastasis.
Several limitations of the present study warrant
mention. First, we did not provide direct evidence that DCs in
lymph nodes induce the proliferation of Tregs and reduce Th1
CD4+ T cells. Second, the association between increasing
neutrophils and decreasing TE cells was not conclusively
determined. Finally, the cell type responsible for the secretion of
IL-8 and MMP2 was not determined. To elucidate the cellular
mechanisms underlying the regulation of antitumor immune response
in TDLNs, separate functional analyses of each subset of T cells,
neutrophils and DCs are needed. Moreover, the molecular mechanisms
of this process remain unclear, but may involve a suppressor of
cytokine signal (SOCS) that regulates the proliferation of
TE or TEM (40).
In conclusion, the present data indicate that
CD4+ T cell and TE subsets in TDLN of gastric
cancer patients are significantly affected by metastasis.
Immunosuppressive cells, such as CD11b+
PD-L1+ DC, neutrophils and Treg cells, exhibit increased
migration to TDLN, in which a subset of CD4+ effector
cells is skewed towards immune tolerance in the tumor
microenvironment. Our findings of the local alteration of immune
cells in TDLN shed light on the immunological mechanisms that occur
in the tumor microenvironment and are expected to aid in the design
of effective immune therapy for patients with gastric cancer.
Acknowledgments
This study was supported by a Grant-in-Aid for
Scientific Research (to H.T. and K.H.).
References
|
1
|
Shurin GV, Ma Y and Shurin MR:
Immunosuppressive mechanisms of regulatory dendritic cells in
cancer. Cancer Microenviron. 6:159–167. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sallusto F, Lenig D, Förster R, Lipp M and
Lanzavecchia A: Two subsets of memory T lymphocytes with distinct
homing potentials and effector functions. Nature. 401:708–712.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sallusto F and Lanzavecchia A:
Heterogeneity of CD4+ memory T cells: Functional modules
for tailored immunity. Eur J Immunol. 39:2076–2082. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Battaglia A, Buzzonetti A, Baranello C,
Ferrandina G, Martinelli E, Fanfani F, Scambia G and Fattorossi A:
Metastatic tumour cells favour the generation of a tolerogenic
milieu in tumour draining lymph node in patients with early
cervical cancer. Cancer Immunol Immunother. 58:1363–1373. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ito N, Suzuki Y, Taniguchi Y, Ishiguro K,
Nakamura H and Ohgi S: Prognostic significance of T helper 1 and 2
and T cytotoxic 1 and 2 cells in patients with non-small cell lung
cancer. Anticancer Res. 25:2027–2031. 2005.PubMed/NCBI
|
|
6
|
Tosolini M, Kirilovsky A, Mlecnik B,
Fredriksen T, Mauger S, Bindea G, Berger A, Bruneval P, Fridman WH,
Pagès F, et al: Clinical impact of different classes of
infiltrating T cytotoxic and helper cells (Th1, th2, treg, th17) in
patients with colorectal cancer. Cancer Res. 71:1263–1271. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Enomoto K, Sho M, Wakatsuki K, Takayama T,
Matsumoto S, Nakamura S, Akahori T, Tanaka T, Migita K, Ito M, et
al: Prognostic importance of tumour-infiltrating memory T cells in
oesophageal squamous cell carcinoma. Clin Exp Immunol. 168:186–191.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kristensen VN, Vaske CJ, Ursini-Siegel J,
Van Loo P, Nordgard SH, Sachidanandam R, Sørlie T, Wärnberg F,
Haakensen VD, Helland Å, et al: Integrated molecular profiles of
invasive breast tumors and ductal carcinoma in situ (DCIS) reveal
differential vascular and interleukin signaling. Proc Natl Acad Sci
USA. 109:2802–2807. 2012. View Article : Google Scholar :
|
|
9
|
Bates GJ, Fox SB, Han C, Leek RD, Garcia
JF, Harris AL and Banham AH: Quantification of regulatory T cells
enables the identification of high-risk breast cancer patients and
those at risk of late relapse. J Clin Oncol. 24:5373–5380. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gnerlich JL, Mitchem JB, Weir JS, Sankpal
NV, Kashiwagi H, Belt BA, Porembka MR, Herndon JM, Eberlein TJ,
Goedegebuure P, et al: Induction of Th17 cells in the tumor
microenvironment improves survival in a murine model of pancreatic
cancer. J Immunol. 185:4063–4071. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mougiakakos D, Johansson CC, Trocme E,
All-Ericsson C, Economou MA, Larsson O, Seregard S and Kiessling R:
Intratumoral forkhead box P3-positive regulatory T cells predict
poor survival in cyclooxygenase-2-positive uveal melanoma. Cancer.
116:2224–2233. 2010.PubMed/NCBI
|
|
12
|
Lee HE, Park DJ, Kim WH, Kim HH and Lee
HS: High FOXP3+ regulatory T-cell density in the
sentinel lymph node is associated with downstream non-sentinel
lymph-node metastasis in gastric cancer. Br J Cancer. 105:413–419.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim M, Grimmig T, Grimm M, Lazariotou M,
Meier E, Rosenwald A, Tsaur I, Blaheta R, Heemann U, Germer CT, et
al: Expression of Foxp3 in colorectal cancer but not in Treg cells
correlates with disease progression in patients with colorectal
cancer. PLoS One. 8:e536302013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nakamura R, Sakakibara M, Nagashima T,
Sangai T, Arai M, Fujimori T, Takano S, Shida T, Nakatani Y and
Miyazaki M: Accumulation of regulatory T cells in sentinel lymph
nodes is a prognostic predictor in patients with node-negative
breast cancer. Eur J Cancer. 45:2123–2131. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Deng L, Zhang H, Luan Y, Zhang J, Xing Q,
Dong S, Wu X, Liu M and Wang S: Accumulation of foxp3+ T
regulatory cells in draining lymph nodes correlates with disease
progression and immune suppression in colorectal cancer patients.
Clin Cancer Res. 16:4105–4112. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Okita Y, Tanaka H, Ohira M, Muguruma K,
Kubo N, Watanabe M, Fukushima W and Hirakawa K: Role of
tumor-infiltrating CD11b+ antigen-presenting cells in
the progression of gastric cancer. J Surg Res. 186:192–200. 2014.
View Article : Google Scholar
|
|
17
|
Carus A, Ladekarl M, Hager H, Nedergaard
BS and Donskov F: Tumour-associated CD66b+ neutrophil
count is an independent prognostic factor for recurrence in
localised cervical cancer. Br J Cancer. 108:2116–2122. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao JJ, Pan K, Wang W, Chen JG, Wu YH, Lv
L, Li JJ, Chen YB, Wang DD, Pan QZ, et al: The prognostic value of
tumor-infiltrating neutrophils in gastric adenocarcinoma after
resection. PLoS One. 7:e336552012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Houghton AM: The paradox of
tumor-associated neutrophils: Fueling tumor growth with cytotoxic
substances. Cell Cycle. 9:1732–1737. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Japanese Gastric Cancer Association:
Japanese classification of gastric carcinoma: 3rd English edition.
Gastric Cancer. 14:101–112. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sano T and Aiko T: New Japanese
classifications and treatment guidelines for gastric cancer:
Revision concepts and major revised points. Gastric Cancer.
14:97–100. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Waugh DJ and Wilson C: The interleukin-8
pathway in cancer. Clin Cancer Res. 14:6735–6741. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shamamian P, Schwartz JD, Pocock BJ, Monea
S, Whiting D, Marcus SG and Mignatti P: Activation of progelatinase
A (MMP-2) by neutrophil elastase, cathepsin G, and proteinase-3: A
role for inflammatory cells in tumor invasion and angiogenesis. J
Cell Physiol. 189:197–206. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bekes EM, Schweighofer B, Kupriyanova TA,
Zajac E, Ardi VC, Quigley JP and Deryugina EI: Tumor-recruited
neutrophils and neutrophil TIMP-free MMP-9 regulate coordinately
the levels of tumor angiogenesis and efficiency of malignant cell
intravasation. Am J Pathol. 179:1455–1470. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Poschke I, De Boniface J, Mao Y and
Kiessling R: Tumor-induced changes in the phenotype of
blood-derived and tumor-associated T cells of early stage breast
cancer patients. Int J Cancer. 131:1611–1620. 2012. View Article : Google Scholar
|
|
26
|
Pagès F, Kirilovsky A, Mlecnik B, Asslaber
M, Tosolini M, Bindea G, Lagorce C, Wind P, Marliot F, Bruneval P,
et al: In situ cytotoxic and memory T cells predict outcome in
patients with early-stage colorectal cancer. J Clin Oncol.
27:5944–5951. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mlecnik B, Tosolini M, Kirilovsky A,
Berger A, Bindea G, Meatchi T, Bruneval P, Trajanoski Z, Fridman
WH, Pagès F, et al: Histopathologic-based prognostic factors of
colorectal cancers are associated with the state of the local
immune reaction. J Clin Oncol. 29:610–618. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dong H, Strome SE, Salomao DR, Tamura H,
Hirano F, Flies DB, Roche PC, Lu J, Zhu G, Tamada K, et al:
Tumor-associated B7-H1 promotes T-cell apoptosis: A potential
mechanism of immune evasion. Nat Med. 8:793–800. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bradley LM, Haynes L and Swain SL: IL-7:
Maintaining T-cell memory and achieving homeostasis. Trends
Immunol. 26:172–176. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wu F, Zhang W, Shao H, Bo H, Shen H, Li J,
Liu Y, Wang T, Ma W and Huang S: Human effector T cells derived
from central memory cells rather than CD8+T cells
modified by tumor-specific TCR gene transfer possess superior
traits for adoptive immunotherapy. Cancer Lett. 339:195–207. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kedzierska K, Valkenburg SA, Doherty PC,
Davenport MP and Venturi V: Use it or lose it: Establishment and
persistence of T cell memory. Front Immunol. 3:3572012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mueller SN, Gebhardt T, Carbone FR and
Heath WR: Memory T cell subsets, migration patterns, and tissue
residence. Annu Rev Immunol. 31:137–161. 2013. View Article : Google Scholar
|
|
33
|
Zhou L, Chong MM and Littman DR:
Plasticity of CD4+ T cell lineage differentiation.
Immunity. 30:646–655. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhu J and Paul WE: Heterogeneity and
plasticity of T helper cells. Cell Res. 20:4–12. 2010. View Article : Google Scholar
|
|
35
|
Brody JR, Costantino CL, Berger AC, Sato
T, Lisanti MP, Yeo CJ, Emmons RV and Witkiewicz AK: Expression of
indoleamine 2,3-dioxygenase in metastatic malignant melanoma
recruits regulatory T cells to avoid immune detection and affects
survival. Cell Cycle. 8:1930–1934. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ostrand-Rosenberg S and Sinha P:
Myeloid-derived suppressor cells: Linking inflammation and cancer.
J Immunol. 182:4499–4506. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fridlender ZG, Sun J, Kim S, Kapoor V,
Cheng G, Ling L, Worthen GS and Albelda SM: Polarization of
tumor-associated neutrophil phenotype by TGF-beta: ‘N1’ versus ‘N2’
TAN. Cancer Cell. 16:183–194. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Schuster S, Hurrell B and Tacchini-Cottier
F: Crosstalk between neutrophils and dendritic cells: A
context-dependent process. J Leukoc Biol. 94:671–675. 2013.
View Article : Google Scholar
|
|
39
|
Schmielau J and Finn OJ: Activated
granulocytes and granulocyte-derived hydrogen peroxide are the
underlying mechanism of suppression of t-cell function in advanced
cancer patients. Cancer Res. 61:4756–4760. 2001.PubMed/NCBI
|
|
40
|
Palmer DC and Restifo NP: Suppressors of
cytokine signaling (SOCS) in T cell differentiation, maturation,
and function. Trends Immunol. 30:592–602. 2009. View Article : Google Scholar : PubMed/NCBI
|