Introduction
Hepato growth factor associated with the metastasis
cellular carcinoma (HCC) is the fifth most common malignancy in the
world, with more than 748,000 new cases diagnosed annually. HCC is
associated with a particularly poor prognosis. HCC ranks as the
second cause of mortality from cancer and is associated with the
shortest survival rate of any cancer (1,2). The
overall poor survival rate of HCC patients is mainly ascribed to
late disease diagnosis, which rules out curative surgery for the
majority of patients (3). The
prognosis for patients eligible for surgical resection remains
dismal, due to the high potential for vascular invasion, metastasis
and recurrence (4). The poor
clinical outcome of HCC patients emphasizes the importance of a
better understanding of the transcriptional activation of oncogenic
signaling pathways and the control of cancer-associated genes.
The SOX family consists of a group of transcription
factors that are defined by a highly conserved high-mobility group
(HMG) DNA-binding domain (5,6).
Knockout experiments demonstrated that SOX genes have a wide range
of roles in development (7).
Recently, a number of links have been found between SOX
transcription factors and human related types of cancer (8). For instance, SOX7 (9–11) and
SOX17 (12,13) act as tumor suppressors in various
types of cancer through suppression of Wnt signaling.
Contrastingly, SOX2 has been reported as a potential oncogene in
breast and lung cancer (14,15).
In the case of SOX18, overexpression of SOX18 has been found in
gastric cancer, as compared to normal gastric tissues (16). Furthermore, SOX18 expression was
found to be correlated with a poor clinical outcome of patients
with ovarian (17), non-small cell
lung cancer (18) or invasive
ductal breast carcinoma (19).
However, little is known concerning the expression pattern and
biological functions of SOX18 in HCC.
In the present study, the SOX18 mRNA level was
significantly increased in HCC tissues. Then, by siRNA-mediated
silencing of SOX18 in HCC cell lines, we found that SOX18 is
involved in the regulation of cell proliferation, cell cycle
progression and metastasis. Furthermore, gene set enrichment
analysis (GSEA) showed that KEGG focal adhesion and chemokine
signaling pathways were correlated with SOX18 expression.
Importantly, we demonstrated that upregulation of SOX18 is
associated with a poor outcome in HCC patients. Collectively, the
present study provides original documentation for the
overexpression of SOX18 in HCC and it may be an effective
therapeutic target for this disease.
Materials and methods
Patients and tissue samples
From 2006 to 2008, 75 patients admitted to Shanxi
Dayi Hospital were enrolled in the present study. In each case, HCC
and adjacent non-tumorous tissues were snap-frozen in liquid
nitrogen and stored at −80°C until RNA extraction. Follow-up was
completed in October 2013. Ethical approval for the present study
was provided by the Independent Ethics Committee of Shanxi Dayi
Hospital. Informed and written consent was obtained from all
patients or their advisers according to the ethics committee
guidelines.
Cell lines
HepG2, BEL-7404, SMC-7721, MHCC-97L and MHCC-97H
cells were obtained from the Cell Bank of Shanghai Biology
Institute, Chinese Academy of Science (Shanghai, China). All
culture media (Life Technologies, Carlsbad, CA, USA) were
supplemented with 10% fetal bovine serum (FBS; Life Technologies),
100 mg/ml penicillin G and 50 µg/ml streptomycin (Life
Technologies). HepG2, BEL-7404, MHCC-97L and MHCC-97H cells were
cultured in DMEM. SMC-7721 cells were cultured in RPMI-1640
medium.
Silencing of SOX18 by small interfering
RNA
The siRNA targeting position 1344–1362
(CUCUCUCAUACGCGUGUAU) of human SOX18 mRNA was synthesized. A
non-specific scramble siRNA sequence was used as the negative
control (NC). The siRNAs were transiently transfected into MHCC-97H
and HepG2 cells using Lipofectamine 2000 (Invitrogen) according to
the manufacturer's instructions. Assays were performed 48 h after
transfection.
Reverse transcription and real-time
PCR
Total RNA was extracted using TRIzol reagent
(Invitrogen) according to the manufacturer's instructions. cDNA was
synthesized by using M-MuLV Reverse Transcriptase (Thermo Fisher
Scientific, Rockford, IL, USA). Real-time PCR was performed using a
standard SYBR-Green PCR kit (Thermo) protocol on an ABI 7300
real-time PCR machine (Applied Biosystems, Foster City, CA, USA).
GAPDH served as an internal control. The gene expression was
calculated using the ∆∆Ct method. All data represent the average of
three replicates. The primers used are listed in Table I.
 | Table IPrimers sequences for the quantitative
PCR. |
Table I
Primers sequences for the quantitative
PCR.
| Primer | Primer sequence | Size (bp) |
|---|
| SOX18 | F:
5′-CGCGTGTATGTTTGGTTC-3′ | |
| (NM_018419.2) | R:
5′-ATGTAACCCTGGCAACTC-3′ | 211 |
| RhoA | F:
5′-GAGTGTTCAGCAAAGACCAAAG-3′ | |
| (NM_001664.2) | R:
5′-TTGCAGCAAGGTTTCACAAG-3′ | 124 |
| IGF1R | F:
5′-GAGCCTCCTGTGAAAGTG-3′ | |
| (NM_000875.4) | R:
5′-GCATCCTGCCCATCATAC-3′ | 175 |
| PDGFB | F:
5′-CTCGATCCGCTCCTTTGATG-3′ | |
| (NM_002608.2) | R:
5′-AGGAAGTTGGCGTTGGTG-3′ | 249 |
| CCL2 | F:
5′-AACCGAGAGGCTGAGACTAAC-3′ | |
| (NM_002982.3) | R:
5′-GGAATGAAGGTGGCTGCTATG-3′ | 125 |
| CCL3 | F:
5′-TTCCGTCACCTGCTCAGAATC-3′ | |
| (NM_002983.2) | R:
5′-TGGCTGCTCGTCTCAAAGTAG-3′ | 188 |
| CCL5 | F:
5′-CCTCGCTGTCATCCTCATTG-3′ | |
| (NM_002985.2) | R:
5′-ACTTGGCGGTTCTTTCGG-3′ | 195 |
| GADPH | F:
5′-CACCCACTCCTCCACCTTTG-3′ | |
|
(NM_001256799.1) | R:
5′-CCACCACCCTGTTGCTGTAG-3′ | 110 |
Western blot analysis
Treated and untreated MHCC-97H and HepG2 cells were
lysed in ice-cold radio immunoprecipitation assay buffer. Equal
amounts of protein were separated via SDS-PAGE gel electrophoresis
and electro-blotted onto a nitrocellulose membrane. Immunodetection
of proteins was performed using specific antibodies. Densitometric
analysis was performed with Image J software. Primary antibodies
were obtained from the following companies: i) SOX18, PDGFB, IGF1R
and CCL3 (Abcam, Cambridge, MA, USA); ii) CCL2, CCL5, RhoA and
GAPDH, CST (Biotech, Danvers, MA, USA).
Cell proliferation assay
Cell proliferation was measured using the Cell
Counting Kit-8 (CCK-8) (Dojindo Lab, Kumamoto, Japan). Briefly, the
treated and untreated MHCC-97H and HepG2 cells were seeded onto
96-well plates. At the indicated time-point, CCK8 solution was
added to each well and incubated for 1 h. The optical density
values (OD) were measured at 450 nm by using a microplate reader
(Bio-Rad Laboratories Inc., Hercules, CA, USA). All experiments
were performed in triplicates and repeated at least three
times.
Cell cycle distribution analysis
Propidium iodide (PI) staining was used to analyze
DNA content. Treated and untreated MHCC-97H and HepG2 cells were
harvested and labeled with PI by using previously described methods
(20). DNA content was analyzed
using a FACScan flow cytometry (BD Biosciences, San Jose, CA, USA)
and the percentages of cells in the G0/G1, S and G2/M phases were
determined with the FlowJo software (Tree Star). Experiments were
performed in triplicate.
Cell apoptosis analysis
The percentage of apototic cells was determined by
double staining with Annexin V-fluorescein isothiocyanate (FITC)
and PI. With or without siRNA transfection for 48 h, both adherent
and floating cells were harvested, washed with PBS, pelleted and
re-suspended in an incubation buffer containing Annexin V-FITC (BD
Biosciences) and PI. The subsequent analysis was performed on a
FACScan flow cytometry.
Boyden chamber assay for migration and
invasion
Quantitative cell migration and invasion assays were
performed using 12-well Boyden chambers (Coring, NY, USA). For the
migration assay, siRNA-transfected cells were serum-starved for 24
h, and then 5×104 cells were seeded into the upper well
of the Boyden chambers, with 500 ml of serum-free medium added to
the lower chamber. After 24 h of incubation, the cells on the upper
surface of the filter were completely removed. The remaining cells
were washed with PBS, fixed in 4% paraformaldehyde and stained with
0.2% crystal violet. The migrated cells were observed under a Leica
inverted microscope (Deerfield, IL, USA) and counted.
For the in vitro invasion assay, the upper
well of the Boyden Chamber was pre-coated with 10 mg/ml Matrigel
(BD Biosciences). The rest of the assay was performed as described
above.
Gene Set Enrichment Analysis (GSEA)
To gain further insight into the biological pathways
involved in HCC pathogenesis through the SOX18 pathway, GESA, a
method of analyzing and interpreting microarray and the data using
biological knowledge (21), was
performed using GSEA version 2.0 from the Broad Institute at MIT,
as previously described (22,23).
RNA-sequencing data of the HCC cohort were downloaded from The
Cancer Genomic Atlas project (TCGA) and analyzed by GSEA. In the
present study, GSEA firstly generated an ordered list of all genes
according to their correlation with SOX18 expression and then a
predefined gene set (signature of gene expression upon perturbation
of certain cancer-related gene) receives an enrichment score (ES),
which is a measure of statistical evidence rejecting the null
hypothesis that its members are randomly distributed in the ordered
list. The expression level of SOX18 was used as phenotype label and
'Metric for ranking genes' was set to Pearson's correlation. The
KEGG gene set biological process database (c2.KEGG. v4.0) from the
Molecular Signatures Database was used for enrichment analysis.
Statistical analysis
Survival curves were obtained by the Kaplan-Meier
method and the differences in survival between low and high SOX18
expression groups were analyzed with the log-rank test. The
two-tailed Student's t-test was used to evaluate statistical
differences between two groups. Statistical significance was set at
P<0.05. Where appropriate, data are expressed as mean ± SD.
Results
Overexpression of SOX18 in HCC tissues is
correlated with reduced survival
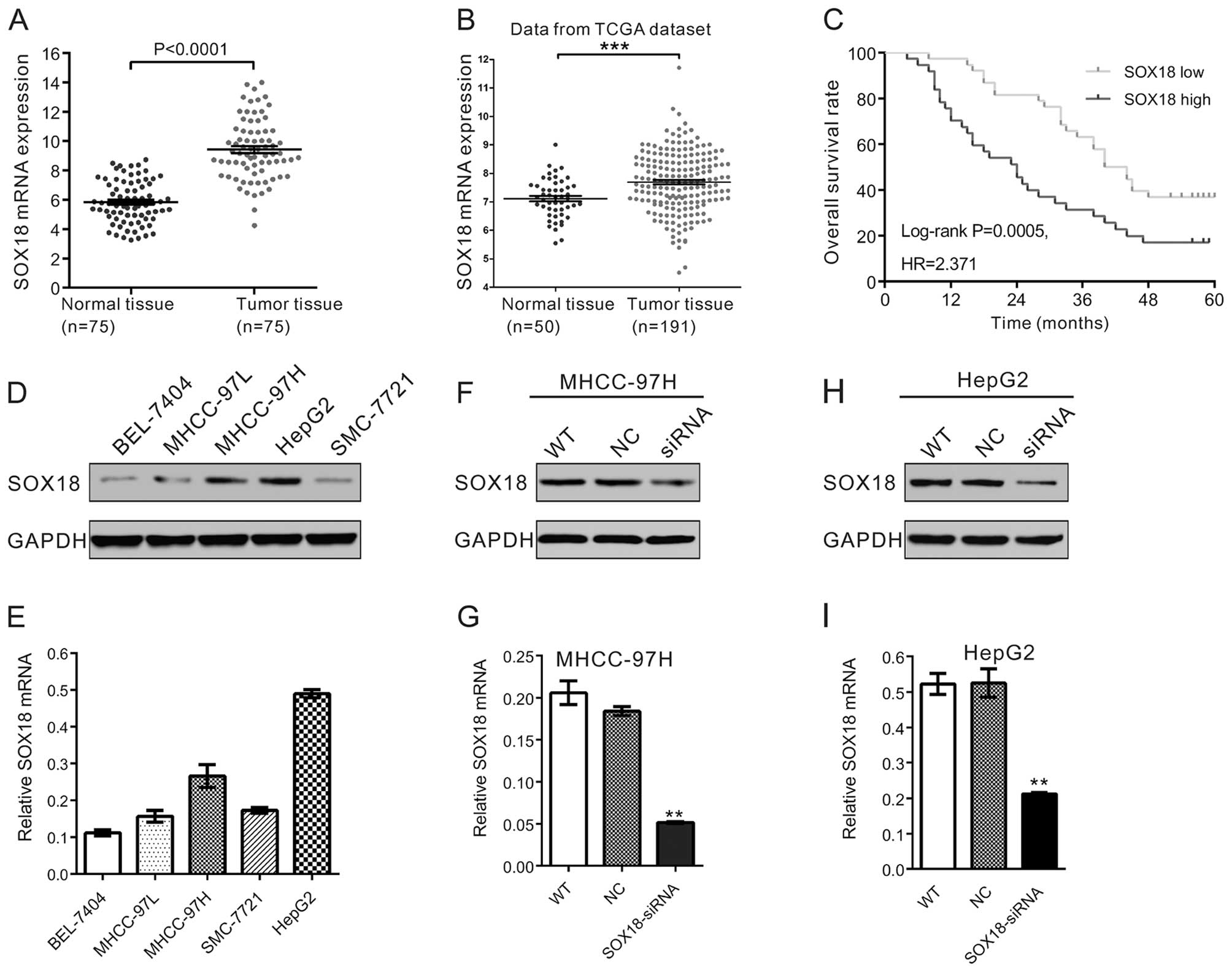
We first measured the SOX18 mRNA level in 75 patient
HCC and adjacent non-tumorous tissues by real-time PCR. As shown in
Fig. 1A, statistical analysis using
the Student's t-test showed that SOX18 mRNA was significantly
overexpressed in the HCC tissues when compared with that in the
normal tissues (P<0.001).
In addition, we re-analyzed high throughput
RNA-sequencing data of the HCC cohort of TCGA and also found a
significant increase in SOX18 expression in the HCC tissues, and
high SOX18 expression was compared with that in the normal tissues
(Fig. 1B).
We next carried out Kaplan-Meier survival analysis
to investigate the clinical outcome of HCC patients with low or
high SOX18 expression. As shown in Fig.
1C, the survival time of the patients with
high-SOX18-expressing tu mors was significantly shorter than that
of patients with low-SOX18-expressing tumors (P<0.01). These
results indicated that SOX18 expression was upregulated in the HCC
tissues and was correlated with poor survival rate of these
patients.
Silencing of SOX 18 by RNA interference
(RNAi)
To investigate the functions of SOX18 overexpression
on HCC, we knocked down its expression in HCC cells by RNAi. We
determined the protein and mRNA levels of SOX18 in five HCC cell
lines, BEL-7404, MHCC-97H, MHCC-97L, HepG2 and SMC-7721, by western
blot analysis and real-time PCR, respectively. Higher protein and
mRNA levels of SOX18 were observed in two cell lines, MHCC-97H and
HepG2 (Fig. 1D and E), which were
selected for the RNAi experiment.
One siRNA targeting human SOX18 (SOX18-siRNA) and a
negative control (NC, a non-specific scramble siRNA) were
synthesized and used to transfect the MHCC-97H and HepG2 cells. The
silencing effect of the siRNA was then evaluated by western blot
analysis and real-time PCR. Our results indicated that SOX18-siRNA
was able to efficiently suppress endogenous SOX18 expression in the
MHCC-97H (Fig. 1F and G) and HepG2
cells (Fig. 1H and I).
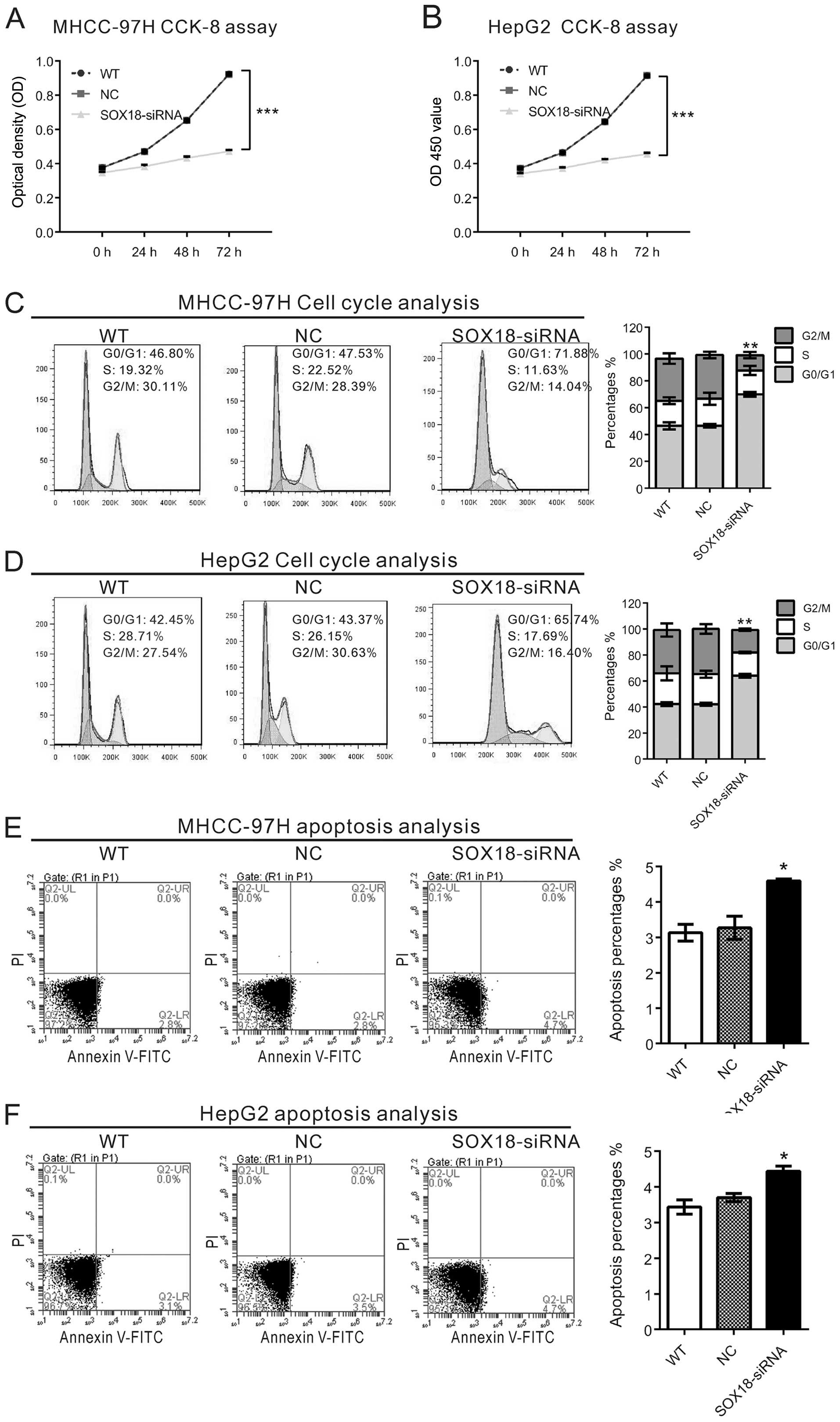
Silencing of SOX18 suppresses the
proliferation, while induces G1-phase arrest and cell apoptosis in
HCC cells
To examine the effects of SOX18 silencing on the
proliferation of HCC cells, the CCK-8 assay was performed. Cell
proliferation of MHCC-97H (Fig. 2A)
and HepG2 cells (Fig. 2B)
transfected with SOX18-siRNA was notably impaired when compared to
cell proliferation in the corresponding WT and NC cells. These
results indicate that SOX18 may promote the proliferation of HCC
cells.
The possible inhibitory effect of SOX18 knockdown on
cell cycle progression was then evaluated. PI staining and flow
cytometric analysis revealed that silencing of SOX18 in the
MHCC-97H (Fig. 2C) and HepG2 cells
(Fig. 2D) caused an increase in
cells in the G1 phase and a corresponding decrease in cells in the
S and G2/M phases. We also assessed the apoptotic function of SOX18
in the HCC cells by Annexin V-FITC/PI staining assay. As shown in
Fig. 2E and F, MHCC-97H and HepG2
cells transfected with SOX18-siRNA exhibited slightly induced cell
apoptosis compared with WT or NC cells. These results imply that
the proliferation-promoting effect of SOX18 is mainly mediated by
promoting cell cycle progression.
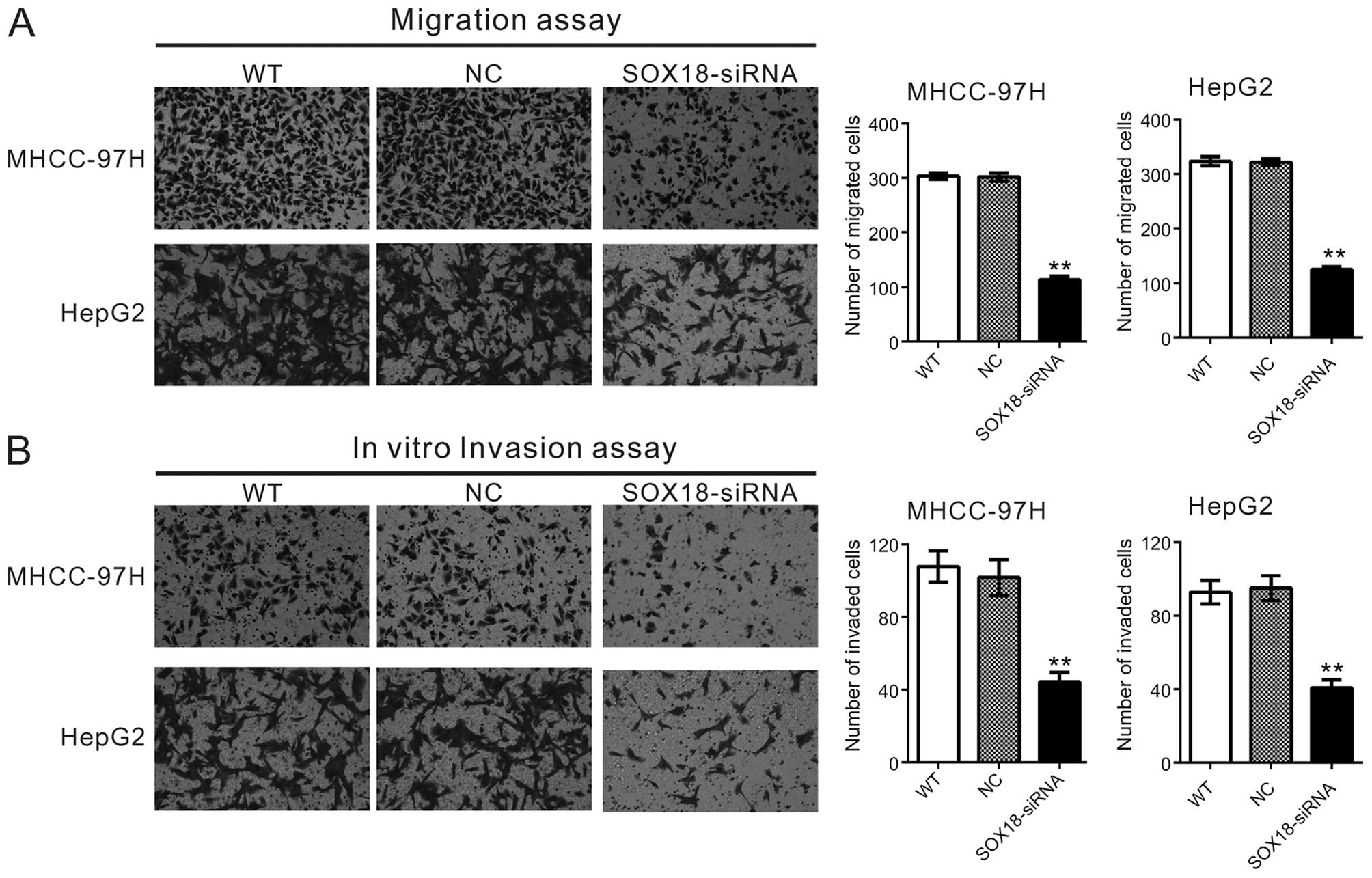
Downregulation of SOX18 inhibits the
motility and invasiveness of HCC cells
To explore the involvement of SOX18 in cell
motility, Transwell assays were carried out to quantitatively
determine the effect of SOX18 on cell migration. As shown in
Fig. 3A, similar numbers of WT and
NC cells migrated to the lower face of the Transwell membrane
(MHCC-97H: WT, 304±6; NC, 301±8; HepG2: WT, 323±8; NC, 322±6),
whereas the SOX18-knockdown cells exhibited a strongly inhibited
motility, with <40% cells migrating (MHCC-97H: 113±6; HepG2:
125±5).
We also investigated whether SOX18 affects the
invasive ability of HCC cells by an in vitro invasion assay.
As shown in Fig. 3B, depletion of
SOX18 dramatically reduced the cell invasive ability when compared
with that of the WT and NC cells. The number of invaded knockdown
cells was ~43% of that of the control cells (MHCC-97H: WT, 108±9;
NC, 102±10; SOX-siRNA, 44±5; HepG2: WT, 93±6; NC, 95±7; SOX-siRNA,
41±4). These data suggest that SOX18 promotes HCC cell
invasion.
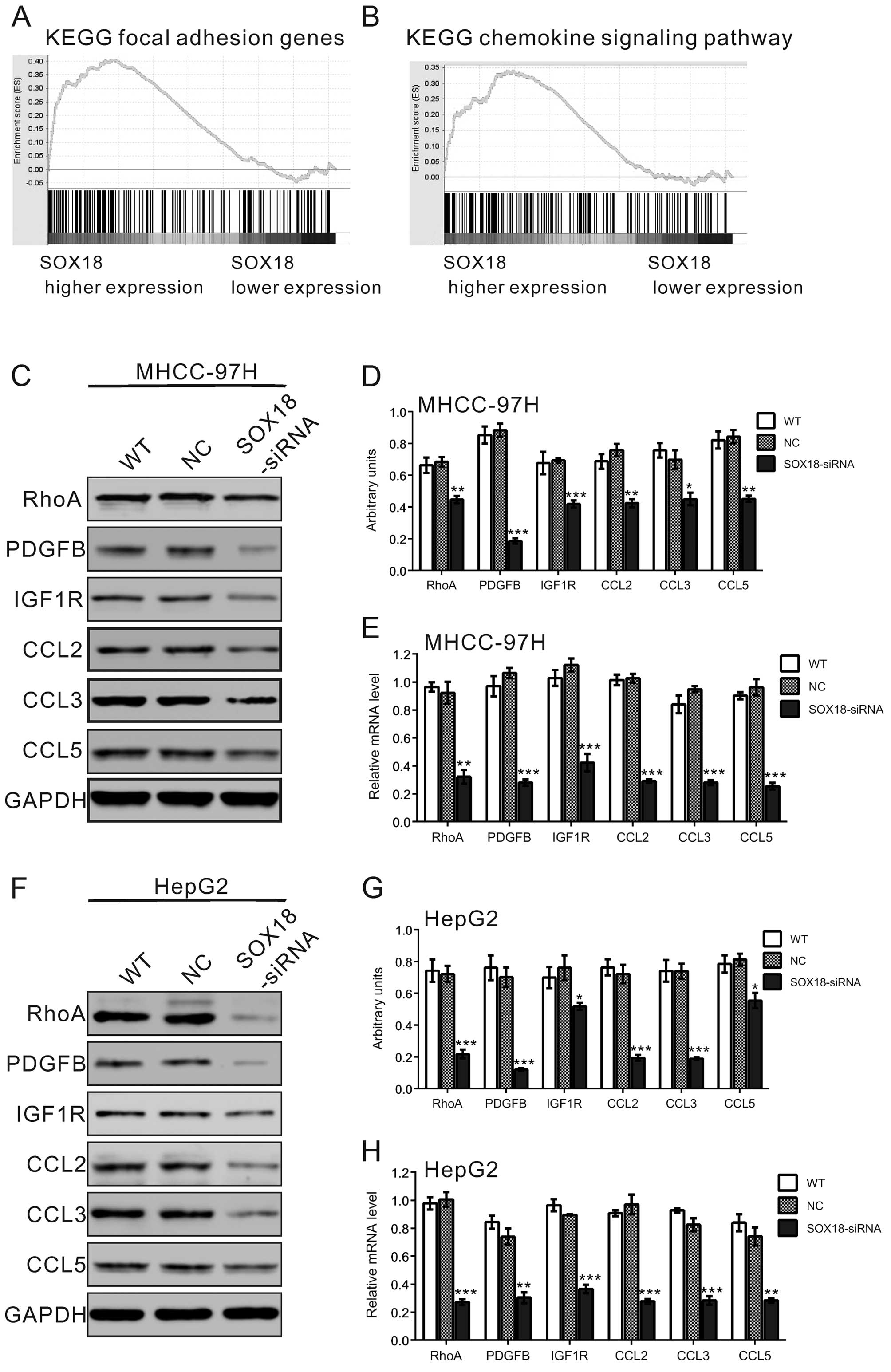
Identification of genes and
signaling-associated biological pathways and processes by GSEA
To probe the SOX18-associated pathways on an
unbiased basis, we performed GSEA using high throughput
RNA-sequencing data of the HCC cohort from The Cancer Genome Atlas
project (TCGA). GSEA is designed to detect coordinated differences
in expression of predefined sets of functionally related genes.
Among all the 188 predefined 'KEGG pathway' gene sets, the focal
adhesion pathway and chemokine signaling pathways were identified
as having a significant association with SOX18 expression in the
HCC dataset (Fig. 4A and B).
SOX18 siRNA regulates the mRNA and
protein expression of RhoA, PDGFB, IGF1R, CCL2, CCL3 and CCL5 in
HCC cells
The effects of SOX18 siRNA on the mRNA and protein
levels of focal adhesion pathway genes (RhoA, PDGFB and IGF1R) and
chemokine signaling pathway genes (CCL2, CCL3 and CCL5) were
investigated. As shown in Fig.
4C–H, SOX18 siRNA treatment in the MHCC-97H and HepG2 cells
significantly decreased the mRNA and protein levels of the detected
genes in comparison with levels in the WT and NC groups.
Discussion
The involvement of SOX genes in various cancers has
been recently confirmed. Since most SOX genes behave as oncogenes
in many human cancers, their targeting has great therapeutic
potential. In the present study, we reported that the elevation of
the SOX18 mRNA level was associated with the poor prognosis of
patients with HCC. The in vitro experiments showed that
knockdown of SOX18 expression inhibited the motile capacity of the
HCC cells. These data indicated the diagnostic and therapeutic
value of SOX18 for HCC.
Several studies (16–19)
have reported the overexpression of SOX18 in gastric, ovarian,
non-small cell lung and breast cancer. In the present study, SOX18
was identified as a potential biomarker for the diagnosis and
prognosis of HCC. SOX18 mRNA was significantly upregulated in the
HCC tissues when compared with adjacent non-tumorous tissues
(Fig. 1A), which was confirmed by
an independent HCC dataset from TCGA (Fig. 1B). More importantly, Kaplan-Meier
survival analysis revealed that a high expression level of SOX18
was associated with a reduced patient survival rate (Fig. 1C).
Previous studies have suggested the promoting effect
of SOX18 on cell proliferation of vascular smooth muscle cells
(24) and MCF-7 breast cancer cells
(25). In line with these findings,
knockdown of SOX18 in the HCC cells significantly impaired cell
growth (Fig. 2). Moreover, flow
cytometric analysis showed that SOX18 knockdown induced G1 phase
arrest and apoptosis of HCC cells (Fig.
2), which may have contributed to the inhibition of
proliferation in the SOX18-knockdown cells. In addition, tumor
metastasis was previously found to be inhibited in SOX18-deficient
mice (26) and dominant-negative
SOX18 shows an inhibitory effect on the migratory ability of MCF-7
cells (25). Consistent with these
studies, we found that SOX18-siRNA treatment significantly
decreased the migration and invasion capabilities of the HCC cells
(Fig. 3), which suggest the role of
SOX18 in promoting the metastasis of HCC.
In order to elucidate the possible mechanism
involved in the SOX18-mediated inhibition of the motility of the
HCC cells, we performed GSEA to identify the associated biological
processes and signaling pathways using high throughput
RNA-sequencing data of the HCC cohort of TCGA (Fig. 4). We found that the KEGG focal
adhesion and chemokine signaling pathways were associated with
SOX18 expression, which indicated that these two pathways play
crucial role in the process of anti-proliferation, anti-migration
and anti-invasion triggered by SOX18-siRNA.
The focal adhesion and chemokine signaling pathways
(27) are involved in various
cellular functions, including cell proliferation, motility,
invasion and mortality. In the present study, the expression of
KEGG focal adhesion genes (RhoA, IGF1R and PDGFB) and KEGG
chemokine signaling genes (CCL2, CCL3 and CCL5) was evaluated in
the SOX18-siRNA-treated HCC cells (Fig.
4). Small GTPase RhoA (28) and
IGF1R (29) have long been
recognized to play an important role in tumorigenesis and tumor
progression. PDGFB is a growth factor associated with the
metastasis of various types of human cancer (30). A growing body of research suggests
the tumor-promoting roles of CCL2 (31,32),
CCL3 (33) and CCL5 (32,34).
Our data showed that SOX18 RNAi significantly downregulated the
expression of detected genes, which indicating that SOX18 may
execute its functions through regulating the expression of focal
adhesion and chemokine signaling genes.
In conclusion, the present study proved for the
first time that SOX18 plays a key role in the proliferation and
metastasis of HCC cells. Moreover, SOX18 may regulate these
biological processes through the focal adhesion and chemokine
signaling pathways, thus providing useful information for the
targeted therapy of HCC. As the SOX18 expression level is
associated with patient survival, inhibition of SOX18 in tumor
tissues may provide an effective therapeutic strategy.
Reference
|
1
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar
|
|
2
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pang RW and Poon RT: From molecular
biology to targeted therapies for hepatocellular carcinoma: The
future is now. Oncology. 72:30–44. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Poon RTP, Fan ST and Wong J: Risk factors,
prevention, and management of postoperative recurrence after
resection of hepatocellular carcinoma. Ann Surg. 232:102000.
View Article : Google Scholar
|
|
5
|
Wilson M and Koopman P: Matching SOX:
Partner proteins and co-factors of the SOX family of
transcriptional regulators. Curr Opin Genet Dev. 12:441–446. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bowles J, Schepers G and Koopman P:
Phylogeny of the SOX family of developmental transcription factors
based on sequence and structural indicators. Dev Biol. 227:239–255.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schepers GE, Teasdale RD and Koopman P:
Twenty pairs of sox: Extent, homology, and nomenclature of the
mouse and human sox transcription factor gene families. Dev Cell.
3:167–170. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dong C, Wilhelm D and Koopman P: Sox genes
and cancer. Cytogenet Genome Res. 105:442–447. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li B, Ge Z, Song S, Zhang S, Yan H, Huang
B and Zhang Y: Decreased expression of SOX7 is correlated with poor
prognosis in lung adenocarcinoma patients. Pathol Oncol Res.
18:1039–1045. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Stovall DB, Wan M, Miller LD, Cao P,
Maglic D, Zhang Q, Stampfer MR, Liu W, Xu J and Sui G: The
regulation of SOX7 and its tumor suppressive role in breast cancer.
Am J Pathol. 183:1645–1653. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang Y, Huang S, Dong W, Li L, Feng Y,
Pan L, Han Z, Wang X, Ren G, Su D, et al: SOX7, down-regulated in
colorectal cancer, induces apoptosis and inhibits proliferation of
colorectal cancer cells. Cancer Lett. 277:29–37. 2009. View Article : Google Scholar
|
|
12
|
Ye YW, Wu JH, Wang CM, Zhou Y, Du CY,
Zheng BQ, Cao X, Zhou XY, Sun MH and Shi YQ: Sox17 regulates
proliferation and cell cycle during gastric cancer progression.
Cancer Lett. 307:124–131. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Du YC, Oshima H, Oguma K, Kitamura T,
Itadani H, Fujimura T, Piao YS, Yoshimoto T, Minamoto T, Kotani H,
et al: Induction and down-regulation of Sox17 and its possible
roles during the course of gastrointestinal tumorigenesis.
Gastroenterology. 137:1346–1357. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen Y, Shi L, Zhang L, Li R, Liang J, Yu
W, Sun L, Yang X, Wang Y, Zhang Y, et al: The molecular mechanism
governing the oncogenic potential of SOX2 in breast cancer. J Biol
Chem. 283:17969–17978. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rudin CM, Durinck S, Stawiski EW, Poirier
JT, Modrusan Z, Shames DS, Bergbower EA, Guan Y, Shin J, Guillory
J, et al: Comprehensive genomic analysis identifies SOX2 as a
frequently amplified gene in small-cell lung cancer. Nat Genet.
44:1111–1116. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Eom BW, Jo MJ, Kook MC, Ryu KW, Choi IJ,
Nam BH, Kim YW and Lee JH: The lymphangiogenic factor SOX 18: A key
indicator to stage gastric tumor progression. Int J Cancer.
131:41–48. 2012. View Article : Google Scholar
|
|
17
|
Pula B, Kobierzycki C, Solinski D,
Olbromski M, NowakMarkwitz E, Spaczynski M, Kedzia W, Zabel M and
Dziegiel P: SOX18 expression predicts response to platinum-based
chemotherapy in ovarian cancer. Anticancer Res. 34:4029–4037.
2014.PubMed/NCBI
|
|
18
|
Jethon A, Pula B, Olbromski M, Werynska B,
Muszczynska-Bernhard B, Witkiewicz W, Dziegiel P and
Podhorska-Okolow M: Prognostic significance of SOX18 expression in
non-small cell lung cancer. Int J Oncol. 46:123–132. 2015.
|
|
19
|
Pula B, Olbromski M, Wojnar A,
Gomulkiewicz A, Witkiewicz W, Ugorski M, Dziegiel P and
Podhorska-Okolow M: Impact of SOX18 expression in cancer cells and
vessels on the outcome of invasive ductal breast carcinoma. Cell
Oncol (Dordr). 36:469–483. 2013. View Article : Google Scholar
|
|
20
|
Papavasiliou FN and Schatz DG:
Cell-cycle-regulated DNA double-strand breaks in somatic
hypermutation of immunoglobulin genes. Nature. 408:216–221. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Subramanian A, Kuehn H, Gould J, Tamayo P
and Mesirov JP: GSEA-P: A desktop application for Gene Set
Enrichment Analysis. Bioinformatics. 23:3251–3253. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen H, Xu J, Hong J, Tang R, Zhang X and
Fang J-Y: Long noncoding RNA profiles identify five distinct
molecular subtypes of colorectal cancer with clinical relevance.
Mol Oncol. 8:1393–1403. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kapoor A, Yao W, Ying H, Hua S, Liewen A,
Wang Q, Zhong Y, Wu CJ, Sadanandam A and Hu B: Yap1 activation
enables bypass of oncogenic Kras addiction in pancreatic cancer.
Cell. 158:185–197. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Garcia-Ramirez M, Martínez-González J,
Juan-Babot JO, Rodriguez C and Badimon L: Transcription factor
SOX18 is expressed in human coronary atherosclerotic lesions and
regulates DNA synthesis and vascular cell growth. Arterioscler
Thromb Vasc Biol. 25:2398–2403. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Young N, Hahn CN, Poh A, Dong C, Wilhelm
D, Olsson J, Muscat GE, Parsons P, Gamble JR and Koopman P: Effect
of disrupted SOX18 transcription factor function on tumor growth,
vascularization, and endothelial development. J Natl Cancer Inst.
98:1060–1067. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Duong T, Proulx ST, Luciani P, Leroux JC,
Detmar M, Koopman P and Francois M: Genetic ablation of SOX18
function suppresses tumor lymphangiogenesis and metastasis of
melanoma in mice. Cancer Res. 72:3105–3114. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Locati M, Deuschle U, Massardi ML,
Martinez FO, Sironi M, Sozzani S, Bartfai T and Mantovani A:
Analysis of the gene expression profile activated by the CC
chemokine ligand 5/RANTES and by lipopolysaccharide in human
monocytes. J Immunol. 168:3557–3562. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Struckhoff AP, Rana MK and Worthylake RA:
RhoA can lead the way in tumor cell invasion and metastasis. Front
Biosci. 16:1915–1926. 2011. View
Article : Google Scholar
|
|
29
|
Adams TE, McKern NM and Ward CW:
Signalling by the type 1 insulin-like growth factor receptor:
Interplay with the epidermal growth factor receptor. Growth
Factors. 22:89–95. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jechlinger M, Sommer A, Moriggl R, Seither
P, Kraut N, Capodiecci P, Donovan M, Cordon-Cardo C, Beug H and
Grünert S: Autocrine PDGFR signaling promotes mammary cancer
metastasis. J Clin Invest. 116:1561–1570. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang J, Patel L and Pienta KJ: CC
chemokine ligand 2 (CCL2) promotes prostate cancer tumorigenesis
and metastasis. Cytokine Growth Factor Rev. 21:41–48. 2010.
View Article : Google Scholar :
|
|
32
|
Soria G and Ben-Baruch A: The inflammatory
chemokines CCL2 and CCL5 in breast cancer. Cancer Lett.
267:271–285. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yang X, Lu P, Fujii C, Nakamoto Y, Gao JL,
Kaneko S, Murphy PM and Mukaida N: Essential contribution of a
chemokine, CCL3, and its receptor, CCR1, to hepatocellular
carcinoma progression. Int J Cancer. 118:1869–1876. 2006.
View Article : Google Scholar
|
|
34
|
Luboshits G, Shina S, Kaplan O, Engelberg
S, Nass D, Lifshitz-Mercer B, Chaitchik S, Keydar I and Ben-Baruch
A: Elevated expression of the CC chemokine regulated on activation,
normal T cell expressed and secreted (RANTES) in advanced breast
carcinoma. Cancer Res. 59:4681–4687. 1999.PubMed/NCBI
|