Introduction
Breast cancer, the second leading cause of
cancer-related mortality among women, has become an increasingly
important public health issue worldwide. It threatens the health of
women all over the world (1). For
example, ~230,480 new cases of invasive breast cancer and 39,520
breast cancer deaths were estimated to occur among US women in
2011, accounting for nearly 1 in 3 cancers diagnosed among women in
the US. The biological features of breast cancer are still not well
understood, however, much has changed in the management of breast
cancer particularly over the last few decades. Several effective
treatments have been explored in dealing with different types of
advanced breast cancer using newly developed targeted therapy, such
as endocrine therapy and trastuzumab treatment in HER-2+
metastatic breast cancer. Therefore, together, oncologists and
clinicians are working diligently to discover more target molecules
of breast cancer and are aimed at developing more individual-based
strategies for breast cancer patients. Consequently, there is a
growing list of such targeting molecules in breast cancers. The
Hedgehog (Hh) signaling pathway has recently been recognized as one
of the most important signaling pathways and a therapeutic target
in cancer.
The Hh signaling pathway has three ligands: sonic
(SHH), Indian (IHH) and desert (DHH) hedgehogs. When the ligands
bind to their Patched (PTC), a 12-transmembrane receptor, the
inhibition of PTC on Smoothened (SMO), a 7-transmembrane protein,
is removed (2,3). Therefore, the signaling pathway is
active, resulting in the activation of the zinc finger
transcription factors Gli1, Gli2 and Gli3, where Gli1 and Gli2
represent the main activators of Hh target genes and Gli3 acts
mostly as a repressor. The exact mechanism of signal transduction
from Smo to the Gli proteins is not yet clear; however, it has been
proposed that regulating Gli transcription factors involves a
multi-protein complex consisting of intraflagellar transport
proteins, protein kinase A, casein kinase, glycogen synthase kinase
3, suppressor of fused and others. Constitutive activation of the
Hh pathway leading to tumorigenesis has been noted in basal cell
carcinomas, medulloblastoma, gastrointestinal tumors and lung and
prostate cancers (4). Breast cancer
is also associated with the aberrant activation of the Hh signaling
pathway (5). A high-resolution
comparative genomic hybridization analysis of breast cancer samples
revealed a frequent loss of PTCH1 (9q22.1-q31) and amplification of
Gli1 (12q13.2-q13.3) chromosomal regions, indicating that Hh
signaling is involved in breast cancer (6,7).
Evidence has shown that expression of SHH, DHH and Gli1 is higher
in various breast cancer cell lines and in 30% of cancer samples
compared with normal mammary epithelial cells or epithelial tissue
(8,9). More importantly, active human SMO
(SmoM2) in the mammary epithelium as well as overexpression of Gli2
both resulted in ductal hyperplasia (10). Furthermore, a genome-wide RNAi
screening demonstrated that Gli2 is necessary for the growth of
breast cancer cell lines and cyclopamine treatment could induce
apoptosis of breast cancer cell lines with elevated SHH, DHH and
Gli1 protein expression (8,9,11).
Although, the data postulating the involvement of this pathway in
human breast cancer are still limited, increasing evidence implies
that the Hh signaling pathway is potentially a useful target in
multi-targeted breast cancer therapy. Modulating its activity may
affect cancer progression and metastasis.
Rab23 is also known to play a key role in negatively
regulating Hh/Gli signaling, since targeted disruption of the
murine Rab23 gene leads to neural tube defects, resembling the
phenotype caused by an excess of Hh signaling, and mutation of the
Rab23 gene causes abnormalities of multiple organs in patients of
two congenital diseases - Carpenter and Gorlin syndromes. Since
over-activation of the Hh signaling pathway is oncogeneic in human
cancers, Rab23, a negative regulator of the Hh signaling pathway,
has been postulated to be a suppressor gene in carcinogenesis. The
abnormal expression of Rab23 has been observed in hepatocellular as
well as thyroid carcinoma (12).
The overexpression of Rab23 has been reported in atrophic
gastritis, a pre-stage of gastric cancer (13). Atrophic gastritis may increase the
risk of gastric cancer, and these findings raise the possibility
that Rab23 upregulation may be involved at an early stage of
gastric carcinogenesis.
To date there have been no studies concerning Rab23
expression in breast cancer. Since Rab23 is a negative regulator of
Hh signaling which can induce malignant carcinoma, upregulation of
Rab23 may be helpful in the treatment of breast cancer. If this is
true, we may be able to identify a new target for the diagnosis and
treatment of breast cancer. In the present study, we demonstrated
that Rab23 was expressed in MCF-7 and MDA-MB-231 cell lines, and
Rab23 activity inhibited breast cancer cell proliferation and DNA
synthesis, and induced apoptosis and downregulated Gli1 and Gli2
expression, which is similar to the effects of cyclopamine on
breast cancer cells in vitro.
Materials and methods
Cell culture
Breast cancer cell lines, MDA-MB-231, MCF-7 and
Bcap37, were obtained from the Cell Resource Center of Shanghai
Institute for Biological Sciences (Chinese Academy of Sciences,
Shanghai, China). They were maintained in RPMI-1640, supplemented
with 10% heat-inactivated (56°C for 30 min) newborn calf serum
(both from Hyclone, Logan, UT, USA) and antibiotics (100 U/ml
penicillin and 100 µg/ml streptomycin). Cells were cultured
at 37°C in a humidified incubator containing 95% air and 5%
CO2. Cells were seeded into 96-, 24- and 6-well plates
or a culture bottle, and serum-starved 24 h before cell
transfection.
Immunofluorescence analysis
Cells (2×105/well) were incubated on
24-well plates (Corning Costar, Cambridge, MA, USA) at 37°C which
were covered with coverslips (1×1 cm). When the cells grew to
occupy ~80% of the coverslips, the cells were fixed in 4%
paraformaldehyde for 30 min at room temperature. After that,
endogenous peroxidase activity was blocked by using 3%
H2O2 in methanol for 30 min at room
temperature. After blocking with 5% normal goat serum containing 3%
BSA in phosphate-buffered saline (PBS) for 1 h at 37°C, the cells
were incubated with the primary antibody, rabbit anti-Rab23 (1:100,
sc-20687; Santa Cruz Biotechnology, Santa Cruz, CA, USA), in a
moist chamber at 4°C overnight. After washing three times with PBS,
the coverslips were incubated with secondary antibodies (1:200,
FITC-conjugated goat anti-rabbit antibodies; Sigma-Aldrich, St.
Louis, MO, USA) for 1 h at 37°C in the dark. The coverslips were
lightly counterstained with Hoechst (1:100; Sigma-Aldrich).
Coverslips were read in a fluorescence microscope at x400
magnification (Nikon, Japan). As a control for specificity of the
primary antibody, coverslips were incubated with normal goat serum
instead of the specific antibody during the first incubation. In
such control sections, coverslip immunofluorescence was not
observed.
Semi-quantitative reverse
transcription-PCR (RT-PCR)
Total RNA was extracted using TRIzol reagent
(Invitrogen, Carlsbad, CA, USA) according to the manufacturer's
instructions. Complementary DNA (cDNA) was synthesized from 5
µg of total RNA with random primers following the
manufacturer's instructions (MBI Fermantas, Vilnius, Lithuania).
Semi-quantitative reverse transcribed-PCR (RT-PCR) was performed in
two steps. First, 5 µg RNA extraction of total RNA was
reverse transcribed with 1 µl reverse transcriptase
(AMV-RT), 2 µl deoxynucleotide triphosphates (dNTPs), 0.5
µl RNase inhibitor, 4 µl MgCl2, 2
µl Rev-Trans, 1 µl primer oligo(dT), and 4.5
µl nuclease-free water for 1 h at 47°C according to the
manufacturer's instructions (Promega, Madison, WI, USA). Second,
PCR amplification was performed with 0.25 µl AmpliTaq DNA
polymerase (Takara, Japan), 1.0 µl each primer, 3 µl
MgCl2 and 10 mM dNTPs in a thermocycler (Bio-Rad,
Hercules, CA, USA). Sequences of the primers used in this study are
listed on Table I. Each RT-PCR
product (10 µl) was analyzed by electrophoresis through a 2%
agarose gel. PCR amplification signals were quantified by
densitometric scanning using Quantity One (Bio-Rad). For primers,
see Table I.
 | Table IRT-PCR primers for analysis. |
Table I
RT-PCR primers for analysis.
| Gene name | Forward primer | Reverse primer |
|---|
| Rab23 | AGG CAC TGG CAA AAA
GGT TA | TA GAC CAC CTT CAG
TGA GGC |
| Gli1 |
CGGGGTCTCAAACTGCCCAGCTT |
GGCTGGGTCACTGGCCCTC |
| Gli2 |
CTAGCATCAGCGAGAACGTG |
AAAGCCTAACTGGCATCCTC |
| GAPDH | ACC ACA GTC CAT GCC
ATC AC | TCC ACC ACC CTG TTG
CTG TA |
Protein extraction and western
blotting
The cells were washed with cold PBS and lysed by the
addition of a lysis buffer containing 1% Nonidet P-40, 50 mM Tris
(pH 7.5), 150 mM NaCl, 0.1% SDS and protease inhibitor cocktail
(Boehringer Mannhein, Lewes, UK) for 20 min at 4°C. Insoluble
materials were removed by centrifugation at 15,000 rpm for 15 min
at 4°C. The supernatant was saved and the protein concentration was
determined using a Bio-Rad protein assay kit (Bio-Rad). Cell
extracts (50 µg/lane) were separated via 10% gel
electrophoresis and electroblotted onto PVDF membranes. Nonspecific
binding sites were blocked by incubating the nitrocellulose sheets
for 1 h in PBS containing 5% low-fat dry milk. Membranes were
probed with the primary anti-Rab23 antibody (1:1,000; Cell
Signaling Technology, Inc., Danvers, MA, USA) overnight at 4°C,
followed by the horseradish peroxidase-conjugated secondary
antibody. Blots were developed using an enhanced chemiluminescence
detection system (ECL; Amersham Pharmacia Biotech) according to the
manufacturer's instructions.
Rab23 gene silencing
In order to knock down protein expression, we
administered specific small interfering RNA (siRNA) against Rab23
with specific siRNA for green fluorescent protein (GFP) as the
control. The following siRNA sequences were applied (Shanghai
GenePharma Co., Ltd., Shanghai, China): siRNA sequence (siRNA
duplex), 5′-CUA CAG AAC AUC AGU GAA ATT-3′ (sense) and 5′-UUU CAC
UGA UGU UCU GUA GTT-3′ (antisense); sequence of the negative
control siRNA, 5′-UUC UCC GAA CGU GUC ACG UTT-3′ (sense) and 5′-ACG
UGA CAC GUU CGG AGA ATT-3′ (anti-sense). Expression levels were
analyzed 48 h after transfection via RT-PCR and
immunofluorescence.
siRNA and cDNA transfection
Human Rab23 and Gli1 expression vectors were kindly
provided by Dr J. Xie (Wells Center for Pediatric Research,
Department of Pediatrics, The Indiana University Simon Cancer
Center, Indiana University, USA). Cells were seeded into a 24- and
a 6-well plate at a density of 2.5×104 cells/well and
incubated for 24 h. Then the cells were transfected with either
Rab23 and Gli1 cDNA or specific Rab23 siRNA by Lipofectamine™ 2000
transfection reagent in Opti-MEM medium (both from Invitrogen)
according to the manufacturer's instructions. All transfection
experiments were carried out in MDA-MB-231 and MCF-7 cells, and
repeated at least 3 times using 2 sets of plasmids prepared
separately. After 48 h of transfection, relative cell
proliferation, cell cycle distribution and apoptosis were
determined by measuring absorbance using bromodeoxyuridine (BrdU)
and flow cytometry (FCM) assays. Each assay was carried out in
triplicate, and normalized relative to the control-transfected
cells.
Incorporation efficiency of BrdU
Cells (1×105/well) were incubated on
24-well plates with coverslips (1×1 cm) at 37°C, after transfection
with Rab23 or siRNA. When the cells occupied ~80% of the
coverslips, the cells were incubated in RPMI-1640 medium
supplemented with 10−5 µmol/l BrdU (Zhongshan
Goldenbridge, Beijing, China) for 15 min at 37°C in a humidified
incubator. The cells were fixed in 4% paraformaldehyde for 30 min
at room temperature. After being blocked with 5% normal goat serum
containing 3% BSA in PBS for 1 h at 37°C, the primary antibody BrdU
(1:100; Santa Cruz Biotechnology) in blocking solution was
incubated with the cells at 4°C overnight. After washing with PBS,
the coverslips were then incubated with the secondary antibody
(1:200; FITC-conjugated goat anti-rabbit antibody) for 1 h at 37°C
in the dark. The slides were lightly counterstained with Hoechst
(1:100; Sigma-Aldrich). Detection of the protein was visualized
using fluorescence microscopy (Nikon, Japan). We randomly chose 5
fields visualized in every coverslip under fluorescence microscopy.
Incorporation efficiency of BrdU in the tumor cells = (the number
of BrdU-positive cells/the number of cells) × 100%.
3-(4,5-Dimethylthiazol-2-y1)-2,5-diphenyltetrazolium bromide (MTT)
assay
The cell viability was determined by MTT assay.
After transfection with the plasmids of Rab23 or siRNA into the
MCF-7 and MDA-MB-231 cells for 24 h, the cells
(5×103/well) were digested and seeded in 96-well
microtiter plates (Costar, Denmark). When cells were incubated for
1, 3 or 5 days, an aliquot (50 µl) of MTT solution (5 mg/ml
in PBS; Sigma-Aldrich) was added to each well, and the plates were
incubated for an additional 4 h at 37°C. MTT solution was aspirated
off, and 150 µl dimethyl sulfoxide (DMSO) was added to each
well to dissolve the formazan crystals formed in the viable cells
for 10 min. The absorbance was read at 490 nm on a Dias automatic
microwell plate reader with DMSO as the blank. Each time point was
performed in 3 wells.
Colony formation assay
MDA-MB-231 and MCF-7 cells (2×104/well)
were incubated on 6-well plates at 37°C. After incubation for 24 h,
the cells were transfected with the plasmids of Rab23 or Gli1.
Twenty-four hours later, the cells were screened by G418 (400
µg/ml; Zhongshan Goldenbridge) for 10 days.
Hexamethylpararosaniline (1% in PBS) was used to stain the
colonies. The colonies with >50 cells were counted by inverted
microscopy. Colony-forming efficiency of the tumor cells = (the
number of colonies/the number of seeded cells) × 100%.
Cell cycle analysis by FCM
MCF-7 and MDA-MB-231 cells were seeded into 6-well
plates and transfected either with Rab23 or siRNA as described
above for 48 h. Then the cells were harvested, washed in PBS, fixed
with 70% ethanol at 4°C for 24 h, followed by DNA staining with
propidium iodide (PI, 1 µg/ml; Sigma-Aldrich) to analyze
cellular DNA content. Cell cycle analysis was performed on a flow
cytometer (Beckman Coulter Inc., Miami, FL, USA).
Apoptosis assay
The apoptotic status of MCF-7 and MDA-MBA-231 cells
was evaluated by measuring the exposure of phosphatidylserine on
the cell membrane using Annexin V-fluorescein isothiocyanate
(Annexin V-FITC) and PI staining by FCM. Breast cancer cells were
plated in 6-well plates. When the cells occupied ~70% of the wells,
the cells were transfected with the plasmids of Rab23. After 24 h
of incubation, the cells were digested and harvested. After
centrifugation, the cell pellets were washed twice with PBS at 4°C.
The cells were incubated with 5 µl Annexin V-FITC and 10
µl PI at room temperature for 15 min in the dark. After the
incubation, 250 µl of 1X binding buffer was added to each
tube. The cells were analyzed immediately by FCM (Beckman
Coulter).
Statistical analysis
All values are expressed as the means ± SEM. The
significance of differences among groups was evaluated by a
Student's t-test for unpaired data or Dunnett's t-test for multiple
comparisons preceded by one-way analysis of variance (GraphPad
Prism 5.0). For all tests, a P-value of <0.05 was considered to
indicate statistical significance.
Results
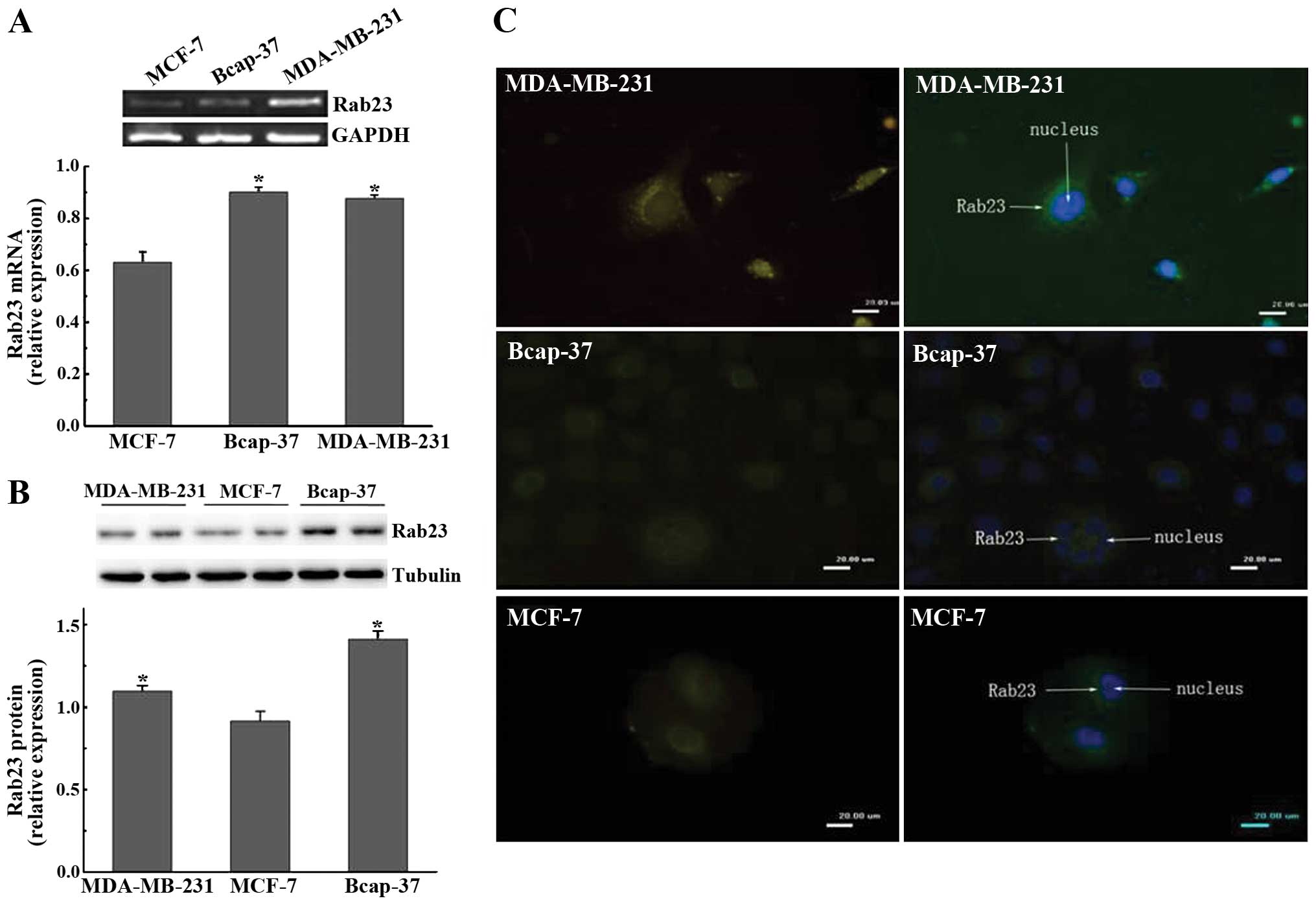
Rab23 is expressed in the breast cancer
cell lines
We selected breast cancer cell lines MDA-MB-23
Bcap37 and MCF-7, and determined whether Rab23 is expressed in
these cell lines. We found that Rab23 mRNA and protein were
expressed in all three cell lines. Expression of Rab23 mRNA and
protein in the MDA-MB-231 and Bcap-37 cells was higher than that in
the MCF-7 cells (P<0.05) (Fig. 1A
and B). From the immunofluorescence results, we found that
Rab23 protein was expressed in the three breast cancer cell lines.
However, its pattern of expression was different. Rab23 protein
fluorescence was observed in the cytoplasm of the MDA-MB-231 cells
but was observed around the cell nucleus in the Bcap-37 and MCF-7
cells (Fig. 1C). Since both Bcap37
and MDA-MB-231 cells are ER-negative, and the expression pattern
had no significant difference, we chose MDA-MB-231 cells to perform
the subsequent experiments.
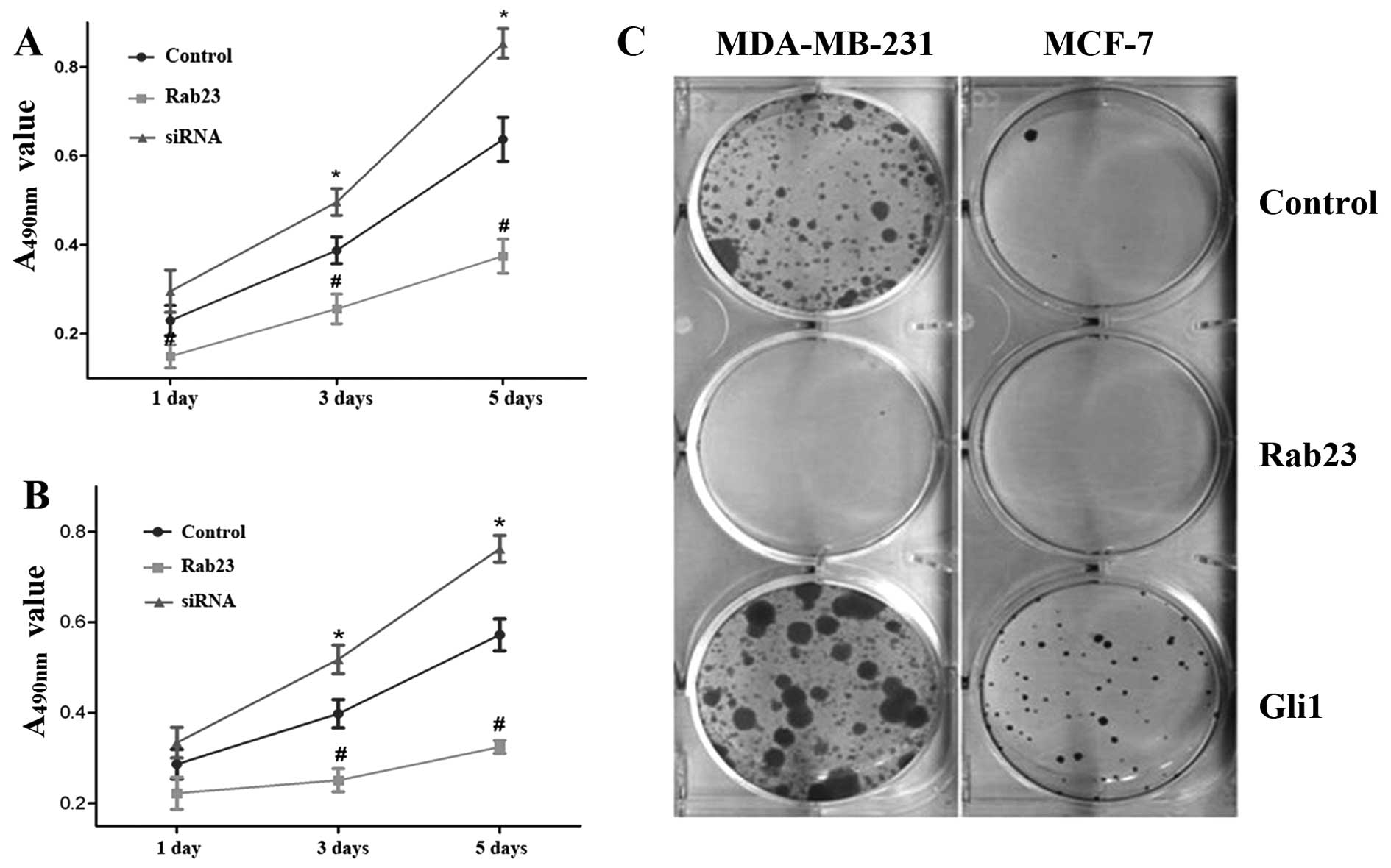
Rab23 inhibits breast cancer cell
viability and proliferation
The cells were transfected with Rab23 siRNA for 48
h. siRNA treatment significantly inhibited Rab23 mRNA expression
compared with the control group in the MDA-MB-231 and MCF-7 cell
lines. The expression of Rab23 (Rab23/GAPDH ratio) decreased from
0.8453±0.05252 to 0.3410±0.06802 and from 0.861±0.05252 to
0.3821±0.06802, respectively, in the MDA-MB-231 and MCF-7 cell
lines. Then we used MTT and colony formation assays to detect the
effect of Rab23 on breast cancer cell viability and proliferation.
MTT assay indicated that the absorbance value at A490 nm
in the Rab23-transfected group was reduced compared with the
control group (P<0.05). However, the absorbance value in the
siRNA group increased (P<0.05) (Fig.
2A and B) after incubation for 1, 3 and 5 days. In the colony
formation assay, in comparison with the control group, the numbers
of colonies formed in the MDA-MB-231 and MCF-7 cell lines
transfected with Rab23 were significantly reduced, while that of
the Gli1-transfected group was increased (Fig. 2C). In the MCF-7 cells, the number of
colonies formed was decreased from 18.33±3.06 to 4.0±2.0 in the
Rab23-transfected cells and was increased from 18.33±3.06 to
74.33±9.71 in the Gli1-transfected cells. In the MDA-MB-231 cells,
the number of colonies changed from 124.7±24.0 to 7.33±3.51 and
124.7±24.0 to 242.3±46.4, respectively (P<0.05). These data
indicate that Rab23 inhibits the growth of breast cancer cells.
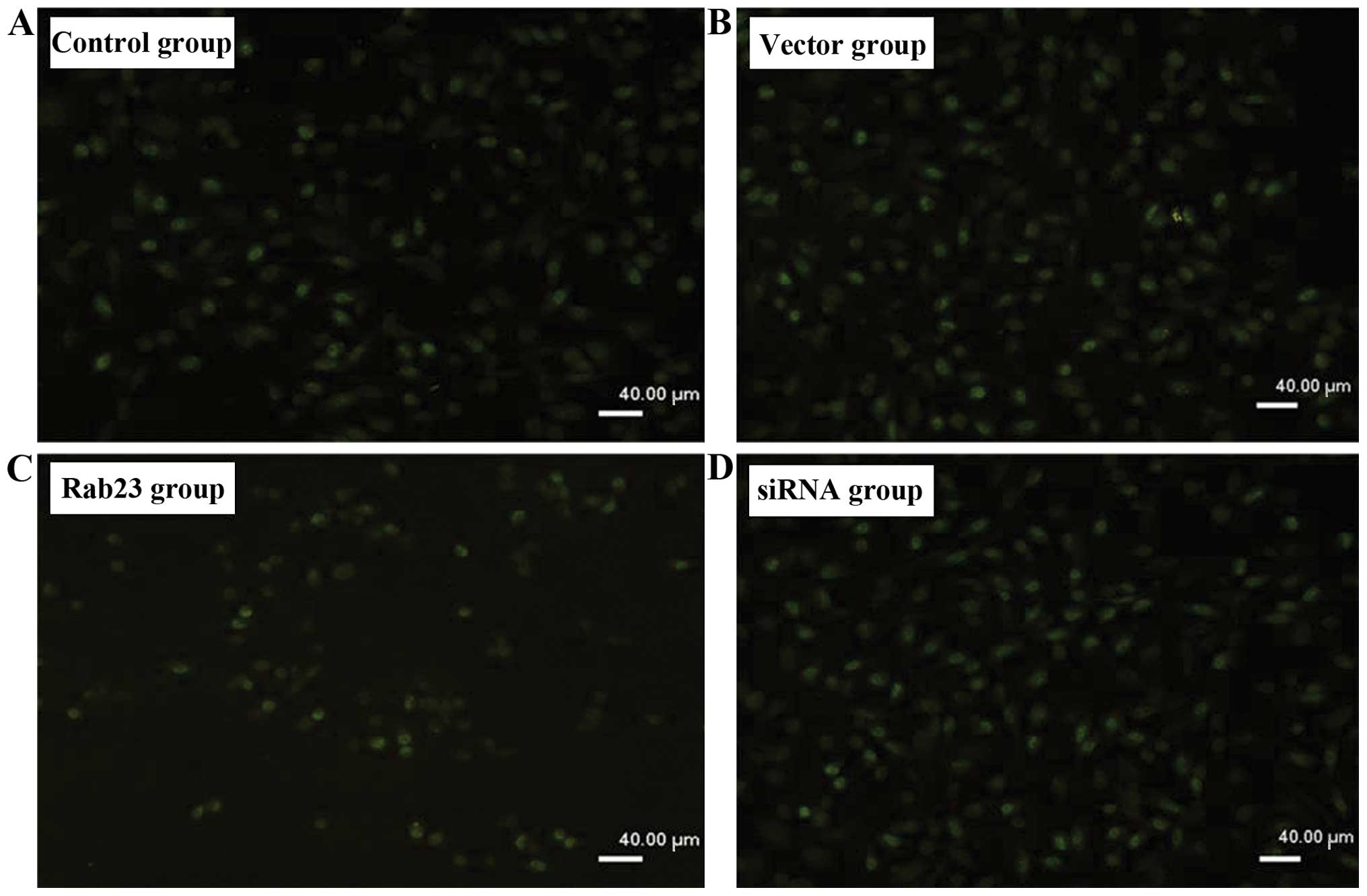
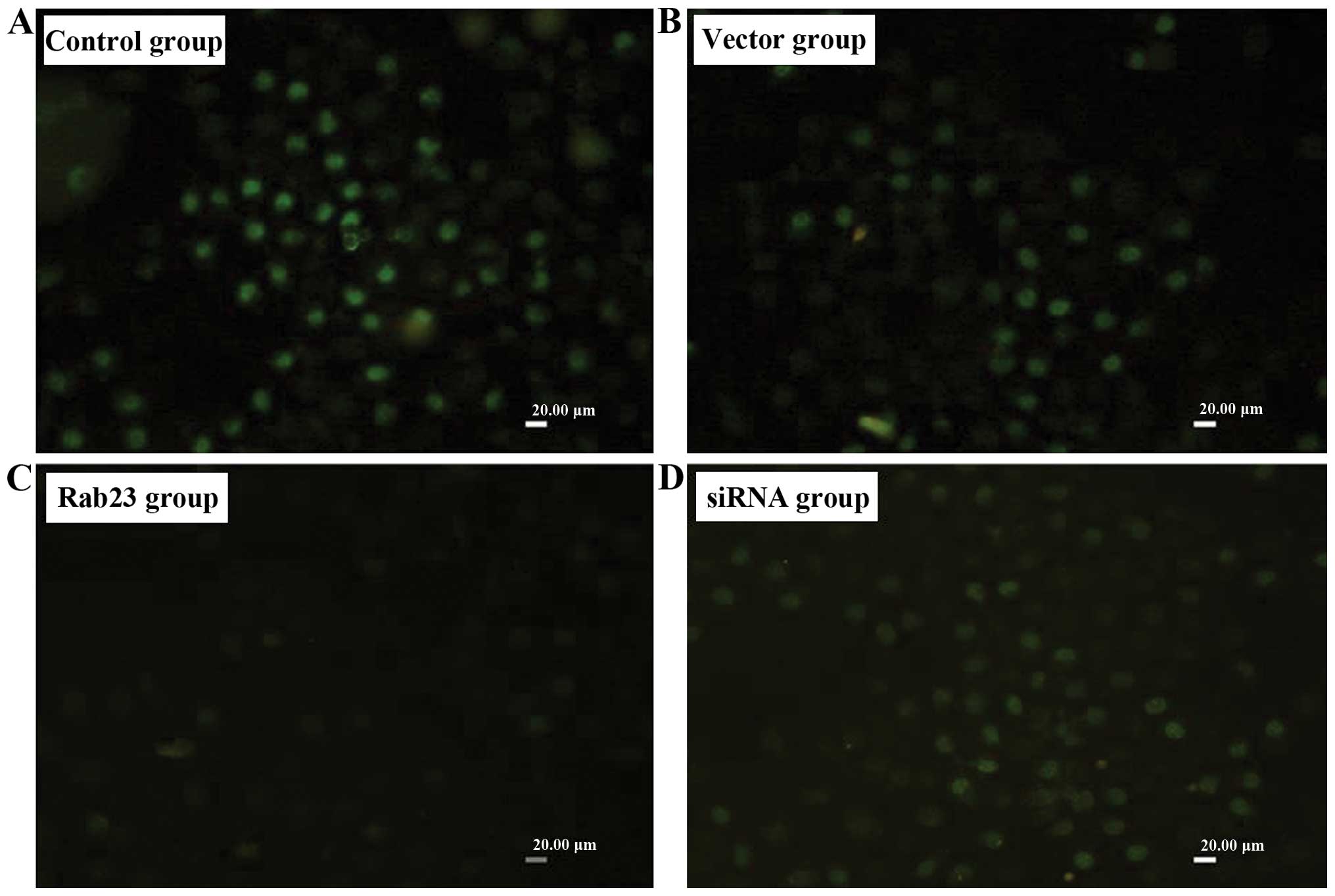
Rab23 inhibits DNA synthesis of breast
cancer cells
We detected the effect of Rab23 on the efficiency of
DNA synthesis in breast cancer cells by using BrdU labeling. After
transfection of the Rab23 plasmids or siRNA into the MDA-MB-231
(Fig. 3) and MCF-7 (Fig. 4) cells at 40 nmol, we found that
incorporation efficiency of BrdU in the group transfected with
Rab23 was lower than that in the control group (P<0.05), while
the transfection efficacy in the siRNA group was higher (P<0.05)
(Table II). There were no
differences between the vector group and the negative control
group.
| Table IIEffect of Rab23 on the incorporation
efficiency of BrdU in the breast cancer cells. |
Table II
Effect of Rab23 on the incorporation
efficiency of BrdU in the breast cancer cells.
| n | Control | Vector | Rab23 | siRNA |
|---|
| MDA-MB-231 | 5 | 32.70±1.956 | 30.10±1.025 | 20.24±1.819a | 48.73±1.55a,b |
| MCF-7 | 5 | 27.88±2.178 | 26.08±3.047 | 17.72±1.767a | 36.20±2.808a,b |
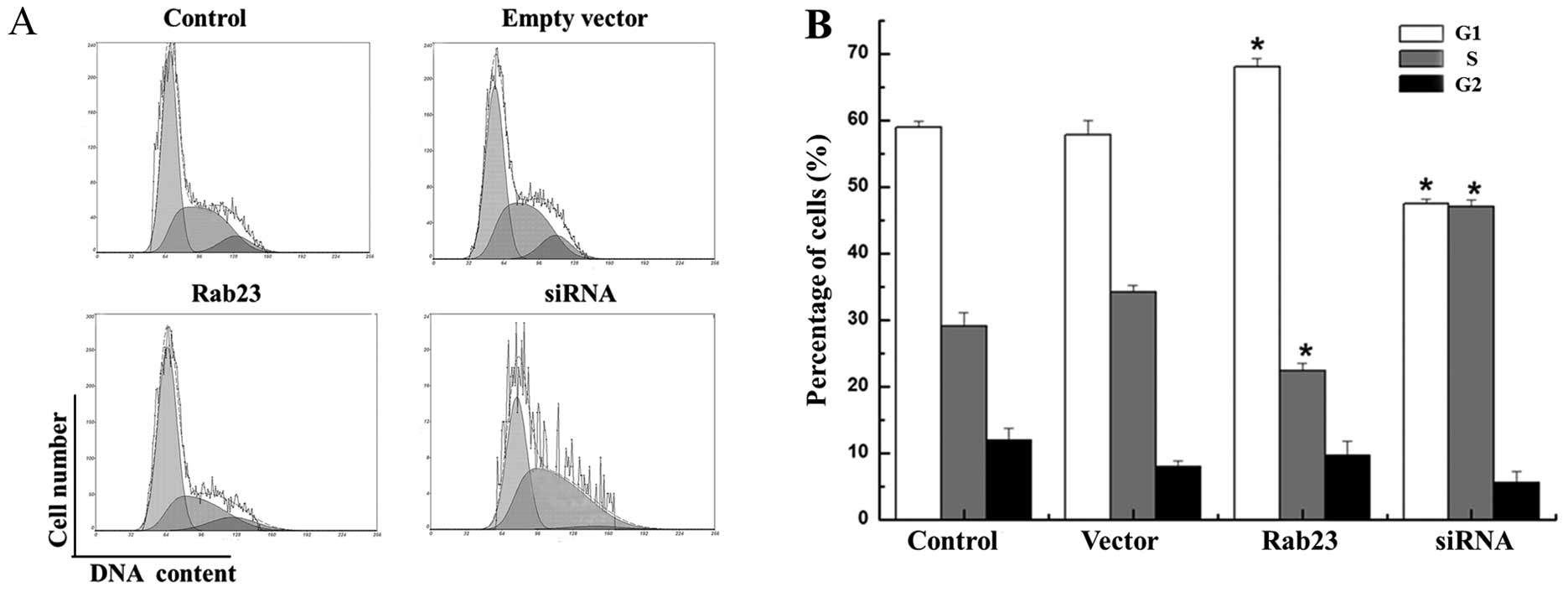
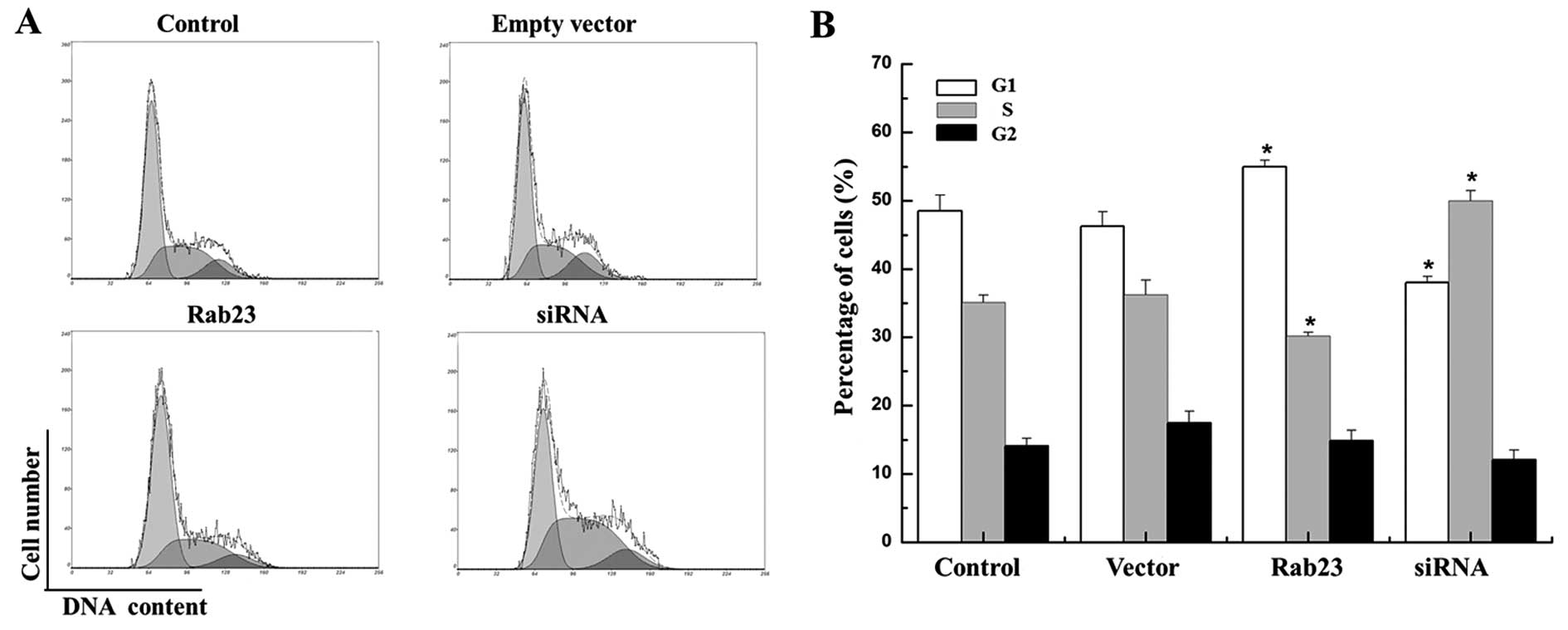
We used FCM to study the effects of Rab23 on the
cell cycle distribution of the MCF-7 and MDA-MB-231 cells. The
results showed that overexpression of Rab23 protein induced changes
in cell cycle progression pattern and caused cell cycle arrest in
the G1 phase. The cell proportion in the G1 phase was significantly
increased from 58.933±1.020 to 68.10±1.210% and from 48.525±2.311
to 55.00±0.949%, respectively in the MCF-7 and MDA-MB-231 cell
lines, which was accompanied by a decrease in the cell proportion
in the S phase from 29.133±1.943 to 22.30±1.168% and from
35.125±1.095 to 30.175±0.559% respectively, compared with the
control cells (P<0.05). Conversely, downregulation of Rab23 by
specific siRNA increased the percentage of cells in the S phase in
both the MCF-7 and MDA-MB-231 cell lines from 29.133±1.943 to
47.00±1.000% and from 35.125±1.095 to 50.00±1.500% respectively,
which was accompanied by a decrease in the cell proportion in the
G1 phase compared with the control cells from 58.933±1.020 to
47.445±0.755% and from 48.525±2.311 to 38.00±0.900%, respectively
(Figs. 5 and 6). These results indicate that Rab23
induced arrest in the G1 phase and a decrease in the S phase
population of the cell cycle, and inhibited cell growth and DNA
synthesis. Thus, Rab23 significantly reduced DNA synthesis in the
breast cancer cells.
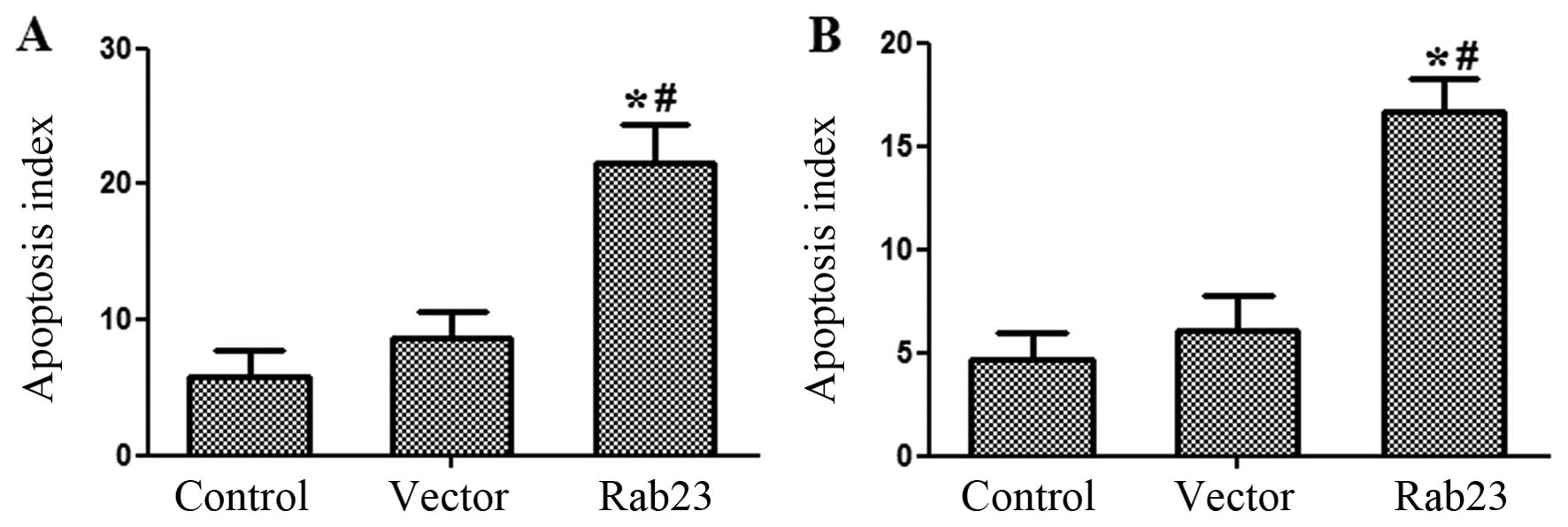
Rab23 increases breast cancer cell
apoptosis
Following FCM apoptosis analysis, it was observed
that Rab23 induced the apoptosis rate of breast cancer cells. After
transfection with Rab23, the proportion of cells in the 2 and 4
phases increased when compared with the control and vector groups.
The percentage of apoptotic MCF-7 cells increased from 4.633±1.290
to 16.70±1.552%, while the cell proportion of MDA-MB-231 cells
increased from 5.867±1.815 to 21.50±2.858% (P<0.05) (Fig. 7). There was no significant
difference between the control and vector group (P>0.05).
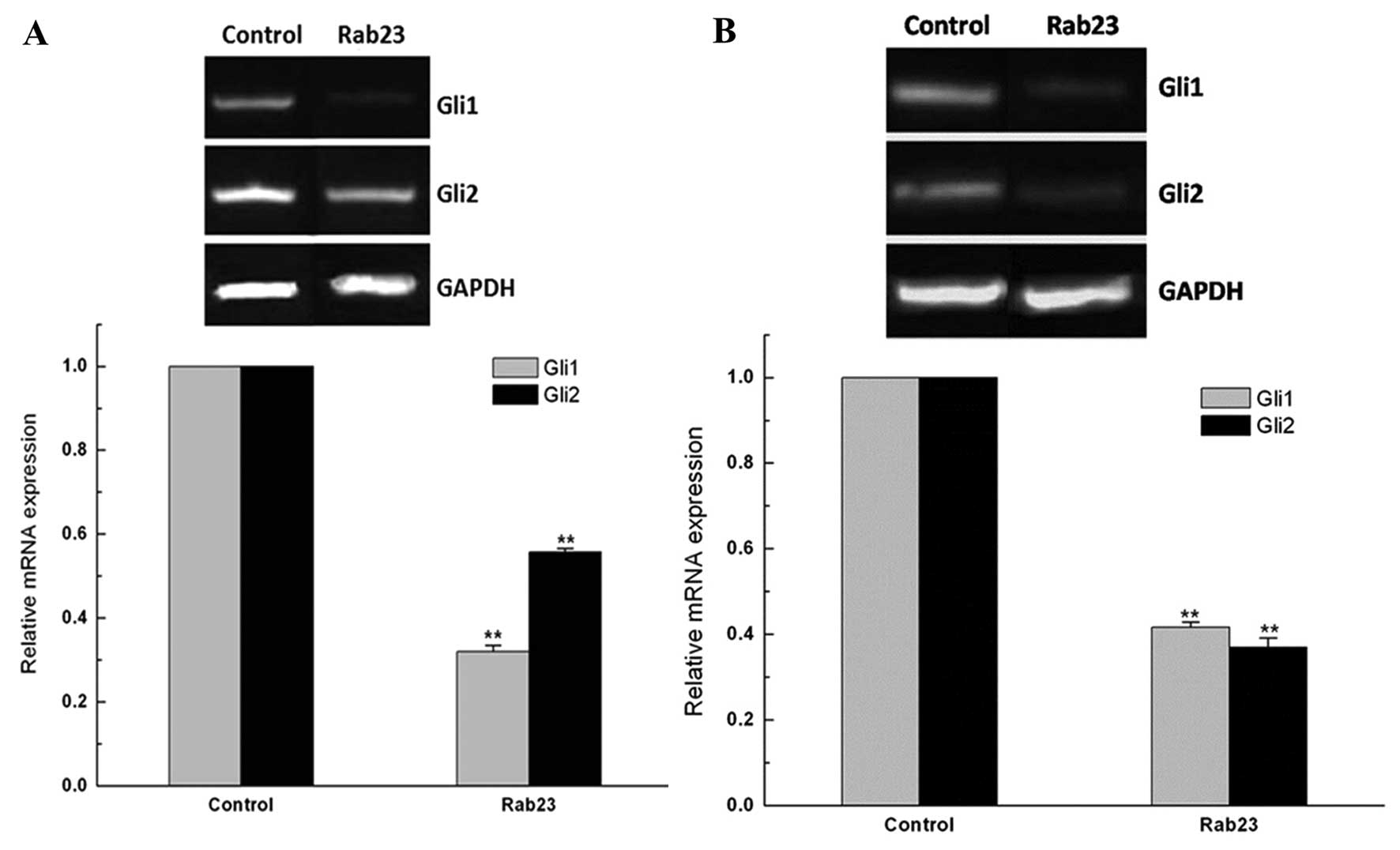
Rab23 downregulates Gli expression
It was found that Rab23 acts as a negative regulator
of the Hh signaling pathway, which drives tumorigenesis in several
types of human cancers. We found that Gli1 and Gli2 mRNA and
protein were expressed in the breast cancer MDA-MB-231 and MCF-7
cells. Following transfection with the Rab23 plasmids, the Gli1 and
Gli2 mRNA levels in the MDA-MB-231 and MCF-7 cells were
significantly decreased compared with levels in the control group
(P<0.01) (Fig. 8).
Discussion
Hh signaling is one of the main pathways in
embryogenesis which is pivotal to the development of most
vertebrate organs and tissues, and its mutation leads to birth
defects and the carcinogenesis of numerous types of tumors.
Recently, data have shown that abnormal Hh signaling activation is
also associated with human tumorigenesis (4). Constitutive activation of the Hh
pathway has been found in several types of tumors. Mutations in Hh
pathway components Ptc1 and Smo lead to pathway constitutive
activation of tumors in the brain, skin, and muscles, which is
considered ligand-independent activation of the Hh pathway
(14,15). On the other hand, ligand-dependent
pathway activation is important for the growth and survival of a
wide variety of cancers, including gastrointestinal tumors,
prostate cancer, hematological malignancies and gliomas (16). These data suggest the significance
of the Hh pathway as novel diagnostic and therapeutic targets.
Further studies have demonstrated that the plant-derived
teratogenic steroidal alkaloid cyclopamine, a potent Hh signaling
antagonist targeting the SMO protein, suppressed the growth of
cancer cells suggesting Hh pathway activation both in vitro
and in vivo (17,18). It has been reported that cyclopamine
inhibits proliferation and induces apoptosis in various human
breast cancer cell lines (19,20).
We also found that cyclopamine effectively inhibits the growth and
proliferation of breast cancer cells which indicates that Hh
signaling may be abnormally activated in breast cancer. Thus,
inhibition of Hh signaling may be a useful method for patients with
breast carcinoma (21).
Recently, Rab23 has also been implicated in
carcinogenesis. However, the role of Rab23 in tumorigenesis is
paradoxical. High Rab23 expression was reported in hepatocellular
and gastric cancer patients and was found to be associated with
tumor size (22). Another report
identified Rab23 as an upregulated gene in nonmalignant diseased
gastric tissues compared with normal gastric mucosa (13). In the neural system, Rab23 may
negatively regulate the SHH signaling pathway by controlling the
subcellular localization of essential SHH components (23,24).
Since Rab23 is a negative regulator of SHH which can induce
malignant carcinoma, we speculated that Rab23 may also contribute
to the tumorigenesis in breast cancer.
We first identified whether Rab23 is expressed in
three breast cancer cell lines and compared the levels among the
cells. We found that Rab23 was expressed at the mRNA and protein
levels but had no difference between the MDA-MB-231 and Bcap-37
cells, while there was lower expression in the MCF-7 cells. The
expression of Rab23 in the Bcap37 and MDA-MB-231 cells, which are
ER-negative, was higher than that in the MCF-7 cells, which are
ER-positive, suggesting that Rab23 expression is possibly related
with ER expression. This requires further investigation.
A previous study found that Rab23 promotes invasion
and its expression is significantly associated with diffuse-type
gastric cancers (dGCs). This is particularly intriguing since dGCs
are phenotypically more invasive than intestinal-type gastric
cancers (22). Our results are
similar since Rab23 expression was higher in the highly metastatic
MDA-MB-231 cells and lower in the low invasive MCF-7 cells. These
studies suggest that Rab23 may play a role in tumor metastasis. We
propose that the difference in Rab23 expression in breast cancers
may be caused by functional changes in the Rab family members. It
is known that Ras is a family of genes encoding small GTPases that
are involved in cellular signal transduction. It plays an important
role in tumorigenesis and tumor metastasis. It may regulate tumor
cell growth, differentiation and survival.
In the present study, we observed the effects of
Rab23 on the proliferation and apoptosis of breast cancer cells.
The results of MTT and in colony forming assays demonstrated that
Rab23 reduced the growth and proliferation of breast cancer cells.
Furthermore, the incorporation efficiency of BrdU and FCM showed
that Rab23 significantly inhibited cell DNA synthesis. The effect
of inhibition was similar to the effects of cyclopamine, a specific
signaling inhibitor for the Hh signaling pathway, in breast cancer
cells in our previous study (21).
Subsequently, FCM apoptosis detection revealed that Rab23 induced
the apoptosis of breast cancer cells. These results suggest that
the inhibitory effect of Rab23 on breast cancer cells may be due to
the inhibition of DNA synthesis and induction of apoptosis in the
breast cancer cells.
Rab23, as a new negative regulator of the SHH
signaling pathway, has been investigated in many laboratories and
it can control the subcellular localization of essential SHH
components. Other studies concerning the effects of Rab23 on
carcinomas confirmed that Rab23 plays a role in antagonizing cancer
cell survival and increasing the basal rate of apoptosis. The exact
cellular mechanism of Rab23 is currently unclear, but data from
mouse genetic studies indicate that Rab23 acts downstream of Smo
and upstream of Gli proteins. Some genetic studies have indicated
that the primary target of Rab23 is the Gli2 activator and that
Rab23 and Gli3 repressor have additive effects on mouse neural
patterning. We demonstrated that Rab23 suppressed Gli1
transcriptional activity in a Su(Fu)-dependent manner (Suppressor
of fused) (25). In order to
explore whether the effect of Rab23 on growth, proliferation, and
apoptosis of breast cancer cells is regulated through the Hh
signaling pathway, we further detected the activated state of the
Hh signaling pathway in this study. We found that expression of
Gli1 and Gli2 appears to be dependent on Rab23. To date, there are
no studies on Gli3 expression in breast cancer cells, and we did
not study Gli3 in breast cancer cells. The exact mechanism of Rab23
regulating Hh will require further investigation. The fact that
Rab23 expression suppresses Gli1 and Gli2 expression does not rule
out the possibility that the effect of Rab23 may be related to the
location of Rab23 in the endocytic pathway and the role in
facilitating vesicular transport and controlling endocytic
progression to lysosomes. In particular, Rab23 expression was
higher in the MDA-MB-231 cells, which are more invasive, suggesting
that there may be multiple mechanisms under which Rab23 has
influence on the proliferation, apoptosis and cell invasion of
breast cancer cells.
In conclusion, Rab23 expression was detected in
breast cancer cells, and Rab23 represents a potent tumor suppressor
in breast cancer, which can inhibit cell growth and proliferation
and induced cell apoptosis of breast cancer cells. These effects
could be due to the inhibition by Rab23 of Gli1 and Gli2 mRNA
expression. The exact molecular mechanism of Rab23 mediation or
modulation of the Hh signaling pathway requires additional
investigation. Rab23 plays an important role in the tumorigenesis
of human breast cancer, and may be a new biological target for the
prognosis and treatment of breast cancer. However, further
investigations, including detection in clinical samples, are needed
to explore and confirm the significance of Rab23 in the
pathogenesis and treatment of breast cancer.
Acknowledgments
This study was supported by grants (no. 31200876 and
no. 31371412) from the National Natural Science Foundation of China
(NSFC).
References
|
1
|
Benson JR, Jatoi I, Keisch M, Esteva FJ,
Makris A and Jordan VC: Early breast cancer. Lancet. 373:1463–1479.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ingham PW and McMahon AP: Hedgehog
signaling in animal development: Paradigms and principles. Genes
Dev. 15:3059–3087. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
McMahon AP, Ingham PW and Tabin CJ:
Developmental roles and clinical significance of hedgehog
signaling. Curr Top Dev Biol. 53:1–114. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Saldanha G: The Hedgehog signalling
pathway and cancer. J Pathol. 193:427–432. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kasper M, Jaks V, Fiaschi M and Toftgård
R: Hedgehog signalling in breast cancer. Carcinogenesis.
30:903–911. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Naylor TL, Greshock J, Wang Y, Colligon T,
Yu QC, Clemmer V, Zaks TZ and Weber BL: High resolution genomic
analysis of sporadic breast cancer using array-based comparative
genomic hybridization. Breast Cancer Res. 7:R1186–R1198. 2005.
View Article : Google Scholar
|
|
7
|
Nessling M, Richter K, Schwaenen C, Roerig
P, Wrobel G, Wessendorf S, Fritz B, Bentz M, Sinn HP, Radlwimmer B,
et al: Candidate genes in breast cancer revealed by
microarray-based comparative genomic hybridization of archived
tissue. Cancer Res. 65:439–447. 2005.PubMed/NCBI
|
|
8
|
Wolf I, Bose S, Desmond JC, Lin BT,
Williamson EA, Karlan BY and Koeffler HP: Unmasking of
epigenetically silenced genes reveals DNA promoter methylation and
reduced expression of PTCH in breast cancer. Breast Cancer Res
Treat. 105:139–155. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mukherjee S, Frolova N, Sadlonova A, Novak
Z, Steg A, Page GP, Welch DR, Lobo-Ruppert SM, Ruppert JM, Johnson
MR, et al: Hedgehog signaling and response to cyclopamine differ in
epithelial and stromal cells in benign breast and breast cancer.
Cancer Biol Ther. 5:674–683. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Moraes RC, Zhang X, Harrington N, Fung JY,
Wu MF, Hilsenbeck SG, Allred DC and Lewis MT: Constitutive
activation of smoothened (SMO) in mammary glands of transgenic mice
leads to increased proliferation, altered differentiation and
ductal dysplasia. Development. 134:1231–1242. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Iorns E, Turner NC, Elliott R, Syed N,
Garrone O, Gasco M, Tutt AN, Crook T, Lord CJ and Ashworth A:
Identification of CDK10 as an important determinant of resistance
to endocrine therapy for breast cancer. Cancer Cell. 13:91–104.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu YJ, Wang Q, Li W, Huang XH, Zhen MC,
Huang SH, Chen LZ, Xue L and Zhang HW: Rab23 is a potential
biological target for treating hepatocellular carcinoma. World J
Gastroenterol. 13:1010–1017. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim KR, Oh SY, Park UC, Wang JH, Lee JD,
Kweon HJ, Kim SY, Park SH, Choi DK, Kim CG, et al: Gene expression
profiling using oligonucleotide microarray in atrophic gastritis
and intestinal metaplasia. Korean J Gastroenterol. 49:209–224.
2007.In Korean. PubMed/NCBI
|
|
14
|
Johnson RL, Rothman AL, Xie J, Goodrich
LV, Bare JW, Bonifas JM, Quinn AG, Myers RM, Cox DR, Epstein EH Jr,
et al: Human homolog of patched, a candidate gene for the basal
cell nevus syndrome. Science. 272:1668–1671. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wicking C, Smyth I and Bale A: The
hedgehog signalling pathway in tumorigenesis and development.
Oncogene. 18:7844–7851. 1999. View Article : Google Scholar
|
|
16
|
Ruiz i Altaba A, Mas C and Stecca B: The
Gli code: An information nexus regulating cell fate, stemness and
cancer. Trends Cell Biol. 17:438–447. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Taipale J, Chen JK, Cooper MK, Wang B,
Mann RK, Milenkovic L, Scott MP and Beachy PA: Effects of oncogenic
mutations in Smoothened and Patched can be reversed by
cyclo-pamine. Nature. 406:1005–1009. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Berman DM, Karhadkar SS, Hallahan AR,
Pritchard JI, Eberhart CG, Watkins DN, Chen JK, Cooper MK, Taipale
J, Olson JM, et al: Medulloblastoma growth inhibition by hedgehog
pathway blockade. Science. 297:1559–1561. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kubo M, Nakamura M, Tasaki A, Yamanaka N,
Nakashima H, Nomura M, Kuroki S and Katano M: Hedgehog signaling
pathway is a new therapeutic target for patients with breast
cancer. Cancer Res. 64:6071–6074. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang X, Harrington N, Moraes RC, Wu MF,
Hilsenbeck SG and Lewis MT: Cyclopamine inhibition of human breast
cancer cell growth independent of Smoothened (Smo). Breast Cancer
Res Treat. 115:505–521. 2009. View Article : Google Scholar
|
|
21
|
Wang HB, Liu YL, Hu YZ and Chi SM: Effect
of cyclopamine on growth and proliferation of human mammary
carcinoma cell Bcap-37. J Med Postgraduates. 20:567–571. 2007.In
Chinese.
|
|
22
|
Hou Q, Wu YH, Grabsch H, Zhu Y, Leong SH,
Ganesan K, Cross D, Tan LK, Tao J, Gopalakrishnan V, et al:
Integrative genomics identifies RAB23 as an invasion mediator gene
in diffuse-type gastric cancer. Cancer Res. 68:4623–4630. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Evans TM, Simpson F, Parton RG and Wicking
C: Characterization of Rab23, a negative regulator of sonic
hedgehog signaling. Methods Enzymol. 403:759–777. 2005. View Article : Google Scholar
|
|
24
|
Eggenschwiler JT, Bulgakov OV, Qin J, Li T
and Anderson KV: Mouse Rab23 regulates hedgehog signaling from
smoothened to Gli proteins. Dev Biol. 290:1–12. 2006. View Article : Google Scholar
|
|
25
|
Chi S, Xie G, Liu H, Chen K, Zhang X, Li C
and Xie J: Rab23 negatively regulates Gli1 transcriptional factor
in a Su(Fu)-dependent manner. Cell Signal. 24:1222–1228. 2012.
View Article : Google Scholar : PubMed/NCBI
|