Introduction
Esophageal cancer (EC) ranks eighth in cancer
incidence with an estimated 456,000 new cases in 2012 (3.2% of the
total patients) and sixth in cancer mortality with an estimated
400,000 deaths (4.9% of the total patients) (1). There are two primary pathological
types of EC, esophageal adenocarcinoma (EAC) and esophageal
squamous cell carcinoma (ESCC), which account for 95% of all cases
of EC (2). ESCC has a predilection
for Asian populations and more than half of all new cases of ESCC
worldwide are diagnosed in China (3). The incidence of EAC has increased by
6-fold in the United States during the last few decades (4). Although there have been advances in
the field of operative methods, chemotherapy (Cis-platinum CDDP)
and radiotherapy (5), the prognosis
of EC patients remain to be determined. The development of new
biomarkers for the early detection and potential therapeutic
targets of EC are important for therapeutic improvement (6).
Serine proteases (plasminogen-plasmin system) is
dysregulated in various human cancers, emerging as an important
mechanism to regulate cell growth, migration, invasion, metastasis
and angiogenesis (7).
Kallikrein-related peptidases (KLKs) are the largest cluster among
all serine proteases within the human genome (8,9). This
cluster consists of a subgroup of 15 homologous secreted trypsin
and chymotrypsin-like serine proteases, encoded by a tightly
clustered multigene family on chromosome 19q13.4 (8). Kallikreins are often co-expressed in
the skin, breast, prostate, pancreas and brain, and secreted by
epithelial cells, from which they enter the bodily fluids such as
sweat, milk, saliva, seminal plasma and cerebrospinal fluid or
pericellular spaces (10).
Kallikrein genes/proteins are aberrantly expressed in many types of
cancer and their expression is often associated with patient
prognosis. Abnormal regulation of KLKs interferes with different
stages of cancer growth, including tumor differentiation,
angiogenesis and metastasis (11).
KLK family members have been reported to be promising
diagnostic/prognostic biomarkers for several cancer types,
including breast, ovarian, prostate and testicular carcinomas. For
example, hK 3 encoded by KLK3, which is well known as
prostate-specific antigen (PSA), used as a marker for prostate
cancer (8).
The kallikrein-related peptidase 10 (KLK10) gene,
also known as normal epithelial cell-specific 1 (NES1), is a member
of the human kallikrein-related family of peptidases (8). The KLK10 gene encodes a 31 kDa
secreted serine protease with 276 amino acids and is expressed in
various tissues and organs such as skin, prostate, fallopian tube,
salivary gland, colon and testis. KLK10 exerts various
physiological functions in tissues and is involved in cancer
progression (12). KLK10 has been
found to be downregulated in breast cancer, prostate tumor,
non-small cell lung cancer and renal cell carcinoma (13–16).
Ectopic KLK10 reduces anchorage-independent growth and enhances
drug sensitivity in liver and breast cancer. Thus KLK10 probably
acts as a tumor suppressor in these cancer types. By contrast,
KLK10 is overexpressed in ovarian cancer, uterine papillary serous
carcinoma, pancreatic ductal adenocarcinoma, colorectal carcinoma
and oral squamous cell carcinoma (11,17–21)
and acts as a tumor promoter. Knockdown of KLK10 inhibits
proliferation and tumorigenicity in pancreatic cancer.
Additionally, the dysregulation of KLK10 expression is useful for
diagnosis and associated with prognosis of various types of cancer.
KLK10 performs as an unfavourable prognostic role in ovarian
cancer, pancreatic ductal adenocarcinoma and gastrointestinal
tumors, or has a favourable prognostic role in breast cancer,
prostate tumor and non-small cell lung cancer, respectively.
KLK10 expression is affected by several factors at
various levels including transcriptional, translational and
post-translational modification. In hormone tightly-regulated
cancers, KLK10 is mainly upregulated by estrogen, androgen and
progestin (22). By contrast, the
downregulation of KLK10 is often associated with hypermethylation
in CpG rich exon 3 (23), instead
of in the promoter region. Additionally, many miRNAs are also
involved in the post-transcriptional regulation of KLK10, and
let-7f, miR-224 and miR-516a in ovarian carcinoma, as well as
miR-21 in prostate cancer, can directly target KLK10 and reduce
KLK10 expression. A previous study also found that KLK10 and KLK
expression was altered by the transcriptional factor specificity
protein 1 (24).
The primary aim of the present study was to examine
the expression changes of KLK10 in clinical EC samples in
comparison with normal tissues, and experimentally demonstrate the
potential biological functions of KLK10 in EC.
Materials and methods
Patients and cell lines
Patients analyzed in this study were diagnosed with
EC during the period 1999–2014 at Anshan Cancer Hospital (Anshan,
Liaoning, China). None of the patients had been administered
chemotherapy or radiotherapy prior to surgery. In total, 83
formalin-fixed, paraffin-embedded specimens were obtained from
resected tumors and 11 adjacent normal specimens. The patient age
ranged from 31 to 78 years with a mean age of 59.8±10.83 years.
Tumors were graded according to the WHO grading system and staged
according to the modified Astler-Coller system (MAC).
The human esophageal squamous cancer cell lines
(Eca-109 and TE-1) were purchased from the Type Culture Collection
of the Chinese Academy of Sciences (Shanghai, China). Eca-109 was
cultured in RPMI-1640, supplemented with 10% fetal bovine serum
(FBS) (both from Gibco-Life Technologies, Carlsbad, CA, USA) and
100 µg/ml each of penicillin and streptomycin
(Invitrogen-Life Technologies, Carlsbad, CA, USA). The TE-1 was
cultured in complete medium of high-glucose Dulbecco's modified
Eagle's medium (DMEM) supplemented with 10% (FBS) (Gibco-Life
Technologies) and 100 µg/ml each of penicillin and
streptomycin (Invitrogen-Life Technologies). The cells were
maintained in a humidified 5% CO2 incubator at 37°C.
Reagents and antibodies
The Cell Counting Kit-8 (CCK-8, cat no. KGA317),
Annexin V-FITC Apoptosis Detection kit (KGA108) and the Cell Cycle
Detection kit (KGA512) were purchased from KeyGen Biotech (Nanjing,
China). cis-Dichlorodiamine platinum (CDDP) was purchased from
Haosen Pharmaceutical Inc. (Jiangsu, China). The antibodies used
for the western blot analysis were: KLK10 (bs-2531R, diluted 1:500)
purchased from Bioss (Beijing, China); β-actin (sc-47778, diluted
1:1,000) from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA);
and active caspase-3 (AJ1131b, diluted 1:1,000) from Abgent (San
Diego, CA, USA). PARP (no. 5625, diluted 1:1,000), CDK2 (no. 2546,
diluted 1:1,000) and cyclin A2 (no. 4656, diluted 1:1,000) were
purchased from Cell Signaling Technology (Danvers, MA, USA). Goat
anti-rabbit, goat anti-mouse IgG, peroxidase-conjugated secondary
antibodies (31460 and 31430, both diluted 1:10,000) and goat
anti-rabbit IgG (H+L) secondary antibody were purchased from
Thermo-Pierce (Rockford, IL, USA).
Immunohistochemistry (IHC)
The cytoplasmic expression of KLK10 in the human
esophageal squamous cancer cell was evaluated as previously
described. The cytoplasmic expression for KLK10 was scored as: i),
0<5% of the squamous cancer cells in the respective lesions;
ii), 5–25% of the squamous cancer cells in the respective lesions
were rated 1; iii), 26–50% of the squamous cancer cells in the
respective lesions were rated 2; iv), 51–75% of the squamous cancer
cells in the respective lesions were rated 3; and v), >75% of
the squamous cancer cells in the respective lesions were rated 4.
The intensity was graded as: i), negative, 0; ii), weak, 1+; iii),
moderate, 2+; and iv), strong, 3+. A final score between 0 and 12
was achieved by multiplication of the extent of positivity and
intensity. Scores of 9–12 were defined as '(+++)̓, scores of 5–8
were defined as '(++)̓, scores of 1–4 were defined as '(+)̓, and
scores of 0 was defined as '(−)̓. (−), (+) were considered negative
and (++), (+++) were considered positive (25). The slides were blindly labeled and
scored by two independent pathologists.
Isolation of total RNA and quantitative
RT-PCR
Total RNA was extracted from EC cells (Eca-109 and
TE-1) using TRIzol-up reagent (Invitrogen-Life Technologies),
according to the manufacturer's instructions. The primers used
were: KLK10 forward; 5′-GCCCGGAGAGTGAAGTACAA-3′ and reverse,
5′GTAAACACCCCACGAGAGGA-3′; β-actin forward,
5′GCATGGAGTCCTGTGGCAT-3′; and reverse 5′-CTAGAAGCATTTGCGGTGG-3′.
mRNA was reverse transcribed to cDNA using a SuperScript™ Two-Step
RT-PCR kit (Takara, Dalian, China) by SureCycler 8800 (Agilent
Technologies, Santa Clara, CA, USA). The PCR amplification was
performed using SYBR-Green PCR Master Mix (Takara) by Stratagene
Mx3005P (Agilent Technologies) under the following conditions: 95°C
for 30 sec, 40 cycles of 95°C for 5 sec and 55°C for 20 sec. The
expression of KLK10 was normalized by using β-actin as an internal
control. The relative expression was analyzed using the
2−ΔΔCt method in MxPro-Mx3005P software.
siRNA transfection
Small interfering RNAs (siRNAs) targeting KLK10 and
siRNA negative control were chemically synthesized (Genepharma,
Shanghai, China). The siRNA sequences used were: KLK10 siRNA 1:
5′-TTGTTGTACTTCACTCTCCGG-3′; KLK10 siRNA 2:
5′-ATGACTTTATTGATCCAGGAC-3′; negative control siRNA:
5′-UUCUCCGAACGUGUCACGUTT-3′. The EC cells were seeded in six-well
plates and incubated overnight for transfection. The medium was
exchanged for serum-free DMEM when the cell confluence reached
60–70%. Complexes of siRNA (final concentration was 20 µM
according to the manufacturer) and Lipofectamine 2000
(Invitrogen-Life Technologies) were added into the medium for 6 h.
Subsequently, the medium was converted with fresh medium
supplemented with 10% cultured FBS. The effectiveness of RNA
interference was assessed by quantitative RT-PCR and western blot
analysis.
Cell counting kit (CCK-8) assay
Cell proliferation was determined using CCK-8 and
KGA317, according to the manufacturer's instructions. Two thousand
cells (Eca-109)/wells and 3,000 cells (TE-1)/wells were seeded in a
96-well cell culture plate, and three replicates were prepared for
each condition. After the cells were grown at different times (1–6
days) in a 37°C incubator the medium was replaced with 100
µl of RPMI-1640/DMEM containing 10 µl of CCK-8
reagent, and the cells were incubated for 2 h at 37°C. The optical
density was then measured using an EnSpire™ 2300 Multilabel reader
(PerkinElmer, Waltham, MA, USA) at 450 nm, after which the mean
values were measured and the growth curves drawn.
Colony formation assay
Eca-109 and TE-1 were used for colony formation
analysis. The cells were seeded in 6-well plates at a density of
500 cells (Eca-109)/800 cells (TE-1) of each well in 2 ml of medium
(RPMI-1640 and DMEM) containing 10% FBS and cultured for 14 days in
a humidified 5% CO2 incubator at 37°C. The colonies were
fixed in 4% paraformaldehyde for 15 min, and stained with a
solution containing 1% crystal violet for 15 min. Subsequently, the
plates were washed with PBS and dried at room temperature.
Cell cycle and apoptosis assays
To evaluate the effects on cell cycle arrest and
CDDP-induced apoptosis by downregulation of KLK10, the cells were
examined using the cell cycle detection kit and the Annexin V-FITC
apoptosis detection kit according to the manufacturer's
instructions. For cell cycle analysis, Eca-109 cells were seeded in
6-well plates (1×105 and 2×105 cells/dish for
analysis of cell cycle arrest and apoptosis, respectively). After
siRNA was transfected for 48 h, a total of 1×106 cells
was pelleted by centrifugation and washed twice with PBS. The cell
pellets were resuspended in 500 µl of ice-cold 70% ethanol
and incubated at −20°C overnight. The fixed cells were centrifuged
and the pellets were washed with PBS. After incubation with 100
µl RNase A (10 µg/ml) for 30 min at 37°C in the dark,
the cells were resuspended in 400 µl PI (50 µg/ml)
and placed at 37°C in the dark for 30 min. The stained cells were
analyzed using an Accuri C6 flow cytometer (Accuri Cytometers Inc.,
Ann Arbor, MI, USA). For the apoptosis analysis, after siRNA were
transfected in Eca-109 cells for 48 h followed by treatment with
CDDP (5 µg/ml) for 48 h the cells were trypsinized, washed
with cold PBS and suspended in PBS. The cells were stained using
the Annexin V-FITC reaction reagent [5 µl of Annexin V-FITC
and propidium iodide (PI)] at 37°C for 30 min in the dark. The
stained cells were analyzed using an Accuri C6 flow cytometer
(Accuri Cytometers Inc.) as previously described (26).
Western blot analysis
After siRNA transfection for 2 days, the cells were
harvested and lysed in RIPA buffer (KGP702) and proteins were
extracted, and supplemented with 1 mM phenylmethylsulfonyl fluoride
(PMSF; KGP610 from KeyGen Biotech). The mixture was centrifuged at
12,000 × g for 15 min and the supernatant was collected. The
protein concentration was determined using the BCA Assay kit
(KGPBCA), and each sample contained 30 µg protein per 10
µl. The protein samples were mixed with loading buffer
(KGP101) and the proteins were separated using 12% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and
transferred to polyvinylidene difluoride (PVDF) membranes (Bio-Rad
Laboratories, Hercules, CA, USA). After soaking in blocking buffer
at room temperature for 2 h, the membranes were incubated at 4°C
overnight with the primary antibody and were subsequently incubated
at 37°C for 2 h with the HRP-conjugated secondary antibody. The
bands were visualized by chemiluminescence, imaged using a ChemiDoc
XRS and analyzed using Image Lab (both from Bio-Rad). All protein
extracts were completed on the ice (26).
Statistical analysis
Data represented at least three independent
experiments and were statistically evaluated by Pearson's
Chi-square test, correction for continuity analysis or Student's
t-test with the statistical analysis software SPSS version 19.0
(IBM). P<0.05 was considered to be statistically
significant.
Results
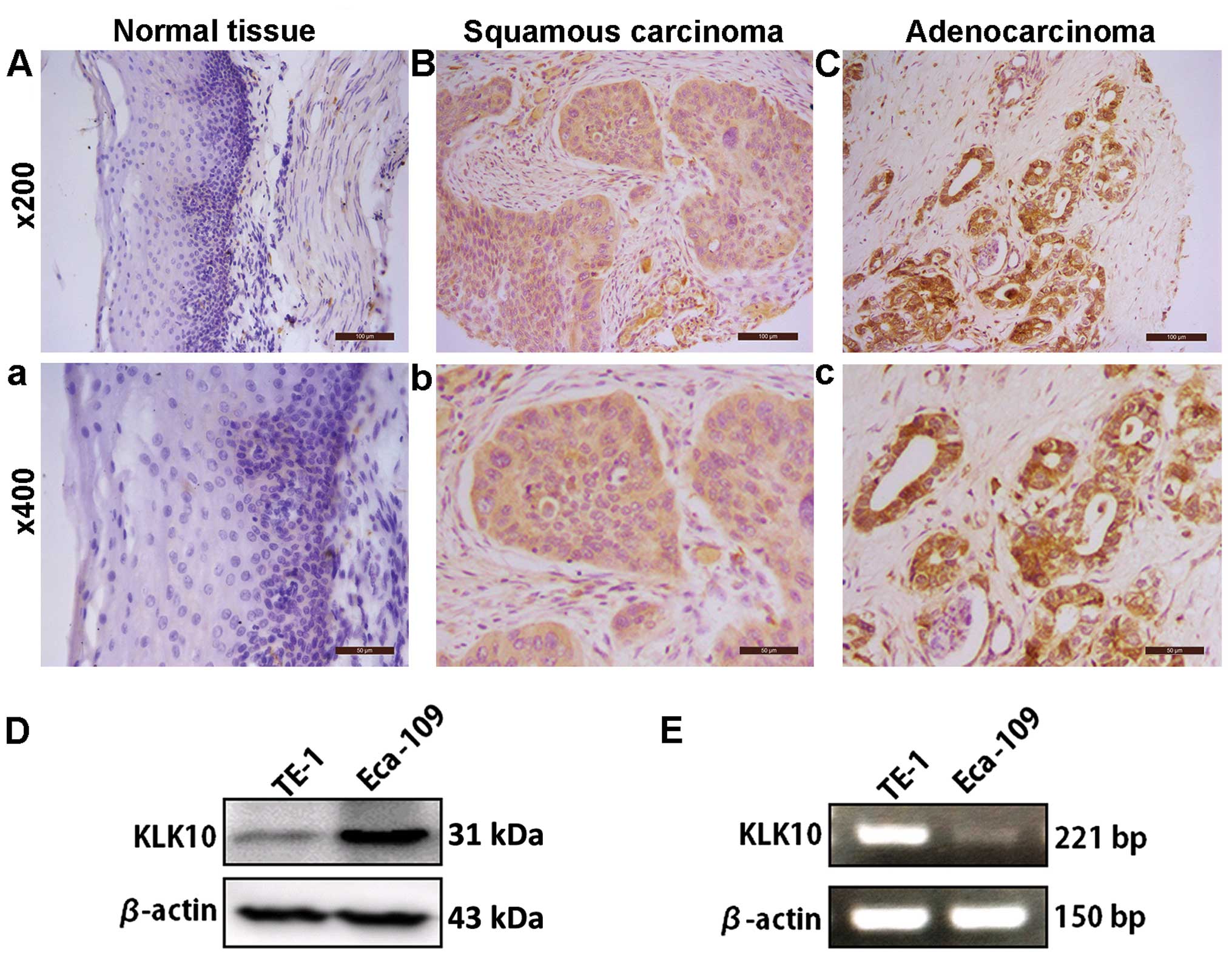
Expression of KLK10 in human EC tissue
samples and cell lines
To detect the expression of KLK10 in human EC tissue
samples, 83 EC specimens were assessed by IHC staining followed by
the Chi-square test and correction for continuity analysis. KLK10
showed strong cytoplasmic staining in the tumor samples, while the
normal epithelium tissues presented no or weekly cytoplasmic
staining (Fig. 1A–C). The data from
IHC analysis indicated KLK10 was markedly increased in tumor
samples, as the positive staining is 67 out of 83 (80.72%) in human
EC and only 3 out of 11 (27.27%) in normal tissues (P=0.001,
Table I). We analyzed the
association between KLK10 expression and other clinical parameters
and found that KLK10 expression correlates with tumor diameter
(P=0.046), although no significant difference was identified in the
pathological types, nodal status and tumor grade (Table II). Then, we detected the
expression of KLK10 in the Eca-109 and TE-1 EC cell lines. The two
cell lines expressed KLK10 protein as shown by western blot
analysis with higher KLK10 in Eca-109, than in TE-1 cells (Fig. 1D). However, its expression in mRNA
levels was opposite to that of the protein expression (Fig. 1E).
 | Table IAssociations between KLK10 expression
and human esophageal tissue. |
Table I
Associations between KLK10 expression
and human esophageal tissue.
| Variable | | Total patients | Expression of KLK10
protein
| Positive (%) | P-value |
|---|
| (−) (+) | (++) (+++) |
|---|
| General | Normal tissue | 11 | 8 | 3 | 27.27 | 0.001a |
| Tumor tissue | 83 | 16 | 67 | 80.72 | |
| Table IIAssociations between KLK10 expression
and clinical parameters. |
Table II
Associations between KLK10 expression
and clinical parameters.
| Variable | Expression of KLK10
| Positive (%) | P-value |
|---|
| (−)(+) | (++)(+++) |
|---|
| Years of age | | | | |
| <60 | 9 | 31 | 77.5 | 0.581a |
| ≥60 | 7 | 36 | 83.7 | |
| Gender | | | | |
| Male | 13 | 57 | 81.4 | 0.708a |
| Female | 3 | 10 | 76.9 | |
| Degree of
differentiation | | | | |
| Well | 6 | 21 | 77.8 | 0.780a |
| Moderately | 8 | 32 | 78.0 | |
| Poorly | 2 | 13 | 86.7 | |
| Tumor diameter
(cm) | | | | |
| <5 | 2 | 26 | 92.9 | 0.046a |
| ≥5 | 14 | 41 | 74.5 | |
| Lymph node
metastasis | | | | |
| Yes | 7 | 34 | 82.9 | 0.782a |
| No | 9 | 33 | 78.6 | |
| Tumor grade | | | | |
| I–IIa | 9 | 31 | 79.5 | 0.776a |
| IIb–III, IV | 7 | 36 | 83.7 | |
| Pathological
types | | | | |
| Squamous cell
carcinoma | 15 | 53 | 78.3 | 0.285b |
|
Adenocarcinoma | 1 | 14 | 92.9 | |
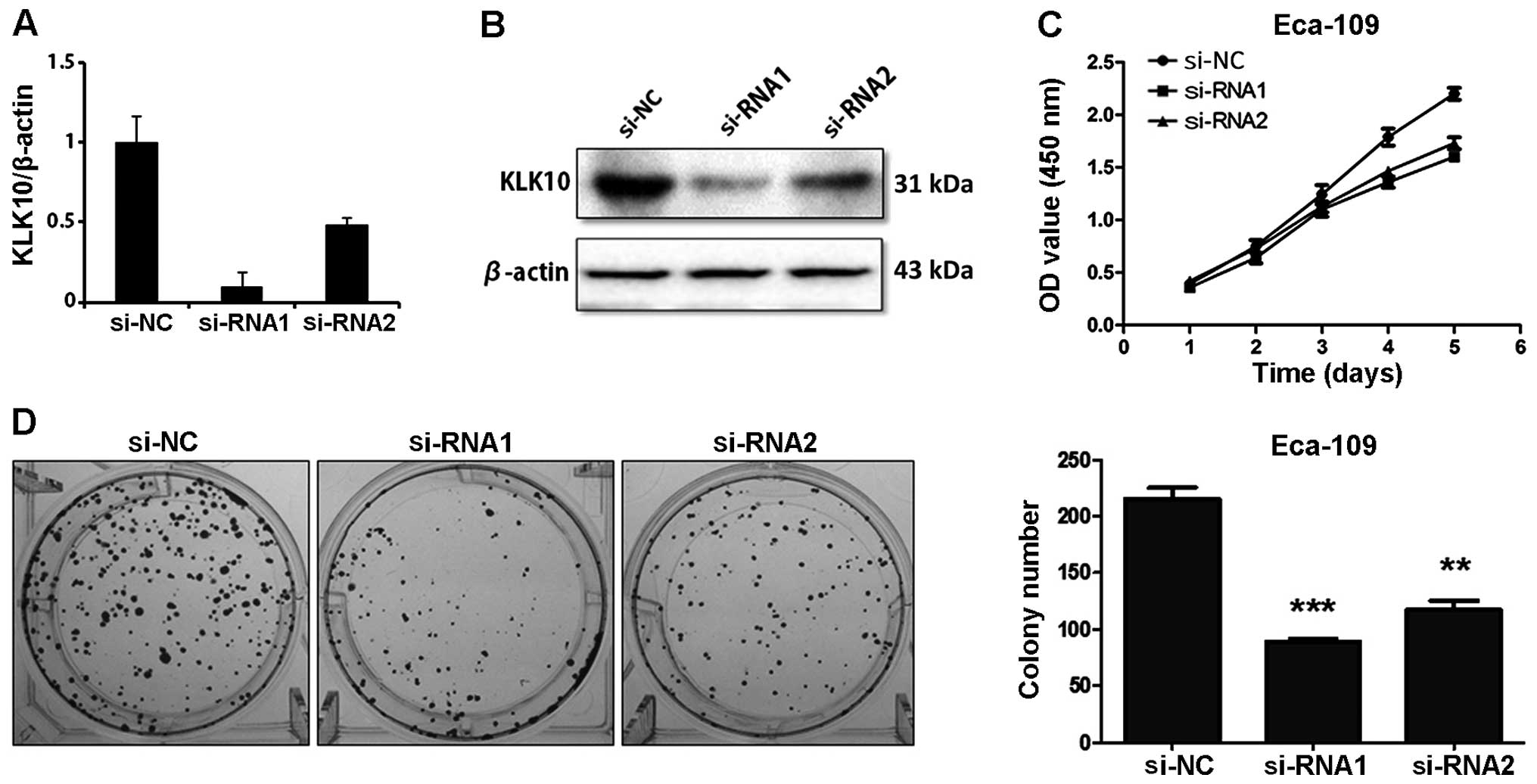
Knockdown of KLK10 inhibits the growth
and colony formation of Eca-109 and TE-1 cells
Two small interference RNAs were used to
downregulate KLK10 expression. As shown by qPCR (Fig. 2A), the mRNA levels of KLK10 were
reduced in KLK10 siRNA-1 and -2 treated cells compared with the
control siRNA-transfected cells, respectively, which was consistent
with KLK10 protein expression (Fig.
2B). In order to investigate the importance of KLK10 in cell
proliferation, a CCK-8 assay was used and we observed a significant
reduction in cell viability in ESCC cells treated with KLK10
siRNAsiRNA-1 and -2. The cells treated with KLK10 siRNAs exhibited
a gradual growth (P<0.05) (Fig.
2C) and formed fewer colonies than the Eca-109 control siRNA
cells (Fig. 2D, left panel). The
bar graph demonstrated that knockdown of KLK10 in Eca-109 cells
decreased the number of colonies as much as ~2-fold relative to the
control (P<0.001 and P<0.01) (Fig. 2D, right panel). These results
suggested the relationship between KLK10 and the cell proliferation
of Eca-109 cells. We also observed a significant reduction of
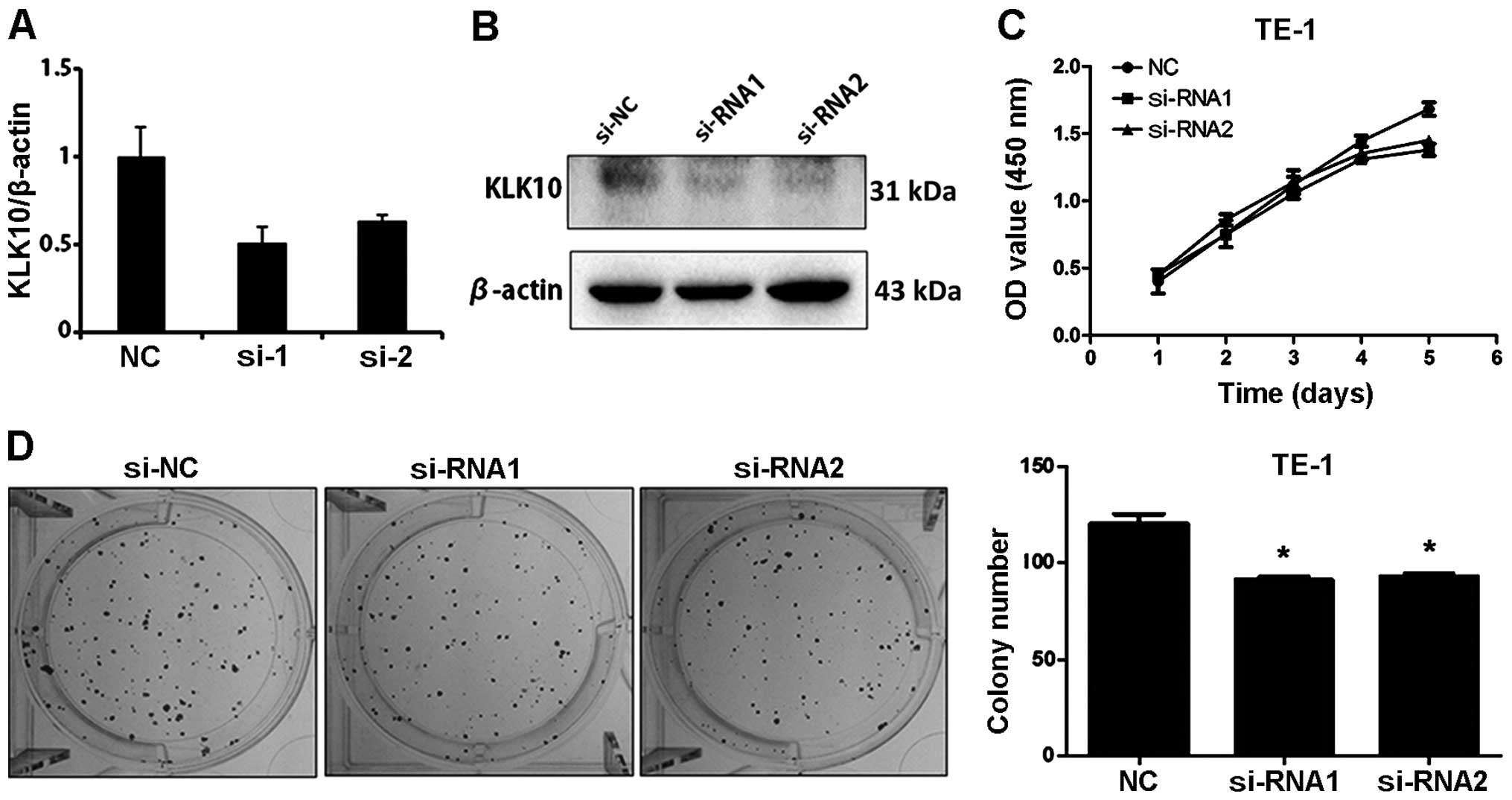
colony formation in KLK10 knockdown TE-1 cells (Fig. 3).
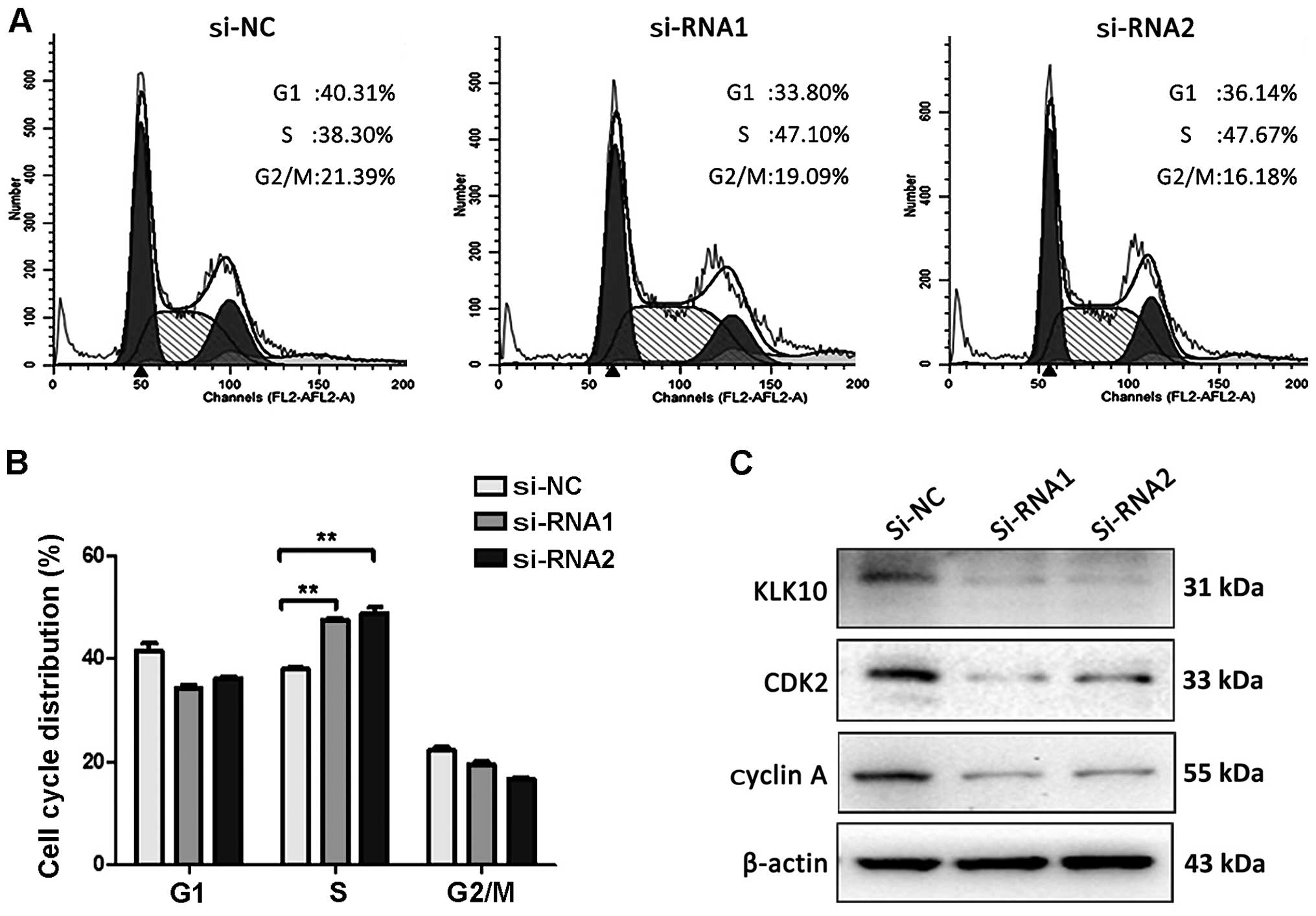
Knockdown of KLK10 leads to S-phase cell
cycle arrest and enhancement of cell cycle regulatory proteins in
Eca-109 cells
Based on the above results, we determined whether
inhibition of cell proliferation after knockdown of KLK10 in
Eca-109 cells was associated with cell cycle regulation. For this
purpose, the cells were harvested and subjected to cell cycle
analysis after transfected for 48 h. We found that the Eca-109
cells were arrested into S phase from 38.3 to 47.1 (siRNA-1) and
47.67% (siRNA-2) in the KLK10 siRNA groups (P<0.01) compared to
the control (Fig. 4A). Regulatory
proteins for S-phase cell cycle were further examined as shown in
Fig 4C, and the knockdown of KLK10
in Eca-109 cells resulted in the reduction of cyclin A and CDK2
expression.
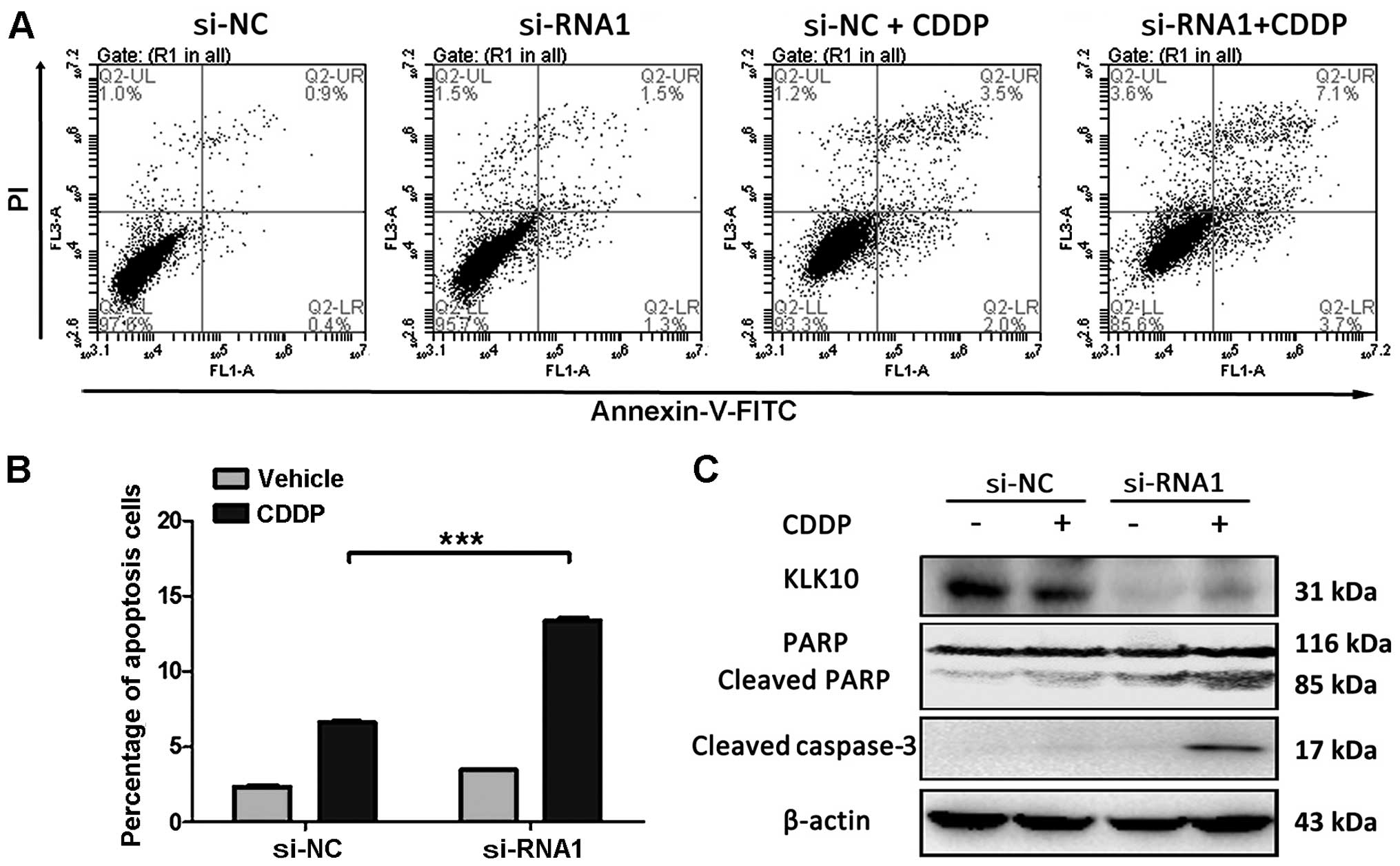
Knockdown of KLK10 promotes CDDP-induced
apoptosis and activation of caspase-3
CDDP is regarded as a first-line chemotherapy for EC
patients, but the drug resistance remains a major clinical
challenge (4). Therefore, we
studied whether knockdown of KLK10 in Eca-109 cells increased the
sensitivity to CDDP. siRNA1 was selected as its higher knockdown
efficiency. Annexin V/PI staining and FACS analysis showed that
apoptosis in KLK10-silenced Eca-109 cells increased to 10.8% as
compared to the control (P<0.001) in response to CDDP. Western
blot analysis revealed that CDDP induces apoptosis via knockdown of
KLK10, significantly increasing protein levels of active caspase-3
and cleaved form of PARP as compared to the control (Fig. 5).
Discussion
Esophageal cancer is difficult to diagnose, and
frequently has a poor prognosis. Therefore, it is of importance to
identify new candidate markers to facilitate the early diagnosis
and treatment of EC (27).
KLK10 codes a secreted serine protease, whose
physiological function remains unknown. Compelling evidence has
indicated that the expression of KLK10 is associated with the
development of severe malignant tumors including steroid
hormone-related cancers and some gastrointestinal tumors (28). KLK10 is considered to be a biomarker
of some malignant tumors. In breast cancer, KLK10 is deregulated
and acts as a tumor suppressor (14). KLK10 may enhance diagnosis
sensitivity and specificity when cooperating with CA125. In NSCLC,
ectopic KLK10 reduces anchorage-independent growth, tumorigenicity
and enhances drug sensitivity (15), whereas in other tumors, such as
prostate cancer, KLK10 acts as a tumor promoter. Knockdown of KLK10
inhibits proliferation and tumorigenicity. In the present study, we
aimed to examine the expression and bio-function of KLK10 in
esophageal cancer.
Previous findings have shown that a high expression
of KLK10 in gastric (29,30) and colorectal cancer (11,31) is
associated with poor prognosis. We hypothesized whether the
expression of KLK10 is abnormal in esophageal cancer. Our results
show an 80.72% positive expression in human EC for KLK10, but only
27.27% in normal tissues. Further analyses revealed that KLK10
expression was inversely correlated with tumor diameter (P=0.046,
Chi-square). The diameter of the esophageal cancer is closely
associated with the prognosis of patients (32,33).
Accordingly, we demonstrated KLK10 is a potential biomarker for the
diagnosis of esophageal cancer.
We observed the expression of KLK10 in mRNA and
protein levels were different between TE-1 and Eca-109 cell lines
used in the present study. TE-1 cells had a higher mRNA expression
level of KLK10, but a relatively lower KLK10 protein expression
level, which may be caused by the frequent regulation at the
transcriptional and post-transcriptional levels by miRNAs (28), as some microRNAs are directly
targeted by KLK10. KLK10 expression may be epigenetically
influenced according to the results. Therefore, mRNA expression
does not represent the protein level of KLK10 in TE-1 cells
transfected with small interference RNAs. Furthermore, inhibition
in the proliferation and colony formation ability of the KLK10
knockdown groups was not obvious. The results suggest that in
certain esophageal carcinoma with a KLK10 high expression, KLK10 is
a potential target for therapy.
DNA synthesis and internal checkpoints during S
phase are activated when DNA double strands are damaged, such as in
fracture (34). ATM/ATR-mediated
Cdc25A degradation acting on cyclin E/A-CDK2 is primarily
responsible for S-phase progression (34). We found that knockdown of KLK10
induced S-phase arrest by flow cytometry and the cyclin A and CDK2
activity was also decreased in Eca-109 cells. Thus, cell cycle
arrest may be caused by cell proliferation inhibition due to
knockdown of KLK10.
Cisplatin resistance remains a serious clinical
challenge in the post-esophageal cancer (5). The results of the present study
suggest that, knockdown of KLK10 can increase the cisplatin
sensitivity of esophageal cancer as the apoptotic ratio was
significantly higher in the KLK10 knockdown group. Therefore, KLK10
is a potential target for the treatment of esophageal cancer by
reversing chemotherapy resistance in esophageal cancer
patients.
In conclusion, to the best of our knowledge, this is
the first study to report that KLK10 was highly expressed in human
EC and that KLK10 is a potential biomarker for EC diagnosis at an
early stage. Knockdown of KLK10 inhibits EC cell proliferation and
reverses chemotherapy resistance in vitro. Future studies
should examine more tissues and in vivo assays to gain
insight into the detailed mechanisms of KLK10-derived EC diagnosis
and therapy.
Acknowledgments
The present study was supported by grants from the
National Natural Science Foundation of China (81102069).
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dawsey SP, Tonui S, Parker RK, Fitzwater
JW, Dawsey SM, White RE and Abnet CC: Esophageal cancer in young
people: A case series of 109 cases and review of the literature.
PLoS One. 5:e140802010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang HY, Spechler SJ and Souza RF:
Esophageal adenocarcinoma arising in Barrett esophagus. Cancer
Lett. 275:170–177. 2009. View Article : Google Scholar :
|
|
4
|
Hong J, Peng D, Chen Z, Sehdev V and
Belkhiri A: ABL regulation by AXL promotes cisplatin resistance in
esophageal cancer. Cancer Res. 73:331–340. 2013. View Article : Google Scholar
|
|
5
|
Grünberger B, Raderer M, Schmidinger M and
Hejna M: Palliative chemotherapy for recurrent and metastatic
esophageal cancer. Anticancer Res. 27(4C): 2705–2714.
2007.PubMed/NCBI
|
|
6
|
Lin C, Song L, Gong H, Liu A, Lin X, Wu J,
Li M and Li J: Nkx2-8 downregulation promotes angiogenesis and
activates NF-κB in esophageal cancer. Cancer Res. 73:3638–3648.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Noel A, Maillard C, Rocks N, Jost M,
Chabottaux V, Sounni NE, Maquoi E, Cataldo D and Foidart JM:
Membrane associated proteases and their inhibitors in tumour
angiogenesis. J Clin Pathol. 57:577–584. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Borgoño CA and Diamandis EP: The emerging
roles of human tissue kallikreins in cancer. Nat Rev Cancer.
4:876–890. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lawrence MG, Lai J and Clements JA:
Kallikreins on steroids: Structure, function, and hormonal
regulation of prostate-specific antigen and the extended kallikrein
locus. Endocr Rev. 31:407–446. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Luo LY, Grass L, Howarth DJ, Thibault P,
Ong H and Diamandis EP: Immunofluorometric assay of human
kallikrein 10 and its identification in biological fluids and
tissues. Clin Chem. 47:237–246. 2001.PubMed/NCBI
|
|
11
|
Talieri M, Alexopoulou DK, Scorilas A,
Kypraios D, Arnogiannaki N, Devetzi M, Patsavela M and Xynopoulos
D: Expression analysis and clinical evaluation of
kallikrein-related peptidase 10 (KLK10) in colorectal cancer.
Tumour Biol. 32:737–744. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Michael IP, Kurlender L, Memari N, Yousef
GM, Du D, Grass L, Stephan C, Jung K and Diamandis EP: Intron
retention: A common splicing event within the human kallikrein gene
family. Clin Chem. 51:506–515. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu XL, Wazer DE, Watanabe K and Band V:
Identification of a novel serine protease-like gene, the expression
of which is down-regulated during breast cancer progression. Cancer
Res. 56:3371–3379. 1996.PubMed/NCBI
|
|
14
|
Sidiropoulos M, Pampalakis G, Sotiropoulou
G, Katsaros D and Diamandis EP: Downregulation of human kallikrein
10 (KLK10/NES1) by CpG island hypermethylation in breast, ovarian
and prostate cancers. Tumour Biol. 26:324–336. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang Y, Song H, Miao Y, Wang R and Chen
L: Frequent transcriptional inactivation of kallikrein 10 gene by
CpG island hypermethylation in non-small cell lung cancer. Cancer
Sci. 101:934–940. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
White NM, Bui A, Mejia-Guerrero S, Chao J,
Soosaipillai A, Youssef Y, Mankaruos M, Honey RJ, Stewart R, Pace
KT, et al: Dysregulation of kallikrein-related peptidases in renal
cell carcinoma: Potential targets of miRNAs. Biol Chem.
391:411–423. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Treeck O, Schüler S, Häring J, Skrzypczak
M, Lattrich C and Ortmann O: icb-1 gene counteracts growth of
ovarian cancer cell lines. Cancer Lett. 335:441–446. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Santin AD, Diamandis EP, Bellone S,
Marizzoni M, Bandiera E, Palmieri M, Papasakelariou C, Katsaros D,
Burnett A and Pecorelli S: Overexpression of kallikrein 10 (hK10)
in uterine serous papillary carcinomas. Am J Obstet Gynecol.
194:1296–1302. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rückert F, Hennig M, Petraki CD, Wehrum D,
Distler M, Denz A, Schröder M, Dawelbait G, Kalthoff H, Saeger HD,
et al: Co-expression of KLK6 and KLK10 as prognostic factors for
survival in pancreatic ductal adenocarcinoma. Br J Cancer.
99:1484–1492. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang YY, Lin YC, Hung HC, Tien WY and
Shieh TY: Polymorphisms in kallikrein7 and 10 genes and oral cancer
risks in Taiwan betel quid chewers and smokers. Oral Dis.
19:824–832. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pettus JR, Johnson JJ, Shi Z, Davis JW,
Koblinski J, Ghosh S, Liu Y, Ravosa MJ, Frazier S and Stack MS:
Multiple kallikrein (KLK 5,7,8, and 10) expression in squamous cell
carcinoma of the oral cavity. Histol Histopathol. 24:197–207.
2009.
|
|
22
|
Luo LY, Grass L and Diamandis EP: Steroid
hormone regulation of the human kallikrein 10 (KLK10) gene in
cancer cell lines and functional characterization of the KLK10 gene
promoter. Clin Chim Acta. 337:115–126. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li B, Goyal J, Dhar S, Dimri G, Evron E,
Sukumar S, Wazer DE and Band V: CpG methylation as a basis for
breast tumor-specific loss of NES1/kallikrein 10 expression. Cancer
Res. 61:8014–8021. 2001.PubMed/NCBI
|
|
24
|
Bin L, Kim BE, Hall CF, Leach SM and Leung
DY: Inhibition of transcription factor specificity protein 1 alters
the gene expression profile of keratinocytes leading to
upregulation of kallikrein-related peptidases and thymic stromal
lymphopoietin. J Invest Dermatol. 131:2213–2222. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hao XP, Pretlow TG, Rao JS and Pretlow TP:
Beta-catenin expression is altered in human colonic aberrant crypt
foci. Cancer Res. 61:8085–8088. 2001.PubMed/NCBI
|
|
26
|
Ling S, Feng T, Ke Q, Fan N, Li L, Li Z,
Dong C, Wang C, Xu F, Li Y, et al: Metformin inhibits proliferation
and enhances chemosensitivity of intrahepatic cholangiocarcinoma
cell lines. Oncol Rep. 31:2611–2618. 2014.PubMed/NCBI
|
|
27
|
Li X, Suo J, Shao S, Xue L, Chen W, Dong
L, Shi J, Fu M, Lu N, Zhan Q, et al: Overexpression of OLC1
promotes tumorigenesis of human esophageal squamous cell carcinoma.
PLoS One. 9:e909582014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
White NM, Chow TF, Mejia-Guerrero S,
Diamandis M, Rofael Y, Faragalla H, Mankaruous M, Gabril M, Girgis
A and Yousef GM: Three dysregulated miRNAs control kallikrein 10
expression and cell proliferation in ovarian cancer. Br J Cancer.
102:1244–1253. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jiao X, Lu HJ, Zhai MM, Tan ZJ, Zhi HN,
Liu XM, Liu CH and Zhang DP: Overexpression of kallikrein gene 10
is a biomarker for predicting poor prognosis in gastric cancer.
World J Gastroenterol. 19:9425–9431. 2013. View Article : Google Scholar
|
|
30
|
Kolin DL, Sy K, Rotondo F, Bassily MN,
Kovacs K, Brezden-Masley C, Streutker CJ and Yousef GM: Prognostic
significance of human tissue kallikrein-related peptidases 6 and 10
in gastric cancer. Biol Chem. 395:1087–1093. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Alexopoulou DK, Papadopoulos IN and
Scorilas A: Clinical significance of kallikrein-related peptidase
(KLK10) mRNA expression in colorectal cancer. Clin Biochem.
46:1453–1461. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang BY, Goan YG, Hsu PK, Hsu WH and Wu
YC: Tumor length as a prognostic factor in esophageal squamous cell
carcinoma. Ann Thorac Surg. 91:887–893. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang BY, Liu CY, Lin CH, Hsu PK, Hsu WH,
Wu YC and Cheng CY: Endoscopic tumor length is an independent
prognostic factor in esophageal squamous cell carcinoma. Ann Surg
Oncol. 19:2149–2158. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Eastman A: Cell cycle checkpoints and
their impact on anticancer therapeutic strategies. J Cell Biochem.
91:223–231. 2004. View Article : Google Scholar : PubMed/NCBI
|