Introduction
Tumor-associated antigen (TAAs)-specific T cells are
detected in many patients with cancer (1,2).
However, these TAA-specific T cells typically fail to control tumor
growth. It is expected that vaccination with TAA enhances
tumor-specific T cell responses, improves antitumor immunity and
provides clinical benefit. The development of cancer vaccines has
been greatly potentiated by the availability of synthetic
tumor-antigen-peptides, although progress has been hampered by the
poor immunogenicity of antigen-peptides.
Liposome has been proposed to be a carrier of
antigens and an adjuvant to induce immune responses (3). We previously reported that antigens
chemically coupled to the surface of liposomes consisting of
unsaturated fatty acids were cross-presented by antigen-presenting
cells (APCs) to cytotoxic T lymphocytes (CTLs) (4). Moreover, we have shown that single
immunization of as low as 280 ng of liposomal peptide along with
CpG conferred complete protection to the immunized mice from a
highly virulent mutant strain, clone 13 of lymphocytic
choriomeningitis virus (LCMV). In addition, surface-linked
liposomal antigens induced long-lived memory CD8+ T
cells in the absence of CD4+ T-cell assistance (5,6). This
form of CTL-based liposomal vaccine was also investigated for tumor
vaccine using an OVA-expressing mouse tumor model. This
investigation suggested surface-coupled liposomal antigens may be
applicable for the development of tumor vaccines (4). However, to the best of our knowledge,
the possibility of a vaccine targeting endogenous tumor antigen by
surface-coupled liposomal antigens remains to be examined.
Telomerase reverse transcriptase (TERT) is a tumor
antigen that is highly expressed in >85% of all human types of
cancer including stem cell-like tumor cells (7–10).
Therefore, TERT antigens have the potential to mediate antitumor
immune responses against a range of tumors (11). In previous studies, epitope peptides
derived from TERT have been shown to elicit CTL responses capable
of recognizing TERT-expressing tumor cells in vitro
(7,8,12).
However, reports of antitumor efficiency in vivo using
natural TERT peptide are limited.
Self-antigen peptides do not necessarily undergo
antigen processing/presentation and serve as dominant epitopes. In
certain situations, the peptides are hidden from immune recognition
and termed 'cryptic' (13).
Dominant peptides show a high affinity for MHC molecules and are
frequently presented by somatic cells, whereas, cryptic peptides
show a low affinity for MHC molecules and are infrequently
presented by somatic cells. Similar to many other TAAs, TERT is a
self-antigen. Therefore, it is thought that central tolerance
against TERT-derived peptides with high affinity for MHC molecules
exists. The central tolerance against TAAs is deemed to be a major
barrier for TAA-specific antitumor immunotherapy. On the other
hand, the CTL repertoire specific for cryptic epitope may escape
from the negative selection process in central tolerance.
Therefore, CTLs against cryptic epitopes may be available for
antitumor immunotherapy. However, it is difficult to identify
cryptic epitopes for immunotherapy due to their poor
immunogenicity. To circumvent this problem, previous investigators
have created modified heteroclitical peptides to enhance their
affinity for HLA-A2 molecules while preserving the amino acids that
interact with the T-cell receptor (TCR), which is specific to the
original cryptic epitope (14–17).
Previous findings showed that vaccination with heteroclitical
peptide derived from cryptic epitopes of TERT inhibited the growth
of tumors in vivo (15,23).
The aim of the present study was to determine the
possibility that the liposome coupled with a peptide derived from
TERT may serve as an effective CTL-based vaccine against tumor.
First, to identify the optimum peptide for coupling on the surface
of liposomes, 20 TERT-derived peptides with HLA-A2.1 binding motif
were selected by computer algorism, the liposome-coupled peptide
was prepared and their ability to provoke peptide specific CD8 T
cell responses in HLA-A*0201-transgenic (HHD) mice was
determined. We identified a novel cryptic epitope with low affinity
for HLA molecules and found that immunization using
liposome-coupled heteroclitical peptide derived from this novel
cryptic epitope elicits CD8 T-cell responses against native peptide
and efficiently inhibits the growth of TERT expressing tumors in
HHD mice. Our results strongly support the use of liposomal vaccine
surface coupled with TERT antigens as a new potential option for
active immunotherapy against tumors.
Materials and methods
Prediction of CTL epitopes
To define potential HLA-A*0201-binding
peptides from human TERT (hTERT), a computer-based program, BIMAs
(http://www-bimas.cit.nih.gov/molbio/hla_bind/), was
used. As shown by the odd numbers in Table I, 10 peptides that have superior
Bimas scores and homologous sequence in murine TERT (mTERT) were
selected for predicted CTL epitopes.
 | Table IBimas score, HLA-A2.1 affinity and
stabilization capacity of hTERT peptides. |
Table I
Bimas score, HLA-A2.1 affinity and
stabilization capacity of hTERT peptides.
| Peptide no.a | Start position | Subsequenceb | BIMAs scorec | RAd | DC50
(h)e |
|---|
| 1 | 540 | ILAKFLHWL | 1745.714 | 2 | 8 |
| 2 | |
ILAKFLHWV | 5683.72 | 2 | >8 |
| 3 | 572 | RLFFYRKSV | 257.342 | 32 | <2 |
| 4 | | YLFFYRKSV | 1183.775 | 4 | 4 |
| 5 | 943 | LLQAYRFHA | 48.984 | 8 | 4 |
| 6 | |
LLQAYRFHV | 685.783 | 4 | >8 |
| 7 | 926 | LQVNSLQTV | 27.573 | 4 | >8 |
| 8 | | LLVNSLQTV | 271.948 | 2 | >8 |
| 9 | 579 | SVWSKLQSI | 17.427 | 32 | 2 |
| 10 | | SLWSKLQSI | 199.162 | 8 | >8 |
| 11 | 945 | QAYRFHACV | 11.747 | 8 | 4 |
| 12 | | QLYRFHACV | 845.752 | 4 | 6 |
| 13 | 944 | LQAYRFHAC | 1.627 | 32 | 4 |
| 14 | |
LQAYRFHAV | 22.777 | 2 | >8 |
| 15 | 23 | PLATFVRRL | 0.822 | >32 | 4 |
| 16 | | YLATFVRRL | 171.771 | 4 | >8 |
| 17 | 847 | GDMENKLFA | 0.576 | >32 | <2 |
| 18 | | GLMENKLFA | 414.505 | >32 | >8 |
| 19 | 925 | DLQVNSLQT | 0.559 | >32 | <2 |
| 20 | | YLQVNSLQT | 34.279 | >32 | <2 |
Modified heteroclitical peptides were produced to
enhance their affinity for HLA-A2 molecules while preserving the
amino acids that interact with the TCR as previously described
(18–21). Heteroclitical modifications were
performed by replacing the amino acid at position 1 with a tyrosine
(Y), position 2 with a leucine (L) or position 9 with a valine (V).
Heteroclitical peptides are shown by the even numbers in Table I. These peptides were synthesized by
Eurofins genomics (Tokyo, Japan).
Animals
The HLA-A*0201-transgenic HHD mice (a
kind gift from Dr F.A. Lemonnier, pasteur Institute, paris, France)
expressed a transgenic HLA-A*0201 monochain in which
human β-2 microglobulin (β2m) is covalently linked to a chimeric
heavy chain composed of HLA-A*0201 (α1 and α2, domains)
and H-2Db (α3, transmembrane and cytoplasmic domains) (18). Six- to 12-week-old mice were used
for all the experiments. The mice were housed in appropriate animal
care facilities at Saitama Medical University (Saitama, Japan) and
handled according to international guidelines. Experimental
protocols were approved by the Animal Research Committee of Saitama
Medical University.
Measurement of peptide relative affinity
(RA) for HLA-A2.1
T2 cells (3×105/ml) were incubated with
various concentrations of peptides ranging from 100 to 0.1
µM in serum-free RpMI-1640 medium (R0) supplemented with 100
ng/ml human β2m (Acris Antibodies, San Diego, CA, USA) at 37°C for
16 h, as previously described (16). The cells were then stained with the
HLA-A2.1 molecule-specific BB7.2 mAb (BioLegend, San Diego, CA,
USA). For each peptide concentration, HLA-A2.1-specific staining
was calculated as the percentage of the staining obtained with 100
µM of the reference peptide HIVpol589 (IVGAETFYV). The RA
was determined as: RA = concentration of each peptide that induces
20% of HLA-A2.1 expression/concentration of the reference peptide
that induces 20% of HLA-A2.1 expression. Lower RA value shows
higher binding affinity for HLA-A2.1.
Assessment of peptide/HLA-A2.1 complex
stability
T2 cells were incubated with 100 µM of each
peptide in R0 supplemented with 100 ng/ml human β2m at 37°C for 16
h, as previously described (16).
The cells were then washed four times to remove free peptides,
incubated with 0.2% GoldiPlug (BD Biosciences Pharmingen, san
Diego, CA, USA) in R0 for 1 h to block the cell surface expression
of newly synthesized HLA-A2.1. After washing and incubation for 0,
2, 4, 6 or 8 h, the cells were stained with anti-HLA-A2.1 antibody
(BB7.2). For each time-point, the peptide-induced HLA-A2.1
expression was calculated as: mean fluorescence of
peptide-preincubated T2 cells - mean fluorescence of T2 cells
treated in similar conditions in the absence of peptide.
DC50 (dissociation complex, DC) was defined as the time
required for the loss of 50% of the HLA-A2.1/peptide complexes
stabilized at t=0.
Cell lines and telomerase detection
Mouse lymphoma cell lines RMA and RMA-HHD, which
were a kind gift from Dr F.A. Lemonnier (22) were cultured in RpMI-1640 medium
supplemented with 10% fetal bovine serum (FBs) (R-10). Telomerase
detection in tumor cells was achieved by flow cytometry as
previously reported using anti-hTERT monoclonal antibody (H-231;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA), which cross
reacts with mTERT (23).
Liposome
The liposomes used in the present study were
provided by NOF Corporation (Tokyo, Japan), and consisted of
dioleoyl phosphatidylcholine (DOPC), dioleoyl
phosphatidyl-ethanolamine (DOPE), dioleoyl phosphatidyl glycerol
(DOPG) and cholesterol in a 4:3:2:7 molar ratio. The crude liposome
solution was passed through a membrane filter with a pore size of
0.2 mm.
Coupling of peptide to liposomes
Liposomal conjugates with TERT peptides were
prepared essentially in the same manner as previously described
(6). The final preparations
contained ≤0.7 mg of the peptide and 10 mg of the liposome/ml
(24).
Immunization
The mice were immunized with indicated doses of
liposomal peptides via subcutaneous injection in the presence of 5
µg/mouse CpG or 100 µg of peptide emulsified in 250
µl of IFA with 100 µg of peptide HBV core 128–140.
These immunizations were performed on day 0 and 7.
Intracellular IFN-γ and CD107a
degranulation analysis
After 5 days following immunization, spleen cells of
immunized mice were incubated with brefeldin A (GolgiPlug; BD
Biosciences) and fluorescein isothiocyanate (FITC)-conjugated
monoclonal antibody (mAb) to CD107a for 5 h at 37°C in the presence
or absence of a relevant peptide. After the incubation, the cells
were washed, labeled with peridinin chlorophyll protein complex
(PerCP)-conjugated anti-CD8α mAb, fixed and permeabilized
(Cytofix-Cytoperm; BD Biosciences) before they were labeled with
phycoerythrin (PE)-conjugated anti-IFN-γ mAb. All the antibodies
were purchased from Biolegend. The stained cells were analyzed on a
FACSCant II flow cytometer (BD Biosciences).
HLA-A2.1 dextramer analysis
To quantify antigen-specific T cells, the spleen
cells were incubated with allophycocyanin (APC)-conjugated HLA
dextramer recognizing 944v peptide derived from hTERT
(944v/dextramer; Immudex, Copenhagen, Denmark) for 20 min at room
temperature. After the incubation, the cells were washed, labeled
with PerCP-conjugated anti-CD8α mAb. The stained cells were
analyzed on a FACSCant II flow cytometer (BD Biosciences).
CTL generation and cytotoxic assay
After 5 days following immunization, the spleen
cells of immunized mice were cultured for 6 days with irradiated
(30 Gy), syngeneic naïve spleen cells pre-pulsed with a relevant
peptide and employed as effector cells in CTL-mediated cytotoxic
assay. TERT-positive RMA-HHD cells were used as target cells.
CTL-mediated cytotoxic assay was performed using a LIVE/DEAD
Cell-mediated Cytotoxicity Assay kit (Life Technologies, Carlsbad,
CA, USA) according to the manufacturer's instructions.
In vivo tumor protection
The HHD mice were vaccinated with liposomal peptide
with 5 µg of Cpg on day 14, 7 and 0. On day 0, the mice were
challenged subcutaneously with 2×104 RMA-HHD cells.
Tumor size was monitored every 3 to 4 days using calipers and the
mice were sacrificed when the tumor mass reached >1,000
mm2 or tumor tissue disintegration occurred.
Statistical analysis
Data were presented as the mean ± SD. Statistical
comparison between groups was based on the Mann-Whitney U test
using Microsoft Excel. Mouse survival time was estimated using the
Kaplan-Meier method and the Log-rank test. P<0.05 was considered
significant.
Results
Selection of CTL epitopes derived from
hTERT for coupling to liposomes
Ten native peptides derived from hTERT were selected
by the BIMAs score (odd numbers in Table I). To improve the binding affinity
for HLA-A2.1, these peptides were subjected to amino acid
substitutions as shown in Table I
(even numbers). Most of the heteroclitical peptides improved their
RA and DC50 value compared with the native peptides
(Table I). To assess the
immunogenicity, these peptides were pooled into four groups
consisting of five peptides (Table
II) and coupled to the surface of liposomes. Each liposomal
peptide pool was subsequently injected into the HHD mice. After 5
days following immunization, the spleen cells of immunized mice
were prepared, stimulated with a relevant peptide and stained for
their expression of surface CD8 and intracellular IFN-γ and
degranulation of CD107a. As shown in Fig. 1, three heteroclitical peptides on
the surface of liposomes (Lip-#8, -#10 and-#14) were able to induce
a significant amount of IFN-γ+ CD107a+
CD8+T cells and the responses were much higher compared
to those induced by the original peptides (Lip-#7, -#9 and-#13).
Lip-#14 was most effective for the induction of peptide-specific
IFN-γ+CD107a+ CD8+ T cells. Thus,
#14 (p944v), the heteroclitical peptide derived from #13 (p944),
was selected as a candidate epitope.
| Table IIContents of peptide groups. |
Table II
Contents of peptide groups.
| Peptide groups | Peptide no. |
|---|
| 1 | 1, 5, 9, 13,
17 |
| 2 | 3, 7, 11, 15,
19 |
| 3 | 2, 6, 10, 14,
18 |
| 4 | 4, 8, 12, 16,
20 |
The heteroclitical peptide coupled to liposome
(Lip-#14) induces a strong CD8 T-cell response with cross
recognition to the native epitope and in vivo primed CTLs
with Lip-#14 recognizes endogenously mTERT-expressing tumor
cells.
The #13 peptide (p944) and #14 peptide (p944v) were,
respectively, coupled to the surface of liposomes (Lip-#13 and
-#14). These liposomal peptides were subsequently injected into the
HHD mice and an intracellular IFN-γ and CD107a degranulation
analysis was conducted to assess antigen-specific CD8 T-cell
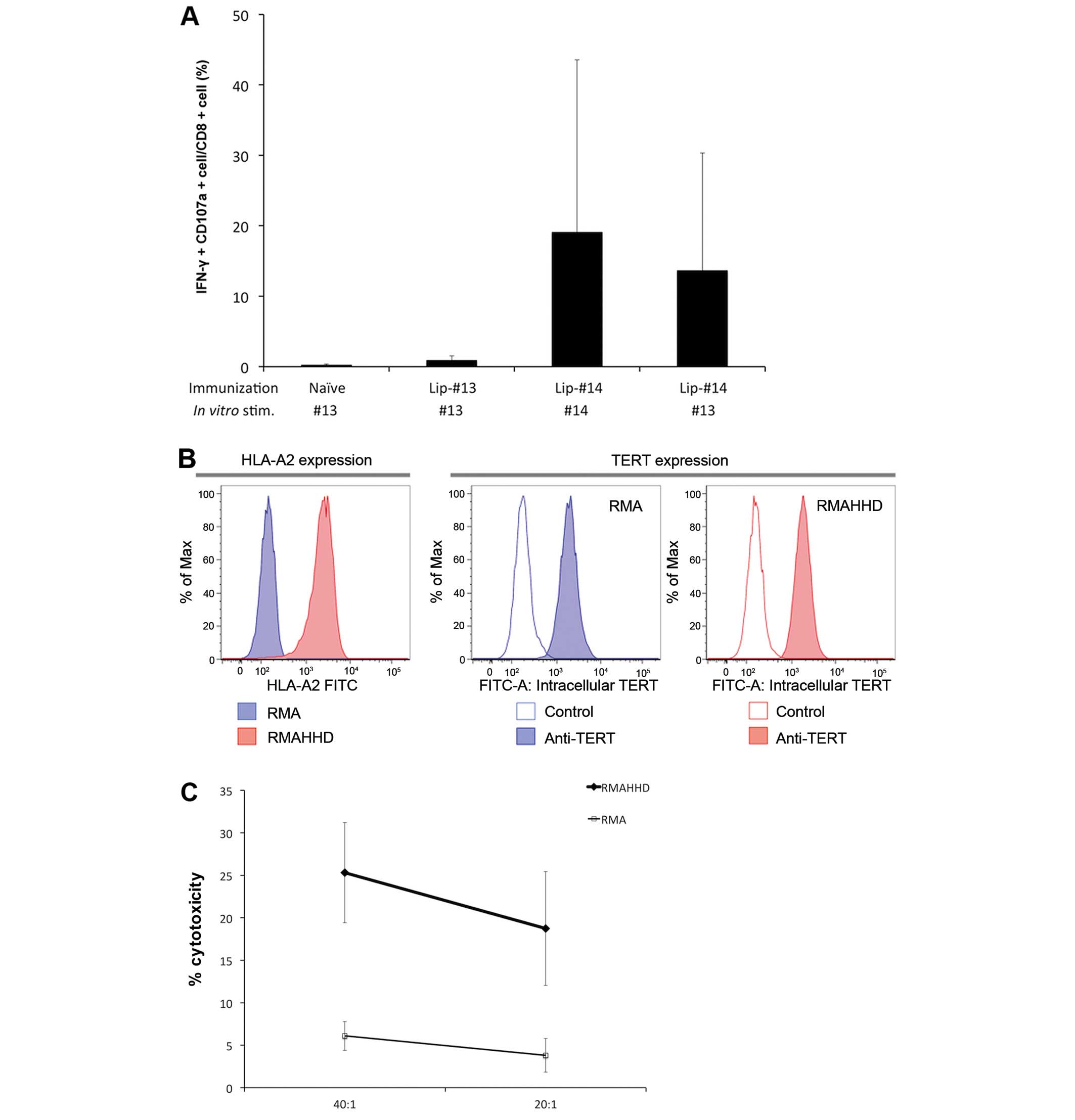
responses against the native epitope #13 (p944). As shown in
Fig. 2A, Lip-#14 induced strong CD8
T-cell responses in vivo. Compared with immunization with
liposomal native #13 peptide, the magnitude of CD8 responses
induced by liposomal #14 peptide was extremely strong. The results
also showed that CD8 T cells that derived from Lip-#14-immunized
mice crossreacted with the native epitope #13 equally to the
heteroclitic epitope #14.
Additionally, to evaluate whether #13 peptide is
naturally processed and presented by tumor cells, #14-specific CTLs
were tested for their ability to lyse endogenously mTERT-expressing
HLA-A2.1+ tumor cells. CTLs specific for #14 were
generated from the spleen cells of mice immunized with Lip-#14 and
their cytolytic activity was tested against RMA-HHD mouse lymphoma
cells. Fig. 2B shows that RMA-HHD
cells express mTERT endogenously while also possessing sufficient
cell surface HLA-A2.1. We found that CTLs specific for #14 were
able to lyse RMA-HHD cells, but no cytotoxicity was observed
against HLA-A2.1-negative RMA cells (Fig. 2C). These results indicated that #13,
the low affinity cryptic epitope, is naturally processed and
presented by HLA-A2.1 molecules on the surface of TERT-expressing
tumor cells and CTLs primed in vivo with Lip-#14
immunization are able to recognize and kill tumor cells in an
HLA-A2.1-restricted manner.
Liposomal #14 peptide induces a stronger
antigen-specific CD8 T-cell response than peptide + IFA
immunization
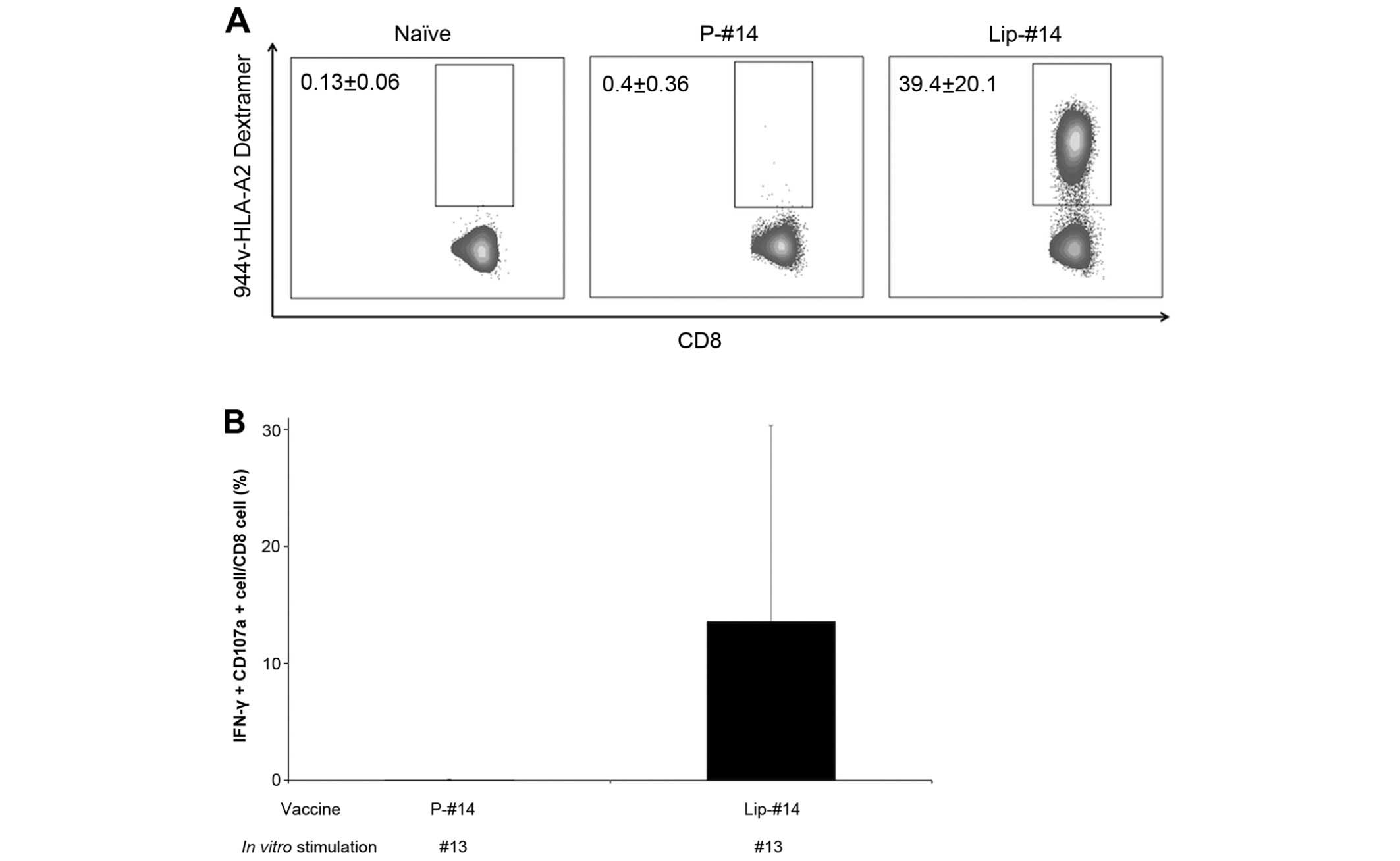
Two vaccines were produced: Lip-#14 and a #14
peptide emulsified in IFA with a MHC class II pan T-helper peptide,
HBVcore128-140 (24) (p-#14). The
immunogenicity of these two forms of TERT 944v was compared by
analyzing spleen cells from the immunized mice using a TERT
944v-HLA-A2 dextramer assay (Fig.
3A), intracellular IFN-γ and CD107a degranulation analysis
(Fig. 3B). We found that an
immunization with Lip-#14 induced greater antigen-specific CD8 T
cell response compared with the p-#14 vaccination. The frequencies
of TERT 944v-HLA-A2 dextramer+ CD8 T cells detected in
HHD mice were ~100-fold higher after Lip-#14 vaccination, compared
to p-#14 vaccination (39.4±20.1 vs. 0.4±0.36%). As dextramer
analysis did not discriminate between anergic and functional T
cells, we also monitored an antigen-specific response by
intracellular IFN-γ and CD107a degranulation analysis. In agreement
with results from the dextramer analysis, the frequencies of
intracellular IFN-γ and CD107a+ CD8 T cells in response
to native the #13 epitope detected in Lip-#14-vaccined HHD mice
were in contrast with the p-#14-vaccined group (Fig. 3B). These results demonstrated that
vaccination with surface-coupled liposomal antigen, which was
modified to enhance their affinity for class I molecules causes
strong immune responses against an epitope that cannot induce
immune responses by ordinary vaccination such as peptide with
IFA.
Previous reports suggest that the efficacy of active
immunotherapy may be associated with the capacity of the vaccine to
promote sustained CD8 T-cell responses in vivo (25). Therefore, we investigated whether
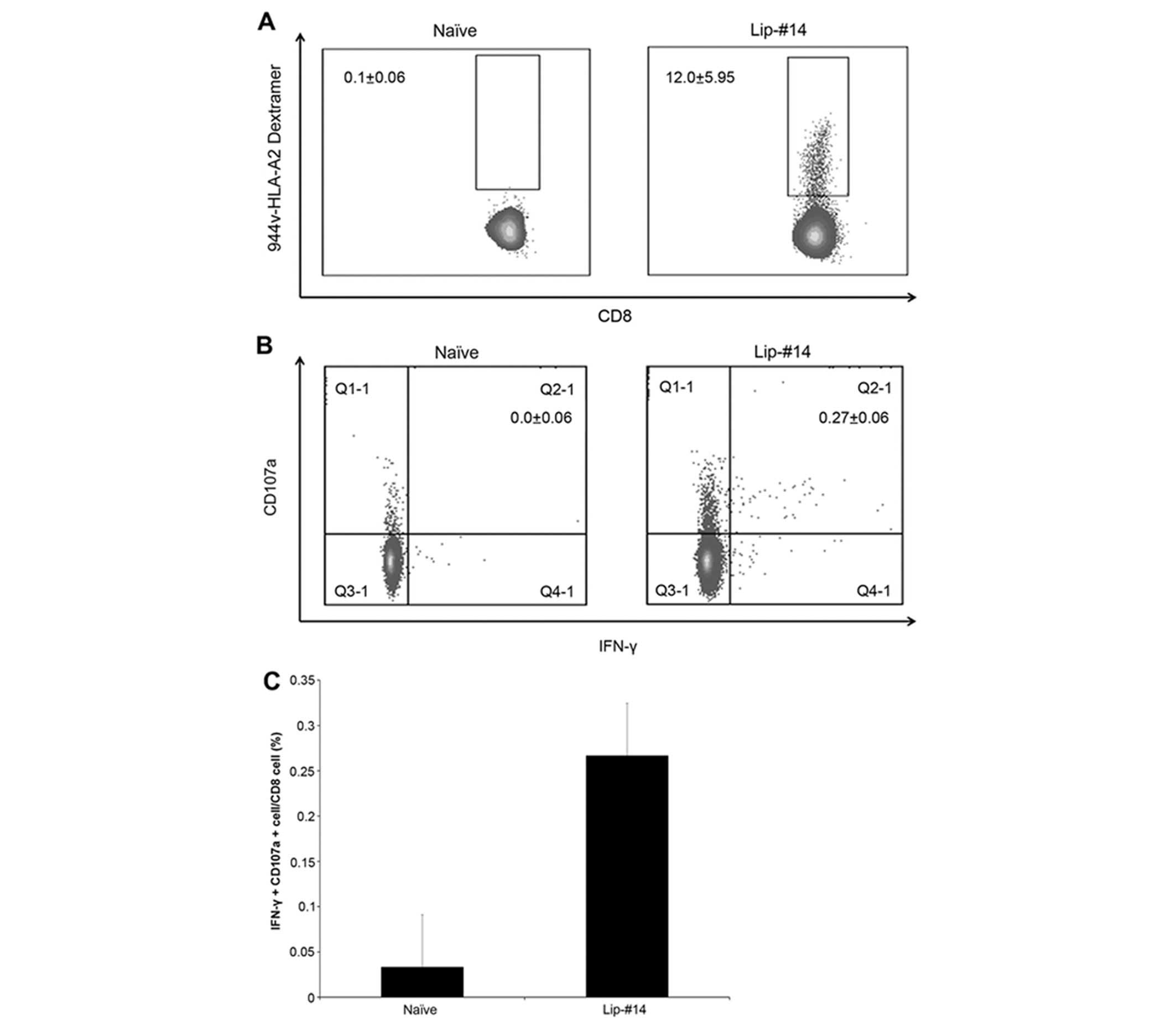
long-lasting #14-specific and #13-crossreactive CD8 T cells could
be elicited in HHD mice immunized with Lip-#14. After 35 days
following immunization, the CD8 responses were measured using a
dextramer assay and flow cytometric analysis using intracellular
IFN-γ and CD107a degranulation. As shown in Fig. 4A, TERT #14-specific CD8 T cells were
detected in mice on day 35 after immunization with Lip-#14. When
the long-lasting CD8 T-cell responses were evaluated by
intracellular IFN-γ and CD107a degranulation assay, we found that
significant IFN-γ+CD107a+CD8 T cells in
response to native #13 peptide can only be detected in HHD mice
injected with Lip-#14 (Fig. 4B and
C). These results suggested that immunization with Lip-#14 is
capable of effectively generating long-lasting memory CD8 T cell
and sustained CD8 T-cell responses against native cryptic
epitope.
Immunization with Lip-#14 induces tumor
growth inhibition
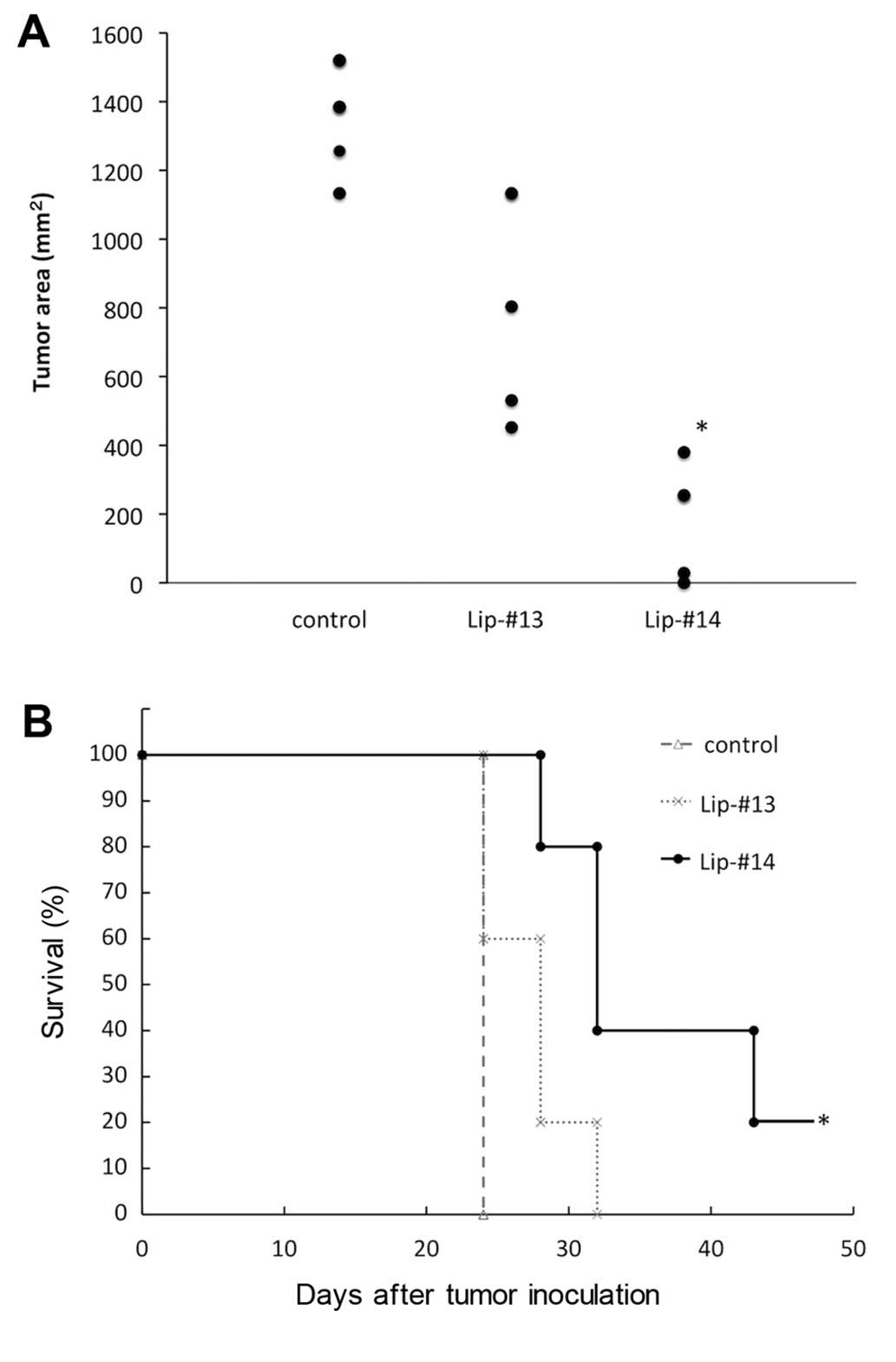
To investigate the antitumor effect of Lip-#14, we
immunized HHD mice with Lip-#13 or -#14 on day 14, 7 and 0. HHD
mice immunized with irrelevant hepatitis C virus core 132–140
peptide coupled to the surface of liposomes were used as the
negative control. On day 0, we challenged HHD mice with
2×104 RMA-HHD lymphoma cells and monitored tumor growth
and survival. The results showed that on day 24 following the
challenge there was no significant difference in tumor size between
Lip-#13 and the control (Fig. 5A).
This result suggested that immunization with Lip-#13 did not induce
sufficient protective antitumor immunity. By contrast, tumor size
in Lip-#14 immunized mice was significantly lower than the control
and Lip-#13 immunized mice (P<0.05, Mann-Whitney U test,
Fig. 5A). Furthermore, Lip-#14
immunization prolonged the survival time of tumor-bearing mice
compared with the control mice (P<0.05, Log-rank test Fig. 5B). There was no significant
difference in survival time between Lip-#13 and the control
(Fig. 5B). Thus, these results
showed that, interference with tumor growth of TERT-expressing
RMA-HHD cells was provided by immunization with Lip-#14.
Discussion
In the present study, we utilized the high homology
between mTERT and hTERT (26),
which enabled us to examine epitopes exhibiting high immunogenicity
on the surface of liposomes and test their immunotherapeutic
potency by immunizing HHD mice and challenging with RMA-HHD
lymphoma cells. In addition, by including a heteroclitic variant to
each epitope candidate, we identified a novel epitope #13 derived
from TERT with low affinity to HLA-A2.1 molecules. The #14, the
heteroclitic peptide of #13, had a high affinity to HLA-A2.1 and
when coupled to the surface of liposome, induced strong and
long-lasting CD8+ T cell responses against both #14 and
the original epitope #13. Those responses were found to provide
protection against endogenously TERT-expressing tumor cells.
As detailed characterization of many tumor cell
expression molecules that act as TAAs is now available (27,28),
immunotherapy has become an increasingly essential component of
cancer therapies (29,30). Active cancer immunotherapy such as a
cancer vaccine is based on fundamental knowledge that TAAs are
presented on MHC molecules for recognition by specific T cells.
However, the heterogeneous expression of most of the characterized
TAAs limits the broad applicability of cancer vaccines. TAAs that
can be broadly applied to various types of cancer, such as
universal TAA, have been limited. Criteria for universal TAA
include: i) expression by the vast majority of cancers, ii)
presence of peptides that bind to MHC molecules, iii) sufficient
natural antigen processing by tumor cells that leads to the
expression of antigen-derived peptides in the groove of MHC, and
iv) recognition by T cell repertoire in an MHC-restricted manner.
Previous studies have shown TERT meets these criteria (31). From these attractive fortes, TERT
has been used as target of cancer vaccine (32). However, similar to most TAA, TERT is
a self-antigen that is expressed on normal tissues, including the
thymus (33). This fact raises the
issue of tolerance of the TERT-specific T-cell repertoire and
consequently the inability to trigger a strong and efficient
antitumor immune response (14).
Previous reports have shown that, self antigen-specific tolerance
influences mainly the T-cell repertoire against dominant
self-epitopes and to a much lesser extent against the cryptic
epitope (34–36). Thus, we examined the non-tolerized
repertoire against low affinity epitopes of TERT as the target of
the antitumor vaccination. Since low affinity epitopes may be
non-immunogenic, we devised two strategies for enhancement of
immunogenicity of the epitope. One was modification of the epitope
sequence to enhance affinity to MHC molecules. The other was
chemically coupling the epitope peptide to the surface of
liposome.
It has been shown that immunization using
heteroclitical peptides derived from low affinity TERT epitopes
emulsified in IFA recruits non-tolerized CTL repertoire and are
more efficient at inducing tumor immunity than high affinity
epitopes (15). However, the #14
peptide, the heteroclitical epitope peptide identified in the
present study, induced only weak CD8 T-cell immune responses
against the native epitope when emulsified in IFA and injected in
HHD mice. By contrast, when coupled to the surface of liposomes, it
induced strong CD8 T-cell immune responses against the native
cryptic epitope #13, while #13 peptide on the surface of liposomes
induced poor CD8 T-cell responses. Therefore, we consider that the
synergistic effect between heteroclitic modification and coupling
to the surface of liposomes provided the strong immunogenicity of
Lip-#14 and induced strong CD8 T cell responses against native
cryptic epitope. Moreover, #1 peptide (540–548), which was reported
to induce CD8 immunity against TERT when emulsified in IFA
(11) had no immunogenicity when
coupled to the surface of the liposomes. These findings suggest
that coupling to the surface of liposomes affects immunogenicity of
peptides. In a previous study, we showed that coupling to the
surface of liposomes alters the immunogenicity of hepatitis C
virus-derived peptides (24). The
mechanisms underlying the alteration of immunogenicity of peptides
by liposome coupling is unclear. We hypothesize that the difference
of the immunogenicity is associated with the difference of antigen
presentation mechanisms. Peptides emulsified in IFA are considered
to fit into the pockets of MHC class I molecules on APCs replacing
the original endogenously synthesized peptides and presented to CD8
T cells (37). On the other hand,
we have shown that antigens chemically coupled to the surface of
liposomes consisting of unsaturated fatty acid (oleoyl liposome)
are cross-presented by APCs (4).
Results from another study have shown that oleoyl liposomes may be
taken up by APCs via at least two pathways: fusion with and
penetration of plasma membranes and pinocytosis (38). These unique antigen presentation
mechanisms may be responsible for the unique immunogenicity of the
peptide antigens.
The efficient antitumor immune responses are often
fraught with autoimmunity. Vitiligo-like depigmentation has been
reported after immunotherapy against malignant melanoma (39). By contrast, adoptive transfer of
p53-specific CTL provides p53-overexpressing tumor growth
inhibition without autoimmunity (40). In the present study, we did not
observe any adverse effect in liposomal antigen-injected animals
despite evidence of the induction of strong TERT-specific CD8
T-cell responses. Previous findings have shown that there are
differences in TERT expression in normal somatic and tumor tissues.
Thus, TERT is a potential safe target for cancer immunotherapy
(41). Gross et al showed
that immunization with heteroclitic low affinity TERT peptide
resulted in anti-TERT T-cell immunity without any adverse effect by
their careful examination in various somatic tissues (15). Recent findings show the safety of
immunotherapy against TERT are accumulating (15,41–43).
However, future investigations should be conducted to address
whether the telomerase inhibition by host immune system activated
by immunotherapy can affect normal tissues.
In conclusion, vaccination with the liposome-coupled
heteroclitical peptide derived from TERT elicits antitumor CD8
T-cell responses evading the influence of tolerance in HHD mice.
Therefore, heteroclitical-modified antigens of cryptic epitope with
low affinity for HLA molecules derived from tumor antigens coupled
to the surface of liposome may play a role as an effective cancer
vaccine candidate.
Acknowledgments
The present study was supported by Grant from the
Ochiai memorial award 2011, the Saitama Medical University Internal
Grant 2012 and the JSPS KAKENHI grant no. 25430156. The authors
would like to thank Hiroe Akatsuka for his technical assistance, as
well as Dr Masanori Matsui and Dr Nobuharu Kobayashi (Depertment of
Microbiology, Saitama Medical University, Saitama, Japan) for their
helpful discussions. We are also grateful to Dr F.A. Lemonnier
(Pasteur Institute, Paris, France) for providing HHD mice and
RMA-HHD cells.
References
|
1
|
Lee PP, Yee C, Savage PA, Fong L,
Brockstedt D, Weber JS, Johnson D, Swetter S, Thompson J, Greenberg
PD, et al: Characterization of circulating T cells specific for
tumor-associated antigens in melanoma patients. Nat Med. 5:677–685.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kawakami Y, Eliyahu S, Delgado CH, Robbins
PF, Sakaguchi K, Appella E, Yannelli JR, Adema GJ, Miki T and
Rosenberg SA: Identification of a human melanoma antigen recognized
by tumor-infiltrating lymphocytes associated with in vivo tumor
rejection. Proc Natl Acad Sci USA. 91:6458–6462. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Peek LJ, Middaugh CR and Berkland C:
Nanotechnology in vaccine delivery. Adv Drug Deliv Rev. 60:915–928.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Taneichi M, Ishida H, Kajino K, Ogasawara
K, Tanaka Y, Kasai M, Mori M, Nishida M, Yamamura H, Mizuguchi J,
et al: Antigen chemically coupled to the surface of liposomes are
cross-presented to CD8+ T cells and induce potent
antitumor immunity. J Immunol. 177:2324–2330. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Takagi A, Matsui M, Ohno S, Duan H, Moriya
O, Kobayashi N, Oda H, Mori M, Kobayashi A, Taneichi M, et al:
Highly efficient antiviral CD8+ T-cell induction by
peptides coupled to the surfaces of liposomes. Clin Vaccine
Immunol. 16:1383–1392. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Taneichi M, Tanaka Y, kakiuchi T and
uchida T: Liposome- coupled peptides induce long-lived memory CD8 T
cells without CD4 T cells. PLoS One. 5:e150912010. View Article : Google Scholar
|
|
7
|
Vonderheide RH, Hahn WC, Schultze JL and
Nadler LM: The telomerase catalytic subunit is a widely expressed
tumor-associated antigen recognized by cytotoxic T lymphocytes.
Immunity. 10:673–679. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Minev B, Hipp J, Firat H, Schmidt JD,
Langlade-Demoyen P and Zanetti M: Cytotoxic T cell immunity against
telomerase reverse transcriptase in humans. Proc Natl Acad Sci USA.
97:4796–4801. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Artandi SE and DePinho RA: Telomeres and
telomerase in cancer. Carcinogenesis. 31:9–18. 2010. View Article : Google Scholar :
|
|
10
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu JP, Chen W, Schwarer AP and Li H:
Telomerase in cancer immunotherapy. Biochim Biophys Acta.
1805:35–42. 2010. View Article : Google Scholar
|
|
12
|
Arai J, Yasukawa M, Ohminami H, Kakimoto
M, Hasegawa A and Fujita S: Identification of human telomerase
reverse transcriptase-derived peptides that induce
HLA-A24-restricted antileukemia cytotoxic T lymphocytes. Blood.
97:2903–2907. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Warnock MG and Goodacre JA: Cryptic T-cell
epitopes and their role in the pathogenesis of autoimmune diseases.
Br J Rheumatol. 36:1144–1150. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Scardino A, Gross DA, Alves P, Schultze
JL, Graff-Dubois S, Faure O, Tourdot S, Chouaib S, Nadler LM,
Lemonnier FA, et al: HER-2/neu and hTERT cryptic epitopes as novel
targets for broad spectrum tumor immunotherapy. J Immunol.
168:5900–5906. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gross DA, Graff-Dubois S, Opolon P, Cornet
S, Alves P, Bennaceur-Griscelli A, Faure O, Guillaume P, Firat H,
Chouaib S, et al: High vaccination efficiency of low-affinity
epitopes in antitumor immunotherapy. J Clin Invest. 113:425–433.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tourdot S, Scardino A, Saloustrou E, Gross
DA, Pascolo S, Cordopatis P, Lemonnier FA and Kosmatopoulos K: A
general strategy to enhance immunogenicity of low-affinity HLA-A2.
1-associated peptides: Implication in the identification of cryptic
tumor epitopes. Eur J Immunol. 30:3411–3421. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Thorn M, Wang M, Kløverpris H, Schmidt
EGW, Fomsgaard A, Wenandy L, Berntsen A, Brunak S, Buus S and
Claesson MH: Identification of a new hTERT-derived
HLA-A*0201 restricted, naturally processed CTL epitope.
Cancer Immunol Immunother. 56:1755–1763. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cole Dk, Edwards ES, Wynn KK, Clement M,
Miles JJ, Ladell K, Ekeruche J, Gostick E, Adams KJ, Skowera A, et
al: Modification of MHC anchor residues generates heteroclitic
peptides that alter TCR binding and T cell recognition. J Immunol.
185:2600–2610. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pinolla-Ibarz J, May RJ, Koronitsvit T,
Gomez M, Kappel B, Zakhaleva V, Zhang RH and Scheinberg DA:
Improved human T-cell responses against synthetic HLA-0201 analog
peptides derived from the WT1 oncoprotein. Leukemia. 20:2025–2033.
2006. View Article : Google Scholar
|
|
20
|
Parkhurst MR, Salgaller ML, Southwood S,
Robbins PF, Sette A, Rosenberg SA and Kawakami Y: Improved
induction of melanoma-reactive CTL with peptides from the melanoma
antigen gp100 modified at HLA-A*0201-binding residues. J
Immunol. 157:2539–2548. 1996.PubMed/NCBI
|
|
21
|
Kubo RT, Sette A, Grey HM, Appella E,
Sakaguchi K, Zhu NZ, Arnott D, Sherman N, Shabanowitz J, Michel H,
et al: Definition of specific peptide motifs for four major HLA-A
alleles. J Immunol. 152:3913–3924. 1994.PubMed/NCBI
|
|
22
|
Pascolo S, Bervas N, Ure JM, Smith AG,
Lemonnier FA and Pérarnau B: HLA-A2.1-restricted education and
cytolytic activity of CD8(+) T lymphocytes from beta2
microglobulin (beta2m) HLA-A2.1 monochain transgenic H-2Db beta2m
double knockout mice. J Exp Med. 185:2043–2051. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Adotévi O, Mollier K, Neuveut C, Dosset M,
Ravel P, Fridman WH, Tartour E, Charneau P, Wain-Hobson S and
Langlade-Demoyen P: Targeting human telomerase reverse
transcriptase with recombinant lentivector is highly effective to
stimulate antitumor CD8 T-cell immunity in vivo. Blood.
115:3025–3032. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Takagi A, Kobayashi N, Taneichi M, Uchida
T and Akatsuka T: Coupling to the surface of liposomes alters the
immunogenicity of hepatitis C virus-derived peptides and confers
sterile immunity. Biochem Biophys Res Commun. 430:183–189. 2013.
View Article : Google Scholar
|
|
25
|
Kaech SM, Wherry EJ and Ahmed R: Effector
and memory T-cell differentiation: Implications for vaccine
development. Nat Rev Immunol. 2:251–262. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Greenberg RA, Allsopp RC, Chin L, Morin GB
and DePinho RA: Expression of mouse telomerase reverse
transcriptase during development, differentiation and
proliferation. Oncogene. 16:1723–1730. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Forni G, Lollini PL, Musiani P and Colombo
MP: Immunoprevention of cancer: Is the time ripe? Cancer Res.
60:2571–2575. 2000.PubMed/NCBI
|
|
28
|
Spurrell EL and Lockley M: Adaptive
immunity in cancer immunology and therapeutics. E Cancer Med Sci.
8:4412014.
|
|
29
|
Blattman JN and Greenberg PD: Cancer
immunotherapy: A treatment for the masses. Science. 305:200–205.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mellman I, Coukos G and Dranoff G: Cancer
immunotherapy comes of age. Nature. 480:480–489. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Patel KP and Vonderheide RH: Telomerase as
a tumor-associated antigen for cancer immunotherapy.
Cytotechnology. 45:91–99. 2004. View Article : Google Scholar
|
|
32
|
Beatty GL and Vonderheide RH: Telomerase
as a universal tumor antigen for cancer vaccines. Expert Rev
Vaccines. 7:881–887. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu K, Schoonmaker MM, Levine BL, June CH,
Hodes RJ and Weng NP: Constitutive and regulated expression of
telomerase reverse transcriptase (hTERT) in human lymphocytes. Proc
Natl Acad Sci USA. 96:5147–5152. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cibotti R, Kanellopoulos JM, Cabaniols JP,
Halle-Panenko O, Kosmatopoulos K, Sercarz E and Kourilsky P:
Tolerance to a self-protein involves its immunodominant but does
not involve its subdominant determinants. Proc Natl Acad Sci USA.
89:416–420. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nanda NK and Sercarz EE: Induction of
anti-self-immunity to cure cancer. Cell. 82:13–17. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Uram JN, Black CM, Flynn E, Huang L,
Armstrong TD and Jaffee EM: Nondominant CD8 T cells are active
players in the vaccine-induced antitumor immune response. J
Immunol. 186:3847–3857. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Townsend ARM, Rothbard J, Gotch FM,
Bahadur G, Wraith D and McMichael AJ: The epitopes of influenza
nucleoprotein recognized by cytotoxic T lymphocytes can be defined
with short synthetic peptides. Cell. 44:959–968. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tanaka Y, Taneichi M, Kasai M, Kakiuchi T
and Uchida T: Liposome-coupled antigens are internalized by
antigen-presenting cells via pinocytosis and cross-presented to CD8
T cells. PLoS One. 5:e152252010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Teulings HE, Limpens J, Jansen SN,
Zwinderman AH, Reitsma JB, Spuls PI and Luiten RM: Vitiligo-like
depigmentation in patients with stage III–IV melanoma receiving
immunotherapy and its association with survival: A systematic
review and meta-analysis. J Clin Oncol. 33:773–781. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Vierboom MP, Nijman HW, Offringa R, van
der Voort EI, van Hall T, van den Broek L, Fleuren GJ, Kenemans P,
Kast WM and Melief CJ: Tumor eradication by wild-type p53-specific
cytotoxic T lymphocytes. J Exp Med. 186:695–704. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Harley CB: Telomerase and cancer
therapeutics. Nat Rev Cancer. 8:167–179. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yan J, Pankhong P, Shin TH, Obeng-Adjei N,
Morrow MP, Walters JN, Khan AS, Sardesai NY and Weiner DB: Highly
optimized DNA vaccine targeting human telomerase reverse
transcriptase stimulates potent antitumor immunity. Cancer Immunol
Res. 1:179–189. 2013. View Article : Google Scholar
|
|
43
|
Kotsakis A, Papadimitraki E, Vetsika EK,
Aggouraki D, Dermitzaki EK, Hatzidaki D, Kentepozidis N, Mavroudis
D and Georgoulias V: A phase II trial evaluating the clinical and
immunologic response of HLA-A2(+) non-small cell lung
cancer patients vaccinated with an hTERT cryptic peptide. Lung
Cancer. 86:59–66. 2014. View Article : Google Scholar : PubMed/NCBI
|