Introduction
Thyroid cancer is the most common endocrine
malignancy, and the number of new cases diagnosed worldwide has
increased in the last decade (1).
Thyroid cancer is classified into undifferentiated and
differentiated cancer, and the vast majority of thyroid cancers
(~80%) are differentiated thyroid cancers (DTCs) (2). DTCs consist of two cellular types:
papillary thyroid cancers (PTCs) and differentiated follicular
thyroid cancers (FTCs). Various effective treatments have been used
for thyroid cancer therapy, such as complete thyroidectomy followed
by radioiodine therapy (3). These
patients have a low mortality rate and excellent prognosis.
However, a subset of patients that develops distant metastases
presents with a high disease recurrence rate and poor prognosis and
the disease may develop into anaplastic thyroid cancer with a fatal
outcome (4,5). Therefore, research on the molecular
mechanisms involved in thyroid carcinogenesis will facilitate the
development of more effective therapy for this cancer.
Ample evidence indicates that aging is the main risk
factor for cancer, and most cancers are diagnosed in individuals
over 55 years of age (6).
Currently, Klotho has recently been identified as a new anti-aging
gene and has gained much attention (7). Klotho (KL), a 1012-amino acid type I
single-pass transmembrane protein, is widely expressed in various
types of tissues, such as brain, kidney, various exocrine and
endocrine tissues, including the thyroid gland (8–10). The
Klotho gene is composed of 5 exons (8,11) and
contains an intracellular domain and an extracellular domain. The
extracellular domain of Klotho is composed of two domains, KL1 and
KL2, with weak homology (10),
while the intracellular domain is short (~10-amino acid long) which
has no known functional domain (6).
The Klotho gene plays roles in various diseases in mice, such as
arteriosclerosis, osteoporosis, endothelial dysfunction,
Parkinsonian gait and cognitive impairment (12–14).
In humans, Klotho is involved in reduced human longevity and
coronary artery disease (15).
Recently, Klotho was identified as a potential tumor suppressor,
and ample evidence suggests that Klotho expression influences human
breast, hepatocarcinogenesis, cervical and lung cancers (7,16–18).
However, the functional role and specific molecular mechanism of
Klotho in thyroid cancer remain unclear.
Ample laboratory data indicate that the Klotho gene
could regulate the expression of STC1 and STC2 in the kidney
(19). Stanniocalcin (STC) is a
secreted glycoprotein hormone and has two isotypes, STC1 and STC2.
Human STC1 is encoded by a single copy gene which is localized at
chromosome 8p11.2-p21 (20). STC1
was originally discovered in mammals and in the endocrine glands of
the kidneys, and stannius corpuscles of bony fishes (21). STC1 is expressed in a wide variety
of tissues, such as the heart, prostate, adrenal gland, lung,
kidney, liver and thyroid gland (22–24).
Multiple lines of evidence show that mammalian STC1 was cloned as a
cancer-related gene (25). Growing
evidence suggests that STC1 expression may play an important role
in human cancers, and high STC1 gene expression has been found in
colorectal, breast, hepatocellular, carcinomas and thyroid cancers
(26–29). In the present study, we investigated
whether Klotho expression is associated with the development of
thyroid cancer via regulation of STC1 protein expression. We found
that Klotho overexpression significantly inhibited human thyroid
cancer cell growth and promoted apoptosis, and suppressed the
expression of STC1.
Materials and methods
Cell culture and transfection
Human thyroid cancer cell lines established from
primary thyroid cancer (FTC133) and distant metastases (FTC238)
(both from ECACC, Wiltshire, UK) were used in this study. Cells
were cultured in RPMI-1640 medium (Invitrogen, Carlsbad, CA, USA)
supplemented with 5% FCS, 1% glutamine and 1%
penicillin-streptomycin. Klotho-overexpressing cells were
constructed with the human Klotho expression plasmid
(pCDNA3.1-Klotho) using Lipofectamine 2000 (Invitrogen) following
the manufacturer's instructions (30).
Cell viability assay
Human thyroid cancer cells in exponential growth
(5×104 cells/well) were seeded into a 96-well sterile
culture plate. Cells were transfected with the Klotho vector or the
control vector for 72 h. Thyroid cancer cell viability was
determined using the CellTiter-Glo Luminescent Cell Viability Assay
kit following the manufacturer's instructions (Promega, Madison,
WI, USA).
MTT assay
Human thyroid cancer cell proliferation was measured
by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
(MTT) assay according to a previously described method (6). The thyroid cancer cells
(5×104 cells/well) were plated in a 96-well culture
plate for 24 h. The cells were then transfected with Klotho and
cultured for 72 h, and 20 μl MTT (10 mg/ml) was then added.
The reaction was terminated after an additional 4 h of incubation.
DMSO (200 μl) was added to the cells for 10 min. The optical
density (OD) value of each well was measured at 570 nm.
Nucleosome ELISA assay for detection of
apoptosis
Human thyroid cancer cell apoptosis was carried out
using the Nucleosome ELISA kit as previously described (31). FTC133 and FTC238 cells were
harvested. Nucleosome ELISAs were performed according to the
manufacturer's instructions (Oncogene Research Products, Cambridge,
MA, USA).
Caspase-3 activity assay
Caspase-3 activity was detected using the
Caspase-Glo®3/7 assay (Promega) according to the
manufacturer's instructions.
Quantitative RT-PCR
Quantitative RT-PCR was performed as previously
described (22). Total RNA was
isolated from human thyroid cancer cells and reverse transcribed
into cDNA using SuperScript™ III Reverse Transcriptase
(Invitrogen). PCR amplification was carried out by an ABI PRISM
7900 thermocycler using SYBR Premix Taq (Applied Biosystems). The
following primer pairs were used for PCR amplification: STC1_F,
human STC1-specific primers were forward,
5′-ATCACATTCCAGCAGGCTTC-3′ and reverse, 5′-CCTGAAGCCATCACTGAGGT-3′;
human Klotho-specific primers were forward,
5′-ACTCCCCCAGTCAGGTGGCGGTA-3′ and reverse,
5′-TGGGCCCGGGAAACCATTGCTGTC-3′. The reaction conditions was as
follows: 95°C for 5 min; followed by 40 cycles at 95°C for 15 sec;
60°C for 45 sec. Relative levels of gene expression are expressed
relative to β-actin and calculated using the 2−ΔΔCT
method (32).
Western blot analysis
Proteins were collected from the cells transfected
with the Klotho vector or the control vector. Cells were washed in
PBS and lysed using RIPA lysis buffer. The concentration of protein
was measured using the BCA kit. Samples (40 μg) were
subjected to SDS-PAGE and transferred to Immobilon-P Transfer
Membranes (Millipore). Western blot anlaysis was conducted using
the following primary and secondary antibodies: rabbit anti-Klotho
polyclonal antibody, rabbit anti-STC1 polyclonal antibody, mouse
anti-β-actin monoclonal antibody, HRP-conjugated rabbit anti-mouse
IgG and HRP-conjugated goat anti-rabbit IgG. Bands were visualized
using an electrochemiluminescence kit (Immulite Pyrilinks-D;
Diagnostic Products Corporation, Los Angeles, CA, USA). Antibodies
used in this study were all obtained from Abcam (Cambridge, MA,
USA).
siRNA
Klotho and STC1 siRNA oligoribonucleotides and
control siRNAs were obtained from Shanghai Sangon Co., Ltd.
(Shanghai, China). Their sequences were as follows: Klotho siRNA
oligoribonucleotides directed against 1274-1298,
5′-ACCAAGAGAGAUGAUGCCAAAUAU-3′ and control siRNAs scrambled,
5′-CACGAGAUAGAGUAGAACCAACUAU-3′; STC1 siRNA oligonucleotides
against 369-392 (33),
5′-GGTGCAGGAAGAGTGCTACAGCAAGTACGTAGGTTGCTGTAGCACTCTTCCTGCACCCTTTTTG-3′.
The DNA oligonucleotides used to generate scrambled siRNA were
5′-GGCGCGCTTTGTAGGATTCGATACGTAAACGAATCCTACAAAGCGCGCTTTTTG-3′. The
cells were transfected with Klotho or STC1 siRNA or scrambled using
Lipofectamine 2000 (Invitrogen) following the manufacturer's
instructions (9).
Statistical analysis
Results from at least three independent experiments
are presented as the mean ± SD. Statistical comparisons were
performed using one-way ANOVA and Dunnett's test. All analyses were
performed using SPSS 13.0 software (SPSS, Chicago, IL, USA).
Results
Overexpression of Klotho inhibits thyroid
cancer cell survival and proliferation
Cell proliferation plays a crucial role in tumor
progression and can regulate the fate of tumors at any given time
(30). Thus, in order to determine
whether Klotho gene and protein expression is associated with
thyroid cancer cell survival and proliferation, we constructed
Klotho-overexpressing cells using pCDNA3.1-Klotho plasmids. As
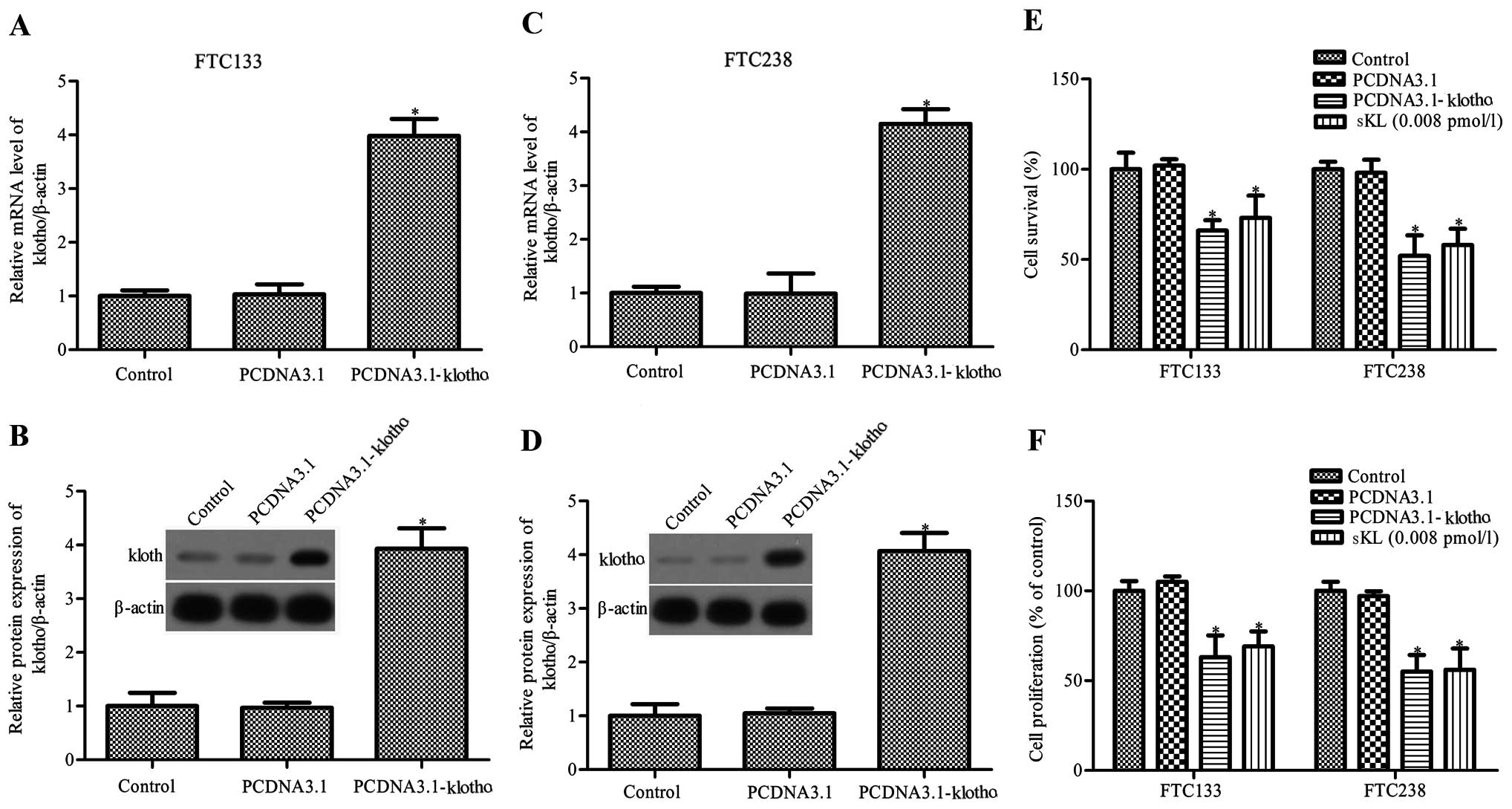
shown in Fig. 1A–D, the mRNA and
protein expression levels of Klotho in the FTC133 and FTC238 cells
were markedly increased after pCDNA3.1-Klotho transfection. FTC133
and FTC238 cells were transfected with pCDNA3.1-Klotho or control
pCDNA3.1, and then the cell viability was assessed by MTT assay.
Klotho expression significantly inhibited FTC133 and FTC238 cell
survival, and decreased FTC133 and FTC238 cell proliferation
(Fig. 1E and F).
Mounting evidence suggests that Klotho may be shed
and could act as a circulating hormone, and it has been
demonstrated that soluble Klotho and conditioned medium derived
from Klotho-overexpressing cells are both active (9,11,34,35).
Therefore, FTC133 and FTC238 cells were next treated with 0.008
pmol/l soluble human KL1 (sKL; PeproTech Inc.). sKL was found to
reduce the viability and proliferation of the FTC133 and FTC238
cells by 73 and 58%, 69 and 56%, respectively (P<0.05, Fig. 1E and F). The results showed that
Klotho and sKL both significantly inhibited the survival and
proliferation of the FTC133 and FTC238 cells.
Overexpression of Klotho increases the
apoptosis of thyroid cancer cells
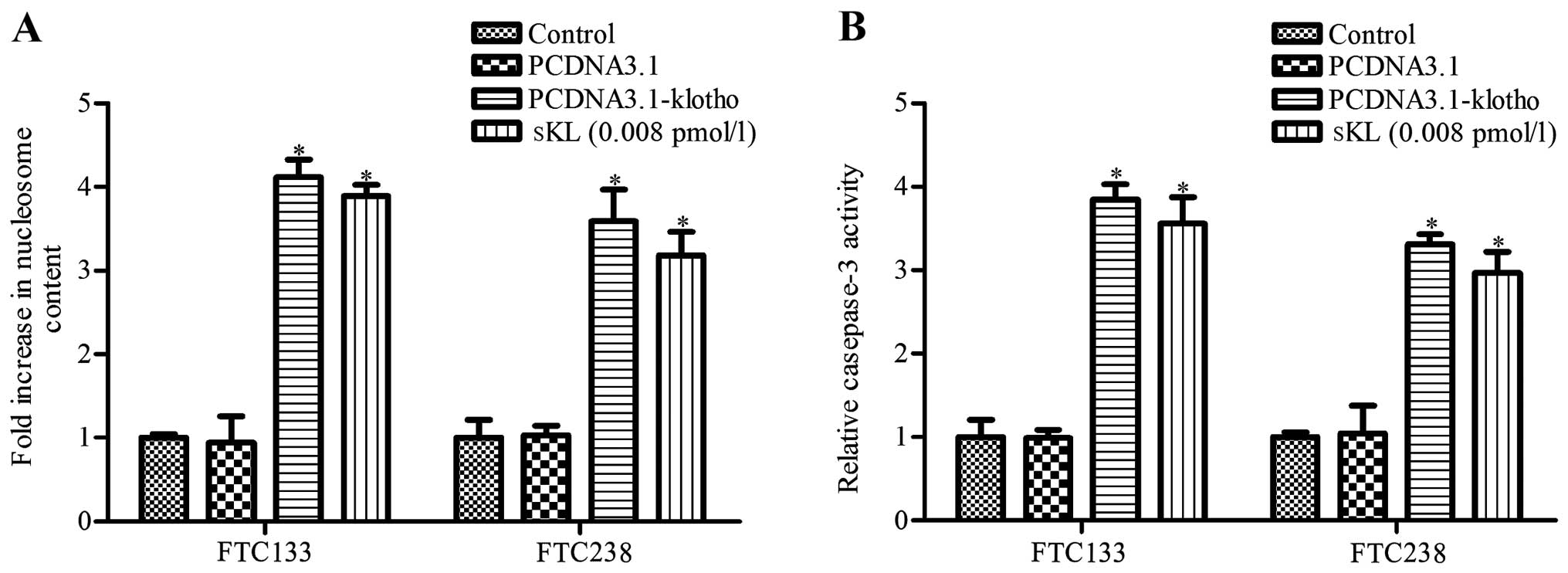
We next performed nucleosome ELISA assay to assess
the apoptosis of the FTC133 and FTC238 cells. The results revealed
that both the Klotho-transfected and sKL treatment group exhibited
more apoptotic cells when compared to that of the control group
(P<0.05, Fig. 2A); no
significant difference was noted between the pCDNA3.1 and control
group (P>0.05, Fig. 2A). We next
determined the activity of caspase-3 in the different groups using
the Caspase-Glo®3/7 assay. The activity of caspase-3 was
much higher in the Klotho-transfected and sKL treatment group when
compared with that of the control group (Fig. 2B). The results indicated that Klotho
and sKL both significantly promoted FTC133 and FTC238 cell
apoptosis.
Loss of Klotho increases human cancer
cell growth and inhibits cell apoptosis
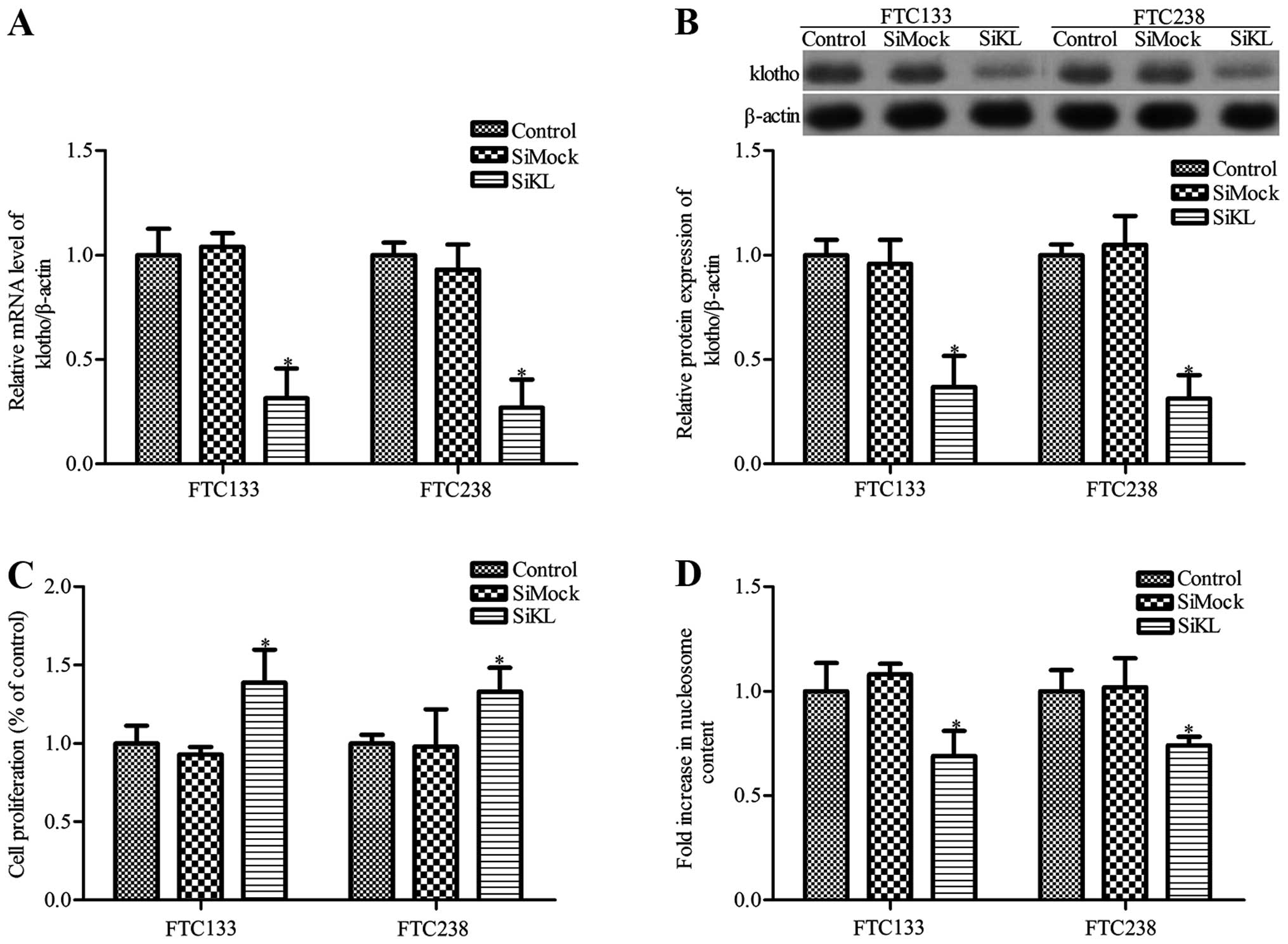
We next investigated the effects of Klotho knockdown
on the growth and apoptosis of thyroid cancer cells. To decrease
the expression of Klotho protein, thyroid cancer cells were
transfected with Klotho siRNA (siKL) or control siRNA (siMock).
RT-PCR and western blot analysis results showed that Klotho
expression in the thyroid cancer FTC133 and FTC238 cells was
markedly decreased following siKL transfection (P<0.05, Fig. 3A and B), whereas there was no
significant difference between the siMock group and control group
(P>0.05). Moreover, cancer cell proliferation was markedly
increased in the siKL-transfected cells compared with the cell
proliferation noted in the control and siMock groups (P<0.05,
Fig. 3C). Simultaneously, cell
apoptosis was also measured. The results showed that siKL
transfection significantly inhibited the apoptosis of the FTC133
and FTC238 cells when compared to the apoptosis noted in the
control group, whereas siMock transfection had no obvious effect on
cell apoptosis (Fig. 3D).
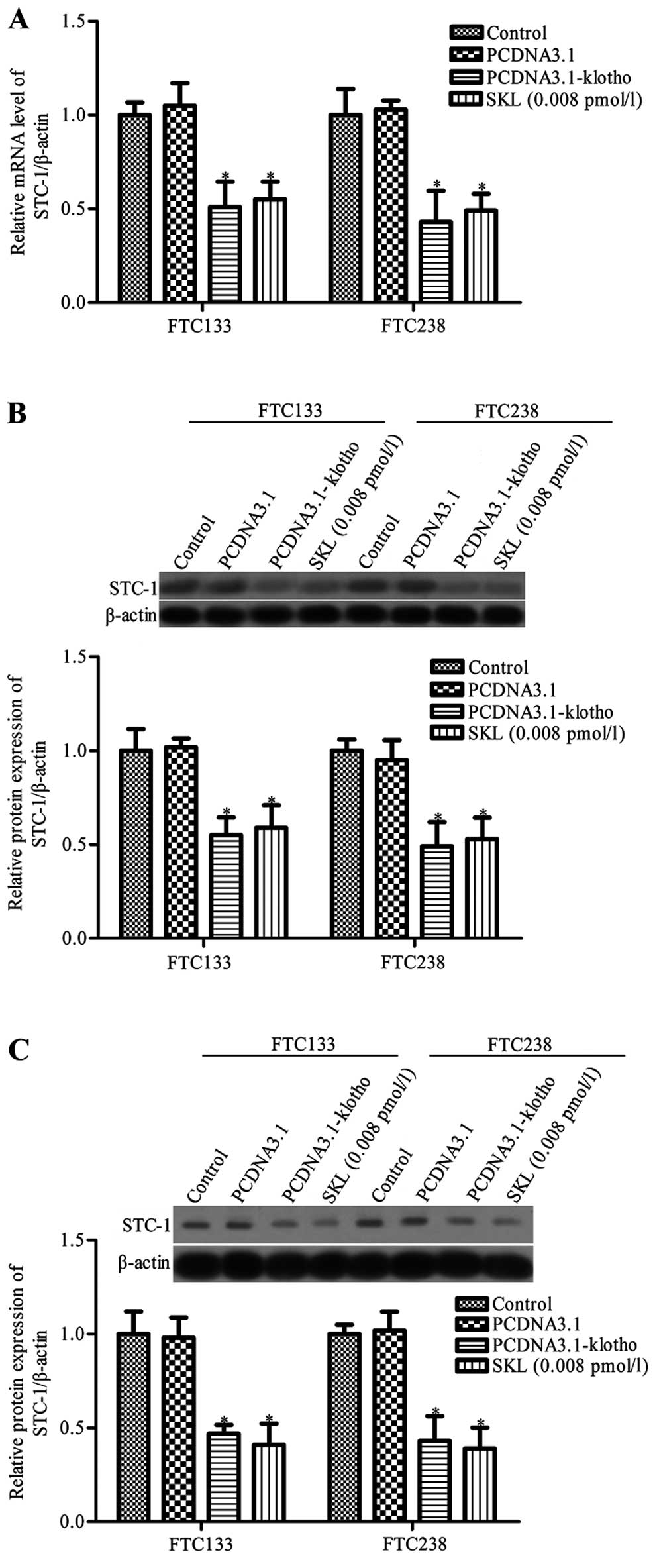
Klotho inhibits STC1 expression in
thyroid cancer cells
STC1 is a secreted glycoprotein hormone and a
molecular marker for micrometastases of various human cancers,
including thyroid cancer (27).
Yahata et al showed that the Klotho gene regulates the
expression of STC1 and STC2 in the kidney (19). To further detect the mechanism of
Klotho-induced thyroid cancer cell apoptosis and growth inhibition,
the effects of Klotho on the expression of STC1 and STC2 in thyroid
cancer cell lines FTC133 and FTC238 were analyzed. RT-PCR and
western blot analysis showed that STC1 mRNA and protein expression
levels were significantly decreased in the Klotho-overexpressing
and sKL-treated thyroid cancer cell lines compared with the control
group (P<0.05, Fig 4A and B).
There was no obvious difference between the control group and
control pcDNA3.1 vector group (P>0.05, Fig. 4A and B). Since STC1 is a secreted
glycoprotein hormone, we also determined the protein expression of
STC1 in cell culture medium. The results revealed that the
secretion of STC1 in the Klotho-overexpressing and sKL-treated
thyroid cancer cell lines was much lower than the secretion in the
control group (P<0.05, Fig.
4C).
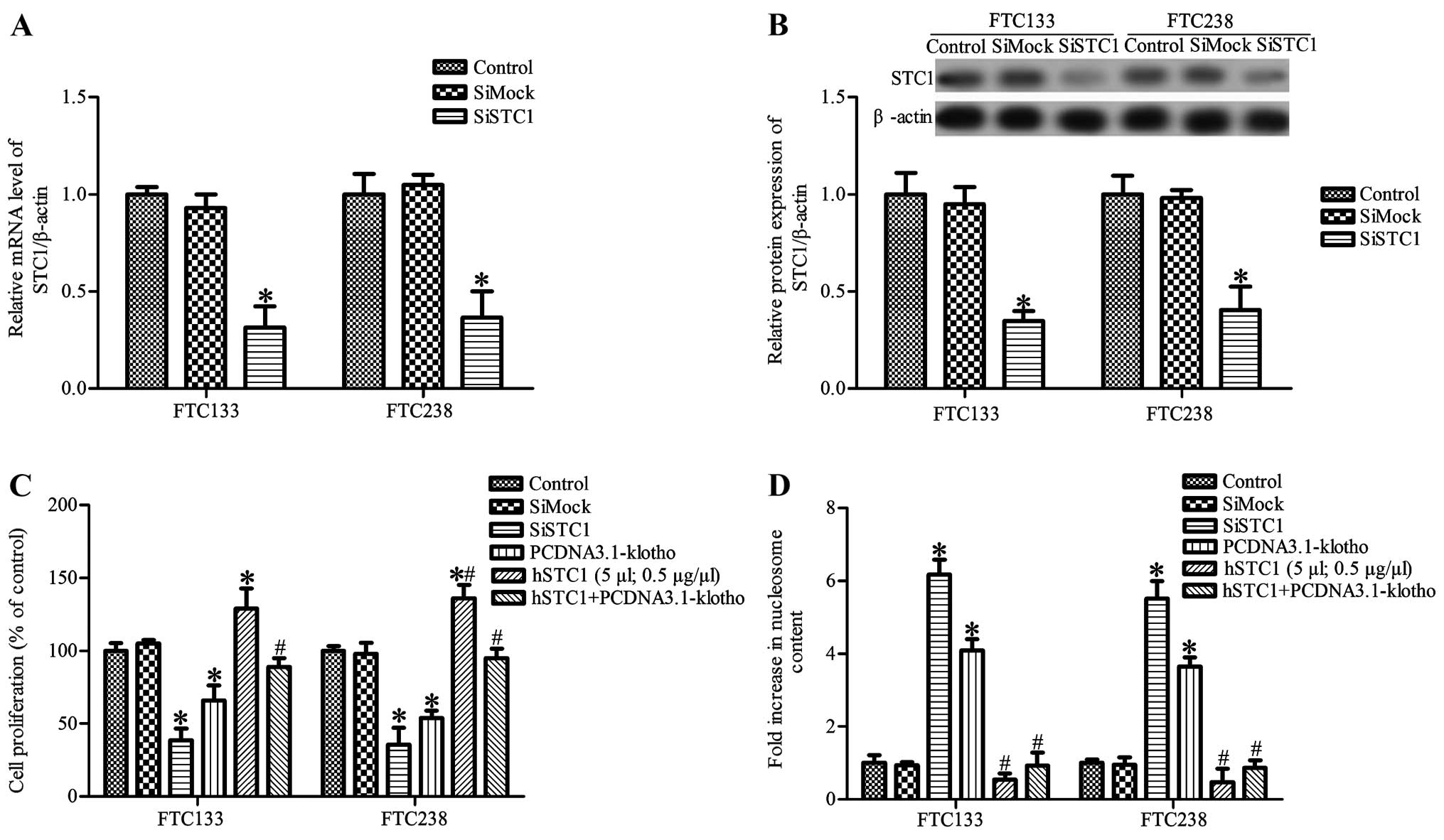
STC1 silencing inhibits cell growth and
promotes cell apoptosis
Thyroid cancer cells were transfected with STC1
siRNA (siSTC1) or control siRNA (siMock), and the level of STC1 in
the FTC133 and FTC238 cells was determined by RT-PCR and western
blot analysis, respectively. As shown in Fig. 5A and B, STC1 mRNA and protein
expression was significantly decreased in the siSTC1-treated
thyroid cancer cells compared with that noted in the control and
siMock groups (P<0.05). We also observed that STC1 silencing
markedly inhibited FTC133 and FTC238 cell proliferation and induced
cell apoptosis (Fig. 5C and D). To
further ascertain whether STC1 is involved in Klotho-induced cell
apoptosis, thyroid cancer cells were treated with 5 µl
recombinant human STC1 (hSTC1, 0.5 µg/µl; BioVender
Research and Diagnostic Products, Czech Republic). The results
showed that hSTC1 treatment significantly promoted FTC133 and
FTC238 cell proliferation and inhibited apoptosis. Moreover, hSTC1
treatment attenuated Klotho-induced inhibiiton of cell
proliferation and promotion of apoptosis (Fig. 5C and D). These results indicate that
Klotho inhibits cancer cell growth and induces apoptosis which is
partially dependent on the downregulation of STC1 expression.
Discussion
It has been reported that STC1 is highly expression
in several human tumors, including colorectal, breast,
hepatocellular carcinomas and thyroid cancers (26–29).
Klotho is known to regulate the expression of STC1. Mice deficient
in Klotho expression exhibit increased renal gene expression of
STC1 and STC2 compared with wild-type mice (19). Moreover, the level of Klotho was
found to be downregulated in breast, lung, colon, pancreatic,
gastric and cervical cancer (36–38).
Although evidence suggests that Klotho acts as a tumor suppressor
in numerous cancers (39), little
is known concerning its potential effect on thyroid cancer.
We reported here that overexpression of Klotho in
the thyroid cancer cell lines FTC133 and FTC238 resulted in a
decreased capacity for cell proliferation and an increased capacity
for cell apoptosis. To further demonstrate the effects of Klotho
overexpression in FTC133 and FTC238 cells, caspase-3 activity was
determined using the Caspase-glo 3/7 assay. Klotho may be shed and
acts as a circulating hormone, and it has been demonstrated that
soluble Klotho and conditioned medium derived from
Klotho-overexpressing cells are both active (9,11,34,35).
We also found that treatment with 0.008 pmol/l sKL significantly
reduced cell growth and promoted cell apoptosis. Loss of Klotho
resulted in the promotion of thyroid cancer cell proliferation and
inhibition of apoptosis. These results indicate that Klotho plays
an important role in thyroid cancer.
Yahata et al showed that the Klotho gene
regulates the expression of STC1 and STC2 in the kidney (19). Moreover, STC1 is upregulated in many
cancers, including thyroid cancer. STC1 is an endocrine hormone
firstly discovered from bony fishes, and is also expressed in
mammals (40). There is mounting
evidence that the mammalian STC1 gene is widely expressed in many
tissues at different levels (mRNA or protein), and may play a
different function. Moreover, evidence suggests that altered
patterns of STC1 expression are tightly involved in the development
of cancer (26). The present study
showed that Klotho overexpression markedly inhibited the expression
of STC1 in thyroid cancer FTC133 and FTC238 cells, which
corroborated the findings reported in the literature. In addition,
FTC133 and FTC238 cells transfected with siKL showed a marked
increase in the expression of STC1. We next established
STC1-silenced cells by STC1 siRNA. The results showed that STC1
knockdown induced the inhibition of proliferation and the promotion
of apoptosis in the FTC133 and FTC238 cells. These results suggest
that the effects of Klotho on FTC133 and FTC238 cell growth are
dependent on the level of STC1. STC1 is a secreted glycoprotein
hormone, and we observed that hSTC1 significantly inhibited Klotho
overexpression-induced cell apoptosis. These data suggest that
Klotho inhibits thyroid cancer cell proliferation and induces cell
apoptosis dependent on STC1.
The present study also had several limitations. The
detailed molecular mechanism of Klotho-mediated cell proliferation
and apoptosis remains unclear. Furthermore, Klotho-induced cell
apoptosis was only tested in thyroid cancer cell lines FTC133 and
FTC238; it will require further validation in other types of
thyroid cancer cell lines and in animal models. In summary, our
study provides initial evidence that Klotho inhibits human
follicular thyroid cancer cell growth and promotes apoptosis
through regulation of the expression of STC1, and may be a new
target for thyroid cancer treatment.
Acknowledgments
This study was supported by grants from the National
Natural Science Foundation of China (81302003), the National
Science and Technology Major Project (No. 2013ZX09303001) and the
Science & Technology Development Fund of Tianjin Education
Commission for Higher education (20120109).
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Xing M: Molecular pathogenesis and
mechanisms of thyroid cancer. Nat Rev Cancer. 13:184–199. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mazzaferri EL and Jhiang SM: Long-term
impact of initial surgical and medical therapy on papillary and
follicular thyroid cancer. Am J Med. 97:418–428. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cornett WR, Sharma AK, Day TA, Richardson
MS, Hoda RS, van Heerden JA and Fernandes JK: Anaplastic thyroid
carcinoma: An Overview. Curr Oncol Rep. 9:152–158. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ain KB, Egorin MJ and DeSimone PA;
Collaborative Anaplastic Thyroid Cancer Health Intervention Trials
(CATCHIT) Group: Treatment of anaplastic thyroid carcinoma with
paclitaxel: Phase 2 trial using ninety-six-hour infusion. Thyroid.
10:587–594. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen B, Wang X, Zhao W and Wu J: Klotho
inhibits growth and promotes apoptosis in human lung cancer cell
line A549. J Exp Clin Cancer Res. 29:99–105. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen L, Liu H, Liu J, Zhu Y, Xu L, He H,
Zhang H, Wang S, Wu Q, Liu W, et al: Klotho endows hepatoma cells
with resistance to anoikis via VEGFR2/PAK1 activation in
hepatocellular carcinoma. PLoS One. 8:e584132013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shiraki-Iida T, Aizawa H, Matsumura Y,
Sekine S, Iida A, Anazawa H, Nagai R, Kuro-o M and Nabeshima Y:
Structure of the mouse klotho gene and its two transcripts encoding
membrane and secreted protein. FEBS Lett. 424:6–10. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wolf I, Levanon-Cohen S, Bose S, Ligumsky
H, Sredni B, Kanety H, Kuro-o M, Karlan B, Kaufman B, Koeffler HP,
et al: Klotho: A tumor suppressor and a modulator of the IGF-1 and
FGF pathways in human breast cancer. Oncogene. 27:7094–7105. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kuro-o M, Matsumura Y, Aizawa H, Kawaguchi
H, Suga T, Utsugi T, Ohyama Y, Kurabayashi M, Kaname T, Kume E, et
al: Mutation of the mouse klotho gene leads to a syndrome
resembling ageing. Nature. 390:45–51. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Matsumura Y, Aizawa H, Shiraki-Iida T,
Nagai R, Kuro-o M and Nabeshima Y: Identification of the human
klotho gene and its two transcripts encoding membrane and secreted
klotho protein. Biochem Biophys Res Commun. 242:626–630. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nagai T, Yamada K, Kim HC, Kim YS, Noda Y,
Imura A, Nabeshima Y and Nabeshima T: Cognition impairment in the
genetic model of aging klotho gene mutant mice: A role of oxidative
stress. FASEB J. 17:50–52. 2003.
|
|
13
|
Nagai R, Saito Y, Ohyama Y, Aizawa H, Suga
T, Nakamura T, Kurabayashi M and Kuroo M: Endothelial dysfunction
in the klotho mouse and downregulation of klotho gene expression in
various animal models of vascular and metabolic diseases. Cell Mol
Life Sci. 57:738–746. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Saito Y, Yamagishi T, Nakamura T, Ohyama
Y, Aizawa H, Suga T, Matsumura Y, Masuda H, Kurabayashi M, Kuro-o
M, et al: Klotho protein protects against endothelial dysfunction.
Biochem Biophys Res Commun. 248:324–329. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Arking DE, Krebsova A, Macek M Sr, Macek M
Jr, Arking A, Mian IS, Fried L, Hamosh A, Dey S, McIntosh I, et al:
Association of human aging with a functional variant of klotho.
Proc Natl Acad Sci USA. 99:856–861. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tao Y, Pinzi V, Bourhis J and Deutsch E:
Mechanisms of disease: Signaling of the insulin-like growth factor
1 receptor pathway - therapeutic perspectives in cancer. Nat Clin
Pract Oncol. 4:591–602. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mattarocci S, Abbruzzese C, Mileo AM,
Visca P, Antoniani B, Alessandrini G, Facciolo F, Felsani A,
Radulescu RT and Paggi MG: Intracellular presence of insulin and
its phosphor-ylated receptor in non-small cell lung cancer. J Cell
Physiol. 221:766–770. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chang B, Kim J, Jeong D, Jeong Y, Jeon S,
Jung SI, Yang Y, Kim KI, Lim JS, Kim C, et al: Klotho inhibits the
capacity of cell migration and invasion in cervical cancer. Oncol
Rep. 28:1022–1028. 2012.PubMed/NCBI
|
|
19
|
Yahata K, Mori K, Mukoyama M, Sugawara A,
Suganami T, Makino H, Nagae T, Fujinaga Y, Nabeshima Y and Nakao K:
Regulation of stanniocalcin 1 and 2 expression in the kidney by
klotho gene. Biochem Biophys Res Commun. 310:128–134. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chang AC, Jeffrey KJ, Tokutake Y,
Shimamoto A, Neumann AA, Dunham MA, Cha J, Sugawara M, Furuichi Y
and Reddel RR: Human stanniocalcin (STC): Genomic structure,
chromosomal localization, and the presence of CAg trinucleotide
repeats. Genomics. 47:393–398. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wagner GF, Hampong M, Park CM and Copp DH:
Purification, characterization, and bioassay of teleocalcin, a
glycoprotein from salmon corpuscles of Stannius. Gen Comp
Endocrinol. 63:481–491. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hayase S, Sasaki Y, Matsubara T, Seo D,
Miyakoshi M, Murata T, Ozaki T, Kakudo K, Kumamoto K, Ylaya K, et
al: Expression of stanniocalcin 1 in thyroid side population cells
and thyroid cancer cells. Thyroid. 25:425–436. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yeung BH, Law AY and Wong CK: Evolution
and roles of stanniocalcin. Mol Cell Endocrinol. 349:272–280. 2012.
View Article : Google Scholar
|
|
24
|
Varghese R, Wong CK, Deol H, Wagner GF and
DiMattia GE: Comparative analysis of mammalian stanniocalcin genes.
Endocrinology. 139:4714–4725. 1998.PubMed/NCBI
|
|
25
|
Chang AC, Janosi J, Hulsbeek M, de Jong D,
Jeffrey KJ, Noble JR and Reddel RR: A novel human cDNA highly
homologous to the fish hormone stanniocalcin. Mol Cell Endocrinol.
112:241–247. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chang AC, Jellinek DA and Reddel RR:
Mammalian stanniocalcins and cancer. Endocr Relat Cancer.
10:359–373. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fujiwara Y, Sugita Y, Nakamori S, Miyamoto
A, Shiozaki K, Nagano H, Sakon M and Monden M: Assessment of
stannio-calcin-1 mRNA as a molecular marker for micrometastases of
various human cancers. Int J Oncol. 16:799–804. 2000.PubMed/NCBI
|
|
28
|
McCudden CR, Majewski A, Chakrabarti S and
Wagner GF: Co-localization of stanniocalcin-1 ligand and receptor
in human breast carcinomas. Mol Cell Endocrinol. 213:167–172. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Okabe H, Satoh S, Kato T, Kitahara O,
Yanagawa R, Yamaoka Y, Tsunoda T, Furukawa Y and Nakamura Y:
Genome-wide analysis of gene expression in human hepatocellular
carcinomas using cDNA microarray: Identification of genes involved
in viral carcinogenesis and tumor progression. Cancer Res.
61:2129–2137. 2001.PubMed/NCBI
|
|
30
|
Chen B, Ma X, Liu S, Zhao W and Wu J:
Inhibition of lung cancer cells growth, motility and induction of
apoptosis by Klotho, a novel secreted Wnt antagonist, in a
dose-dependent manner. Cancer Biol Ther. 13:1221–1228. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shih A, Davis FB, Lin HY and Davis PJ:
Resveratrol induces apoptosis in thyroid cancer cell lines via a
MAPK- and p53-dependent mechanism. J Clin endocrinol Metab.
87:1223–1232. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar
|
|
33
|
Liu G, Yang G, Chang B, Mercado-Uribe I,
Huang M, Zheng J, Bast RC, Lin SH and Liu J: Stanniocalcin 1 and
ovarian tumorigenesis. J Natl Cancer Inst. 102:812–827. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Abramovitz L, Rubinek T, Ligumsky H, Bose
S, Barshack I, Avivi C, Kaufman B and Wolf I: KL1 internal repeat
mediates klotho tumor suppressor activities and inhibits bFGF and
IGF-I signaling in pancreatic cancer. Clin Cancer Res.
17:4254–4266. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen CD, Podvin S, Gillespie E, Leeman SE
and Abraham CR: Insulin stimulates the cleavage and release of the
extracellular domain of Klotho by ADAM10 and ADAM17. Proc Natl Acad
Sci USA. 104:19796–19801. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lee J, Jeong DJ, Kim J, Lee S, Park JH,
Chang B, Jung SI, Yi L, Han Y, Yang Y, et al: The anti-aging gene
KLOTHO is a novel target for epigenetic silencing in human cervical
carcinoma. Mol Cancer. 9:1092010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang L, Wang X, Wang X, Jie P, Lu H, Zhang
S, Lin X, Lam EK, Cui Y, Yu J, et al: Klotho is silenced through
promoter hypermethylation in gastric cancer. Am J Cancer Res.
1:111–119. 2011.PubMed/NCBI
|
|
38
|
Pan J, Zhong J, Gan LH, Chen SJ, Jin HC,
Wang X and Wang LJ: Klotho, an anti-senescence related gene, is
frequently inactivated through promoter hypermethylation in
colorectal cancer. Tumour Biol. 32:729–735. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu H, Fergusson MM, Castilho RM, Liu J,
Cao L, Chen J, Malide D, Rovira II, Schimel D, Kuo CJ, et al:
Augmented Wnt signaling in a mammalian model of accelerated aging.
Science. 317:803–806. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yeung HY, Lai KP, Chan HY, Mak NK, Wagner
GF and Wong CK: Hypoxia-inducible factor-1-mediated activation of
stanniocalcin-1 in human cancer cells. Endocrinology.
146:4951–4960. 2005. View Article : Google Scholar : PubMed/NCBI
|