Introduction
The cancer stem cell (CSC) theory states that only a
small population of cells within a tumor is tumorigenic and has the
ability for self-renewal (1). CSCs
are thought to be related to tumor recurrence and treatment
resistance; even though we treat non-CSCs by surgery or
chemotherapy, relapse or metastasis may result if CSCs should
resist (2). This theory was first
suggested in leukemia, and many studies investigating CSCs are now
performed in various types of solid tumors such as glioblastoma,
colon cancer, breast cancer and prostate cancer (3–7).
Undifferentiated-cell (normal stem cell) markers such as Nanog,
OCT3/4 and SOX2 are known to be expressed in a small population of
cancer cells (8–17). Increasing evidence suggests that
tumor tissues are not homogeneous but heterogeneous supporting the
CSC hypothesis.
Gremlin 1 is one of the bone morphogenetic protein
(BMP) antagonists and especially antagonizes with BMP2, BMP4 and
BMP7 (18,19). BMPs are members of the transforming
growth factor (TGF)-β superfamily and play an essential role during
development and differentiation while Gremlin 1 is also related to
differentiation in combination with BMPs and its dysregulation
results in developmental diseases (20–23).
Gremlin 1 is overexpressed in various types of human cancers
(24–26). Recently several studies have
demonstrated that CSCs prevent the BMP differentiating effect by
secreting Gremlin 1 in glioblastoma, suggesting that Gremlin 1
plays a role in maintenance of CSC properties (27,28).
In gynecological malignancies such as ovarian, endometrial and
cervical cancers, various techniques to isolate cells with CSC-like
properties have been reported. For instance, CD44- or
CD133-positive cells are known to be more tumorigenic than negative
ones in ovarian cancer (29,30).
Similar findings have been suggested for side population (SP) cells
in endometrial cancer and ALDH1-positive cells in cervical cancer
(31–36). Gremlin 1 is also reported to be
expressed in cervical cancer compared to normal tissues of the
cervix (37). A few studies have
investigated the significance of BMPs in cervical cancer (38–40),
however, there are no studies on the clinico-pathological
significance of Gremlin 1 in cervical cancer nor on the function of
Gremlin 1 in CSC-like cells of cervical cancer.
Viewing the importance of Gremlin 1 in CSC
maintenance and expression of Gremlin 1 in cervical cancer tissue,
we hypothesized that Gremlin 1 may be correlated with the prognosis
of cervical cancer through maintaining CSC properties. In the
present study, we first demonstrated the clinicopathological
significance of Gremlin 1 in stage I–II cervical cancer patients
and the in vitro promoting effects of Gremlin 1 on CSC
properties in cervical cancer cells.
Materials and methods
Patients
The present study was approved by the Institutional
Ethics Committee, and written informed consent was obtained from
each patient. Patients with a diagnosis of cervical cancer (stage
I–II) who underwent primary surgery at the University of Tokyo
Hospital from 2005 to 2014 were enrolled. A total of 104 samples
were obtained.
RNA extraction and RT-quantitative PCR
(RT-qPCR)
Cervical cancer tissues (100–200 mg) were collected
and snap frozen with liquid nitrogen and preserved at −80°C.
Tissues were homogenized using MagNA Lyser Instrument (Roche
Diagnostics, Mannheim, Germany) and total RNA was extracted with a
Tissue Total RNA kit (Favorgen Biotech Corp., Ping-Tung, Taiwan)
according to the manufacturer's protocols. First-strand cDNA was
synthesized (reverse transcription reactions) from 1 µg of
total RNA using ReverTra Ace (Toyobo, Osaka, Japan). RT-qPCR was
performed with SYBR-Green PCR Master Mix (Roche Diagnostics)
according to the manufacturer's instructions. β-actin was used as a
housekeeping gene, and the results are represented as fold-change
relative to β-actin expression (2−ΔΔCt). The sequences
of the primer pairs used were as follows: Gremlin 1, forward,
5′-TCATCAACCGCTTCTGTTACGGC-3′ and reverse,
5′-CAGAAGGAGCAGGACTGAAAGG-3′; Nanog, forward
5′-GCTGAGATGCCTCACACGGAG-3′ and reverse
5′-TCTGTTTCTTGACCGGGACCTTGTC-3′; OCT3/4, forward
5′-TGGAGAAGGAGAAGCTGGAGCAAAA-3′ and reverse
5′-GGCAGATGGTCGTTTGGCTGAATA-3′; SOX2, forward
5′-GGAAATGGGAGGGGTGCAAAAGAGG-3′ and reverse
5′-TTGCGTGAGTGTGGATGGGATTGGTG-3′; β-actin, forward
5′-CTGGAACGGTGAAGGTGACA-3′ and reverse
5′-AAGGGACTTCCTGTAACAACGCA-3′. Denaturation was performed at 95°C
for 2 min, followed by 35 cycles at 98°C for 10 sec, at 65°C for 10
sec and at 68°C for 8 sec. Each experiment was performed in
triplicate and repeated three times.
Cell culture
The human squamous cell carcinoma cell line CaSki
(derived from a metastatic site; small intestine) was obtained from
the American Type Culture Collection (ATCC, Manassas, VA, USA) and
cultured in adherent conditions in Dulbecco's modified Eagle's
medium (DMEM; Wako Pure Chemical Industries Ltd., Osaka, Japan)
supple mented with 10% fetal bovine serum (FBS; Invitrogen Life
Technologies, Carlsbad, CA, USA) and sub-cultured by 0.25%
trypsin/EDTA (Wako Pure Chemical Industries Ltd.) detachment. For
the sphere-forming assays, DMEM/F12 (Invitrogen Life Technologies)
supplemented with 20 ng/ml human recombinant epidermal growth
factor (EGF; Wako Pure Chemical Industries Ltd.), 10 ng/ml basic
fibroblastic growth factor (bFGF; ReproCELL, Inc., Kanagawa, Japan)
and 0.3% bovine serum albumin (BSA; Sigma-Aldrich Co., St. Louis,
MO, USA) (sphere medium) was used. Each medium contained no
antibiotics. All cells were grown and treated in a humidified
atmosphere at 37°C and 5% CO2. When 60–70% confluence
was reached, the medium was replaced with DMEM containing 0.5% FBS
and 1,000 ng/ml human recombinant Gremlin 1 (R&D Systems Inc.,
Minneapolis, MN, USA) or vehicle for another 24 h and total RNA was
extracted.
Flow cytometry
The ALDH enzymatic activity of the cells was
measured using the ALDEFLUOR kit (StemCell Technologies, Vancouver,
BC, Canada) according to the manufacturer's protocol. CaSki cells
(1×106 cells) were suspended in ALDEFLUOR assay buffer
containing ALDH substrate. The brightly fluorescent ALDH-positive
cells were detected using FACSCalibur flow cytometer (BD
Biosciences, San Jose, CA, USA). As a negative control, cells were
stained under identical conditions after treatment with the
specific ALDH inhibitor N,N-diethylaminobenzaldehyde (DEAB). For
obtaining consistent results, the following conditions were
applied; 2×105 cells were seeded onto a 6-well
multi-plate dish in triplicate for 48 h and medium was switched to
DMEM containing 3% FBS and Gremlin 1 (1,000 ng/ml) or vehicle for
another 24 h and flow cytometry was performed as described above.
Each experiment was repeated three times.
Sphere-forming assay
CaSki cells were cultured under the conditions
described for flow cytometry, and then the sphere-forming assay was
performed. Dissociated single cells (1,000 cells) were seeded onto
ultra-low attachment multi-plate (24-well) dish (Corning Inc.,
Corning, NY, USA) in quadruplicate and cultured in sphere medium
for another 7 days. After that, spheroids larger than 100 µm
in diameter were counted, and the number of spheroids was compared
between the Gremlin 1-exposed cells and the control (vehicle). Each
experiment was repeated three times.
Statistical analysis
Known clinicopathological prognostic factors and
expression of Gremlin 1 mRNA were analyzed by univariate Cox
proportional hazards regression models, and the relative risk (RR)
with 95% confidence intervals (CIs) was evaluated. Logistic
regression analysis was performed to investigate the relationship
between Gremlin 1 mRNA expression level and recurrence. Using this
result, receiver operating characteristic (ROC) curves were used to
assess the criterion value. Kaplan-Meier curves were used to
estimate the probability of OS and PFS, and the log-rank test was
used to compare survival between the two groups (high Gremlin 1
mRNA or low Gremlin 1 mRNA levels). A χ2 test was used
to evaluate the relationship between expression of Gremlin 1 and
each clinicopathological factor. For the in vitro study, the
Student's t-test was used to compare the data obtained for the
controls. P<0.05 was considered to indicate a statistically
significant difference. JMP® (SAS Institute) was used
for statistical analysis.
Results
Expression of Gremlin 1 in cervical
cancer tissues is significantly correlated with progression-free
survival (PFS) and recurrence
Characteristics of the 104 patients whose cancer
tissues were collected are shown in Table I. Expression of Gremlin 1 mRNA was
not a significant prognostic factor for overall survival (OS) in
the univariate analysis (Table II)
but was a prognostic factor for PFS in both univariate and
multivariate analyses as well as histology (non-squamous carcinoma)
(Table III). Expression of
Gremlin 1 mRNA was also significantly correlated with recurrence in
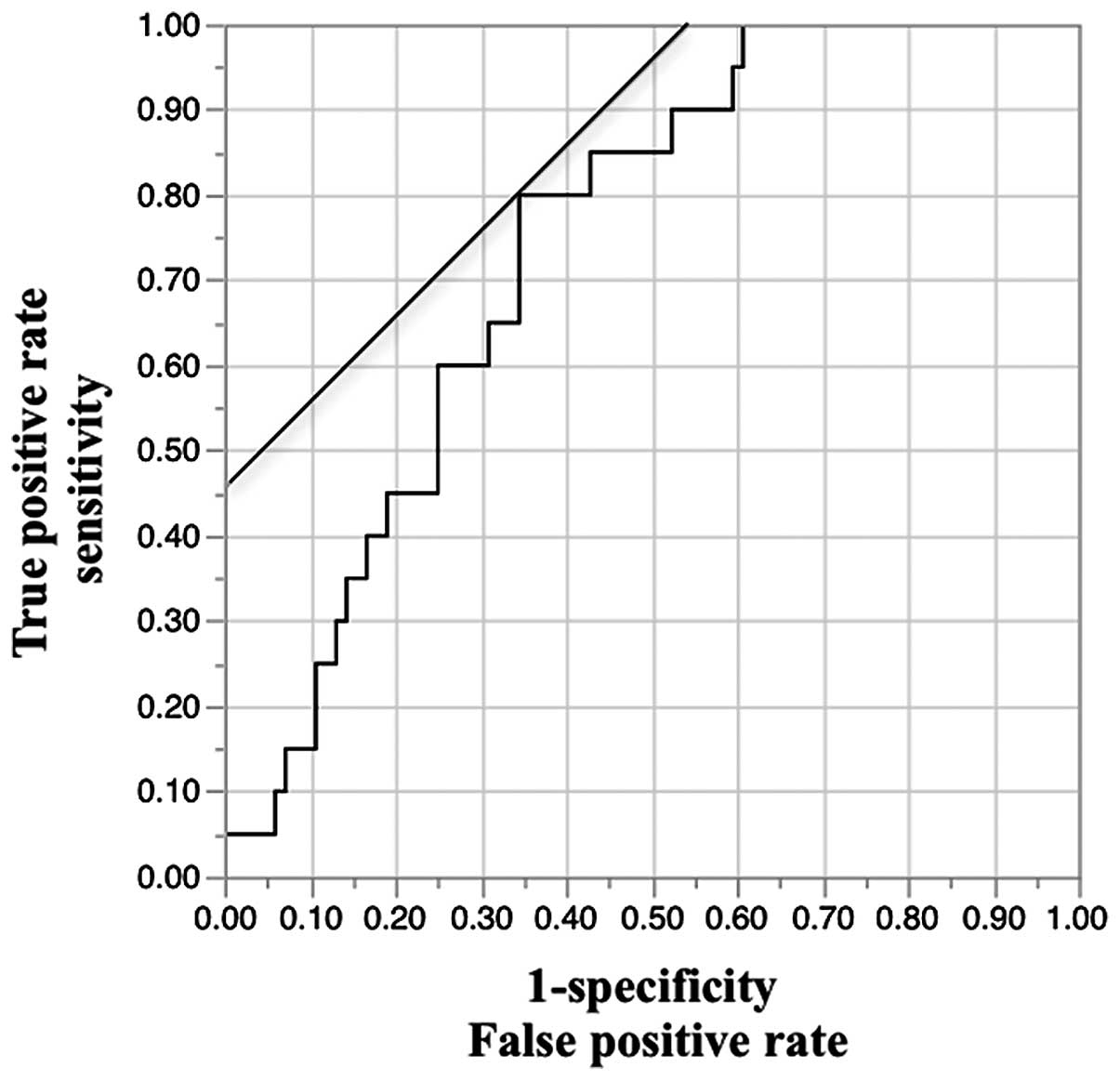
the logistic regression analysis. Cut-off value for high and low
expression was set using ROC curves; high expression of Gremlin 1
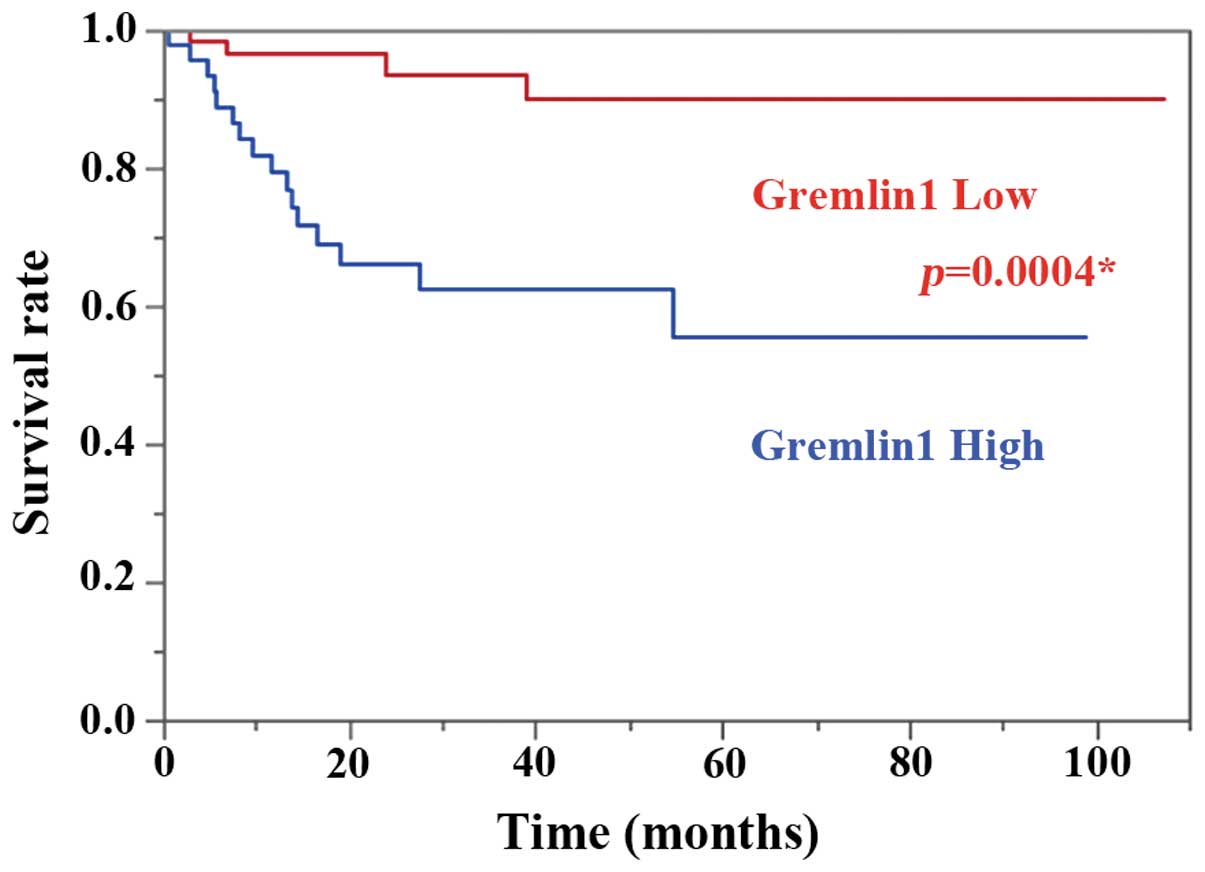
mRNA, >0.20; low expression, <0.20 (Fig. 1). Kaplan-Meier curves for OS and PFS
in patients with high and low Gremlin 1 mRNA expression are shown
in Fig. 2. High expression of
Gremlin 1 mRNA was a significant prognostic factor for PFS
(log-rank test, P=0.0004).
 | Table ICharacteristics of the cervical
cancer patients. |
Table I
Characteristics of the cervical
cancer patients.
| Features | Cases (n) | Percentage (%) |
|---|
| Mean age (range) in
years | 44.0
(22.9–75.1) |
| FIGO stage |
| Stage I | 69 | 66 |
| Stage II | 35 | 34 |
| Histology |
| SCC | 59 | 56 |
| Non-SCC | 45 | 44 |
| Tumor size |
| ≤4 cm | 46 | 44 |
| >4 cm | 58 | 56 |
| Pm invasion |
| Negative | 79 | 75 |
| Positive | 25 | 25 |
| Stromal
invasion |
| ≤2/3 | 48 | 46 |
| >2/3 | 56 | 54 |
| Margin |
| Negative | 98 | 94 |
| Positive | 6 | 6 |
| LVSI |
| Negative | 43 | 41 |
| Positive | 61 | 59 |
| LN metastasis |
| Negative | 73 | 70 |
| Positive | 31 | 30 |
| Recurrence |
| Negative | 84 | 81 |
| Positive | 20 | 19 |
| Table IIPredictive factors for overall
survival (OS). |
Table II
Predictive factors for overall
survival (OS).
| Features | Univariate
| Multivariate
|
|---|
| Relative risk (95%
CI) | P-value | Relative risk (95%
CI) | P-value |
|---|
| Stage (II vs.
I) | 2.91
(0.60–16.07) | 0.1782 | 1.44
(0.28–9.15) | 0.6624 |
| Histology (NS vs.
SCC) | 3.13
(0.66–22.04) | 0.1508 | 4.11
(0.86–29.21) | 0.0758 |
| Bulky (>4 cm)
tumor (positive vs. negative) | 5.14
(0.86–97.49) | 0.0739 | 1.81
(0.28–35.39) | 0.5679 |
| Pm invasion
(positive vs. negative) | 0.61
(0.032–3.81) | 0.6404 | – | – |
| Stromal invasion
(>2/3 vs. ≤2/3) | 1.34
(0.27–7.41) | 0.7114 | – | – |
| Margin (positive
vs. negative) | 3.26
(0.16–20.46) | 0.3458 | – | – |
| LVSI (positive vs.
negative) | 1.28e9
(2.23-#) | 0.0064a | 1.04e9
(1.31–111) | 0.0064a |
| LN metastasis
(positive vs. negative) | 2.02
(0.38–9.65) | 0.3796 | – | – |
| Expression of
Gremlin 1 | 0.39
(0.017–1.54) | 0.3354 | – | – |
| Table IIIPredictive factors for
progression-free survival (PFS). |
Table III
Predictive factors for
progression-free survival (PFS).
| Features | Univariate
| Multivariate
|
|---|
| Relative risk (95%
CI) | P-value | Relative risk (95%
CI) | P-value |
|---|
| Stage (II vs.
I) | 3.30
(1.36–8.43) | 0.0082a | 1.84
(0.69–5.27) | 0.3247 |
| Histology (NS vs.
SCC) | 2.57
(1.05–6.86) | 0.0379a | 4.01
(1.55–11.51) | 0.0077a |
| Bulky (>4 cm)
tumor (positive vs. negative) | 4.86
(1.63–20.84) | 0.0030 | 2.85
(0.86–13.10) | 0.1021 |
| Pm invasion
(positive vs. negative) | 2.20
(0.86–5.33) | 0.0959 | – | – |
| Stromal invasion
(>2/3 vs. ≤2/3) | 1.59
(0.65–4.24) | 0.3108 | – | – |
| Margin (positive
vs. negative) | 1.06
(0.05–5.18) | 0.9491 | – | – |
| LVSI (positive vs.
negative) | 6.84
(1.97–43.04) | 0.0011a | 3.86
(0.92–26.41) | 0.0538 |
| LN metastasis
(positive vs. negative) | 2.37
(0.97–5.79) | 0.0565 | – | – |
| Expression of
Gremlin 1 mRNA | 1.43
(1.05–1.78) | 0.0281a | 1.41
(1.01–1.78) | 0.0036a |
High expression of Gremlin 1 in cervical
cancer tissues is significantly correlated with a bulky tumor
The correlations between expression of Gremlin 1
mRNA and each prognosis factor are listed in Table IV. Of note, high expression of
Gremlin 1 mRNA was significantly correlated with a bulky (>4 cm)
tumor (χ2 test, P=0.0494).
| Table IVCorrelation between expression of
Gremlin 1 mRNA (high or low) and each risk clinicopathological
factor. |
Table IV
Correlation between expression of
Gremlin 1 mRNA (high or low) and each risk clinicopathological
factor.
| Features | Relative risk (95%
CI) | P-value |
|---|
| Stage (II vs.
I) | 1.38
(0.81–2.37) | 0.2325 |
| Histology (NS vs.
SCC) | 1.14
(0.37–1.77) | 0.5415 |
| Bulky (>4 cm)
tumor (positive vs. negative) | 1.40
(1.001–1.971) | 0.0494a |
| Pm invasion
(positive vs. negative) | 1.96
(0.97–3.95) | 0.0534 |
| Stromal invasion
(>2/3 vs. ≤2/3) | 1.05
(0.74–1.51) | 0.7600 |
| Margin (positive
vs. negative) | 2.62
(0.50–13.68) | 0.2345 |
| LVSI (positive vs.
negative) | 1.04
(0.75–1.43) | 0.8076 |
| LN metastasis
(positive vs. negative) | 1.59
(0.88–2.87) | 0.0189 |
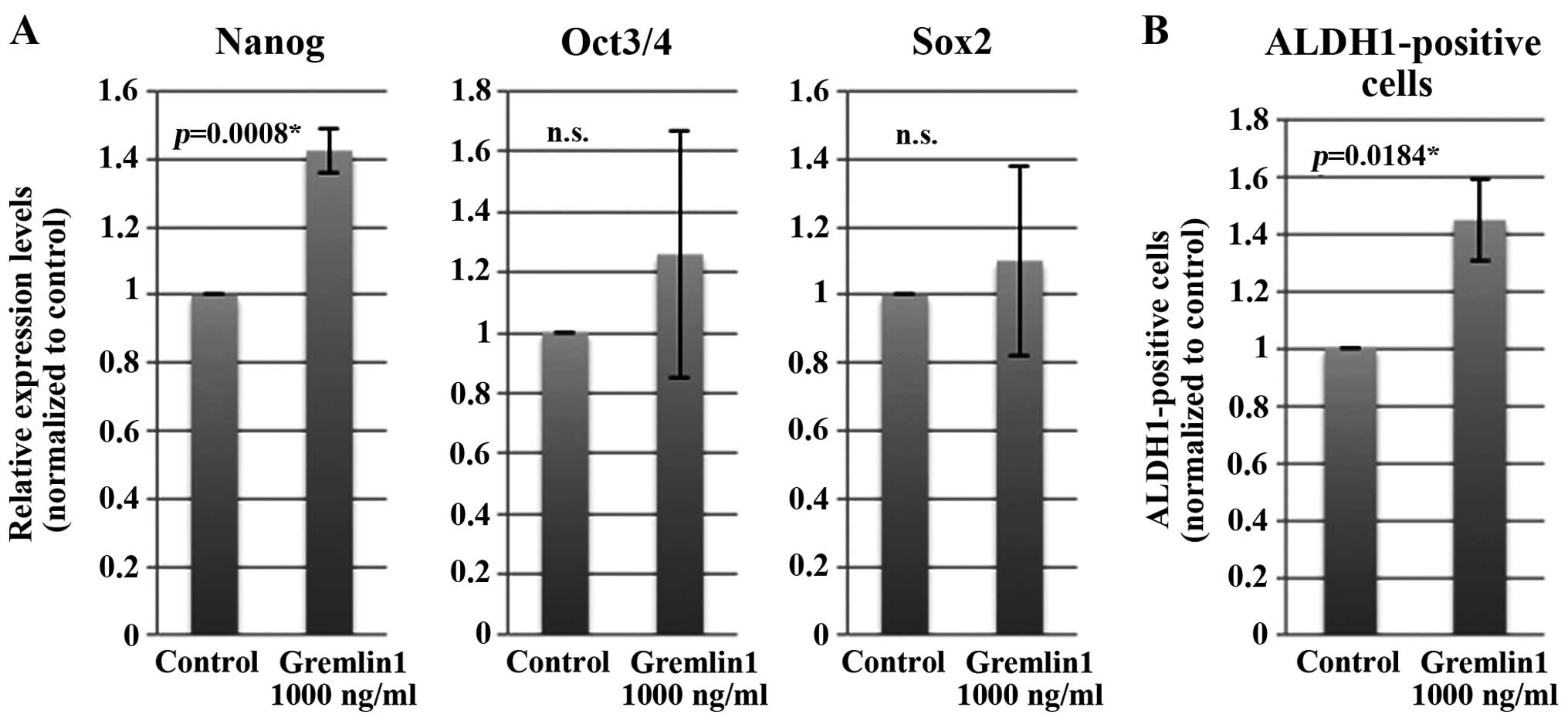
Exposure of CaSki cells to Gremlin 1
increases Nanog mRNA expression levels and the percentage of
ALDH1-positive cells in vitro
We then speculated whether the clinical significance
of Gremlin 1 expression is attributed to CSC-like properties. We
decided to use a CaSki cell in vitro model as CaSki cells
are derived from a recurrent and metastatic site. Cancer stem cells
are thought to share normal stem cell features and express
undifferentiated-cell markers such as Nanog, Oct3/4 and Sox2.
ALDH1-positive cells are reported to be a marker of cancer
stem-like cells in various types of solid tumors including cervical
cancer. After culturing CaSki cells for 48 h, we exposed cells to
Gremlin 1 (1,000 ng/ml) or vehicle control for 24 h and assessed
these markers. Exposing CaSki cells to Gremlin 1 significantly
increased the expression of Nanog mRNA but not Oct3/4 and Sox2 mRNA
(Fig. 3A). Likewise, exposing CaSki
cells to Gremlin 1 significantly (~1.41-fold) increased the
population of ALDH1-positive cells compared to the control
(Fig. 3B). These results were not
obtained when the concentration of Gremlin 1 was reduced to 500
ng/ml (data not shown).
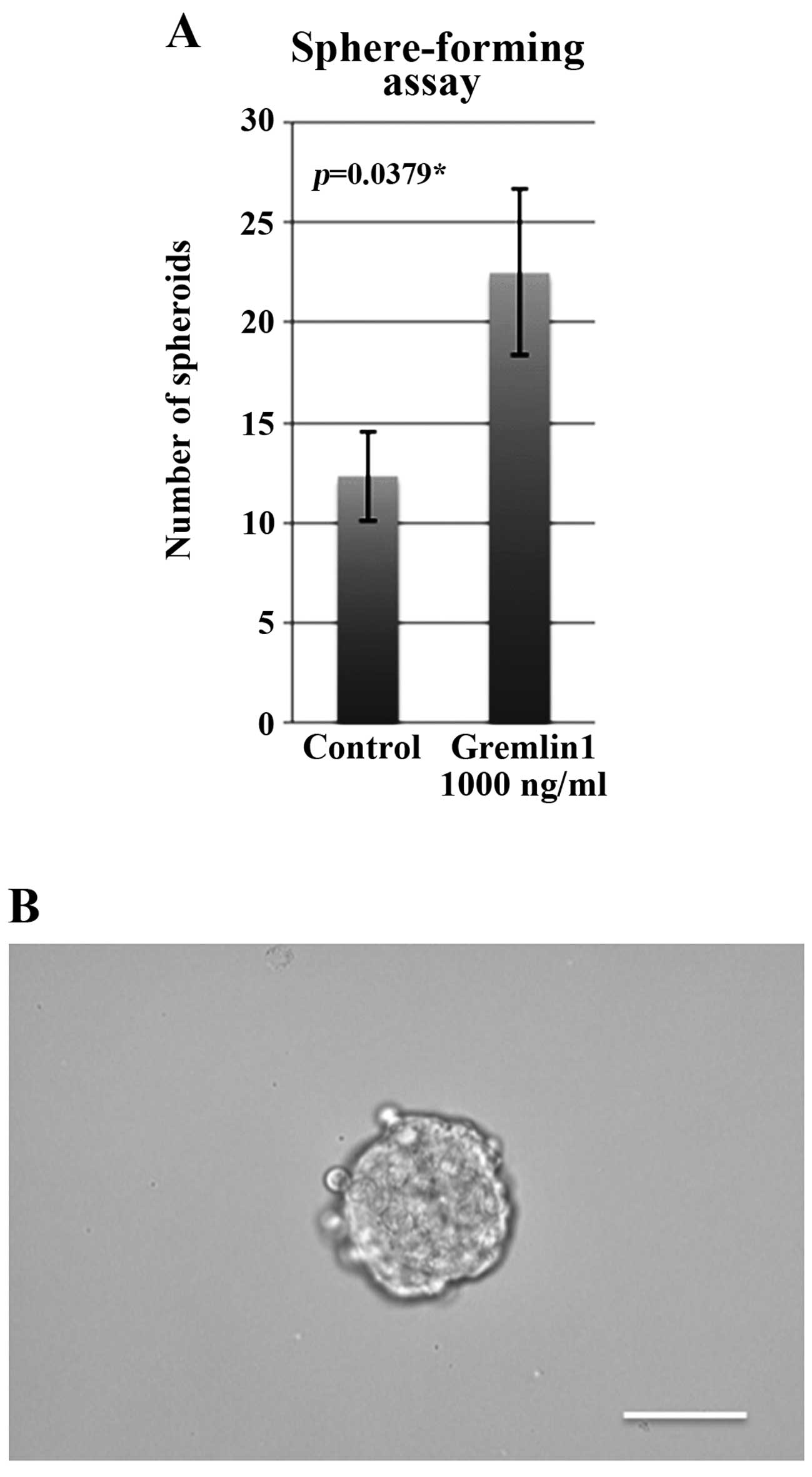
Exposing CaSki cells to Gremlin 1
increases sphere-forming ability
We then investigated the sphere-forming ability of
Gremlin 1-exposed cells. Significantly more spheroids were formed
when 1,000 cells of Gremlin 1-exposed cells were seeded compared to
the control (Fig. 4A). A
representative counted spheroid (>100 µm in diameter) is
shown in Fig. 4B.
Discussion
Here, we investigated the clinical significance of
Gremlin 1 in cervical cancer and then investigated its effects on
cancer cells in vitro.
The CSC theory states that only a small population
of cells within a tumor is tumorigenic and has the ability for
self-renewal. Increasing evidence suggests that tumor tissues are
not homogeneous but heterogeneous and supports the CSC hypothesis
in cervical cancer as well. Recently, Gremlin 1 has been suggested
to be secreted by CSCs in glioblastoma and prevents CSCs from
differentiating by its antagonistic effect on BMPs.
First, we showed that high expression of Gremlin 1
mRNA in cervical cancer tissues is a significant prognostic factor
for PFS and clinical recurrence. High expression of Gremlin 1 mRNA
was significantly correlated with a bulky (>4 cm) tumor, which
is one of the most malignant features of cervical cancer. However,
these findings should not be interpreted to conclude that cases of
poor prognosis have a larger population of CSCs. Gremlin 1 was
found to be secreted from stromal cancer-associated fibroblast
(CAFs) in colon cancer and its expression was localized to invasion
fronts (24–26). On the other hand, Gremlin 1 was
found to be secreted by CSCs in glioblastoma and was thought to
prevent CSCs from differentiating by antagonizing the effects
ofBMPs (27,28). Although there are no common views
about the distribution pattern of Gremlin 1 expression in cervical
cancer (37), we may safely say
that a microenvironment of high expression of Gremlin 1 is
favorable for CSC maintenance or invasion.
In an in vitro study, we revealed the effects
of Gremlin 1 on CSC-like properties. The exposure of CaSki cells to
Gremlin 1 (1,000 ng/ml) increased: i) Nanog mRNA expression levels
(but not Oct3/4 and Sox2 mRNA expression levels), ii) the
population of ALDH-positive cells (1.41-fold higher compared with
the control) and iii) sphere-forming ability when 1,000 Gremlin
1-exposed cells were seeded.
The effects of Gremlin 1 may have been obtained by
preventing CSCs from differentiating or promoting non-CSCs to
de-differentiate. We investigated changes in the population of
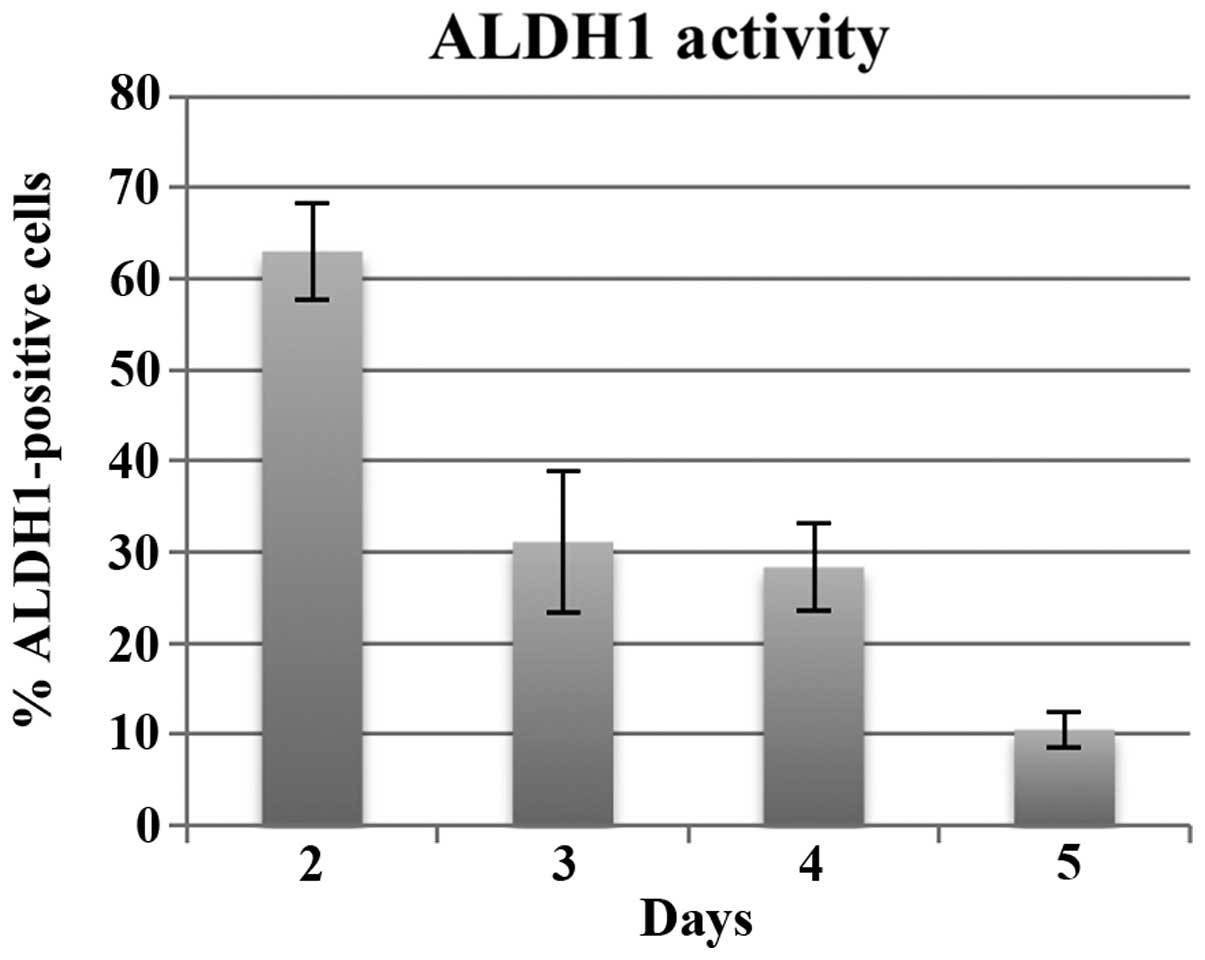
ALDH1-positive CaSki cells in normal culture medium at each day
after 48 h of seeding (Fig. 5).
After 48 h of seeding, >60% of adherent cells were
ALDH1-positive and those percentages were reduced to ~30% upon
reaching confluency, which are compatible with previous reports
that the ALDH1-positive population in CaSki cells are 20–30%
(32). In our experiments of
Gremlin 1-exposure, the cells were ~60–70% confluent after 48 h of
seeding (when we exposed cells to Gremlin 1) and became 90–100%
confluent when assessed after 72 h of seeding. Considering this
fact and a previous report that ALDH1-negative cells do not become
ALDH-positive (32), the effects of
Gremlin 1 that we observed were achieved by preventing CSCs from
differentiating to some extent resulting in increased
sphere-forming ability.
Gremlin 1 specifically antagonizes BMP2, BMP4 and
BMP7 function among the various members of BMPs (19). We performed the same experiments
using BMP2, BMP4 and BMP7 (500 ng/ml). Only exposure of CaSki cells
to BMP7 significantly reduced the expression of Nanog mRNA
expression. However, the population of ALDH1-positive cells was not
significantly different even between BMP7-exposed cells and the
vehicle control (data not shown). This fact suggests that the
BMP/Gremlin 1 axis is complex and we should not only consider high
and low expression of BMPs or Gremlin 1 but the combination is
important. Gremlin 1 is thought to have various effects on non-BMP
signaling (41). We performed the
same experiments using LDN-193189 [a small molecule inhibitor of
BMP type I receptors ALK2 and ALK3 (42–44)]
to confirm that our obtained results of Gremlin 1 were via BMP
signaling. Exposure of CaSki cells to LDN-193189 increased the
expression of Nanog mRNA in a dose-dependent manner and
significantly high expression of Nanog mRNA was observed compared
to the vehicle control when the concentration of LDN-193189 was 100
nM (data not shown).
In conclusion, we demonstrated the clinical
significance of Gremlin 1 in cervical cancer and insights into its
effects on CSC-like properties in vitro were shown. In
cervical cancer, it is suggested that Gremlin 1 may play a role in
clinical recurrence and maintaining CSC-like properties.
References
|
1
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer, and cancer stem cells. Nature.
414:105–111. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Meacham CE and Morrison SJ: Tumour
heterogeneity and cancer cell plasticity. Nature. 501:328–337.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dick JE and JE D: Stem cell concepts renew
cancer research. Blood. 112:4793–4807. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen X, Rycaj K, Liu X and Tang DG: New
insights into prostate cancer stem cells. Cell Cycle. 12:579–586.
2013. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Skibinski A and Kuperwasser C: The origin
of breast tumor heterogeneity. Oncogene. 475:1–8. 2015.
|
|
6
|
Triscott J, Rose Pambid M and Dunn SE:
Concise review: Bullseye: Targeting cancer stem cells to improve
the treatment of gliomas by repurposing disulfiram. Stem Cells.
33:1042–1046. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zeki SS, Graham TA and Wright NA: Stem
cells and their implications for colorectal cancer. Nat Rev
Gastroenterol Hepatol. 8:90–100. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu CWLC, Li CH, Peng YJ, Cheng YW, Chen
HW, Liao PL, Kang JJ and Yeng MH: Snail regulates Nanog status
during the epithelial-mesenchymal transition via the
Smad1/Akt/GSK3β signaling pathway in non-small-cell lung cancer.
Oncotarget. 5:3880–3894. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang D, Lu P, Zhang H, Luo M, Zhang X, Wei
X, Gao J, Zhao Z and Liu C: Oct-4 and Nanog promote the
epithelial-mesenchymal transition of breast cancer stem cells and
are associated with poor prognosis in breast cancer patients.
Oncotarget. 5:10803–10815. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Iv Santaliz-Ruiz LE, Xie X, Old M, Teknos
TN and Pan Q: Emerging role of nanog in tumorigenesis and cancer
stem cells. Int J Cancer. 135:2741–2748. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kregel S, Szmulewitz RZ and Vander Griend
DJ: The pluripotency factor Nanog is directly upregulated by the
androgen receptor in prostate cancer cells. Prostate. 74:1530–1543.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Loh YH, Wu Q, Chew JL, Vega VB, Zhang W,
Chen X, Bourque G, George J, Leong B, Liu J, et al: The Oct4 and
Nanog transcription network regulates pluripotency in mouse
embryonic stem cells. Nat Genet. 38:431–440. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Miyazawa K, Tanaka T, Nakai D, Morita N
and Suzuki K: Immunohistochemical expression of four different stem
cell markers in prostate cancer: High expression of NANOG in
conjunction with hypoxia-inducible factor-1α expression is involved
in prostate epithelial malignancy. Oncol Lett. 8:985–992.
2014.PubMed/NCBI
|
|
14
|
Kumazawa S, Kajiyama H, Umezu T, Mizuno M,
Suzuki S, Yamamoto E, Mitsui H, Sekiya R, Shibata K and Kikkawa F:
Possible association between stem-like hallmark and radioresistance
in human cervical carcinoma cells. J Obstet Gynaecol Res.
40:1389–1398. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu XF, Yang WT, Xu R, Liu JT and Zheng
PS: Cervical cancer cells with positive Sox2 expression exhibit the
properties of cancer stem cells. PLoS One. 9:e870922014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shen L, Huang X, Xie X, Su J, Yuan J and
Chen X: High expression of SOX2 and OCT4 indicates radiation
resistance and an independent negative prognosis in cervical
squamous cell carcinoma. J Histochem Cytochem. 62:499–509. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang L, Guo H, Lin C, Yang L and Wang X:
Enrichment and characterization of cancer stem like cells from a
cervical cancer cell line. Mol Med Rep. 9:2117–2123.
2014.PubMed/NCBI
|
|
18
|
Khokha MK, Hsu D, Brunet LJ, Dionne MS and
Harland RM: Gremlin is the BMP antagonist required for maintenance
of Shh and Fgf signals during limb patterning. Nat Genet.
34:303–307. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Church RH, Krishnakumar A, Urbanek A,
Geschwindner S, Meneely J, Bianchi A, Basta B, Monaghan S, Elliot
C, Strömstedt M, et al: Gremlin 1 preferentially binds to bone
morphogenetic protein-2 (BMP-2) and BMP-4 over BMP-7. Biochem J.
466:55–68. 2015. View Article : Google Scholar
|
|
20
|
Wang RN, Green J, Wang Z, Deng Y, Qiao M,
Peabody M, Zhang Q, Ye J, Yan Z, Denduluri S, et al: Bone
morphogenetic protein (BMP) signaling in development and human
diseases. Genes Dis. 1:87–105. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ducy P and Karsenty G: The family of bone
morphogenetic proteins. Kidney Int. 57:2207–2214. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ciuclan L, Sheppard K, Dong L, Sutton D,
Duggan N, Hussey M, Simmons J, Morrell NW, Jarai G, Edwards M, et
al: Treatment with anti-gremlin 1 antibody ameliorates chronic
hypoxia/SU5416-induced pulmonary arterial hypertension in mice. Am
J Pathol. 183:1461–1473. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Worthley DL, Churchill M, Compton JT,
Tailor Y, Rao M, Si Y, Levin D, Schwartz MG, Uygur A, Hayakawa Y,
et al: Gremlin 1 identifies a skeletal stem cell with bone,
cartilage, and reticular stromal potential. Cell. 160:269–284.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Karagiannis GS, Musrap N, Saraon P, Treacy
A, Schaeffer DF, Kirsch R, Riddell RH and Diamandis EP: Bone
morphogenetic protein antagonist gremlin-1 regulates colon cancer
progression. Biol Chem. 396:163–183. 2015. View Article : Google Scholar
|
|
25
|
Karagiannis GS, Treacy A, Messenger D,
Grin A, Kirsch R, Riddell RH and Diamandis EP: Expression patterns
of bone morphogenetic protein antagonists in colorectal cancer
desmoplastic invasion fronts. Mol Oncol. 8:1240–1252. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mulvihill MS, Kwon YW, Lee S, Fang LT,
Choi H, Ray R, Kang HC, Mao JH, Jablons D and Kim IJ: Gremlin is
overexpressed in lung adenocarcinoma and increases cell growth and
proliferation in normal lung cells. PLoS One. 7:e422642012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Seoane J: Gremlins sabotage the mechanisms
of cancer stem cell differentiation. Cancer Cell. 25:716–717. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yan K, Wu Q, Yan DH, Lee CH, Rahim N,
Tritschler I, DeVecchio J, Kalady MF, Hjelmeland AB and Rich JN:
Glioma cancer stem cells secrete Gremlin 1 to promote their
maintenance within the tumor hierarchy. Genes Dev. 28:1085–1100.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Curley MD, Therrien VA, Cummings CL,
Sergent PA, Koulouris CR, Friel AM, Roberts DJ, Seiden MV, Scadden
DT, Rueda BR, et al: CD133 expression defines a tumor initiating
cell population in primary human ovarian cancer. Stem Cells.
27:2875–2883. 2009.PubMed/NCBI
|
|
30
|
Zhang S, Balch C, Chan MW, Lai HC, Matei
D, Schilder JM, Yan PS, Huang TH and Nephew KP: Identification and
characterization of ovarian cancer-initiating cells from primary
human tumors. Cancer Res. 68:4311–4320. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Friel AM, Sergent PA, Patnaude C, Szotek
PP, Oliva E, Scadden DT, Seiden MV, Foster R and Rueda BR:
Functional analyses of the cancer stem cell-like properties of
human endometrial tumor initiating cells. Cell Cycle. 7:242–249.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu SY and Zheng PS: High aldehyde
dehydrogenase activity identifies cancer stem cells in human
cervical cancer. Oncotarget. 4:2462–2475. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bortolomai I, Canevari S, Facetti I, De
Cecco L, Castellano G, Zacchetti A, Alison MR and Miotti S: Tumor
initiating cells: Development and critical characterization of a
model derived from the A431 carcinoma cell line forming spheres in
suspension. Cell Cycle. 9:1194–1206. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
López J, Valdez-Morales FJ,
Benítez-Bribiesca L, Cerbón M and Carrancá AG: Normal and cancer
stem cells of the human female reproductive system. Reprod Biol
Endocrinol. 11:532013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Maddox J, Shakya A, South S, Shelton D,
Andersen JN, Chidester S, Kang J, Gligorich KM, Jones DA, Spangrude
GJ, et al: Transcription factor Oct1 is a somatic and cancer stem
cell determinant. PLoS Genet. 8:e10030482012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yao T, Wu Z, Liu Y, Rao Q and Lin Z:
Aldehyde dehydrogenase 1 (ALDH1) positivity correlates with poor
prognosis in cervical cancer. J Int Med Res. 42:1038–1042. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Namkoong H, Shin SM, Kim HK, Ha SA, Cho
GW, Hur SY, Kim TE and Kim JW: The bone morphogenetic protein
antagonist gremlin 1 is overexpressed in human cancers and
interacts with YWHAH protein. BMC Cancer. 6:742006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liu CY, Chao TK, Su PH, Lee HY, Shih YL,
Su HY, Chu TY, Yu MH, Lin YW and Lai HC: Characterization of LMX-1A
as a metastasis suppressor in cervical cancer. J Pathol.
219:222–231. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tsuchida R, Osawa T, Wang F, Nishii R, Das
B, Tsuchida S, Muramatsu M, Takahashi T, Inoue T, Wada Y, et al:
BMP4/Thrombospondin-1 loop paracrinically inhibits tumor
angiogenesis and suppresses the growth of solid tumors. Oncogene.
33:3803–3811. 2014. View Article : Google Scholar
|
|
40
|
Cassar L, Li H, Pinto AR, Nicholls C,
Bayne S and Liu JP: Bone morphogenetic protein-7 inhibits
telomerase activity, telomere maintenance, and cervical tumor
growth. Cancer Res. 68:9157–9166. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Mitola S, Ravelli C, Moroni E, Salvi V,
Leali D, Ballmer-Hofer K, Zammataro L and Presta M: Gremlin is a
novel agonist of the major proangiogenic receptor VEGFR2. Blood.
116:3677–3680. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Fotinos A, Nagarajan N, Martins AS, Fritz
DT, Garsetti D, Lee AT, Hong CC and Rogers MB: Bone morphogenetic
protein-focused strategies to induce cytotoxicity in lung cancer
cells. Anticancer Res. 34:2095–2104. 2014.PubMed/NCBI
|
|
43
|
Sanvitale CE, Kerr G, Chaikuad A, Ramel
MC, Mohedas AH, Reichert S, Wang Y, Triffitt JT, Cuny GD, Yu PB, et
al: A new class of small molecule inhibitor of BMP signaling. PLoS
One. 8:e627212013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Vogt J, Traynor R and Sapkota GP: The
specificities of small molecule inhibitors of the TGFß and BMP
pathways. Cell Signal. 23:1831–1842. 2011. View Article : Google Scholar : PubMed/NCBI
|