Introduction
Lung cancer occurs and develops as a complicated
result of an accumulation of various endogenous and exogenous
effects (1,2). Gene alterations participate in lung
cancer genesis. ING5 inhibits cancer aggressiveness via preventing
EMT and is a potential prognostic biomarker for lung cancer
(3). Fibulin-5 is a metastasis
suppressor in lung cancer by modulating the tumor microenvironment
to suppress Wnt/β-catenin signaling (4). Fra-1 enhances lung cancer epithelial
cell motility and invasion by inducing the activity of MMPs, in
particular MMP-2 and MMP-9, and EGFR-activated signaling (5). It has been shown that inactivation of
tumor suppressor genes and activation of oncogenes play a
significant role in carcinogenesis. However, the etiology of lung
cancer remains poorly understood.
Fos-related antigen-1 (Fra-1, also known as FOSL1)
is a member of the activator protein-1 (AP-1) transcription factor
superfamily (6,7). Fra-1 can positively regulate
transcription. Fra-1 activity is regulated transcriptionally as
well as post-translationally (8,9). It is
overexpressed in a variety of cancers, including lung, breast,
colon and brain. High Fra-1 levels are associated with enhanced
cell proliferation, migration, invasion and survival (6–9). Fra-1
plays a role in the progression and prognosis of non-small cell
lung cancer (NSCLC) (10). Fra-1
can promote motility, invasion, and anchorage-independent growth of
lung epithelial cells in vitro, but is insufficient for
tumor formation (11). Although
some evidence has been reported, the mechanism of Fra-1 in
malignancy is not fully understood. Meanwhile, at present, few
studies have investigated the relationship between Fra-1 and lung
cancer. Thus, research on the effect and mechanism of Fra-1 in lung
cancer is warranted.
TP53 is a crucial transcription factor and an
important sensor of cellular stress under genotoxic, pathological,
chemotoxic and even normal physiological conditions (12,13).
MDM2 is a negative regulator of TP53. Abnormalities in the TP53
gene and overexpression of MDM2 are commonly observed in
malignancies. The MDM2-P53 feedback loop plays an important role in
tumor progression (14,15). Inactivation of the TP53
tumor-suppressor gene is a common mechanism that cancer cells
require to proliferate and escape pre-programmed cell death
(16).
In this study, we examined the expression levels of
Fra-1 in lung cancer tissues. At the same time, we studied the
influence of Fra-1 on lung cancer and explored the possible
mechanism.
Materials and methods
Cell culture
One identified general human lung cancer cell line,
H460, was cultured in RPMI-1640 medium (Hyclone, Logan, UT, USA)
supplemented with 10% fetal bovine serum (FBS; Gibco by Life
Technologies™, Grand Island, NY, USA), 100 U/ml penicillin and 100
µg/ml streptomycin (GE Healthcare Life Sciences, Logan, UT,
USA) at 37°C in the presence of 5% CO2.
Patient samples
Ten participants were recruited at the Xiangya
Hospital, Central South University (Changsha, Hunan, China).
Consent forms were obtained from individual patients, and
experimental protocols were approved by the Institutional Review
Board of Xiangya Hospital. All subjects enrolled in the study were
Chinese. All clinical and biological data were available for the
samples (Table I). Lung cancer and
corresponding non-tumor normal tissues were collected, and each
biopsy sample was divided into two sections; one was submitted for
routine histological diagnosis, and the remaining section was used
for qPCR, immunohistochemistry and western blot experiments.
 | Table ICharacteristics of the lung cancer
patients. |
Table I
Characteristics of the lung cancer
patients.
| Samples | Age (years) | Gender | Histological
diagnosis |
|---|
| 1 | 58 | Male | Lung squamous cell
cancer |
| 2 | 56 | Male | Lung squamous cell
cancer |
| 3 | 62 | Male | Lung squamous cell
cancer |
| 4 | 58 | Male | Lung squamous cell
cancer |
| 5 | 67 | Male | Lung squamous cell
cancer |
| 6 | 55 | Male | Lung squamous cell
cancer |
| 7 | 65 | Male | Lung squamous cell
cancer |
| 8 | 72 | Female | Lung squamous cell
cancer |
| 9 | 57 | Male | Lung squamous cell
cancer |
| 10 | 56 | Male | Lung squamous cell
cancer |
Total RNA extraction and quantitative
real-time PCR analysis
Total RNA was extracted from the lung cancer and
corresponding non-tumor normal tissues using TRIzol reagent, and
cDNA synthesis was carried out using the RevertAid First Strand
cDNA Synthesis kit (both from CWBio, Beijing, China) according to
the manufacturer's recommendations. Quantitative real-time PCR
(qRT-PCR) was carried out with GoTaq qPCR Master Mix (Promega,
Madison, WI, USA). For detection of Fra-1 mRNA expression levels,
GAPDH was amplified in parallel as an internal control. The
sequences of the primers used for qPCR were as follows: Fra-1
forward, 5′-gcatgggctaaggatttgaa-3′and reverse,
5′-tcccaaatttagcctgttgg-3′; and GAPDH forward, 5′-cga
ccactttgtcaagctca-3′ and reverse, 5′-actgagtgtggcagggactc-3′. The
expression of mRNA was assessed by evaluated threshold cycle (CT)
values. The CT values were normalized with the expression levels of
GAPDH, and the relative amount of mRNA specific to each of the
target genes was calculated using the 2−ΔΔCT method
(17–22). qPCR was carried out with the Bio-Rad
CFK96™ Real-Time system (Bio-Rad, Hercules, CA, USA). The data were
analyzed by Bio-Rad CFK manager software (Bio-Rad). Expression of
mRNA was assessed by the evaluated threshold cycle (CT) values and
GAPDH was used as an internal control.
Immunohistochemistry (IHC) and evaluation
of staining
Immunohistochemistry was conducted using the
peroxidase anti-peroxidase technique following a microwave antigen
retrieval procedure. The antibody for Fra-1 was purchased
from Boster Biotechnology (Wuhan, China). The antibody against
Fra-1 (1:100) was overlaid on lung cancer and corresponding
non-tumor normal tissue sections and incubated overnight at 4°C.
Secondary antibody incubation (Santa Cruz Biotechnology, Inc.,
Santa Cruz, CA, USA) was performed at room temperature for 30 min.
Color reaction was developed by using 3,3′-diaminobenzidine
tetrahydrochloride (DAB) chromogen solution. All slides were
counterstained with hematoxylin. Positive control slides were
included in every experiment in addition to the internal positive
controls. The specificity of the antibody was determined with a
matched IgG isotype antibody as a negative control.
Sections were evaluated in a blinded manner by two
investigators in an effort to provide a consensus on staining
patterns by light microscopy (Olympus). Fra-1 staining was assessed
according to the methods described by Hara and Okayasu (23) with minor modifications. Each case
was rated according to a score that added a scale of intensity of
staining to the area of staining. At least 10 high-power fields
were chosen randomly, and >1,000 cells were counted for each
section. The intensity of staining was graded on the following
scale: 0, no staining; 1+, mild staining; 2+, moderate staining;
3+, intense staining. The area of staining was evaluated as
follows: 0, no staining of cells in any microscopic fields; 1+,
<30% of tissue stained positive; 2+, between 30 and 60% stained
positive; 3+, >60% stained positive. The minimum score when
summed (extension + intensity) was, therefore, 0, and the maximum,
6. A combined staining score (extension + intensity) of ≤2 was
considered to be a negative staining (low staining); a score
between 3 and 4 was considered to be moderate staining; whereas a
score between 5 and 6 was considered to be strong staining. An
optimal cut-off level was identified as follows: a staining index
score of 0–2 was used to define tumors with negative expression and
3–7 indicated positive expression of these two proteins. Agreement
between the two evaluators was 95%, and all scoring discrepancies
were resolved through discussion between the two evaluators.
Construction of the pEGFP-N1-Fra-1
vector
The coding region of the Fra-1 gene was generated by
PCR with the primer pair 5′-atactcgaatgaacctggccatcagcat-3′ and
5′-gcggaattctcacagggacatgaaatccg-3′. PCR was performed under the
following conditions: 1 cycle for 5 min at 94°C; 30 cycles for 45
sec at 94°C, 45 sec at 55°C, and 90 sec at 72°C, and ended with 10
min at 72°C. The fragments were cloned into the TA vector (Promega)
and used to transform E. coli JM109 (Takara, Dalian, China).
Following selection and propagation, the pure plasmid DNA was
prepared by standard methods. The DNA fragments were removed from
the TA vector by restriction enzyme digestion with XhoI and
EcoR1 (Promega) to subclone into the pEGFP-N1 vector. The
fusion sequences were verified by DNA sequencing using ABI
3730.
Cell transfection
Cell transfection was accomplished using
Lipofectamine, according to the manufacturer's instructions
(Invitrogen, USA). Cells (2×105) were plated into each
well of a 6-well plate 24 h prior to the transfection. For each
transfection, 2 µg of pEGFP-N1-Fra-1 plasmid and pEGFP-N1
vector plasmid were transfected into the H460 cells, respectively.
The plasmids were diluted with 100 µl of serum-free media,
and 4 µl Lipofectamine was added into 100 µl
serum-free media. The two solutions were combined, mixed gently and
incubated at room temperature for 30 min. Then 200 µl of the
mixture and 200 µl of serum-free media were added into each
well. The cells were then incubated at 37°C for 24 h, followed by
replacing the transfection media with fresh complete culture media.
After an additional 48 h of culture, the cells were harvested for
the following cell apoptosis, intracellular reactive oxygen species
(ROS) measurement, and western blot experiments.
Effect of Fra-1 on lung cancer cell
apoptosis
Cell apoptosis was analyzed by flow cytometric
analysis using a MoFlo™ XDP High-Performance Cell Sorter (Beckman
Coulter, Brea, CA, USA), propidium iodide (PI) and Hoechst 33342
double staining (Nanjing KeyGen Biotech Co., Ltd., Nanjing, China).
Briefly, H460 cells were transfected transiently with
pEGFP-N1-Fra-1 plasmid and the pEGFP-N1 vector plasmid,
respectively. After 48 h, the cells were collected in an Eppendorf
tube and washed twice with PBS by centrifugation. The supernatants
were discarded. To detect apoptosis, 500 µl PBS, 5 µl
Hoechst 33342 and 5 µl PI were added to each tube, and the
contents of the tube were mixed in the dark at room temperature for
15 min, followed by FCM testing (Beckman Coulter). Data were
acquired and analyzed with Summit v5.2 software (Beckman
Coulter).
Detection of mitochondrial membrane
potential by JC-1
The impact of Fra-1 was measured by flow cytometry
using the sensitive and relatively mitochondrion-specific
lipophilic cationic probe fluorochrome JC-1. JC-1 accumulates to
form J-aggregates and emits red fluorescence in the mitochondria
with a high membrane potential, yet dissociates into monomers and
emits green fluorescence in those that lose cross-membrane
electrochemical gradient. The cells were suspended in 1 ml of warm
staining buffer at ~1×106 cells/ml and incubated at 37°C
for 5 min. Then 1 µl of 2 mM JC-1 was added (2 µM
final concentration) and the cells were incubated at 37°C in 5%
CO2, for 15 to 30 min. The cells were pelleted by
centrifugation, resuspended by gently flicking the tubes, and 500
µl PBS was added to each tube. Cells were analyzed with
MoFlo™ XDP High-Performance Cell Sorter. Data were acquired and
analyzed with Summit v5.2 software.
Intracellular ROS measurement
The production of intracellular ROS was measured by
performing flow cytometry using the oxidation-sensitive probe,
2′,7′-dichlorofluorescein diacetate (DCFH-DA; Applygen, Beijing,
China). Briefly, 10 mM DCFH-DA stock solution (in methanol) was
diluted 4,000-fold in cell culture medium without serum or other
additives to yield a 2.5-mM working solution. After exposure of
human umbilical vein endothelial cells (HUVECs) to silica
nanoparticles for 3 and 24 h, respectively, the cells in 6-well
plates were washed twice with PBS and incubated in 2 ml working
solution of DCFH-DA at 37°C in the dark for 30 min. Then the cells
were washed twice with cold PBS and resuspended in the PBS for
analysis of intracellular ROS by FACS (Beckman Coulter).
Intracellular calcium ion
(Ca2+) concentration assay
Intracellular Ca2+ concentration was
measured by means of the fluorescent Ca2+ chelator
Fura-2 AM, which permeates into cells where it is cut into Fura-2,
resorting within cells. Fura-2 combines with intracellular
Ca2+ to form a fluorescent compound, whose fluorescent
intensity was determined at an excitation wavelength of 340 nm and
an emission wavelength of 510 nm in FACS (Beckman Coulter). After
treatment, the cells were harvested and rinsed with PBS. The
harvested cells were suspended in PBS and incubated with 5
µM Fura-2 AM for 60 min at 37°C. During the session of
incubation with Fura-2 AM, the cell cultures were mildly shaken at
intervals of 10 min aimed to facilitate the combination of Fura-2
and Ca2+ to form the fluorescent compound. Then, the
cells were washed twice and resuspended in PBS for FACS measurement
(Beckman Coulter). Data were acquired and analyzed with Summit v5.2
software.
Western blot analysis
The lung cancer tissues, corresponding non-tumor
normal tissues and H460 cells were lysed in RIPA buffer (CWBio),
and total protein concentration was determined using the
Pierce® BCA protein assay kit (Thermo Scientific, Inc.,
Rockford, IL, USA). Extracts containing 50 µg of proteins
were separated on 10% SDS-PAGE gels and electroblotted onto
nitrocellulose membranes (Hyclone Laboratories). The membranes were
inhibited using Tris-buffered saline/Tween-20 (25 mM Tris-HCl, 150
mM NaCl, pH 7.5 and 0.05% Tween-20) containing 5% non-fat milk
followed by overnight incubation at 4°C with the primary antibodies
(rabbit anti-MDM2 antibody, 1:200 and rabbit anti-P53 antibody,
1:200; Boster Biotechnology). Following three washes, the membranes
were incubated with horseradish peroxidase-conjugated secondary
antibodies (catalog no. sc-2491, dilution 1:5,000; Santa Cruz
Biotechnology, Inc.) and the specific signals were visualized using
an ECL detection system. The anti-GAPDH antibody (1:3,000; Santa
Cruz Biotechnology, Inc.) was used as a loading control.
Statistical analysis
Differences in non-parametric variables were
analyzed by the Mann-Whitney U test. Differences in the
quantitative variables between groups were analyzed by Student's
t-test using SPSS 11.0 program (SPSS, Chicago, IL, USA). A
probability value (p) <0.05 was considered to indicate a
statistically significant result.
Results
Fra-1 expression is upregulated in lung
cancer tissues
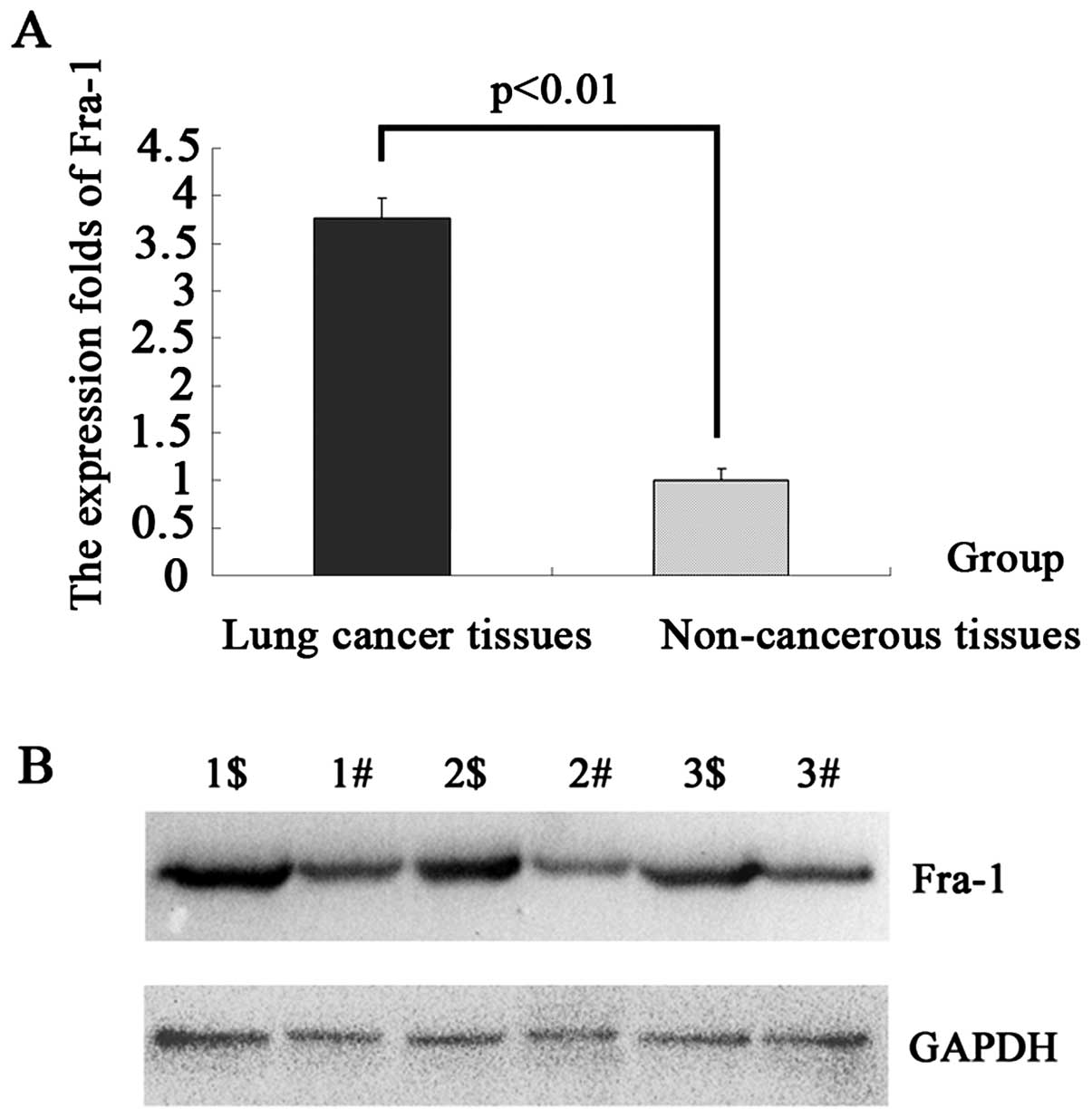
To detect the mRNA expression levels of the Fra-1
molecule in lung cancer and the adjacent non-cancerous tissues, 10
samples of each were selected to perform qPCR of the Fra-1 gene.
The data were analyzed using the 2−ΔΔCT method, and the
fold change in the expression of this gene relative to the internal
control gene, GAPDH, was analyzed. The expression of the Fra-1 gene
was higher in the lung cancer samples compared with the adjacent
non-cancerous tissues and the normalized Fra-1 gene expression in
lung cancer was upregulated by 3.76-fold (p=0.0094<0.01)
(Table II, Fig. 1A).
| Table IIAssessment of the mRNA expression
levels of Fra-1 in lung cancer and adjacent non-cancerous tissues
by qPCR. |
Table II
Assessment of the mRNA expression
levels of Fra-1 in lung cancer and adjacent non-cancerous tissues
by qPCR.
| Gene | Sample | n | Fra-1 CT (mean ±
SD) | GAPDH CT (mean ±
SD) | ΔCT (mean ±
SD) | ΔΔCT (mean ±
SD) | Folda |
|---|
| Fra-1 | Lung cancer | 10 | 26.15±1.14 | 19.24±0.64 | 6.91±0.43 | −1.91±0.42 | 3.76 |
| Non-cancerous
tissues | 10 | 28.23±1.17 | 19.41±0.73 | 8.82±0.39 | | (2.62–4.99) |
To determine whether the Fra-1 gene is expressed at
a higher level in lung cancer compared with adjacent non-cancerous
tissues, the protein expression levels of Fra-1 were further
examined by western blot analysis in 1 to 3 samples (Fig. 1B). In comparison with the adjacent
non-cancerous tissues, the expression level was identified to be
higher in the lung cancer tissues, which corresponded with the qPCR
results.
To confirm the pattern of Fra-1 in lung cancer,
immunohistochemistry (IHC) was carried out with antibodies against
the Fra-1 protein in lung cancer and the adjacent non-cancerous
tissues. Fra-1 was identified as being differentially expressed
between lung cancer tissues vs. the adjacent non-cancerous tissues.
IHC showed a similar pattern in protein expression with the western
blot results. A total of 50.09% (13/22) of lung cancer tissues had
high Fra-1 expression and 13.64% (3/22) of the adjacent
non-cancerous tissues had a high score. The distribution of low
scores was 9.09% (2/22) and 63.64% (14/22) in the lung cancer and
the adjacent non-cancerous tissues, respectively (p=0.012<0.05)
(Table III). This corresponded
with the qPCR results.
| Table IIIDifference in Fra-1 expression
between the lung cancer and the adjacent non cancerous tissues. |
Table III
Difference in Fra-1 expression
between the lung cancer and the adjacent non cancerous tissues.
| n | Score
| P-value |
|---|
| Low (0–2) | Moderate (3–4) | High (5–6) |
|---|
| Lung cancer | 22 | 2 (9.09%) | 7 (31.82%) | 13 (50.09%) | =0.012<0.05 |
| Non-cancerous
tissues | 22 | 14 (63.64%) | 5 (22.73%) | 3 (13.64%) | |
Fra-1 inhibits the apoptosis of lung
cancer cells in vitro
We found that Fra-1 was highly expressed in the lung
cancer tissues by qPCR, western blot and IHC technologies. To
elucidate the function of Fra-1 in the apoptosis of lung cancer
cells, the lung cancer cell line, H460, was transfected with the
plasmid pEGFP-N1/Fra-1 or the control vector. After determining
Fra-1 protein by qPCR technologies, we performed a Hoechst 33342/PI
double-staining experiment to test the rate of apoptosis in the
H460, H460/vector, and H460/Fra-1 cells. A considerable decrease in
apoptotic cells was observed in the H460/Fra-1 cells (6.29±0.41%)
compared with the H460/vector (11.73±0.85%) and H460 cells
(12.04±0.92%) (Fig. 2A).
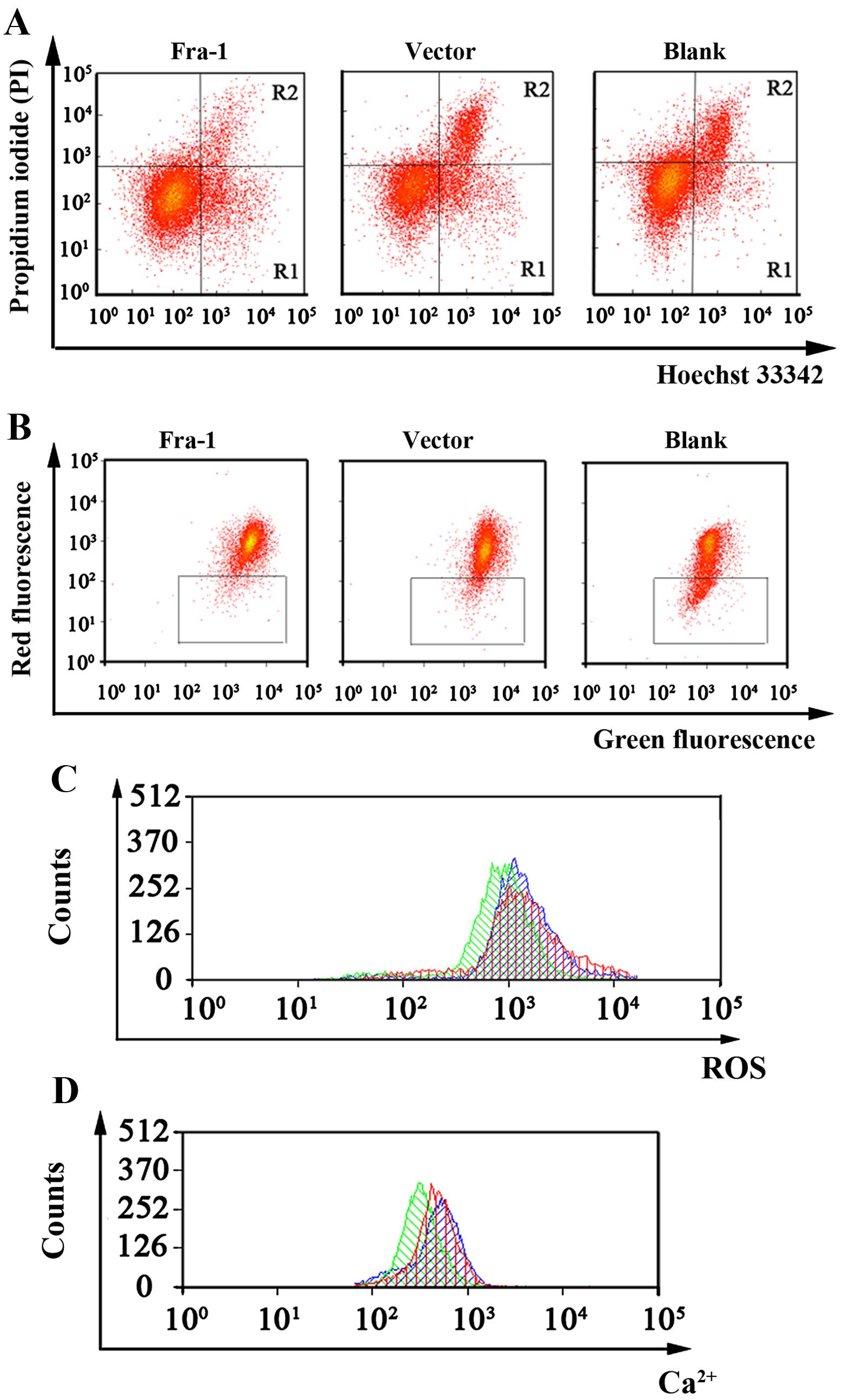 | Figure 2Assessment of cell apoptosis,
mitochondrial membrane potential (ΔΨm), intracellular reactive
oxygen species (ROS) and calcium ion Ca2+ concentration
in H460, H460/vector and H460/Fra-1 cells. (A) Cell apoptosis of
the H460, H460/vector and H460/Fra-1 cells was analyzed by flow
cytometry. (B) ΔΨm in the H460, H460/vector and H460/Fra-1 cells
was analyzed by flow cytometry. (C) ROS in the H460, H460/vector
and H460/Fra-1 cells were analyzed by flow cytometry. Green
indicates H460/Fra-1 cells, red indicates H460/vector cells, blue
indicates H460 cells. (D) Ca2+ concentrations in the
H460, H460/vector and H460/Fra-1 cells were assessed by flow
cytometry. Green indicates H460/Fra-1 cells, red indicates
H460/vector cells, blue indicates H460 cells. Data are
representative of three independent experiments. |
Fra-1 affects mitochondrial membrane
potential (ΔΨm), ROS and Ca2+ concentrations in lung
cancer cells in vitro
Cell apoptosis is closely related with a reduction
in ΔΨm and an increase in intracellular ROS and Ca2+
concentrations. Thus, we tested the influence of Fra-1 on these
three parameters. Our results showed that overexpression of Fra-1
in lung cancer H460 cells, led to an increase in ΔΨm and inhibited
cell apoptosis (Fig. 2B). ROS
results showed that overexpression of Fra-1 in lung cancer H460
cells, led to a decrease in intracellular ROS (Fig. 2C). Ca2+ experimental
results showed that overexpression of Fra-1 in lung cancer H460
cells, led to a decrease in Ca2+ concentrations and
suppressed the apoptosis of the lung cancer cells (Fig. 2D).
Fra-1 is correlated with dysregulation of
the P53 signaling pathway in lung cancer tissues in vivo
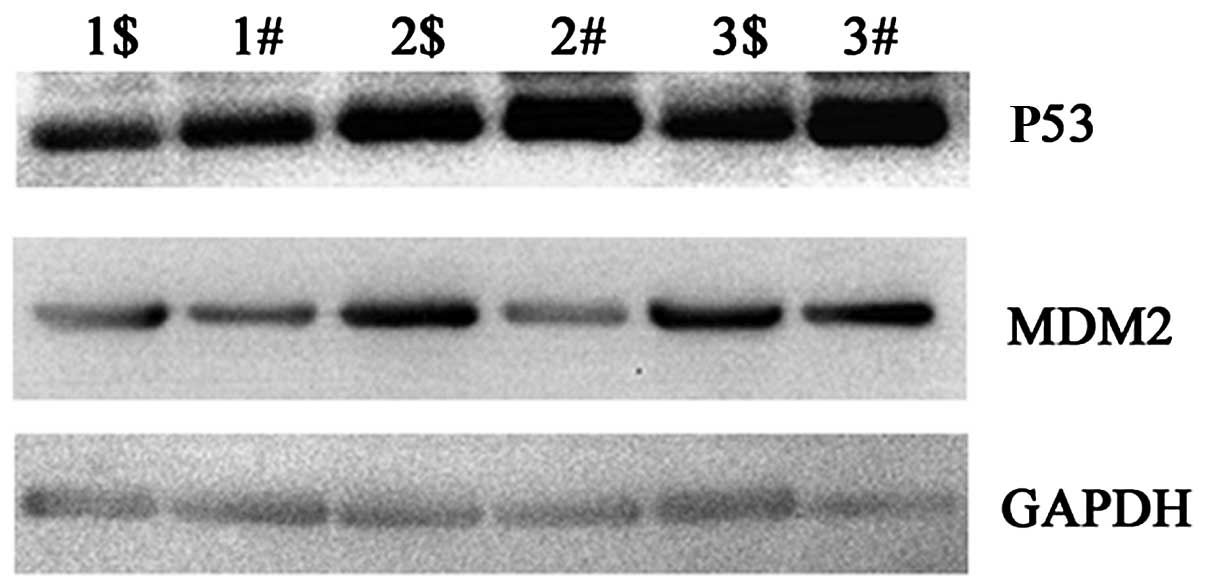
To uncover the possible mechanism of Fra-1 in lung
cancer, we tested the expression levels of key molecules in the P53
signaling pathway by western blot technology. P53 was downregulated
in the lung cancer tissues compared with the level in the adjacent
non-cancerous tissues, while MDM2 was upregulated in the lung
cancer tissues (Fig. 3). Combined
with the above result for which Fra-1 was upregulated in lung
cancer, we propose that Fra-1 is correlated with dysregulation of
the P53 signaling pathway in lung cancer tissues in
vitro.
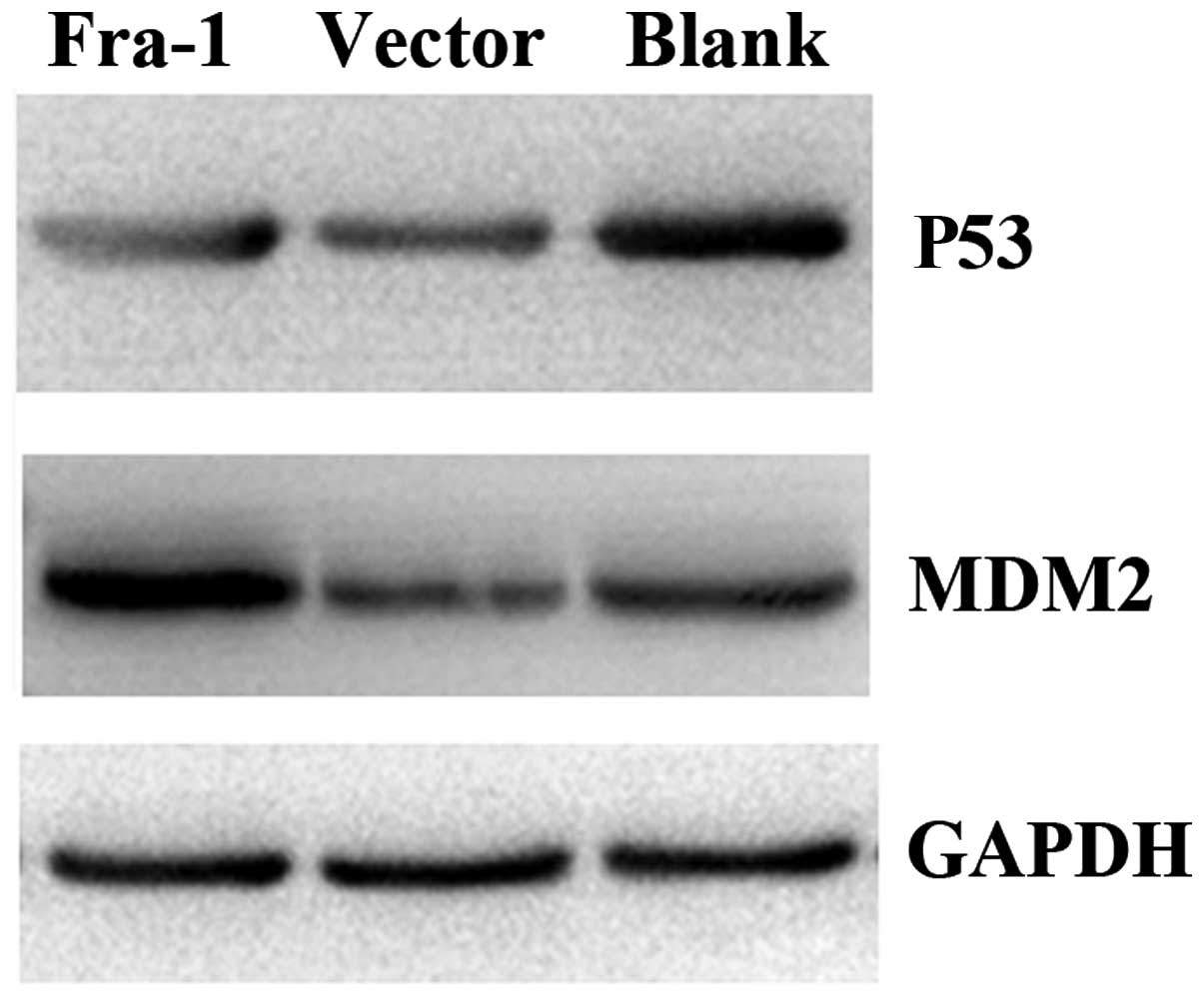
Fra-1 overexpression affects the
expression of P53 and MDM2 in vitro
To confirm whether Fra-1 affects the expression of
P53 and MDM2 in vivo, the H460 cells were transfected with
the plasmid pEGFP-N1/Fra-1 or control vector. We harvested the
cells and tested the expression levels of P53 and MDM2 proteins
in vivo. P53 was downregulated in the H460 cells in which
Fra-1 was overexpressed, while MDM2 was upregulated (Fig. 4). Our results revealed that Fra-1
overexpression affected the expression of P53 and MDM2 in
vivo.
Discussion
Lung cancer is the leading cause of cancer-related
death worldwide with a 5-year survival rate of <15%, despite
significant advances in both diagnostic and therapeutic approaches
(1,2). Lung cancer occurs and develops as a
complicated result of an accumulation of various endogenous and
exogenous effects. Gene alterations participate in lung cancer
genesis (24–26). Thus, identifying molecular
aberrations in lung cancer may improve our understanding of lung
carcinogenesis and help us to subdivide patients into biologically
and clinically relevant subgroups, as well as develop novel
therapeutic strategies.
In this study, we found that the expression of the
Fra-1 gene was higher in the lung cancer samples compared with that
in the adjacent non-cancerous tissues and the normalized Fra-1 gene
expression in lung cancer was upregulated by 3.76-fold. A similar
trend was noted by western blot experiments. IHC showed a similar
pattern in protein expression with the qPCR results and western
blot results. A high score of Fra-1 was noted in 50.09% (13/22) of
the lung cancer tissues and 13.64% (3/22) of the adjacent
non-cancerous tissues. Motrich et al found that both Fra-1
and c-Fos were overexpressed in >95% of human ductal breast
carcinoma biopsies examined in contrast with very low or
undetectable levels in normal tissue (27). Fra-1 is highly expressed in the
muscle-invasive form of carcinoma of the bladder (28). Fos-related activator-1 is
overexpressed in oral squamous cell carcinoma and is associated
with tumor lymph node metastasis (29). Our results corresponded with the
previous findings.
To uncover the potential mechanism of Fra-1 in lung
cancer, we studied the effect of Fra-1 on the apoptosis of lung
cancer H460 cells. Our results showed that there was a considerable
decrease in apoptotic cells in the H460 cells with Fra-1
overexpession. Henckels and Prywes found that Fra-1 overexpression
was sufficient to increase cell motility and anchorage-independent
growth (30). High levels of Fra-1
in cells where elevated ERK activity leads to protein stabilization
provide survival signals for tumor cells removed from the
extracellular matrix (31). Fra-1
may influence the metastatic potential of breast cancer cells by
altering the expression of adhesion molecules, resulting in
increased adherence to endothelial cells under flow conditions
(32). Collectively, the results
suggested that Fra-1 may affect cell proliferation, invasion and
apoptosis.
Cell apoptosis is closely related with a reduction
in ΔΨm and an increase in intracellular ROS and Ca2+
concentrations (33–38). In this study, we found that
overexpression of Fra-1 in lung cancer H460 cells, led to an
increase in ΔΨm and a decrease in intracellular ROS and
Ca2+ concentrations. Yang and Meyskens found that
alterations in AP-1 transcription signaling, mediated by changes in
AP-1 dimeric composition and reduced intracellular ROS levels,
substantially contributed to the phenotypic changes induced by
resveratrol (39). The rise in
intracellular calcium concentration (Ca2+) causes
mitochondrial Ca2+ overload, thereby triggering
apoptosis (40).
Furthermore, we found that Fra-1 was correlated with
dysregulation of the P53 signaling pathway in lung cancer tissues
in vitro, and Fra-1 overexpression affected the expression
of MDM2 and P53 in vivo. Degradation of P53 is regulated by
its interaction with specific E3 ubiquitin ligases, the best known
one being encoded by MDM2 (41).
Damage to the P53-dependent mechanism is often caused by
overexpression of MDM2, which codes for a P53-regulating protein
(42). Combined with the above
results revealing that Fra-1 is highly expressed in lung cancer, we
inferred that Fra-1 is upregulated in lung cancer tissues and
inhibits the apoptosis of lung cancer cells by the P53 signaling
pathway.
Acknowledgments
This study was supported by the National Natural
Science Foundation of China (81272975, 81172302, 81402270 and
81402307); Key Project of Hunan Provincial Natural Science
Foundation (12JJ2044); Project of the Hunan Provincial Natural
Science Foundation (12JJ3121); Project of the Hunan Provincial
Development and Reform Commission; the Planned Science and
Technology Project of Hunan Province (2010FJ3088 and 2012FJ2014);
and Hunan Provincial Innovation Foundation for Postgraduates.
Abbreviations:
|
Fra-1
|
Fos-related antigen-1
|
|
AP-1
|
activator protein-1
|
|
TP53
|
tumor protein p53
|
|
ROS
|
reactive oxygen species
|
|
DAB
|
3,3′-diaminobenzidine
tetrahydrochloride
|
|
FBS
|
fetal bovine serum
|
|
PI
|
propidium iodide
|
|
DCFH-DA
|
2′,7′-dichlorofluorescein
diacetate
|
|
IHC
|
immunohistochemistry
|
|
GADPH
|
glyceraldehyde-3-phosphate
dehydrogenase
|
References
|
1
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wood SL, Pernemalm M, Crosbie PA and
Whetton AD: Molecular histology of lung cancer: From targets to
treatments. Cancer Treat Rev. 41:361–375. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang F, Zhang X, Meng J, Zhao Y, Liu X,
Liu Y, Wang Y, Li Y, Sun Y, Wang Z, et al: ING5 inhibits cancer
aggressiveness via preventing EMT and is a potential prognostic
biomarker for lung cancer. Oncotarget. 6:16239–16252. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen X, Song X, Yue W, Chen D, Yu J, Yao Z
and Zhang L: Fibulin-5 inhibits Wnt/β-catenin signaling in lung
cancer. Oncotarget. 6:15022–15034. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Adiseshaiah P, Vaz M, Machireddy N,
Kalvakolanu DV and Reddy SP: A Fra-1-dependent, matrix
metalloproteinase driven EGFR activation promotes human lung
epithelial cell motility and invasion. J Cell Physiol. 216:405–412.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rege TA and Hagood JS: Thy-1 as a
regulator of cell-cell and cell-matrix interactions in axon
regeneration, apoptosis, adhesion, migration, cancer, and fibrosis.
FASEB J. 20:1045–1054. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Scognamiglio G, D'Antonio A, Rossi G,
Cavazza A, Camerlingo R, Pirozzi G, La Mantia E, Anniciello AM,
Morabito A, Cantile M, et al: CD90 expression in atypical
meningiomas and meningioma metastasis. Am J Clin Pathol.
141:841–849. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kitayama J, Emoto S, Yamaguchi H, Ishigami
H, Yamashita H, Seto Y, Matsuzaki K and Watanabe T: CD90(+)CD45(−)
intraperitoneal mesothelial-like cells inhibit T cell activation by
production of arginase I. Cell Immunol. 288:8–14. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ohmura-Kakutani H, Akiyama K, Maishi N,
Ohga N, Hida Y, Kawamoto T, Iida J, Shindoh M, Tsuchiya K,
Shinohara N, et al: Identification of tumor endothelial cells with
high aldehyde dehydrogenase activity and a highly angiogenic
phenotype. PLoS One. 9:e1139102014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ma K, Chang D, Gong M, Ding F, Luo A, Tian
F, Liu Z and Wang T: Expression and significance of FRA-1 in
non-small-cell lung cancer. Cancer Invest. 27:353–359. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Adiseshaiah P, Lindner DJ, Kalvakolanu DV
and Reddy SP: FRA-1 proto-oncogene induces lung epithelial cell
invasion and anchorage-independent growth in vitro, but is
insufficient to promote tumor growth in vivo. Cancer Res.
67:6204–6211. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Oikonomou D, Hassan K, Kaifi JT, Fiegel
HC, Schurr PG, Reichelt U, Aridome K, Yekebas EF, Mann O, Kluth D,
et al: Thy-1 as a potential novel diagnostic marker for
gastrointestinal stromal tumors. J Cancer Res Clin Oncol.
133:951–955. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jiang J, Zhang Y, Chuai S, Wang Z, Zheng
D, Xu F, Zhang Y, Li C, Liang Y and Chen Z: Trastuzumab (herceptin)
targets gastric cancer stem cells characterized by CD90 phenotype.
Oncogene. 31:671–682. 2012. View Article : Google Scholar
|
|
14
|
Goldar S, Khaniani MS, Derakhshan SM and
Baradaran B: Molecular mechanisms of apoptosis and roles in cancer
development and treatment. Asian Pac J Cancer Prev. 16:2129–2144.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Paul I and Jones JM: Apoptosis block as a
barrier to effective therapy in non small cell lung cancer. World J
Clin Oncol. 5:588–594. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Beesoo R, Neergheen-Bhujun V, Bhagooli R
and Bahorun T: Apoptosis inducing lead compounds isolated from
marine organisms of potential relevance in cancer treatment. Mutat
Res. 768:84–97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
18
|
Zhou Y, Wang W, Zheng D, Peng S, Xiong W,
Ma J, Zeng Z, Wu M, Zhou M, Xiang J, et al: Risk of nasopharyngeal
carcinoma associated with polymorphic lactotransferrin haplotypes.
Med Oncol. 29:1456–1462. 2012. View Article : Google Scholar
|
|
19
|
Xiao S, Zhou Y, Yi W, Luo G, Jiang B, Tian
Q, Li Y and Xue M: Fra-1 is downregulated in cervical cancer
tissues and promotes cervical cancer cell apoptosis by p53
signaling pathway in vitro. Int J Oncol. 46:1677–1684.
2015.PubMed/NCBI
|
|
20
|
Zheng D, Liao S, Zhu G, Luo G, Xiao S, He
J, Pei Z, Li G and Zhou Y: CD38 is a putative functional marker for
side population cells in human nasopharyngeal carcinoma cell lines.
Mol Carcinog. Jan 28–2015.Epub ahead of print. View Article : Google Scholar
|
|
21
|
Liao S, Xiao S, Zhu G, Zheng D, He J, Pei
Z, Li G and Zhou Y: CD38 is highly expressed and affects the
PI3K/Akt signaling pathway in cervical cancer. Oncol Rep.
32:2703–2709. 2014.PubMed/NCBI
|
|
22
|
Zhu W, Li J, Su J, Li J, Li J, Deng B, Shi
Q, Zhou Y and Chen X: FOS-like antigen 1 is highly expressed in
human psoriasis tissues and promotes the growth of HaCaT cells in
vitro. Mol Med Rep. 10:2489–2494. 2014.PubMed/NCBI
|
|
23
|
Hara A and Okayasu I: Cyclooxygenase-2 and
inducible nitric oxide synthase expression in human astrocytic
gliomas: Correlation with angiogenesis and prognostic significance.
Acta Neuropathol. 108:43–48. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Guo M, Alumkal J, Drachova T, Gao D,
Marina SS, Jen J and Herman JG: CHFR methylation strongly
correlates with methylation of DNA damage repair and apoptotic
pathway genes in non-small cell lung cancer. Discov Med.
19:151–158. 2015.PubMed/NCBI
|
|
25
|
Chiu CH, Ho HL, Doong H, Yeh YC, Chen MY,
Chou TY and Tsai CM: MLH1 V384D polymorphism associates with poor
response to EGFR tyrosine kinase inhibitors in patients with EGFR
L858R-positive lung adenocarcinoma. Oncotarget. 6:8407–8417. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Van Nostrand JL, Brisac A, Mello SS,
Jacobs SB, Luong R and Attardi LD: The p53 target gene SIVA enables
non-small cell lung cancer development. Cancer Discov. 5:622–635.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Motrich RD, Castro GM and Caputto BL: Old
players with a newly defined function: Fra-1 and c-Fos support
growth of human malignant breast tumors by activating membrane
biogenesis at the cytoplasm. PLoS One. 8:e532112013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sayan AE, Stanford R, Vickery R,
Grigorenko E, Diesch J, Kulbicki K, Edwards R, Pal R, Greaves P,
Jariel-Encontre I, et al: Fra-1 controls motility of bladder cancer
cells via transcriptional upregulation of the receptor tyrosine
kinase AXL. Oncogene. 31:1493–1503. 2012. View Article : Google Scholar
|
|
29
|
Zhang L, Pan HY, Zhong LP, Wei KJ, Yang X,
Li J, Shen GF and Zhang Z: Fos-related activator-1 is overexpressed
in oral squamous cell carcinoma and associated with tumor lymph
node metastasis. J Oral Pathol Med. 39:470–476. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Henckels E and Prywes R: Fra-1 regulation
of matrix metallopeptidase-1 (MMP-1) in metastatic variants of
MDA-MB-231 breast cancer cells. F1000Res. 2:2292013.PubMed/NCBI
|
|
31
|
Vial E and Marshall CJ: Elevated ERK-MAP
kinase activity protects the FOS family member FRA-1 against
proteasomal degradation in colon carcinoma cells. J Cell Sci.
116:4957–4963. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Oliveira-Ferrer L, Kürschner M, Labitzky
V, Wicklein D, Müller V, Lüers G, Schumacher U, Milde-Langosch K
and Schröder C: Prognostic impact of transcription factor Fra-1 in
ER-positive breast cancer: Contribution to a metastatic phenotype
through modulation of tumor cell adhesive properties. J Cancer Res
Clin Oncol. 141:1715–1726. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hacker K and Medler KF: Mitochondrial
calcium buffering contributes to the maintenance of Basal calcium
levels in mouse taste cells. J Neurophysiol. 100:2177–2191. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ogura T, Margolskee RF and Kinnamon SC:
Taste receptor cell responses to the bitter stimulus denatonium
involve Ca2+ influx via store-operated channels. J
Neurophysiol. 87:3152–3155. 2002.PubMed/NCBI
|
|
35
|
Akabas M, Dodd J and al-Awqati Q:
Identification of electrophysiologically distinct subpopulations of
rat taste cells. J Membr Biol. 114:71–78. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Pulkkinen V, Manson ML, Säfholm J, Adner M
and Dahlén SE: The bitter taste receptor (TAS2R) agonists
denatonium and chloroquine display distinct patterns of relaxation
of the guinea pig trachea. Am J Physiol Lung Cell Mol Physiol.
303:L956–L966. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ruiz-Avila L, McLaughlin SK, Wildman D,
McKinnon PJ, Robichon A, Spickofsky N and Margolskee RF: Coupling
of bitter receptor to phosphodiesterase through transducin in taste
receptor cells. Nature. 376:80–85. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li Y, Su X, Wang X, Leung AW, Xu C, Wang P
and Liu Q: Cytotoxic effect of protoporphyrin IX to human leukemia
U937 cells under ultrasonic irradiation. Cell Physiol Biochem.
33:1186–1196. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yang S and Meyskens FL Jr: Alterations in
activating protein 1 composition correlate with phenotypic
differentiation changes induced by resveratrol in human melanoma.
Mol Pharmacol. 67:298–308. 2005. View Article : Google Scholar
|
|
40
|
Liu KH, Yang ST, Lin YK, Lin JW, Lee YH,
Wang JY, Hu CJ, Lin EY, Chen SM, Then CK, et al: Fluoxetine, an
antidepressant, suppresses glioblastoma by evoking AMPAR-mediated
calcium-dependent apoptosis. Oncotarget. 6:5088–5101. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chumakov PM: Function of the p53 gene:
Choice between life and death. Biochemistry (Mosc). 65:28–40.
2000.
|
|
42
|
Chipuk JE and Green DR: Dissecting
p53-dependent apoptosis. Cell Death Differ. 13:994–1002. 2006.
View Article : Google Scholar : PubMed/NCBI
|