Introduction
Lung cancer is the leading cause of cancer death
among various types of malignant tumours, with the highest
morbidity and mortality rates worldwide (1). According to tumour histologic
features, lung cancer can be divided into several subtypes, of
which non-small cell lung cancer (NSCLC) occupies >80% of the
total lung cancer cases (2).
Although current therapies have achieved certain progress, the
disease still shows poor outcomes and low five-year survival rates
(<15%) (3,4). Therefore, elucidating the pathological
mechanism to provide a theoretical basis for developing potential
and effective therapies for NSCLC is imperative.
In recent years, microRNAs (miRNAs), a class of
18–22 nucleotide-long small non-coding RNAs, have been recognised
as novel regulators for gene expression; miRNAs
post-transcriptionally modulate gene expression by targeting the
3′-untranslated region (UTR) of the targets (5,6). Thus,
miRNAs regulate various cellular processes and participate in the
pathogenesis of many diseases (7,8).
Certain miRNAs are dysregulated in almost all cancer types; these
miRNAs do not only contribute to the development and progression of
cancers, but also represent novel targets for cancer therapy
(9–11). Therefore, miRNAs can serve as novel
diagnostic biomarkers and potential candidate therapeutic
targets.
Nucleosome-binding protein 1 (NSBP1) (also named
high-mobility group nucleosome-binding domain 5) has been widely
studied because of its ability to modulate gene transcription by
binding to chromatin (12). NSBP1
is broadly distributed in many tissues and is mainly located in the
nucleus (13,14). NSBP1 is currently recognised as a
candidate oncogene in various cancers, including prostate cancer
(15), gliomas (16), bladder cancer (17), breast cancer (18) and osteosarcoma (19). In addition, silencing of NSBP1
suppresses lung cancer cell proliferation (20). However, the molecular basis of NSBP1
in regulating lung cancer remains unclear. Nevertheless, NSBP1 may
be a potential candidate target for developing cancer therapies,
including lung cancer.
miRNAs are regulators for gene silencing,
representing novel therapeutic targets. However, to date, the
specific miRNAs that can target and regulate NSBP1 have not been
recognised. Thus, the present study aimed to identify a novel miRNA
that specifically targets and modulates NSBP1 expression in NSCLC.
Bioinformatics analysis revealed that miR-326 had a putative
binding site within the 3′-UTR of NSBP1. Their substantial
relationship was further verified by dual-luciferase reporter
assay, real-time quantitative polymerase chain reaction (RT-qPCR)
and western blot analysis. As expected, suppression of NSBP1 by
miR-326 overexpression mimicked the effect of NSBP1 silencing by
small interfering RNA (siRNA) on NSCLC cell proliferation and
invasion. Suppression of NSBP by miR-326 overexpression or NSBP1
siRNA both inhibited the expression levels of cyclin B1 and MMP9,
which are associated with tumour cell proliferation and invasion.
Further data indicated that miR-326 and NSBP1 expression levels
were inversely correlated in NSCLC tissues. Taken together, our
study suggested a novel and critical functional significance of
miR-326 in NSCLC, and this miRNA was a promising therapeutic
candidate for NSCLC by suppressing NSBP1.
Materials and methods
Cell cultures
Human NSCLC cell lines NCI-H157 and A549 were
obtained from American Type Culture Collection (Manassas, VA, USA).
Human keratinocyte cell line HaCaT and human embryonic kidney cell
line HEK293T were purchased from Type Culture Collection of the
Chinese Academy of Sciences (Shanghai, China). These cells were
maintained in DMEM (Invitrogen, Carlsbad, CA, USA) plus 10% fetal
calf serum (FCS; Gibco, Rockville, MD, USA) and 1%
penicillin/streptomycin (Sigma, St. Louis, MO, USA) and maintained
in a humidified incubator containing 95% air/5% CO2 at
37°C.
RT-qPCR
Total RNA was extracted using miRNeasy mini kit
(Qiagen, Dusseldorf, Germany) according to the manufacturer's
instructions. For mRNA expression analysis, cDNA was reverse
transcribed by M-MLV reverse transcriptase (Takara, Dalian,
Shanghai). For miRNA expression analysis, cDNA was synthesised with
miScript reverse transcription kit (Qiagen). Primer sequences were
as follows: NSBP1, forward 5′-GCAGTCAGGCAGTGACTGCCTTCG-3′ and
reverse 5′-CCCTTTTCTGTGGCATCTTC-3′; GAPDH, forward
5′-CAGTCAGCCGCATCTTCTTTT-3′ and reverse 5′-GTGACCAGGCGCCCAATAC-3′;
miR-326, forward 5′-ACTGTCCTTCCCTCTGGGC-3′ and reverse
5′-AATGGTTGTTCTCCACTCTCTCTC-3′; U6 small nuclear RNA, forward
5′-GCTTCGGCAGCACATATACTAAAAT-3′ and reverse,
5′-CGCTTCACGAATTTGCGTGTCAT-3′. GAPDH was used as an internal
control for mRNA quantification. U6 small nuclear RNA was used as
an internal control for miRNA quantification. RT-qPCR analysis was
performed using SYBR-Green Master Mix (Bio-Rad, Hercules, CA, USA).
The relative gene expression was analysed using the
2−ΔΔCt method and normalised to the internal
controls.
Western blot analysis
Cells were lysed in RIPA lysis buffer, and the
protein concentration was measured using a BCA kit (Beyotime
Biotechnology, Haimen, China). Equal amounts of proteins from
different samples were separated on 12.5% sodium dodecyl sulphate
(SDS) polyacrylamide gel electrophoresis. The separated protein
bands on SDS polyacrylamide gel were electro-transferred onto a
nitrocellulose membrane (Bio-Rad). After blocking with 5% skim milk
powder, the membrane was blotted with NSBP1, cyclin B1, MMP9 and
GAPDH antibodies (Cruz Biotechnology, Inc., Santa Cruz, CA, USA) at
4°C overnight. Horseradish peroxidase-conjugated secondary
antibodies (Santa Cruz Biotechnology, Inc.) diluted in 1:2,000 were
added and incubated for 1 h at room temperature. Finally, the
immunoreactive bands were developed with the enhanced
chemiluminescence method. The grey value of protein bands was
quantified with Image-Pro Plus 6.0 software (Media Cybernetics,
Inc., Rockville, MD, USA), and relative protein expression data
were normalised to GAPDH.
Cell transfection
NSBP1-specific siRNA and control non-specific siRNA
(NC siRNA) were purchased from Cruz Biotechnology, Inc., and then
transfected into cells according to the manufacturer's
instructions. In brief, cells were plated into six-well plates at
2×105 cells/well overnight. About 1 μg of siRNA
was diluted into 100 μl of transfection medium, and 6
μl of transfection reagent was diluted into 100 μl of
transfection medium. Both solutions were then mixed together for 45
min at room temperature. The siRNA transfection reagent mixture was
added to each well with 0.8 ml of transfection medium and incubated
for 7 h. The medium was replaced with fresh normal growth medium
and cultured for another 48 h. The interfering efficiency was
detected by western blot analysis. miR-326 mimics and non-specific
controls (miR-NC) were obtained from Shanghai GenePharma Co., Ltd.
(Shanghai, China). NSCLC cells were seeded in six-well plates
overnight and then transfected with 50 nM miR-326 mimics or miR-326
inhibitor using Lipofectamine 2000 (Invitrogen) according to the
manufacturer's instruction. The transfection efficiency was
subsequently detected by RT-qPCR analysis after transfection for 48
h.
Cell proliferation assay
Cell proliferation of NSCLC cells was measured by
the 3-(4,5-dimethyl-2-thiazolyl)-2,5-di-phenyl-2-H-tetrazolium
bromide (MTT) method. In brief, NSCLC cells were seeded into
96-well cell plates at a density of 1×105 cells/well and
grown for 24 h. The cells were then transfected with miR-NC or
miR-326 mimics for 48 h. The old medium was replaced by fresh
medium containing 20 μl of MTT (5 mg/ml in PBS; Sigma) and
cultured for 4 h. The medium was discarded, and 200 μl of
dimethyl sulphoxide was added to dissolve crystal formazan. The
optical density (OD) of each reaction solution at 490 nm was
measured with an enzyme immunoassay analyser (Bio-Tek Instruments,
Winooski, VT, USA).
Cell invasion assay
Tumour cell invasion ability was detected by
Transwell invasion assay. The Transwell filter was precoated with
Matrigel (BD Bioscience, San Jose, CA, USA) on the upper surface of
a polycarbonic membrane. NSCLC cells were transfected with miR-326
mimics or miR-NC for 48 h and then starved overnight. A total of
1×105 cells were added in the top chamber with
serum-free medium, and the medium containing 10% FCS was added to
the lower chamber. After 48 h, non-migrated cells in the upper
chamber were removed by a cotton swab, and the migrated cells in
the lower chamber were fixed with 95% ethanol and stained with 4
g/l trypan blue solution. Cells were observed under a microscope
(Olympus, Tokyo, Japan), and positive staining cells were counted
in five random fields.
Cell cycle analysis
Cell cycle distribution was detected by flow
cytometry. In brief, NSCLC cells were serum-starved for 24 h and
transfected with NSBP1 siRNA or miR-326 mimics for 48 h.
Thereafter, cells were harvested, washed with ice-cold PBS and
fixed with 70% ethanol. Propidium iodide containing RNase
(Molecular Probes, Eugene, OR, USA) was added to the cells and
incubated for 30 min in the dark. FACScan flow cytometry (Becton
Dickinson, Franklin Lakes, NJ, USA) was used to detect the cell
cycle distribution.
Dual-luciferase reporter assay
The cDNA fragments from NSBP1 containing an miR-326
binding site were inserted into pmirGLO vector (Promega, Madison,
WI, USA). HEK293T cells were seeded into six-well plates at
2×105 cells/well overnight. About 10 ng of pmirGLO-NSBP1
recombinant vectors and 50 nM miR-326 mimics were introduced into
HEK293T cells using Lipofectamine 2000 (Invitrogen). The
transfected cells were lysed after 48 h of transfection. Firefly
luciferase activity and Renilla luciferase activity were quantified
by the dual-luciferase reporter method (Promega).
NSCLC specimen collection
Twenty pairs of NSCLC tissues and matched adjacent
non-tumour tissues were provided by Xi'an Central Hospital of Xi'an
Jiaotong University. The tissues were collected from NSCLC patients
who had undergone surgical resections and stored at −80°C for use.
The use of clinical tissues was approved by the hospital's
Institutional Human Experiment and Ethic Committee with informed
consent from the patients.
Statistical analysis
Data are presented as the mean ± standard deviation
(SD). Statistical analyses were carried out using SPSS software
(version 11.5; SPSS Inc., Chicago, IL, USA), and statistical
differences were analysed using Student's t-test or one-way ANOVA.
Correlation analysis was performed using Spearman's rank
correlation coefficients. P<0.05 was considered to indicate a
statistically significant difference.
Results
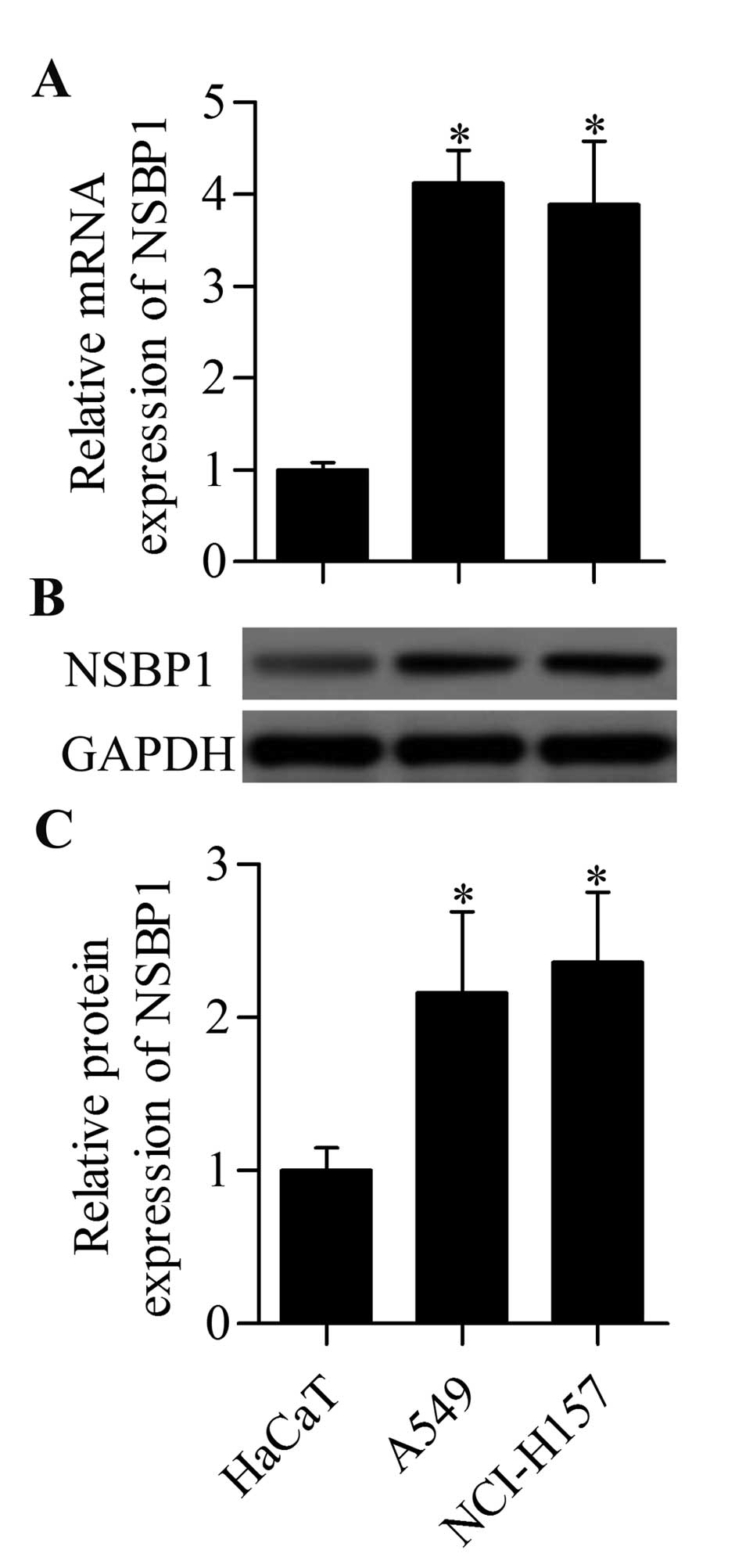
NSBP1 is highly expressed in human NSCLC
cells
To explore the potential role of NSBP1 in human
NSCLC, the expression patterns of NSBP1 in the NSCLC cell lines
were examined by RT-qPCR and western blot analysis. The results
showed that the mRNA expression of NSBP1 was significantly higher
in A549 and NCI-H157 cells than that in control HaCaT cells
(Fig. 1A). Moreover, the NSBP1
protein expression was also highly upregulated in A549 and NCI-H157
cells (Fig. 1B and C). These
results suggested a critical role of NSBP1 in NSCLC cells.
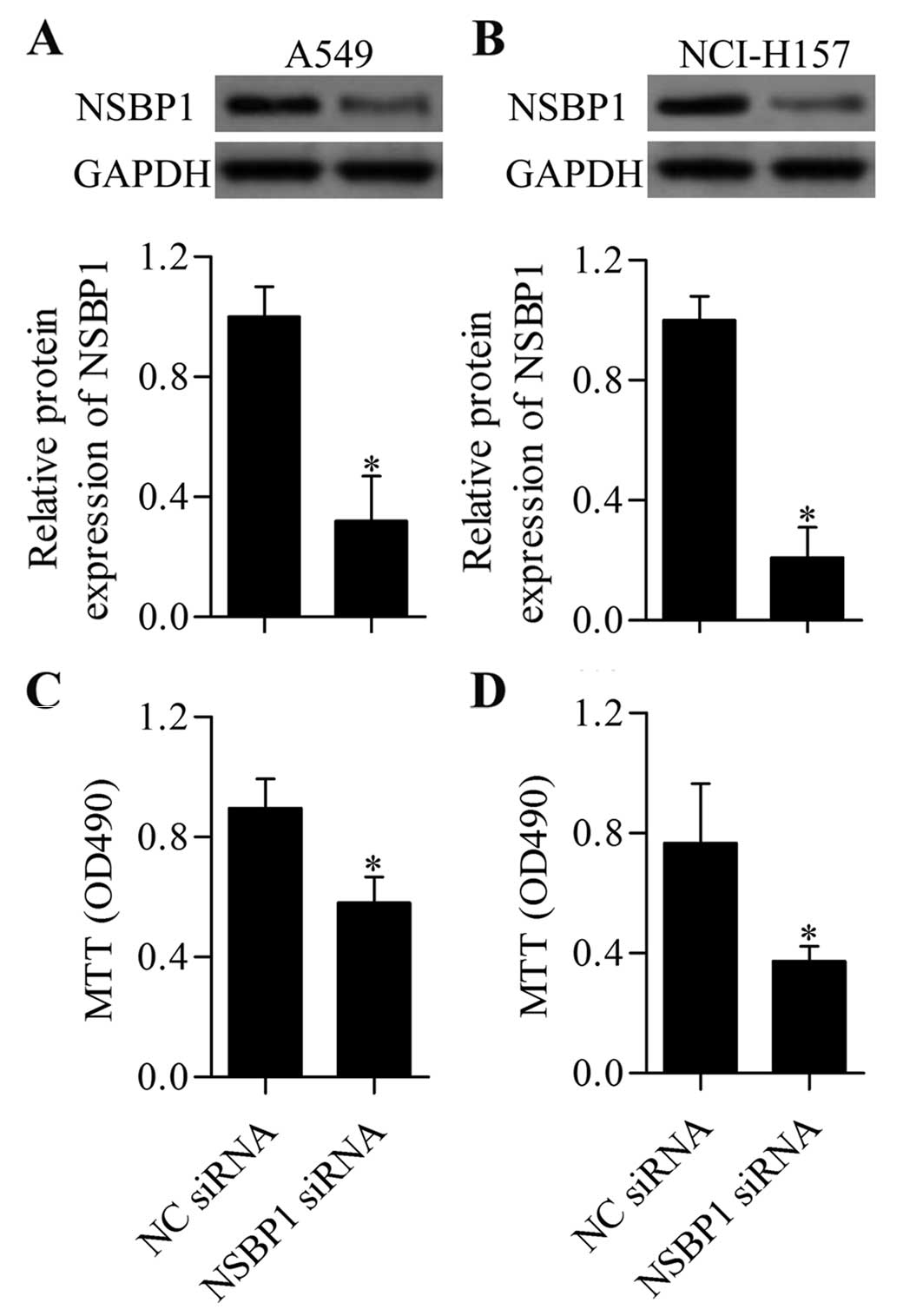
Knockdown of NSBP1 inhibits NSCLC cell
proliferation and invasion
To understand the functional significance of NSBP1
in regulating the biological processes of NSCLC cells, we performed
loss-of-function experiments of NSBP1 by transfection with specific
siRNA targeting NSBP1. The transfection efficiency was detected by
western blot analysis, which showed that NSBP siRNA effectively
downregulated the expression of NSBP1 in A549 (Fig. 2A) and NCI-H157 (Fig. 2B) cells. Subsequently, we measured
the effect of NSBP1 suppression on NSCLC cell proliferation. MTT
assay showed that knockdown of NSBP1 significantly inhibited A549
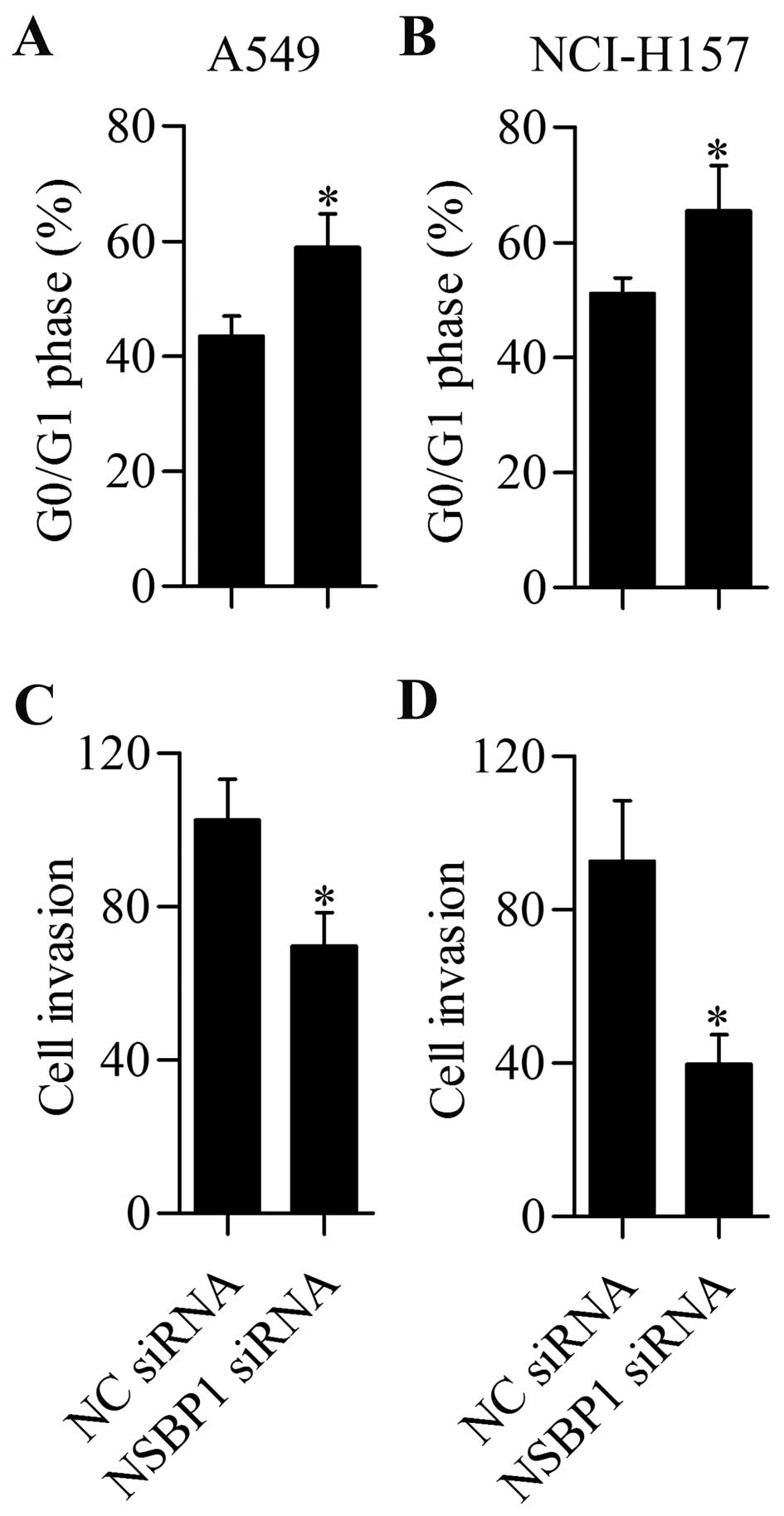
(Fig. 2C) and NCI-H157 (Fig. 2D) cell proliferation. Furthermore,
cell cycle distribution showed that knockdown of NSBP1 apparently
induced G0/G1 cell cycle arrest of A549 (Fig. 3A) and NCI-H157 (Fig. 3B) cells. To further investigate the
biological effect of NSBP1 in NSCLC cells, we examined the effect
of NSBP1 knockdown on NSCLC cell invasion. We found that
suppression of NSBP1 significantly repressed the invasive ability
of A549 (Fig. 3C) and NCI-H157
(Fig. 3D) cells. In summary, these
results indicated that NSBP1 participated in the regulation of
NSCLC cell proliferation and invasion.
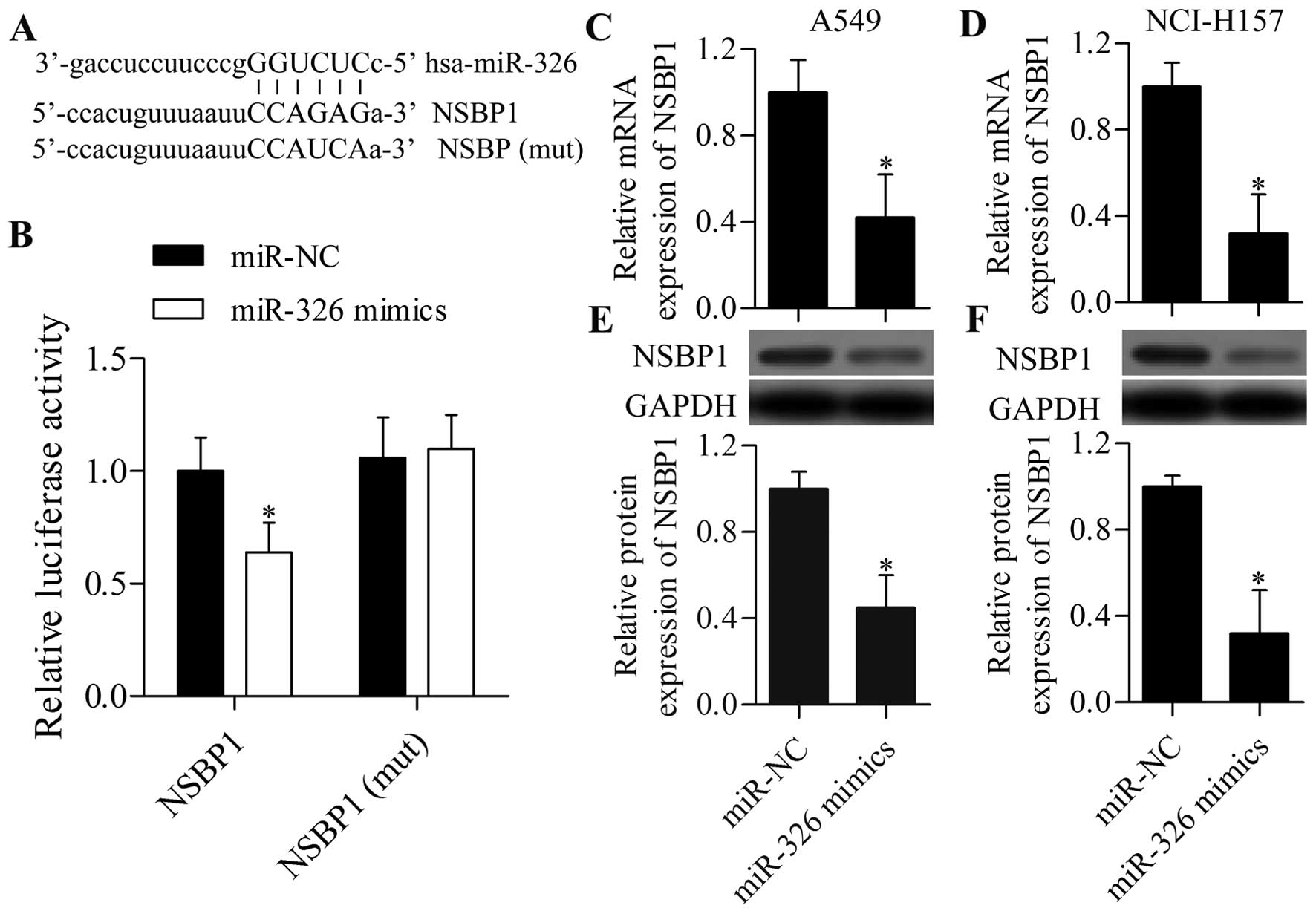
miR-326 targets the 3′-UTR of NSBP1 and
regulates NSBP1 expression
An increasing number of studies have suggested that
miRNAs have the potential to treat lung cancer via targeting
specific genes (21). To date, the
miRNAs specifically targeting NSBP1 have not been well
characterised. In the present study, we searched for specific
miRNAs that could target and regulate NSBP, and investigated
whether NSBP1 expression can be regulated by miRNAs. Bioinformatics
analysis demonstrated that miR-326 had a predicted binding site in
the 3′-UTR of NSBP1 (Fig. 4A). We
then performed a dual-luciferase reporter experiment to verify
their direct relationship. The results showed that overexpression
of miR-326 markedly repressed the relative luciferase activity of
pmirGLO-NSBP1, but it did not affect the relative luciferase
activity of pmirGLO-NSBP1 (mut)-transfected cells (Fig. 4B).
To investigate whether miR-326 can regulate NSBP1
expression in NSCLC cells, we transfected miR-326 mimics into NSCLC
cells and detected NSBP1 expression using RT-qPCR and western blot
analysis. RT-qPCR analysis showed that miR-326 overexpression
significantly suppressed the mRNA expression of NSBP1 in A549
(Fig. 4C) and NCI-H157 (Fig. 4D) cells. Western blot analysis
further displayed that the protein expression of NSBP1 was also
markedly repressed by miR-326 mimics in A549 (Fig. 4E) and NCI-H157 (Fig. 4F) cells. These results indicated
that miR-326 could target the 3′-UTR of NSBP1 and repress its
expression in NSCLC cells.
Overexpression of miR-326 suppresses
NSCLC cell proliferation and invasion
Given the inhibitory effect of miR-326 on NSBP1
expression, miR-326 might have a biological effect on NSCLC cells.
To test this hypothesis, we examined the biological effect of
miR-326 on cell proliferation and invasion in NSCLC cells.
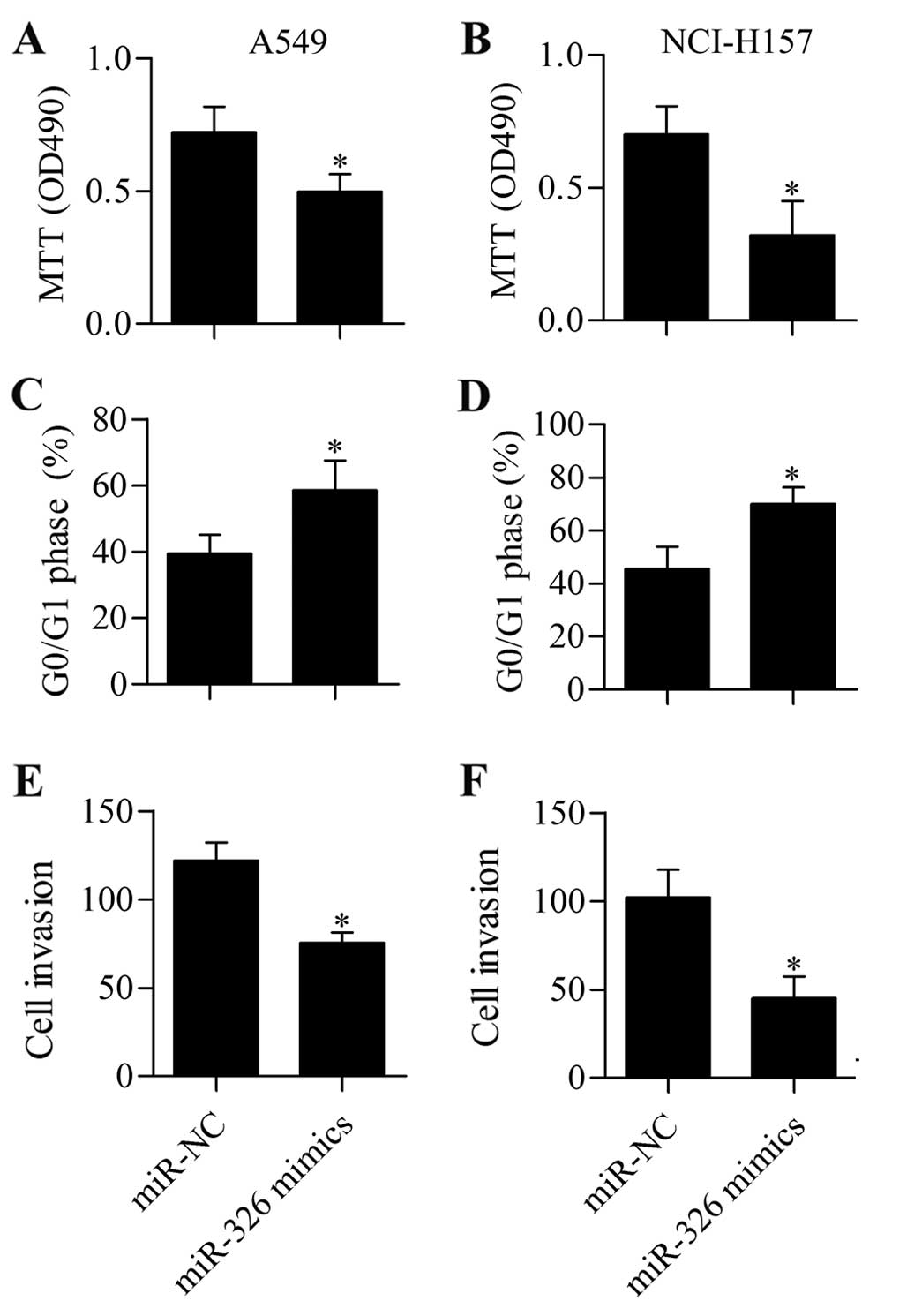
Interestingly, MTT assay demonstrated that transfection of miR-326
significantly inhibited cell proliferation of A549 (Fig. 5A) and NCI-H157 (Fig. 5B) cells. Moreover, miR-326
overexpression induced G0/G1 cell cycle arrest in A549 (Fig. 5C) and NCI-H157 (Fig. 5D) cells. The results of Transwell
assay suggested that miR-326 mimic-transfected cells showed
significantly decreased cell invasive capacity (Fig. 5E and F).
NSBP1 gene silencing induced by siRNA or
miR-326 overexpression represses cyclin B1 and MMP9 in NSCLC
cells
To further understand the molecular basis of NSBP1
gene silencing induced by NSBP1 siRNA or miR-326 in regulating
NSCLC cell proliferation and invasion, we detected the regulatory
effect of NSBP1 gene silencing on cyclin B1 and MMP9, which were
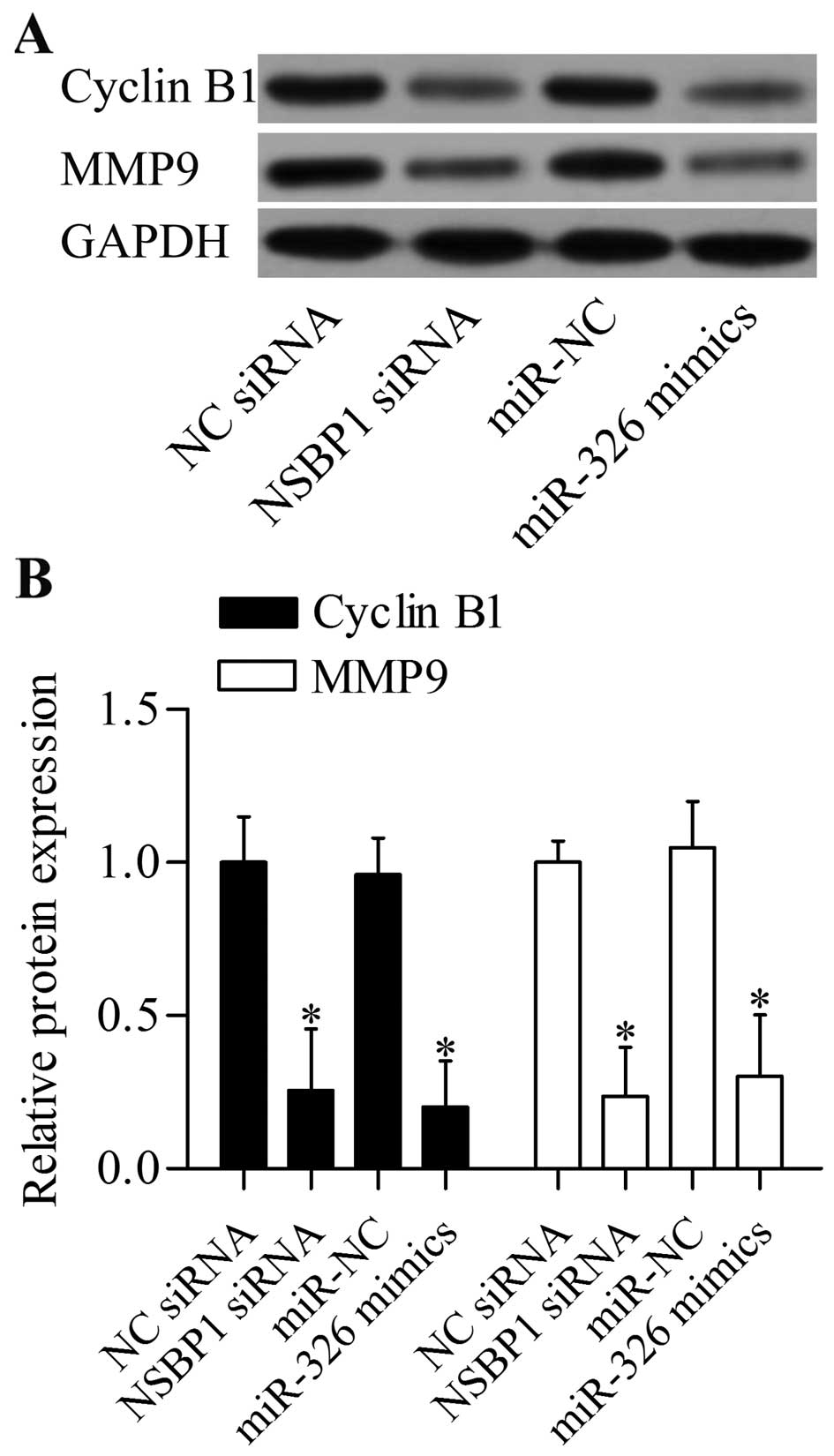
involved in regulating cancer cell proliferation and invasion. We
found that suppression of NSBP1 by siRNA or miR-326 inhibited the
protein expression levels of cyclin B1 and MMP9 in A549 (Fig. 6A and B).
miR-326 regulates cyclin B1 and MMP9
expression through NSBP1
To investigate whether miR-326 overexpression
suppressed cyclin B1 and MMP9 directly through NSBP1, we performed
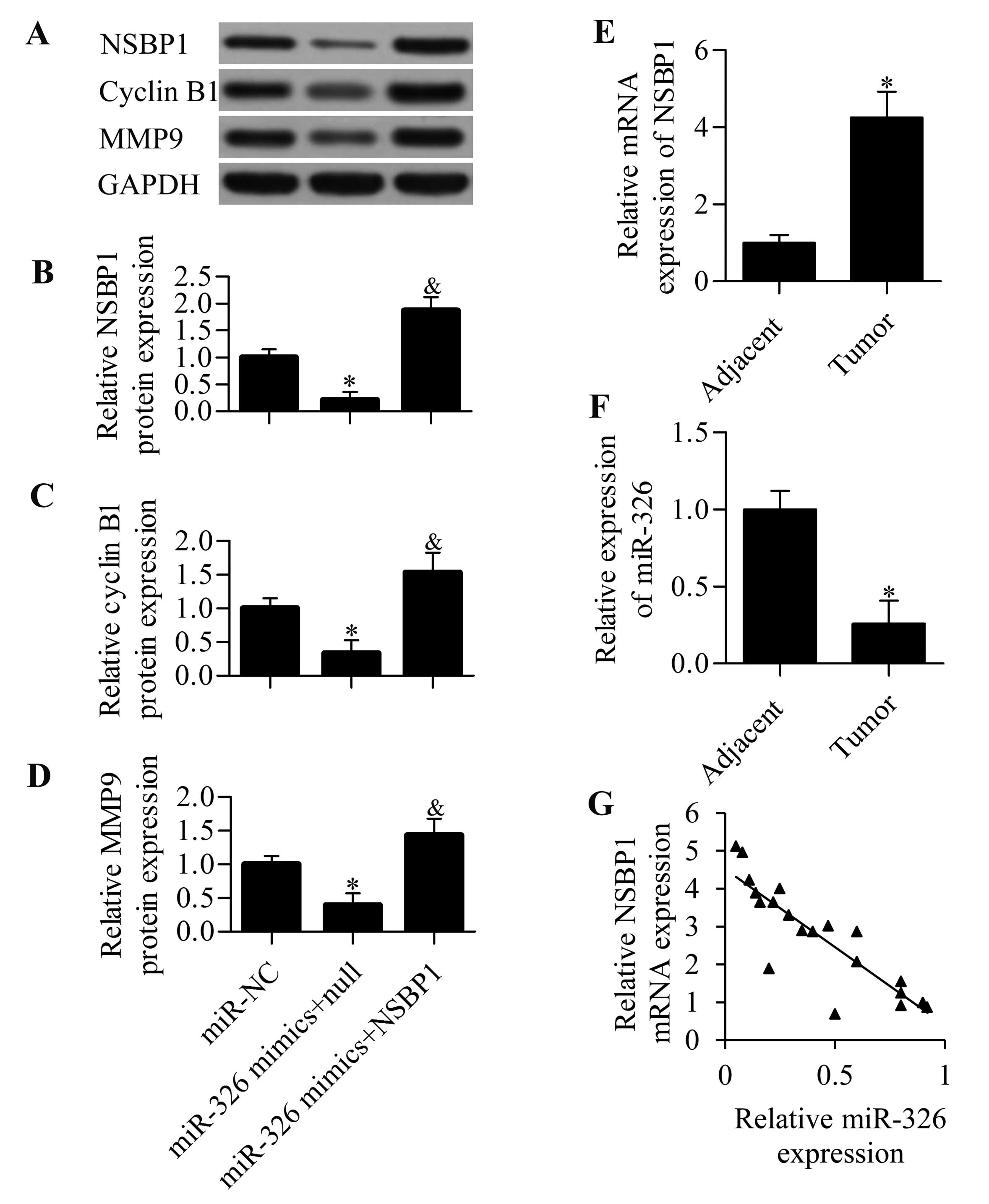
a rescue experiment by overexpression of NSBP1. We found that
transfection of NSBP expression vectors harbouring no 3′-UTR of
NSBP1 significantly restored NSBP1 expression, which was decreased
by the transfection of miR-326 mimics (Fig. 7A and B). NSBP1 overexpression
restored the decreased protein expression levels of cyclin B1 and
MMP9 induced by miR-326 overexpression (Fig. 7A, C and D), indicating that miR-326
inhibited cyclin B1 and MMP9 expression through NSBP1.
miR-326 expression is inversely
correlated with NSBP1 expression in NSCLC specimens
To gain insight into the functional significance of
miR-326 and NSBP1 expression in NSCLC, we uncovered their
expression levels in clinical NSCLC tissues and elucidated their
potential relationship. We found that NSBP1 mRNA expression was
significantly increased in NSCCL specimens (Fig. 7E), whereas miR-326 expression was
markedly decreased (Fig. 7F)
compared with that in adjacent non-tumour tissues. We correlated
NSBP1 mRNA expression and miR-326 expression using correlation
analysis, which demonstrated that miR-326 expression was remarkably
inversely correlated with NSBP1 expression in NSCLC tissues
(Fig. 7G) (r=−0.8756, P<0.001),
implying that decreased miR-326 might contribute to the increased
NSBP1 expression during NSCCL development and progression.
Discussion
The present study showed that suppression of NSBP1
by miR-326 could inhibit NSCLC cell proliferation and invasion. We
demonstrated that NSBP1, which functioned as an oncogene in NSCLC,
could be targeted by miR-326. Overexpression of miR-326
significantly inhibited NSBP1 expression, which was analogous to
NSBP1 siRNA. Our study suggested that miR-326 was a novel
therapeutic target for treating NSCCL by suppressing NSBP1.
NSBP1 plays a critical role in modulating gene
transcription (12). Thus,
dysregulation of NSBP1 may be associated with disease generation
and development. Increasing evidence revealed that NSBP1 is
extensively involved in tumourigenesis. Jiang et al reported
that knockdown of NSBP1 by RNA interference inhibits prostate
cancer cell proliferation in vitro and in vivo
(15). Knockdown of NSBP1 causes
cell cycle arrest and promotes cell apoptosis of prostate cancer
cells possibly by inhibiting cyclin B1 and Bcl-2 expression levels
(15). Similarly, Zhang et
al showed that NSBP1 gene silencing results in increased
mitochondria-mediated cell apoptosis in prostate cancer cells
(22). Furthermore, NSBP1 is
overexpressed in glioma clinical tissues, and knockdown of NSBP1
induces cell cycle arrest in the G1 phase, inhibits cell
proliferation and promotes cell apoptosis in glioma cells in
vitro (16). Moreover, NSBP1
was demonstrated to be overexpressed in bladder cancer (17), clear cell renal cell carcinoma
(23), osteosarcoma (19) and breast cancer (18). These studies also reported that
NSBP1 regulated cancer cell proliferation, apoptosis and invasion.
In the present study, we found that NSBP1 was highly expressed in
NSCLC cells, and knockdown of NSBP1 by NSBP1 siRNA significantly
inhibited cell proliferation and invasion of NSCLC cells.
Furthermore, knockdown of NSBP1 suppressed cyclin B1 and MMP9
expression levels, which might be the molecular basis for NSBP1
knockdown-induced decreased cell proliferation and invasion of
NSCLC cells. Our results were consistent with the findings of Chen
et al, who reported that knockdown of NSBP1 by siRNA
inhibits cell proliferation and induces cell cycle arrest in the G1
phase in lung cancer cells (20).
Notably, high expression of NSBP1 was also found to be associated
with drug resistance and ionising radiation in cancer cells
(24,25). Collectively, our data, as well as
the aforementioned findings, support the notion that NSBP1 could be
a candidate target for developing cancer therapies.
Recent emerging studies have proposed miRNAs as
novel therapeutic tools for cancer treatment by negatively
modulating target gene expression (5,6,9–11).
These reports prompted us to identify specific miRNAs that could
target and regulate NSBP1 expression. Intriguingly, we found that
miR-326 possessed a predicted binding site in the 3′-UTR of NSBP1,
implying that miR-326 may directly modulate NSBP1 expression.
Dual-luciferase activity validated the interaction between miR-326
and NSBP1 3′-UTR. Further experiments using RT-qPCR and western
blot analysis confirmed that miR-326 inhibited NSBP1 expression in
NSCLC cells. Strikingly, overexpression of miR-326 markedly
inhibited cell proliferation and invasion of NSCLC cells, which
mimicked the effect of NSBP1 siRNA, suggesting that miR-326 exerted
a tumour suppressive function by inhibiting NSBP1 expression.
Several studies have reported the potential therapeutic function of
miR-326 against tumours. Kefas et al revealed that
transfection of miR-326 significantly decreases cell growth and
cell tumourigenicity of glioma cells (26,27).
Similarly, miR-326 expression is low in glioma tissues, and low
miR-326 expression is associated with advanced pathological grade
of gliomas (28). In pancreatic
ductal adenocarcinoma, miR-326 significantly decreased in patients,
and patients with relatively high miR-326 expression displayed
long-term survival or without venous invasion, implying that
miR-326 is a tumour suppressor in pancreatic ductal adenocarcinoma
(29). Moreover, miR-326 was shown
to exert a tumour suppressive role in glioma (30) or colorectal cancer (31) by targeting the Nin one binding
protein that can inhibit cancer cell proliferation, migration and
invasion, and promote cell apoptosis. Interestingly, miR-326 was
found to suppress cancer cell invasion by targeting the disintegrin
and metalloprotease 17 in lung adenocarcinoma (32). In line with these findings, we also
found that miR-326 was frequently downregulated in NSCLC tissues,
and overexpression of miR-326 inhibited cancer cell proliferation
and invasion. These findings suggested that miR-326 functioned as a
tumour suppressor.
Taken together, our study demonstrated a direct
target relationship between NSBP1 and miR-326 through which miR-326
inhibited cell proliferation and invasion of NSCLC cells. Loss of
NSBP1 induced by NSBP1 siRNA or miR-326 overexpression repressed
the expression levels of cyclin B1 and MMP9, which contribute to
cancer cell proliferation and invasion. Conversely, overexpression
of NSBP1 apparently rescued the decreased cyclin B1 and MMP9
expression levels induced by miR-326 overexpression, further
confirming that miR-326 exerted its tumour suppressive function by
targeting NSBP1. Most importantly, we observed that miR-326
expression was inversely correlated with NSBP1 expression in NSCLC
clinical specimens, further confirming its functional significance
in NSCLC. Therefore, our study suggested that miR-326-NSBP1 could
be employed as a promising candidate target for developing novel
anticancer therapeutics for NSCLC.
Abbreviations:
|
miRs
|
microRNAs
|
|
UTR
|
untranslated region
|
|
NSBP1
|
nucleosome-binding protein
|
|
NSCLC
|
non-small cell lung cancer
|
|
MMP9
|
matrix metalloproteinase-9
|
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Travis WD, Brambilla E, Noguchi M,
Nicholson AG, Geisinger R, Yatabe Y, Beer DG, Powell CA, Riely GJ,
Van Schil PE, et al: International Association for the study of
lung cancer/American Thoracic Society/European Respiratory Society
International multidisciplinary classification of lung
adenocarcinoma. J Thorac Oncol. 6:244–285. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Claassens L, van Meerbeeck J, Coens C,
Quinten C, Ghislain I, Sloan EK, Wang XS, Velikova G and Bottomley
A: Health-related quality of life in non-small cell lung cancer: An
update of a systematic review on methodologic issues in randomized
controlled trials. J Clin Oncol. 29:2104–2120. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hu Z, Chen X, Zhao Y, Tian T, Jin G, Shu
Y, Chen Y, Xu L, Zen K, Zhang C, et al: Serum microRNA signatures
identified in a genome-wide serum microRNA expression profiling
predict survival of non-small cell lung cancer. J Clin Oncol.
28:1721–1726. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Winter J, Jung S, Keller S, Gregory RI and
Diederichs S: Many roads to maturity: MicroRNA biogenesis pathways
and their regulation. Nat Cell Biol. 11:228–234. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rottiers V, Najafi-Shoushtari SH, Kristo
F, Gurumurthy S, Zhong L, Li Y, Cohen DE, Gerszten RE, Bardeesy N,
Mostoslavsky R, et al: MicroRNAs in metabolism and metabolic
diseases. Cold Spring Harb Symp Quant Biol. 76:225–233. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Aigner A: MicroRNAs (miRNAs) in cancer
invasion and metastasis: Therapeutic approaches based on
metastasis-related miRNAs. J Mol Med Berl. 89:445–457. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Esquela-Kerscher A and Slack FJ: Oncomirs
- microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lu J, Getz G, Miska EA, Alvarez-Saavedra
E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA,
et al: MicroRNA expression profiles classify human cancers. Nature.
435:834–838. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hock R, Furusawa T, Ueda T and Bustin M:
HMG chromosomal proteins in development and disease. Trends Cell
Biol. 17:72–79. 2007. View Article : Google Scholar
|
|
13
|
Shirakawa H, Landsman D, Postnikov YV and
Bustin M: NBP-45, a novel nucleosomal binding protein with a
tissue-specific and developmentally regulated expression. J Biol
Chem. 275:6368–6374. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
King LM and Francomano CA:
Characterization of a human gene encoding nucleosomal binding
protein NSBP1. Genomics. 71:163–173. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jiang N, Zhou LQ and Zhang XY:
Downregulation of the nucleosome-binding protein 1 (NSBP1) gene can
inhibit the in vitro and in vivo proliferation of prostate cancer
cells. Asian J Androl. 12:709–717. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Qu J, Yan R, Chen J, Xu T, Zhou J, Wang M,
Chen C, Yan Y and Lu Y: HMGN5: A potential oncogene in gliomas. J
Neurooncol. 104:729–736. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wahafu W, He ZS, Zhang XY, Zhang CJ, Yao
K, Hao H, Song G, He Q, Li XS and Zhou LQ: The nucleosome binding
protein NSBP1 is highly expressed in human bladder cancer and
promotes the proliferation and invasion of bladder cancer cells.
Tumour Biol. 32:931–939. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Weng M, Song F, Chen J, Wu J, Qin J, Jin T
and Xu J: The high-mobility group nucleosome-binding domain 5 is
highly expressed in breast cancer and promotes the proliferation
and invasion of breast cancer cells. Tumour Biol. 36:959–966. 2015.
View Article : Google Scholar
|
|
19
|
Zhou X, Yuan B, Yuan W, Wang C, Gao R and
Wang J: The expression and clinical significance of high mobility
group nucleosome binding domain 5 in human osteosarcoma. Tumour
Biol. 35:6539–6547. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen P, Wang XL, Ma ZS, Xu Z, Jia B, Ren
J, Hu YX, Zhang QH, Ma TG, Yan BD, et al: Knockdown of HMGN5
expression by RNA interference induces cell cycle arrest in human
lung cancer cells. Asian Pac J Cancer Prev. 13:3223–3228. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Barger JF and Nana-Sinkam SP: MicroRNA as
tools and therapeutics in lung cancer. Respir Med. 109:803–812.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang XY, Guo ZQ, Ji SQ, Zhang M, Jiang N,
Li XS and Zhou LQ: Small interfering RNA targeting HMGN5 induces
apoptosis via modulation of a mitochondrial pathway and Bcl-2
family proteins in prostate cancer cells. Asian J Androl.
14:487–492. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ji SQ, Yao L, Zhang XY, Li XS and Zhou LQ:
Knockdown of the nucleosome binding protein 1 inhibits the growth
and invasion of clear cell renal cell carcinoma cells in vitro and
in vivo. J Exp Clin Cancer Res. 31:222012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang C, Gao R, Wang J, Yuan W, Wang C and
Zhou X: High-mobility group nucleosome-binding domain 5 increases
drug resistance in osteosarcoma through upregulating autophagy.
Tumour Biol. 35:6357–6363. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Su B, Shi B, Tang Y, Guo Z, Yu X, He X, Li
X, Gao X and Zhou L: HMGN5 knockdown sensitizes prostate cancer
cells to ionizing radiation. Prostate. 75:33–44. 2015. View Article : Google Scholar
|
|
26
|
Kefas B, Comeau L, Floyd DH, Seleverstov
O, Godlewski J, Schmittgen T, Jiang J, diPierro CG, Li Y, Chiocca
EA, et al: The neuronal microRNA miR-326 acts in a feedback loop
with notch and has therapeutic potential against brain tumors. J
Neurosci. 29:15161–15168. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kefas B, Comeau L, Erdle N, Montgomery E,
Amos S and Purow B: Pyruvate kinase M2 is a target of the
tumor-suppressive microRNA-326 and regulates the survival of glioma
cells. Neuro-oncol. 12:1102–1112. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang S, Lu S, Geng S, Ma S, Liang Z and
Jiao B: Expression and clinical significance of microRNA-326 in
human glioma miR-326 expression in glioma. Med Oncol. 30:3732013.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang ZL, Bai ZH, Wang XB, Bai L, Miao F
and Pei HH: miR-186 and 326 predict the prognosis of pancreatic
ductal adenocarcinoma and affect the proliferation and migration of
cancer cells. PLoS One. 10:e01188142015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhou J, Xu T, Yan Y, Qin R, Wang H, Zhang
X, Huang Y, Wang Y, Lu Y, Fu D, et al: MicroRNA-326 functions as a
tumor suppressor in glioma by targeting the Nin one binding protein
(NOB1). PLoS One. 8:e684692013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wu L, Hui H, Wang LJ, Wang H, Liu QF and
Han SX: MicroRNA-326 functions as a tumor suppressor in colorectal
cancer by targeting the nin one binding protein. Oncol Rep.
33:2309–2318. 2015.PubMed/NCBI
|
|
32
|
Cai M, Wang Z, Zhang J, Zhou H, Jin L, Bai
R and Weng Y: Adam17, a target of miR-326, promotes EMT-induced
cells invasion in lung adenocarcinoma. Cell Physiol Biochem.
36:1175–1185. 2015. View Article : Google Scholar : PubMed/NCBI
|