Introduction
Breast cancer is the second most commonly diagnosed
type of cancer, and the leading cause of cancer-related death in
American women. In the United States, 240,000 mostly postmenopausal
women are diagnosed with the disease every year (1). A number of recent independent clinical
trials and studies have shown that postmenopausal women undergoing
combined estrogen and progestin hormone-replacement therapy (HRT)
have an increased risk of developing metastatic breast cancer
compared with women taking only estrogen (2–5).
Combination HRT is commonly prescribed to women with an intact
uterus to alleviate postmenopausal symptoms. The progestin
component is added to minimize the risk of endometrial hyperplasia,
which may precede endometrial cancer (6,7).
Recent laboratory studies have shown that progestin stimulates
proliferation of normal and neoplastic breast cancer cells
(8,9), correlating with clinical findings for
combination HRT use.
Studies designed to elucidate the mechanism(s)
underlying the increased incidence of breast cancer associated with
combination HRT have shown that progestins induce the potent
angiogenic factor VEGF, in human breast cancer cells (10–12).
Furthermore, such studies have demonstrated that both natural and
synthetic progestins increase the synthesis and secretion of VEGF
in breast cancer cells that express mutant p53 tumor-suppressor
protein, but not wild-type p53 (12,13).
Locally produced VEGF acts in a paracrine manner to stimulate both
endothelial and tumor epithelial cells; the latter cells are also
stimulated by VEGF in an autocrine fashion (14). In addition, progestins such as
medroxyprogesterone acetate (MPA), a commonly prescribed component
of HRT, have been implicated in reactivating breast cancer stem
cell subpopulations in hormone-responsive cell lines (12,15).
Progestins are also believed to act in concert with RANKL to
increase cellular proliferation (9,16), and
increase tumor vasculature (12,17),
thereby providing an enriched environment for tumor growth and
metastasis.
Previous studies in our laboratory have shown that
progestins drive 7,12-dimethylbenz(a)anthracene (DMBA)-induced
hormone-dependent mammary tumors in Sprague-Dawley rats (17–19).
We have exploited this finding to establish an in vivo model
for testing progestin antagonists in an inclusive microenvironment.
Although the exact mechanism behind progestin-accelerated tumor
growth in the DMBA model is not fully understood, evidence suggests
that progestin-induced VEGF production resulting in increased
angiogenesis is likely responsible (17–19).
Previous studies have reported that antiprogestins (both synthetic,
such as RU-486, and naturally-occurring compounds, such as
apigenin) block the incidence and growth of mammary tumors in the
progestin-accelerated DMBA-induced model (17,19–21).
These studies suggest that progestin-accelerated DMBA-induced
mammary tumor incidence is largely influenced by the inherent
ability of progestins to increase production of VEGF, resulting in
decreased latency and increased tumor incidence, burden, and
multiplicity. For this reason, the model provides an excellent
means of studying hormone-dependent breast cancer and is
particularly suitable for identifying naturally-occurring,
non-toxic antagonists of progestin-induced VEGF.
Luteolin
[2-(3,4-dihydroxyphenyl)-5,7-dihydroxy-4H-1-benzopyran-4-one] (LU)
is a low molecular weight, naturally-occurring flavonoid commonly
found in fruits and vegetables. A number of studies show that LU
possesses a myriad of anticancer functions and that it suppresses
tumor development in several types of human cancer more effectively
than other flavonoids (22–24). Studies show that LU inhibits VEGF
production and that it has antiprogestin capabilities (25–29),
demonstrating its potential effectiveness against
hormone-responsive cancers. Collectively, these findings suggest
that LU has important chemopreventive properties. Herein, we
provide evidence supporting the ability of LU to substantially
inhibit MPA-accelerated tumor latency, incidence and growth in the
DMBA-induced mammary tumor model. We also show that LU possibly
arrests the growth of mammary tumors by suppressing VEGF production
and angiogenesis, vital components of breast tumor formation and
development.
Materials and methods
Animals
All surgical and experimental procedures were
approved by the University of Missouri-Columbia Institutional
Animal Care and Use Committee (IACUC). Intact adult female
Sprague-Dawley rats (45–55-day old) were purchased from Harlan
Breeders (Indianapolis, IN, USA) and maintained under 12-h
light/dark cycles with ad libitum access to food (LabDiet
5008; LabDiet, St. Louis, MO, USA) and water in accordance with
guidelines established by the Association for Assessment and
Accreditation of Laboratory Animal Care International (AAALAC).
Luteolin
LU was purchased from Indofine Chemical Company (cat
no. L-101; Hillsborough, NJ, USA) and dissolved in sterile filtered
dimethyl sulfoxide (DMSO, cat no. D2650; Sigma-Aldrich, St. Louis,
MO, USA). Solutions of LU were prepared weekly, aliquoted for daily
use, and stored at −20°C until use.
Experimental design
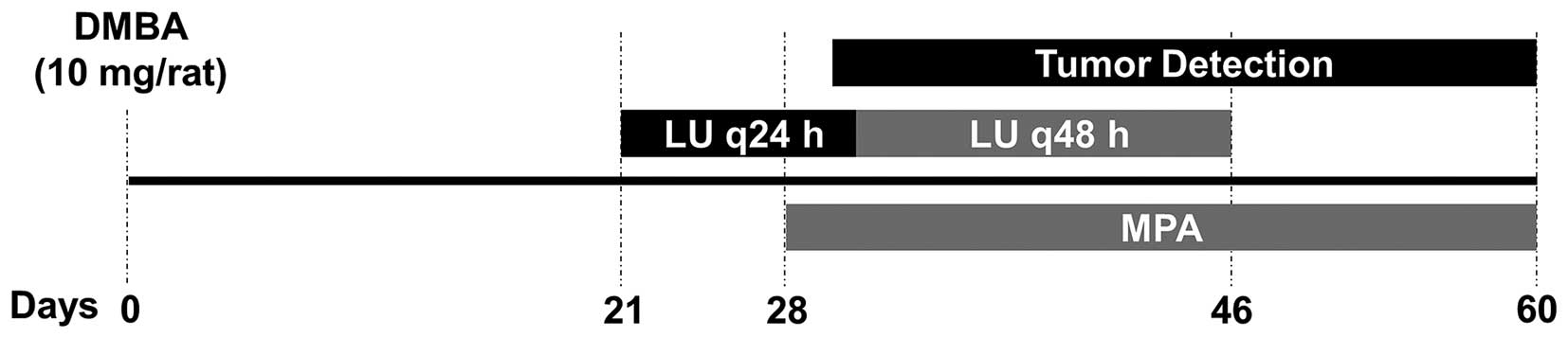
We modified the protocol of DMBA-induced mammary
tumor formation described previously (17,19)
(summarized in Fig. 1). Animals
were given a 1 ml bolus of 10 mg of DMBA (cat no. D3254;
Sigma-Aldrich) dissolved in peanut oil by oral gavage (day 0).
Three weeks post-DMBA administration (day 21), animals were divided
into 5 treatment groups (n=10–12 animals/group). Animals in the
control group and those given only MPA were administered DMSO by
intraperitoneal injection. The animals given LU (1, 10, or 25
mg/kg) received injections of the flavonoid in DMSO every 24 h for
10 days, followed by another 8 injections at 48-h intervals. LU
doses were selected based on previously reported in vivo
studies (30–32). Four weeks post-DMBA administration
(day 28), 25 mg 60-day release MPA or placebo pellets (cat no.
P-161; Innovative Research of America, Sarasota, FL, USA) were
implanted subcutaneously on the dorsal part of the neck. Animals
were weighed twice a week and, starting on day 29, palpated every
other day to detect tumor latency and incidence. On day 59, all
animals were sacrificed and tumor number and volume (1/2 L x
W2) (33,34) determined. Tumors and contralateral
inguinal mammary gland tissue devoid of tumors were retained for
analysis.
Histology and immunohistochemical
analysis
Immunohistochemical staining of mammary and tumor
tissue was performed following previously described procedures
(17,19). The following polyclonal antibodies
were used: anti-VEGF antibody (1:100 dilution, cat no. SC-152;
Santa Cruz Biotechnology Dallas, TX, USA); and anti-Ki67 antigen
antibody (1:400 dilution, cat no. RB1510-P; Thermo Scientific
Waltham, MA, USA). Cell death immunohistochemistry was determined
using a Roche (Basel, Switzerland) terminal deoxynucleotidyl
transferase-mediated dUTP nick end labeling (TUNEL) detection kit.
Histological samples were analyzed and quantified using Fovea Pro
3.0 (Reindeer Graphics) and ImageJ. Images were captured at ×20 and
×40 magnification and threshold intensity was adjusted for
measurement in pixels. Tumors and representative contralateral
inguinal mammary gland tissues were excised from animals in each
treatment group, fixed in formaldehyde, and embedded in paraffin
for immunohistochemistry. One section from each individual tumor
and mammary gland was placed on the corresponding slide for each of
the immunohistochemical stains and 4 random fields captured from
every section to minimize errors due to differences in cellularity.
All the mammary tumors were collected and assessed for IHC
biomarkers. The availability of tumor sections was dependent upon
tumor occurrence (while 6–9 tumors developed in control, MPA and
MPA + 10 mg/kg LU groups, only 2 and 3 animals developed tumors in
the 1 and 25 mg/kg LU-treated animal group, respectively). Regions
of staining within tumors and areas of mammary hyperplasia in
contralateral inguinal mammary gland tissue were recorded. For
analysis of mammary gland tissue, only four glands were used in
each group. Fovea Pro 3.0 was used to quantitate the percent area
of VEGF, Ki67 and TUNEL staining in tumor tissue using the color
threshold feature in ImageJ. This facilitated precise
discrimination between positive/negative cells and background.
CD31, a blood vessel marker, was used to quantitate blood vessels
in excised tumor tissue. Three CD31-labeled ×10 sections were taken
from each tumor to minimize intratumoral variation, as previously
described (12). The total number
of vessels were counted in each section and then averaged per
corresponding tumor. Data was then reported as means per treatment
group with each group having an n≥3; except for the group given 25
mg/kg LU, which contained 1 tumor.
Statistical analysis
Tumor latency was analyzed using the LIFETEST
procedure in SAS software (9.4) to determine differences in
time-to-event. Pairwise comparisons between groups were made using
the Wilcoxon log-rank test in which the time-to-event represents
time to appearance of the first tumor in each animal. Tumor-free
animals were censored upon death or termination of the study (day
59). All other tumor burdened animals, regardless of survival, were
uncensored. Tumor incidence and number of tumors per category (up
to 300 mm3 or >300 mm3) were compared
pairwise using the general linear model (GENMOD) procedure in SAS
software to determine the differences in least squared means among
groups (logit link function and a binomial distribution). A logit
link with a distribution binomial p cannot be equal to 0 [logit =
ln (p/1-p)], thus the log of 0 is undefined. In the group treated
with MPA + 25 mg/kg LU, there were 0 tumors formed in >300
mm3 group, therefore a '1' was added to this group for
statistical analysis (i.e., 1 tumor in 10 animals, instead of 0 in
10 animals). No adjustment was made for multiple comparisons in the
LIFETEST or GENMOD procedures. Immunohistochemical data were
analyzed using an ANOVA followed by an all pairwise multiple
comparison test (Student-Newman-Keuls test) in SigmaPlot 12.5. IHC
analysis of VEGF in the groups given MPA and MPA + LU 25 mg/kg was
by t-test in SigmaPlot 12.5. In the group given 25 mg/kg LU (tumor
tissue was n=1), analysis of VEGF IHC by the Student-Newman-Keuls
multi-range test met the critical value between MPA and MPA + 25
mg/kg LU due to the large difference between treatment groups,
resulting in statistical significance. This test assumes that MPA +
25 mg/kg LU is a true representation of the mean (% area). Similar
statistical significance was not reached with the MPA + 25 mg/kg LU
groups for other markers analyzed by IHC. For all comparisons,
P≤0.05 was regarded as statistically significant.
Results
Luteolin suppresses development of
progestin-accelerated DMBA-induced mammary tumors
Using our well-established model of DMBA-induced
mammary tumors (17–19), we examined the potential of LU to
prevent MPA-driven tumor development. Three weeks after DMBA
administration and 1 week prior to implantation of the MPA pellet
(day 21), various doses of LU were administered to determine its
ability to impede MPA-dependent tumor development by preventing the
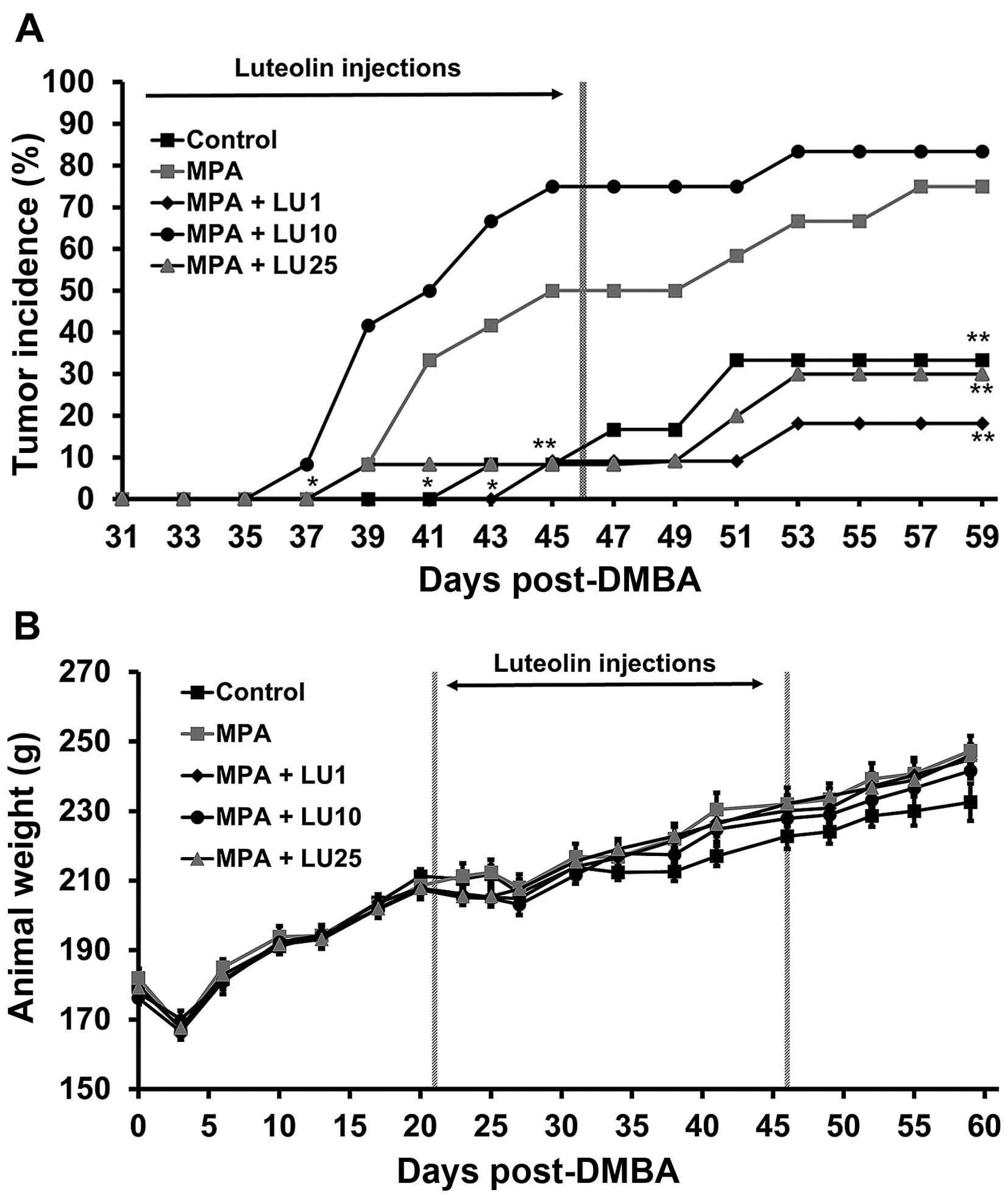
progression of neoplastic lesions to frank tumors. MPA reduced
tumor latency in DMBA-treated rats compared with controls
(DMBA-treated rats implanted with placebo pellets) (Fig. 2A; P<0.05). Interestingly, the
latency curve for the 10 mg/kg LU group was similar to that of the
MPA group, with no significant difference between the two profiles.
In contrast, in those animals given MPA + either 1 or 25 mg/kg LU,
time-to-event data (latency) increased significantly (LIFETEST;
*P<0.05) compared with animals given MPA alone.
At termination of LU treatment (day 46), tumor
incidence increased in animals treated with MPA alone and MPA + 10
mg/kg LU compared with controls and those administered 1 or 25
mg/kg LU + MPA (P<0.05) (Fig.
2A). Following cessation of LU treatment, tumor incidence in
animals receiving the flavonoid at a dose of 1 or 25 mg/kg remained
relatively low until the end of the experiment on day 59. As a
result, tumor incidence at day 59 in groups given 1 or 25 mg/kg LU
and controls was significantly reduced compared with that in
animals treated with MPA alone (Fig.
2A). Interestingly, administration of 10 mg/kg LU appeared to
have little or no inhibitory effect on MPA-driven tumor incidence
(Fig. 2A). Animal weights were not
significantly affected by LU at even the highest dose used (25
mg/kg) throughout these studies (Fig.
2B), indicating that the flavonoid had little or no
toxicity.
Even though more than one tumor developed in a few
animals (two animals in control, two in MPA, one in MPA + LU1, one
in MPA + LU10, and 0 in MPA + LU25 groups) there was no overall
significant difference in tumor multiplicity between groups. The
majority of tumors formed in this animal model were ductal
carcinomas with cribriform, papillary or a combination of
cribriform and papillary patterns. Ductal carcinomas were also the
predominant type of neoplasm detected in LU-treated rats and there
was no observable trend for a particular classification of neoplasm
as a response to the different treatments.
Luteolin suppresses progestin-driven
mammary tumor growth
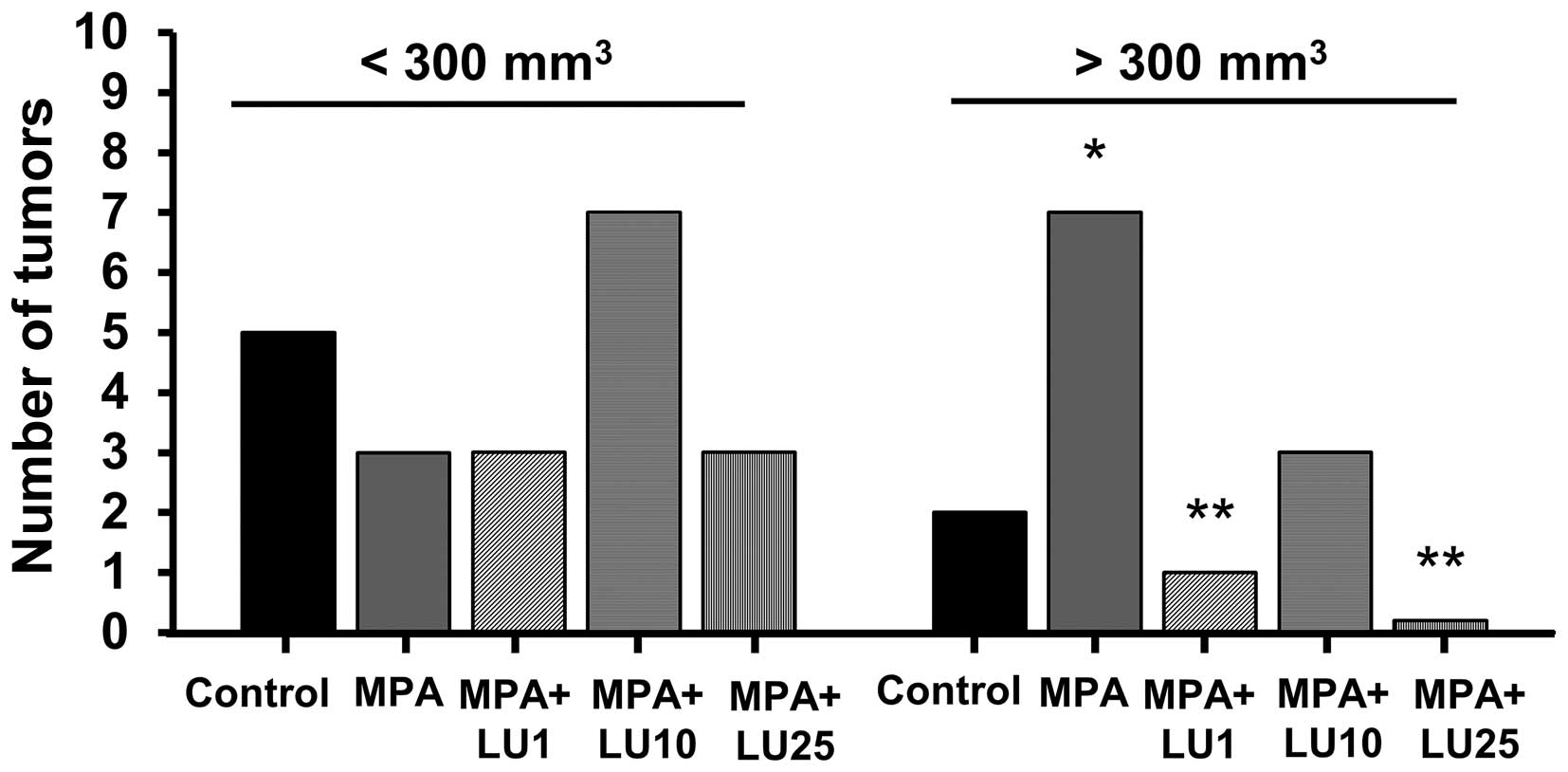
Due to the biological variance in the volume of
tumors within and among animal treatment groups (data not shown),
they were divided into two size groups, separating them into small
and large tumors (small, up to 300 mm3; large >300
mm3).
No statistical differences were observed among the
various treatment groups with respect to the numbers of small
tumors (<300 mm3) occurring in experimental animals
(Fig. 3), though more small tumors
developed in the group given MPA + 10 mg/kg LU, reflecting a higher
incidence of tumors in this group compared with other LU-treated
groups (Fig. 2A). However, the
number of large (>300 mm3) tumors arising in animals
receiving only MPA, was significantly higher than in the control
group (Fig. 3). Administration of 1
or 25 mg/kg LU significantly reduced the number of large tumors
compared with the number observed in the MPA-treated group
(Fig. 3), suggesting that LU
interfered with MPA-driven tumor volume increases. Interestingly,
although by day 59 no difference was observed in tumor incidence
between animals given MPA alone and those administered MPA + 10
mg/kg LU (Fig. 2A), more of the
tumors in the latter group were small (<300 mm3).
This finding suggests that a dose of 10 mg/kg LU, while not
affecting tumor incidence, suppresses MPA-driven tumor growth and
prevents the development of small tumors into larger ones.
Luteolin promotes mammary tumor
regression
Our initial results demonstrated that doses of 1 or
25 mg/kg LU most effectively suppressed progestin-dependent
increases in tumor incidence and growth. In the 1 mg/kg LU
treatment group, a total of only 4 tumors were detected (in 2 of 11
animals), while just 3 tumors were observed in the group given 25
mg/kg LU [in 3 of 10 animals (Fig.
2A)].
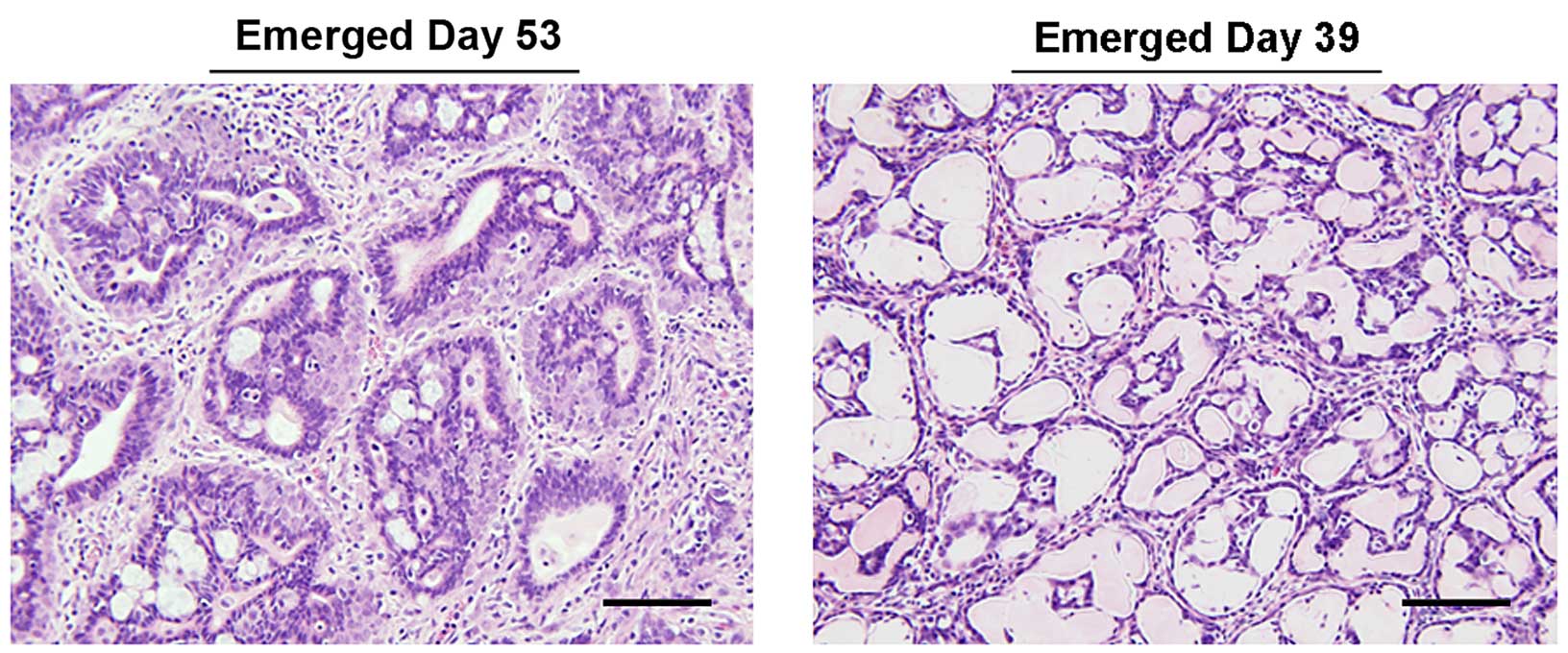
In the 25 mg/kg group, only 2 of the 3 tumors were
present during the last week of LU treatment. These tumors were
initially palpated on days 39 and 51, while the third tumor was
first palpated on day 53. Tumors detected on days 51 and 53
developed well after termination of LU treatment. The tumor
detected on day 53 contained a hypercellular stromal compartment
surrounded by nests of cribriform, hyperplastic glandular tissue
(Fig. 4). The first tumor which
arose on day 39 during LU treatment had decreased in size by the
time it was excised and examined on day 59 (Fig. 4). This mass was composed of
tightly-packed tubular structures with empty lumens and lined with
a flattened epithelium. The cause of this change is not known, but
has features suggestive of epithelial atrophy. The tumor detected
on day 51 was too small and not collectable at the end of the
experiment. Consequently, the tumor that emerged on day 53
(Fig. 4) was used alone for all
subsequent immunohistochemical analysis of tissues representing the
25 mg/kg LU treatment group.
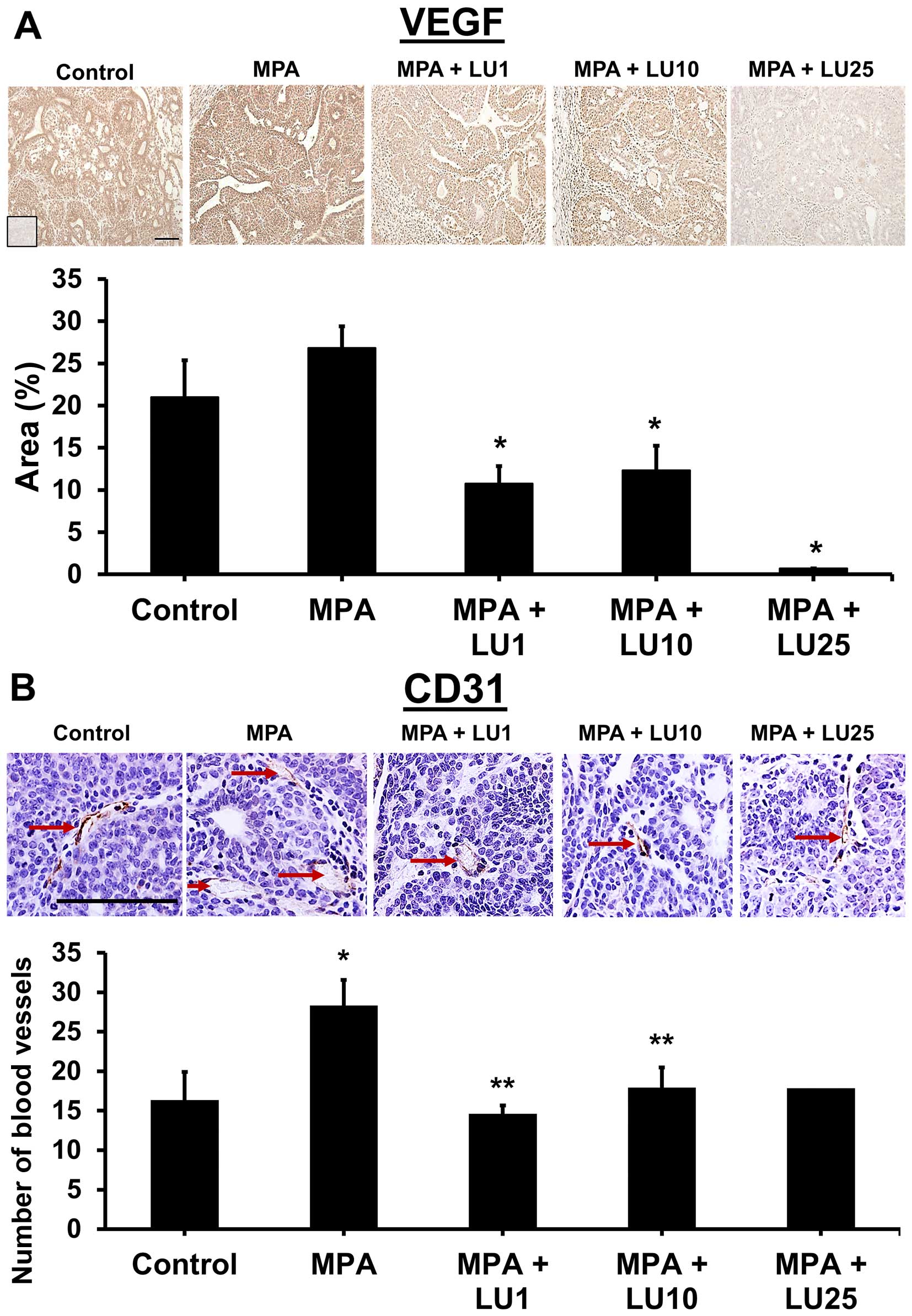
Luteolin reduces expression of VEGF and
CD-31, markers of angiogenesis, in mammary tumor tissue
In previous studies, we showed that continuous
production of VEGF by breast cancer cells is a vital component of
MPA-dependent angiogenesis and subsequent tumor development
(12,17,19).
In the current studies, we postulated that LU would reduce
progestin-accelerated tumor growth by suppressing MPA-induced VEGF
levels, thereby increasing tumor latency and reducing tumor
number.
Assessment of the immunohistochemical data
pertaining to the expression of specific markers showed that LU
significantly reduced levels of tumor-associated VEGF in all
treatment groups (1, 10, and 25 mg/kg) compared with MPA alone
(Fig. 5A). Non-tumor-associated
levels of VEGF in mammary glands was significantly reduced by the
highest dose of LU (25 mg/kg) compared with controls (data not
shown). MPA alone had no significant effect on VEGF levels in
either tumor or mammary gland tissues though there was a trend
towards higher levels of VEGF within tumors derived from MPA
treated animals (Fig. 5A and data
not shown). Tumors from MPA-treated animals exhibited a higher
number of blood vessels compared with controls and all three doses
of LU significantly suppressed tumor blood vessel formation
(Fig. 5B).
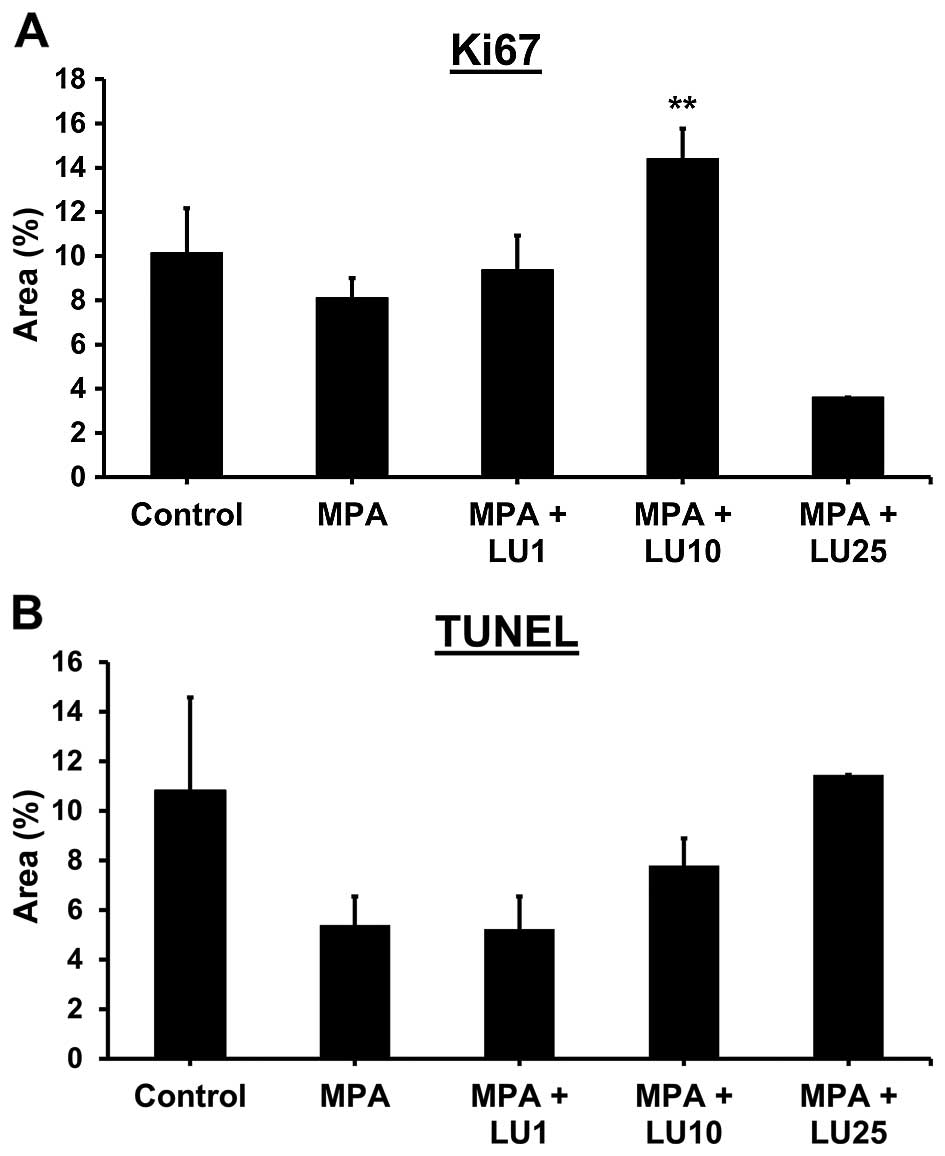
Luteolin increases expression of Ki67, a
marker of proliferation, in mammary tumor tissue
Levels of the proliferation marker Ki67 were
significantly higher within end-point tumor tissues derived from
animals given 10 mg/kg LU compared with either MPA alone, MPA + 1
mg/kg LU or controls (Fig. 6A).
Expression of Ki67 was reduced in tumors obtained from the group
given 25 mg/kg LU compared with either MPA alone or controls,
however, this effect was not statistically significant (Fig. 6A). In contrast, compared with
controls, levels of Ki67 were significantly higher in non-tumor
mammary gland tissue collected from animals given the lowest dose
of LU (1 mg/kg) but not in the other treatment groups (data not
shown).
TUNEL assays of tumor tissue demonstrated no
significant differences in levels of apoptosis between groups
treated with LU (Fig. 6B). However,
significantly less cell death occurred in non-tumor mammary gland
tissue obtained from animals administered 25 mg/kg LU, compared
with controls (data not shown). Taken together, these data suggest
that the reduced levels of cell death and increased proliferation
observed at the end of the study in some animals given LU may occur
in response to the lifting of selective inhibitory pressure on day
46 when LU injections were terminated.
Immunohistochemical analysis of tissue derived from
mammary tumors and contra-lateral non-tumor mammary glands showed
that in the latter, LU was unable to prevent and/or reverse the
formation of hyperplastic lesions arising in response to MPA
(please see images in Fig. 5) (data
not shown). These data suggest that LU exerts its effects and
prevents MPA-induced tumor development in breast tissue at a stage
subsequent to the formation of precancerous lesions.
Discussion
The consumption of combination HRT, which contains
both estrogen and progestin, puts millions of postmenopausal women
at higher risk of developing breast cancer compared with those
taking estrogen alone (2–5). It is therefore imperative that we
develop new and effective safe synthetic and/or naturally-occurring
compounds with antiprogestin activity that can be taken in
conjunction with combination HRT regimens to eliminate the
progestin-dependent increase in breast cancer risk. With this in
mind, we conducted studies to investigate the ability of the
flavonoid LU to act as a preventive compound in MPA-driven breast
cancer, given that LU has been shown to have anticarcinogenic
properties in other test systems (22,24).
Using an established progestin-dependent DMBA mammary tumor model
previously developed in our laboratory (17–19),
we determined that LU suppresses the development of
progestin-driven mammary tumors.
Surprisingly, when LU was used as a chemopreventive
agent, a biphasic response was observed, whereby both low and high
doses (1 and 25 mg/kg), but not an intermediate dose (10 mg/kg), of
the flavonoid decreased MPA-induced mammary tumor incidence and
increased mammary tumor latency. These observations resulted in a
non-monotonic U-shaped dose-response curve (35). It should be noted that treatment
with 10 mg/kg LU, in the absence of MPA, did not cause induction of
tumors or toxicity to animals (data not shown), indicating that the
flavonoid does not have any inherent tumor-stimulating properties.
Importantly, no adverse effects were observed with any of the doses
of LU used in these studies.
Having determined that LU is an effective means by
which to block the development of progestin-dependent tumors, we
sought to elucidate the mechanisms responsible by examining VEGF,
blood vessel density and Ki67 expression, as well as conducting
TUNEL assays in sections from mammary tumors and contralateral
inguinal mammary glands. Because LU treatment stopped two weeks
prior to the termination of the study, these results represent the
lasting effect of LU on the targeted tissues.
Tumor incidence was low in animals given 1 and 25
mg/kg LU, an observation which may be explained by a sustained loss
of VEGF within tumor tissue resulting in an inability of
preneoplastic lesions to form frank tumors, as previously observed
with apigenin treatment (19). It
is likely that loss of VEGF, which is potently angiogenic, resulted
in reduced blood vessel density in regressing tumors, indicating
disruption of tumor nourishment. Although LU caused a sustained
downregulation of tumor-associated VEGF that was independent of
dosage, it had no effect on VEGF levels in non-tumor mammary gland
tissue. The ability of LU to significantly reduce both VEGF
production and tumor volume in all three dosage groups (1, 10, and
25 mg/kg) is most likely explained by the flavonoid acting as both
an antiprogestin and possibly an estrogen/antiestrogen (23,29,36–38).
This proposed mechanism could explain why tumor volumes in the 10
mg/kg group were reduced even though the incidence of tumors in the
same group was high. Since LU did not influence progesterone
receptor levels (data not shown), its inhibitory effect may be due
to its ability to attenuate the post-ligand binding signal
transduction pathway normally known to promote VEGF production in
tumor cells. For example, hypoxia-inducible factor α (HIF-1α) plays
an important role in progesterone receptor-mediated VEGF induction
(39). Thus, inhibition or
downregulation of HIF-1α may suppress VEGF activity. It is also
possible that LU may modify the activity of progesterone receptor
post-transcriptionally, as shown by others (40). These possibilities remain to be
explored.
Ki67 and TUNEL measurements provided little insight
into the mechanism through which LU prevents tumor formation in the
MPA-driven DMBA-induced mammary tumor model. While neither Ki67 nor
TUNEL signals changed markedly in response to doses of 1 and 25
mg/kg LU, Ki67 expression was significantly increased in tumor
tissue obtained from animals given a dose of 10 mg/kg LU,
suggesting that in this group, circulating levels of LU caused
tumor cell proliferation, which in turn resulted in LU losing its
ability to control the formation of MPA-driven tumors. Such effects
of flavonoids have been reported previously (19,21).
It is likely that our inability to gain meaningful data for Ki67
and TUNEL assays is due to the time lag between cessation of LU
treatment and tumor collection. During this period when the
suppressive pressure of LU was removed, tumor cell proliferation
most likely increased, while apoptosis decreased. In future studies
we will address this time-lag by collecting tumors at the time LU
treatment is terminated. Suppression of VEGF expression and
consequent disruption of angiogenesis by LU may explain why these
tumors remained smaller compared with those under the influence of
MPA alone, even though tumor incidence was equivalent.
It is well known that progestins increase VEGF in
hormone-responsive tumor cells (12,17,19,41).
Recent evidence suggests that progestins not only provide a
microenvironment conducive to growth and metastasis, but that they
also enrich the tumor cell population (42,43).
It is therefore imperative that we improve the available
therapeutic anti-progestin options. The minimal increase in
tumor-associated VEGF observed in the MPA group in this study
likely occurred as a result of the length of time involved in the
experiment. Due to a large number of variously sized (both large
and small) tumors present in the first place, maximum tumor growth
may have already occurred prior to the end of the experiment,
influencing final VEGF levels. Alternatively, seasonal variations
may have caused fluctuations in levels of VEGF (44). Nevertheless, it is interesting to
note that LU brought about a persistent and significant reduction
in VEGF production within mammary tumors, and that suppression of
VEGF was independent of dosage. To our knowledge, this is the first
study of any lasting effect of LU (i.e., persistent inhibition of
VEGF) even when cessation of LU administration occurred at a point
well before the end of the experiment. These observations suggest
that LU may cause epigenetic changes in the VEGF gene, though this
remains to be determined. Importantly, these findings indicate that
LU has the ability to prevent tumors from establishing a
microenvironment conducive to growth. Our observations also suggest
that LU may suppress cancer stem cells since tumor incidence
remained low even when LU supplementation was discontinued. Thus
in vitro studies to examine the effects of LU on the
self-renewal properties of stem-like cells are warranted.
Considering that treatment with 25 mg/kg LU was nontoxic, resulted
in the lowest total number of tumors (3 tumors in the 25 mg/kg
group vs. 4 tumors in the 1 mg/kg group), and largely counteracted
the effects of MPA on DMBA-induced mammary cancer by reducing both
tumor volume and incidence, we propose that the flavonoid should be
further evaluated as a naturally-occurring chemopreventive
compound. LU possesses important antitumor properties that may well
be extremely advantageous to women who are either undergoing
combination HRT or who have already been exposed to this type of
therapy. Further studies are justified to elucidate fully the
effects of LU in vivo and to gain a better understanding of
its potential for human use. In future animal studies, the effects
of orally administered LU should be examined, in order to determine
the preventive properties of the flavonoid when ingested as a
dietary supplement. Such studies would serve as a means of
assessing its potential use in humans.
Acknowledgments
The present study was supported by a peer-reviewed
COR award from the University of Missouri College of Veterinary
Medicine (Columbia, MO) and by funds from generous donors to the
Ellis Fischel Cancer Center, University of Missouri (Columbia, MO).
We would like to thank Mr Jason Lee for help with the figures.
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chlebowski RT, Hendrix SL, Langer RD,
Stefanick ML, Gass M, Lane D, Rodabough RJ, Gilligan MA, Cyr MG,
Thomson CA, et al WHI Investigators: Influence of estrogen plus
progestin on breast cancer and mammography in healthy
postmenopausal women: The Women's Health Initiative Randomized
Trial. JAMA. 289:3243–3253. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Beral V; Million Women Study
Collaborators: Breast cancer and hormone-replacement therapy in the
Million Women Study. Lancet. 362:419–427. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rossouw JE, Anderson GL, Prentice RL,
LaCroix AZ, Kooperberg C, Stefanick ML, Jackson RD, Beresford SA,
Howard BV, Johnson KC, et al Writing Group for the Women's Health
Initiative Investigators: Risks and benefits of estrogen plus
progestin in healthy postmenopausal women: Principal results from
the Women's Health Initiative randomized controlled trial. JAMA.
288:321–333. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fahlén M, Fornander T, Johansson H,
Johansson U, Rutqvist LE, Wilking N and von Schoultz E: Hormone
replacement therapy after breast cancer: 10 year follow up of the
Stockholm randomised trial. Eur J Cancer. 49:52–59. 2013.
View Article : Google Scholar
|
|
6
|
Horn LC, Schnurrbusch U, Bilek K,
Hentschel B and Einenkel J: Risk of progression in complex and
atypical endometrial hyperplasia: Clinicopathologic analysis in
cases with and without progestogen treatment. Int J Gynecol Cancer.
14:348–353. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pike MC, Peters RK, Cozen W, Probst-Hensch
NM, Felix JC, Wan PC and Mack TM: Estrogen-progestin replacement
therapy and endometrial cancer. J Natl Cancer Inst. 89:1110–1116.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Brisken C: Progesterone signalling in
breast cancer: A neglected hormone coming into the limelight. Nat
Rev Cancer. 13:385–396. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gonzalez-Suarez E, Jacob AP, Jones J,
Miller R, Roudier-Meyer MP, Erwert R, Pinkas J, Branstetter D and
Dougall WC: RANK ligand mediates progestin-induced mammary
epithelial proliferation and carcinogenesis. Nature. 468:103–107.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hyder SM, Murthy L and Stancel GM:
Progestin regulation of vascular endothelial growth factor in human
breast cancer cells. Cancer Res. 58:392–395. 1998.PubMed/NCBI
|
|
11
|
Liang Y, Besch-Williford C, Brekken RA and
Hyder SM: Progestin-dependent progression of human breast tumor
xenografts: A novel model for evaluating antitumor therapeutics.
Cancer Res. 67:9929–9936. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liang Y, Benakanakere I, Besch-Williford
C, Hyder RS, Ellersieck MR and Hyder SM: Synthetic progestins
induce growth and metastasis of BT-474 human breast cancer
xenografts in nude mice. Menopause. 17:1040–1047. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liang Y, Wu J, Stancel GM and Hyder SM:
p53-dependent inhibition of progestin-induced VEGF expression in
human breast cancer cells. J Steroid Biochem Mol Biol. 93:173–182.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liang Y and Hyder SM: Proliferation of
endothelial and tumor epithelial cells by progestin-induced
vascular endothelial growth factor from human breast cancer cells:
Paracrine and autocrine effects. Endocrinology. 146:3632–3641.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Joshi PA, Jackson HW, Beristain AG, Di
Grappa MA, Mote PA, Clarke CL, Stingl J, Waterhouse PD and Khokha
R: Progesterone induces adult mammary stem cell expansion. Nature.
465:803–807. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schramek D, Leibbrandt A, Sigl V, Kenner
L, Pospisilik JA, Lee HJ, Hanada R, Joshi PA, Aliprantis A,
Glimcher L, et al: Osteoclast differentiation factor RANKL controls
development of progestin-driven mammary cancer. Nature. 468:98–102.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Benakanakere I, Besch-Williford C, Schnell
J, Brandt S, Ellersieck MR, Molinolo A and Hyder SM: Natural and
synthetic progestins accelerate
7,12-dimethylbenz[a]anthracene-initiated mammary tumors and
increase angiogenesis in Sprague-Dawley rats. Clin Cancer Res.
12:4062–4071. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Benakanakere I, Besch-Williford C, Carroll
CE and Hyder SM: Synthetic progestins differentially promote or
prevent 7,12-dimethylbenz(a)anthracene-induced mammary tumors in
Sprague-Dawley rats. Cancer Prev Res (Phila). 3:1157–1167. 2010.
View Article : Google Scholar
|
|
19
|
Mafuvadze B, Benakanakere I, López Pérez
FR, Besch-Williford C, Ellersieck MR and Hyder SM: Apigenin
prevents development of medroxyprogesterone acetate-accelerated
7,12-dimethylbenz(a) anthracene-induced mammary tumors in
Sprague-Dawley rats. Cancer Prev Res (Phila). 4:1316–1324. 2011.
View Article : Google Scholar
|
|
20
|
Mafuvadze B, Liang Y, Besch-Williford C,
Zhang X and Hyder SM: Apigenin induces apoptosis and blocks growth
of medroxyprogesterone acetate-dependent BT-474 xenograft tumors.
Horm Cancer. 3:160–171. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mafuvadze B, Cook M, Xu Z, Besch-Williford
CL and Hyder SM: Effects of dietary apigenin on tumor latency,
incidence and multiplicity in a medroxyprogesterone
acetate-accelerated 7,12-dimethylbenz(a)anthracene-induced breast
cancer model. Nutr Cancer. 65:1184–1191. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lin Y, Shi R, Wang X and Shen HM:
Luteolin, a flavonoid with potential for cancer prevention and
therapy. Curr Cancer Drug Targets. 8:634–646. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
López-Lázaro M: Distribution and
biological activities of the flavonoid luteolin. Mini Rev Med Chem.
9:31–59. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Seelinger G, Merfort I, Wölfle U and
Schempp CM: Anti-carcinogenic effects of the flavonoid luteolin.
Molecules. 13:2628–2651. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bagli E, Stefaniotou M, Morbidelli L,
Ziche M, Psillas K, Murphy C and Fotsis T: Luteolin inhibits
vascular endothelial growth factor-induced angiogenesis; inhibition
of endothelial cell survival and proliferation by targeting
phosphatidylinositol 3′-kinase activity. Cancer Res. 64:7936–7946.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pratheeshkumar P, Son YO, Budhraja A, Wang
X, Ding S, Wang L, Hitron A, Lee JC, Kim D, Divya SP, et al:
Luteolin inhibits human prostate tumor growth by suppressing
vascular endothelial growth factor receptor 2-mediated
angiogenesis. PLoS One. 7:e522792012. View Article : Google Scholar
|
|
27
|
Selvendiran K, Koga H, Ueno T, Yoshida T,
Maeyama M, Torimura T, Yano H, Kojiro M and Sata M: Luteolin
promotes degradation in signal transducer and activator of
transcription 3 in human hepatoma cells: An implication for the
antitumor potential of flavonoids. Cancer Res. 66:4826–4834. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Luo H, Jiang BH, King SM and Chen YC:
Inhibition of cell growth and VEGF expression in ovarian cancer
cells by flavonoids. Nutr Cancer. 60:800–809. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nordeen SK, Bona BJ, Jones DN, Lambert JR
and Jackson TA: Endocrine disrupting activities of the flavonoid
nutraceuticals luteolin and quercetin. Horm Cancer. 4:293–300.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fang J, Zhou Q, Shi XL and Jiang BH:
Luteolin inhibits insulin-like growth factor 1 receptor signaling
in prostate cancer cells. Carcinogenesis. 28:713–723. 2007.
View Article : Google Scholar
|
|
31
|
Zhou Q, Yan B, Hu X, Li XB, Zhang J and
Fang J: Luteolin inhibits invasion of prostate cancer PC3 cells
through E-cadherin. Mol Cancer Ther. 8:1684–1691. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Samy RP, Gopalakrishnakone P and
Ignacimuthu S: Anti-tumor promoting potential of luteolin against
7,12-dimethylbenz(a) anthracene-induced mammary tumors in rats.
Chem Biol Interact. 164:1–14. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Faustino-Rocha A, Oliveira PA,
Pinho-Oliveira J, Teixeira-Guedes C, Soares-Maia R, da Costa RG,
Colaço B, Pires MJ, Colaço J, Ferreira R, et al: Estimation of rat
mammary tumor volume using caliper and ultrasonography
measurements. Lab Anim (NY). 42:217–224. 2013. View Article : Google Scholar
|
|
34
|
Tomayko MM and Reynolds CP: Determination
of subcutaneous tumor size in athymic (nude) mice. Cancer Chemother
Pharmacol. 24:148–154. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Vandenberg LN, Colborn T, Hayes TB,
Heindel JJ, Jacobs DR Jr, Lee DH, Shioda T, Soto AM, vom Saal FS,
Welshons WV, et al: Hormones and endocrine-disrupting chemicals:
Low-dose effects and nonmonotonic dose responses. Endocr Rev.
33:378–455. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Markaverich BM, Shoulars K and Rodriguez
MA: Luteolin regulation of estrogen signaling and cell cycle
pathway genes in MCF-7 human breast cancer cells. Int J Biomed Sci.
7:101–111. 2011.PubMed/NCBI
|
|
37
|
Le Bail JC, Varnat F, Nicolas JC and
Habrioux G: Estrogenic and antiproliferative activities on MCF-7
human breast cancer cells by flavonoids. Cancer Lett. 130:209–216.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zand RSR, Jenkins DJ and Diamandis EP:
Steroid hormone activity of flavonoids and related compounds.
Breast Cancer Res Treat. 62:35–49. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Carroll CE, Liang Y, Benakanakere I,
Besch-Williford C and Hyder SM: The anticancer agent YC-1
suppresses progestin-stimulated VEGF in breast cancer cells and
arrests breast tumor development. Int J Oncol. 42:179–187.
2013.
|
|
40
|
Knutson TP and Lange CA: Tracking
progesterone receptor-mediated actions in breast cancer. Pharmacol
Ther. 142:114–125. 2014. View Article : Google Scholar :
|
|
41
|
Hyder SM, Chiappetta C and Stancel GM:
Pharmacological and endogenous progestins induce vascular
endothelial growth factor expression in human breast cancer cells.
Int J Cancer. 92:469–473. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cittelly DM, Finlay-Schultz J, Howe EN,
Spoelstra NS, Axlund SD, Hendricks P, Jacobsen BM, Sartorius CA and
Richer JK: Progestin suppression of miR-29 potentiates
dedifferentiation of breast cancer cells via KLF4. Oncogene.
32:2555–2564. 2013. View Article : Google Scholar
|
|
43
|
Horwitz KB, Dye WW, Harrell JC, Kabos P
and Sartorius CA: Rare steroid receptor-negative basal-like
tumorigenic cells in luminal subtype human breast cancer
xenografts. Proc Natl Acad Sci USA. 105:5774–5779. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Löscher W, Mevissen M and Häussler B:
Seasonal influence on 7,12-dimethylbenz[a]anthracene-induced
mammary carcinogenesis in Sprague-Dawley rats under controlled
laboratory conditions. Pharmacol Toxicol. 81:265–270. 1997.
|