Introduction
Hypoxia within solid tumors may have a profound
impact in malignant progression. Hypoxia within the tumor mass is
an independent marker of poor prognosis for patients with a variety
of cancers including cervical cancers, soft tissue sarcoma and head
neck cancers (1–3). Hypoxic stress underlies a number of
biologically important processes in which cellular migration and
invasion occur (4). Hypoxia may
play an important role in promoting tumor metastasis and invasion
into the extracellular matrix (5).
Hypoxia upregulates the expression of the urokinase plasminogen
activator receptor (uPAR) which is a component of a protease system
implicated in tumor invasion and metastasis (6). The uPAR is activated by binding with
its ligand, urokinase plasminogen activator (uPA), allowing the
conversion of inactive plasminogen into plasmin that in turn can
degrade extracellular matrix proteins, facilitating invasion by
tumor cells (7).
Hypoxia-inducible factor (HIF)-1 is a heterodimeric
transcriptional complex that plays a pivotal role in the regulation
of cellular utilization of oxygen and is an essential regulator of
angiogenesis in solid tumor and ischemic disorders. HIF-1 is
composed of the bHLH-PAS proteins HIF-1α and aryl hydrocarbon
receptor nuclear translocator (ARNT). HIF-1 mediates the
transcriptional response to oxygen deprivation by binding to HIF-1
response elements (HRE) within the promoters or enhancers of genes
involved in glycolysis, glucose transport, erythropoiesis and
angiogenesis (8,9). The HIF-1 activity is critical for
cancer development and HIF-1 is essential for proliferation,
survival or differentiation of multiple tumor tissues (9). In light of the above considerations,
we investigated how uPAR expression is induced by hypoxia in
cervical cancer cell lines. We also sought to determine the
regulator of uPAR expression during hypoxia. HIF-1 is a possible
candidate for the regulator of uPAR expression.
Materials and methods
Samples
The samples were obtained from patients undergoing
surgery or biopsy at Tokyo Medical University Hospital with
informed consent. We used ten normal cervices and 35 cervical
cancers. To extract total RNA, the tissues were finely minced into
small pieces with scissors, washed in phosphate-buffered saline
(PBS), snap-frozen and stored at −80°C. For immunohistochemistry
analysis, the tissues were formalin-fixed and embedded in
paraffin.
Cell culture
The human cervical cancer cell lines, CaSki and CA,
were maintained in Modified Eagle Medium (MEM; Invitrogen,
Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (FBS).
For hypoxic exposure, these cells (1.0×107 cells) were
plated in 10 ml of medium in 100-mm dishes and incubated overnight
in 1% O2 in a water-jacketed CO2 incubator
(NAPCO, Winchester, VA, USA) at 37°C in a humidified atmosphere
with 5% CO2 for indicated times. In experiments using
transfected cells, each transfection was performed before exposure
to hypoxia.
DNA plasmids
Sequence analysis of the human uPAR promoter
revealed one putative binding site for HIF-1, the HRE motifs
5′-BACGTSSK-3′. The putative HIF binding site at position −34 to
−39 was identified on the sense strand (10). Luciferase reporter constructs,
puPAR-141-luc, containing the uPAR promoter which is produced by
PCR were prepared by ligation into pGL3-basic vector (Fig. 3A). We prepared the puPAR-141mt-luc,
where the putative HRE is mutated by replacement of ACG with TTT
using PCR-based site-specific mutagenesis. HIF-1α and ARNT
constructs were kindly provided by Dr L. Eric Huang (Departments of
Neurosurgery and Oncological Sciences, University of Utah,
USA).
Invasion assays
For invasion assays, 12-mm-diameter Transwell
polycarbonate filters (12-mm pore size; Costar) in a modified
Boyden chamber were coated with Matrigel (100 ml; Sigma, St. Louis,
MO, USA) at 1:20 dilution in serum-free medium and air-dried for 24
h. Cells (5×104) in the complete medium (200 ml) were
seeded into the inner chamber. A total of 600 ml of the medium was
added to the lower chamber, and the plate was incubated at 37°C in
a 5% CO2/95% air incubator (20% O2). For
hypoxic treatment, plates containing Boyden chambers were placed in
1% O2 in a water-jacketed CO2 incubator
(NAPCO) at 37°C in a humidified atmosphere with 5% CO2
overnight. Cells on the lower surface of the filter were scraped
with a rubber scraper into the medium from the lower chamber,
pelleted, resuspended in the medium (50 ml), and counted using a
hemocytometer. Each condition was performed in quadruplicates, and
the experiments were carried out twice.
Real-time RT-PCR analysis
Total RNA was isolated using Isogen reagent (Nippon
Gene, Tokyo, Japan) and quantified by A260/A280 measurement using
an Ultraspec 3000 (Amersham Biosciences, Piscataway, NJ, USA).
Total RNA (50 mg) was reverse-transcribed into cDNA using an RT-PCR
kit (Stratagene, La Jolla, CA, USA) according to the manufacture's
recommendations.
Real-time PCR was performed for the quantitative
estimation of the DNA level. PCR reactions (20 ml) were set-up with
the final concentrations of 5 mM MgCl2, SYBR-Green
Master Mix (2 ml; Roche Applied Science, Mannheim, Germany), cDNA
(1:10 dilution; 5 ml), and forward and reverse primers (0.3 mM
each). The primer sequences were as follows:
5′-CAACACCACCAAATGCAACG-3′ (forward), and
5′-GGTTTTTCGGTTCGTGAGTG-3′ (reverse). The reactions were then
cycled in a LightCycler (Roche Applied Science) with the following
parameters: denaturation for 1 cycle at 95°C for 10 sec, 45 cycles
(temperature transition of 20°C/sec) of 95°C for 10 sec, 62°C for
10 sec, and 72°C for 6 sec. Fluorescence readings were taken at
72°C and, melting curve analysis was performed with continuous
fluorescence reading. The LightCycler software generated a standard
curve (from measurements taken during the exponential phase of the
amplification) that enabled the amount of each gene in each test
sample to be determined.
Western blot analysis
CaSki and CA cells were seeded at 2.5×106
cells/150 mm dish, and incubated overnight at 37°C. Subsequently,
cells were exposed to hypoxic conditions. Cells were harvested and
lysed on ice for 30 min. In lysis buffer [10 mM Tris (pH 8.0), 1 mM
EDTA, 400 mM NaCl, 10% glycerol, 0.5% NP40, 5 mM sodium fluoride,
0.1 mM phenylmetylsulphonyl fluoride and 1 mM DTT], containing
complete protease inhibitor cocktail (Boehringer Mannheim,
Indianapolis, IN, USA). The lysate was centrifuged at 14,000 rpm
for 15 min and the soluble fractions were collected. Protein
concentrations were measured using Bio-Rad protein assay kit
(Bio-Rad Laboratories, Hercules, CA, USA). Equal amounts of protein
(40 mg) were loaded onto a 4–12% SDS-polyacrylamide gel and
subjected to electrophoresis at 200 V for 50 min. The protein was
transferred onto a polyviniylidene difluoride membrane and probed
with an anti-uPAR antibody (FL-290; Santa Cruz Biotechnology, Santa
Cruz, CA, USA), anti-HIF-1α antibody (NB100-105; Novus Biologicals,
Littleton, CO, USA), and anti-actin antibody (C4; Boehringer
Mannheim). The same blot was probed with different antibodies after
stripping the membrane. Each protein was detected by horseradish
peroxidase-conjugated secondary antibody coupled with enhanced
chemiluminescence western blotting detection reagents (Amersham
Biosciences). Each band intensity was normalized to the intensity
of the actin band.
Transfection and luciferase assays
Each cell line was seeded at 5.0×105
cells/35-mm dish and incubated overnight at 37°C in a 5%
CO2 incubator. For each transfection, empty or
expression vectors (1.0 mg) along with the promoter-luciferase DNA
(0.3 mg) were mixed in Opti-MEM (0.2 ml) and a precipitate was
formed using Lipofectamine 2000 (both from Invitrogen) according to
the manufacturer's instructions. Cells were washed with Opti-MEM,
and complexes were applied to the cells. After transfection for 24
h, cells were harvested and extracts were prepared with the Glo
Lysis Buffer (Promega, Madison, WI, USA). Luciferase activity was
measured in extracts from triplicate samples using the Bright-Glo
Luciferase Assay system (Promega).
Electrophoretic mobility shift
assays
The HIF-1 protein was synthesized in vitro in
the presence of unlabeled amino acids using the pcDNA3-HIF-1α and
pcDNA3-ARNT expression constructs with the coupled
transcription/translation system (TNT) from Promega (11). Translated products were analyzed by
western blotting using anti-HIF-1α antibody (NB100-105; Novus
Biologicals).
Oligonucleotides containing the HIF-1 consensus
DNA-binding site from uPAR promoter (HRE/uPAR) and the HIF-1
binding site from the erythropoietin promoter (HRE) were purchased
as single-stranded DNAs from Genosys Biotechnologies (Woodlands,
TX, USA). Double-stranded oligonucleotides were prepared by
annealing complementary oligonucleotides, in a buffer containing 10
mM Tris (pH 8.0), 500 mM NaCl and 1 mM EDTA. The sequences of the
complementary pairs were as follows:
5′-AAGGAGAGAAGACGTGCAGGGACCCC-3′ and
5′-GGGGTCCCTGCACGTCTTCTCTCCTT-3′ (HRE/uPAR);
5′-TCTGTACGTGACCACACTCACCTC-3′ and 5′-GAGGTGAGTGTGGTCACGTACAGA-3′
(HRE). Equimolar amounts of the complementary oligonucleotides were
mixed in a 1.5-ml microcentrifuge tube and placed in a heat block
at 95°C. The heat block was allowed to cool to room temperature,
and the samples were desalted on a G-25 Microspin column
(Amersham). The double-stranded oligo-nucleotides were end-labeled
with 32P using T4 polynucleitide kinase and
[γ-32P]-ATP. For electrophoretic mobility shift
analysis, end-labeled double stranded oligonucleotides, 5,000 cpm,
were incubated with HIF-1 protein (2 ml) prepared by in
vitro transcription/translation at room temperature (22°C) for
30 min in the presence of a binding buffer containing 10% glycerol,
20 mM HEPES (pH 7.5), 25 mM KCl, 2 mM DTT, 2 mM MgCl2,
0.4 % NP-40 and 1 mg sheared salmon sperm DNA. When competition
assays were performed, an unlabeled HIF-1 consensus sequence
oligonucleotide was incubated with proteins and buffer for 5 min
before the addition of each labeled oligonucleotide. For supershift
assays, 0.5 mg of HIF-1α antibody (OZ15; Lab Vision Corporation,
Fremont, CA, USA) was incubated with the binding mixtures for 5 min
before the addition of the labeled oligonucleotide. Samples (20 ml)
were loaded onto a 5% non-denaturing polyacrylamide gel and
subjected to electrophoresis at 150 V for 1 h using 0.5X
Tris-borate EDTA [1X Tris-borate EDTA: 89 mM Tris, 8 mM boric asid
and 2 mM EDTA (pH 8.3)] as running buffer. After electrophoresis,
gels were transferred to Whatman 3 MM paper and exposed to Kodak
XAR film with intensifying screens at −80°C.
siRNA transfection
Gene silencing was achieved by transient
transfection of siRNA oligonucleotides encoding specific sequences
for HIF-1α according to the manufacturer's instructions (Santa Cruz
Biotechnology). In brief, CaSki and CA were plated overnight in
antibiotic-free medium to reach 50% confluency. Cells were
transfected the following morning using Lipofectamine 2000 with 100
pmol/ml of siRNA oligonucleotides. Control siRNA was transfected to
exclude non-specific effects. After exposure to 1% O2
for 24 h, cells were harvested for protein.
Immunohistopathological analysis
Immunohistochemistry was performed on representative
formalin-fixed, paraffin embedded sections. Each tissue block was
sectioned at 4-mm thickness, deparaffinized in xylene and
rehydrated in graded alcohols. Primary antibodies against uPAR
(CD87, clone HD-uPAR-13.1; 1:50 dilution; American Diagnostica,
Stamford, CT, USA) and HIF-1α (NB100-105, clone H1alpha67; 1:500
dilution; Novus Biologicals) were used. Pretreatment for unmasking
of antigens was carried out either via protease XXIV (Sigma
Chemicals, Perth, WA, USA) for 10 min at room temperature for uPAR,
or by autoclaving for 10 min at 110°C for HIF-1α.
Results
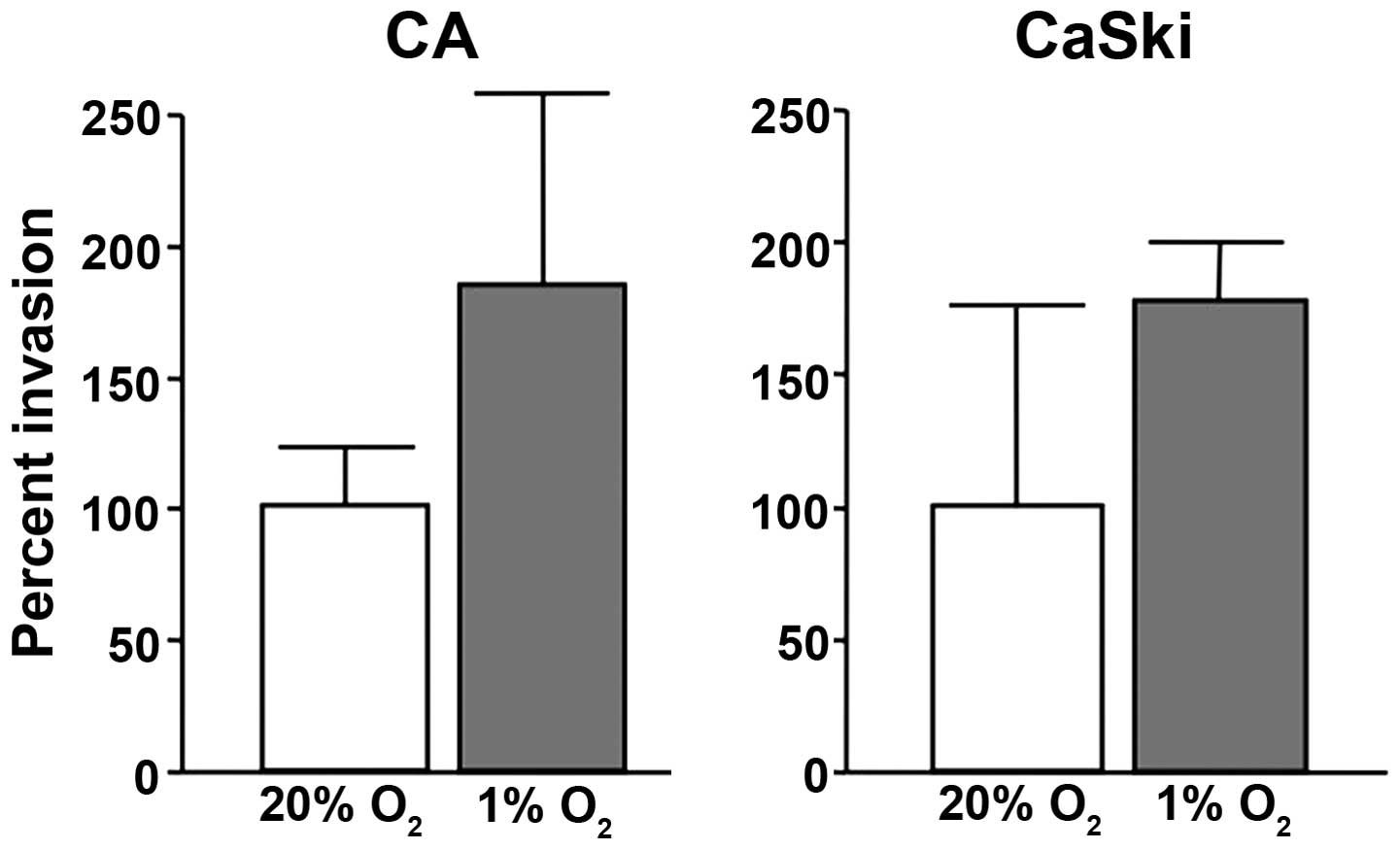
Hypoxia mediates invasion
To determine the functional correlates of
hypoxia-induced uPAR expression and invasion, we first compared the
invasion of cervical cancer cells through a reconstituted basement
membrane (Matrigel). The cervical cancer cell line, CaSki and CA,
were incubated in hypoxic (1% O2) and normoxic
conditions (20% O2), respectively. Invasion assays
showed that these cells were 2-fold more invasive under hypoxic
conditions than under normoxic conditions (Fig. 1).
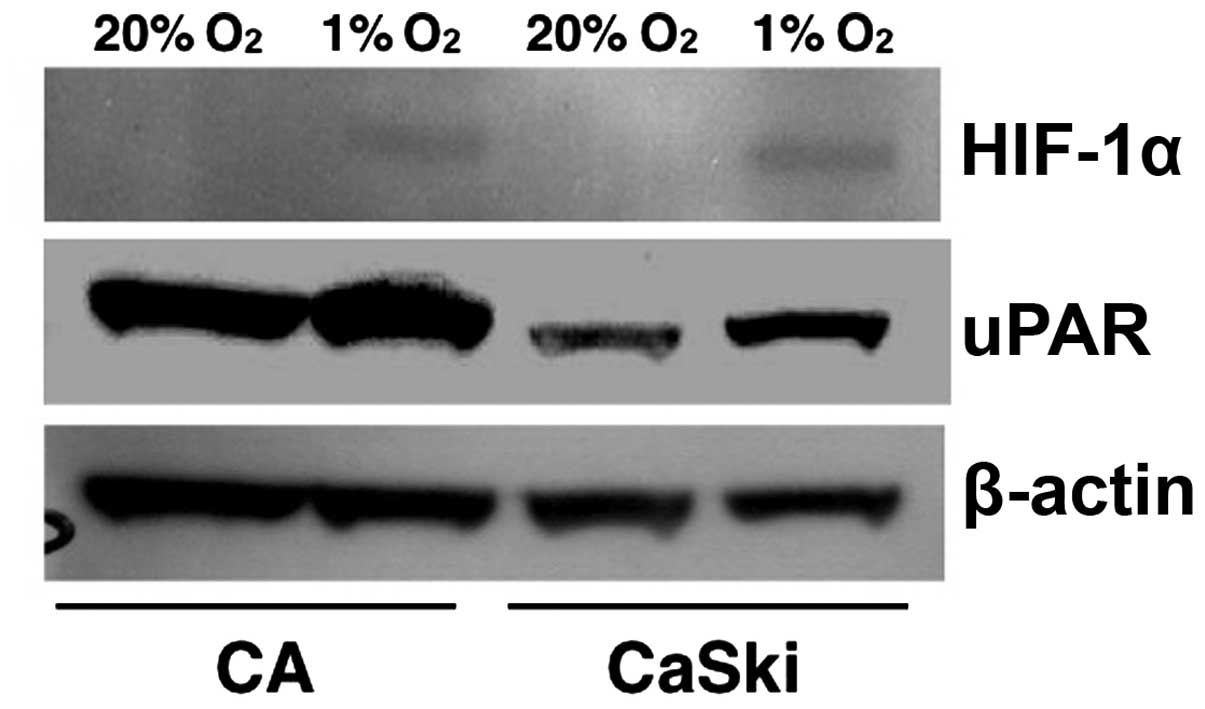
Hypoxia induces endogenous uPAR
expression
To analyze whether there is a correlation between
HIF-1α and uPAR, we examined the effect of hypoxia on HIF-1α and
uPAR expression in cervical cancer cell line, CaSki and CA. Whole
cell extracts prepared from controls and from cells exposed to 1%
O2 for 24 h were subjected to western blot analysis. As
shown in Fig. 2, both HIF-1α and
uPAR protein expressions were increased significantly under hypoxic
conditions. These results indicate that hypoxia induces the
endogenous expressions of uPAR.
HIF-1 transactivates the uPAR
promoter
To examine whether the induction of uPAR expression
by hypoxia is mediated via a transcriptional mechanism, CaSki and
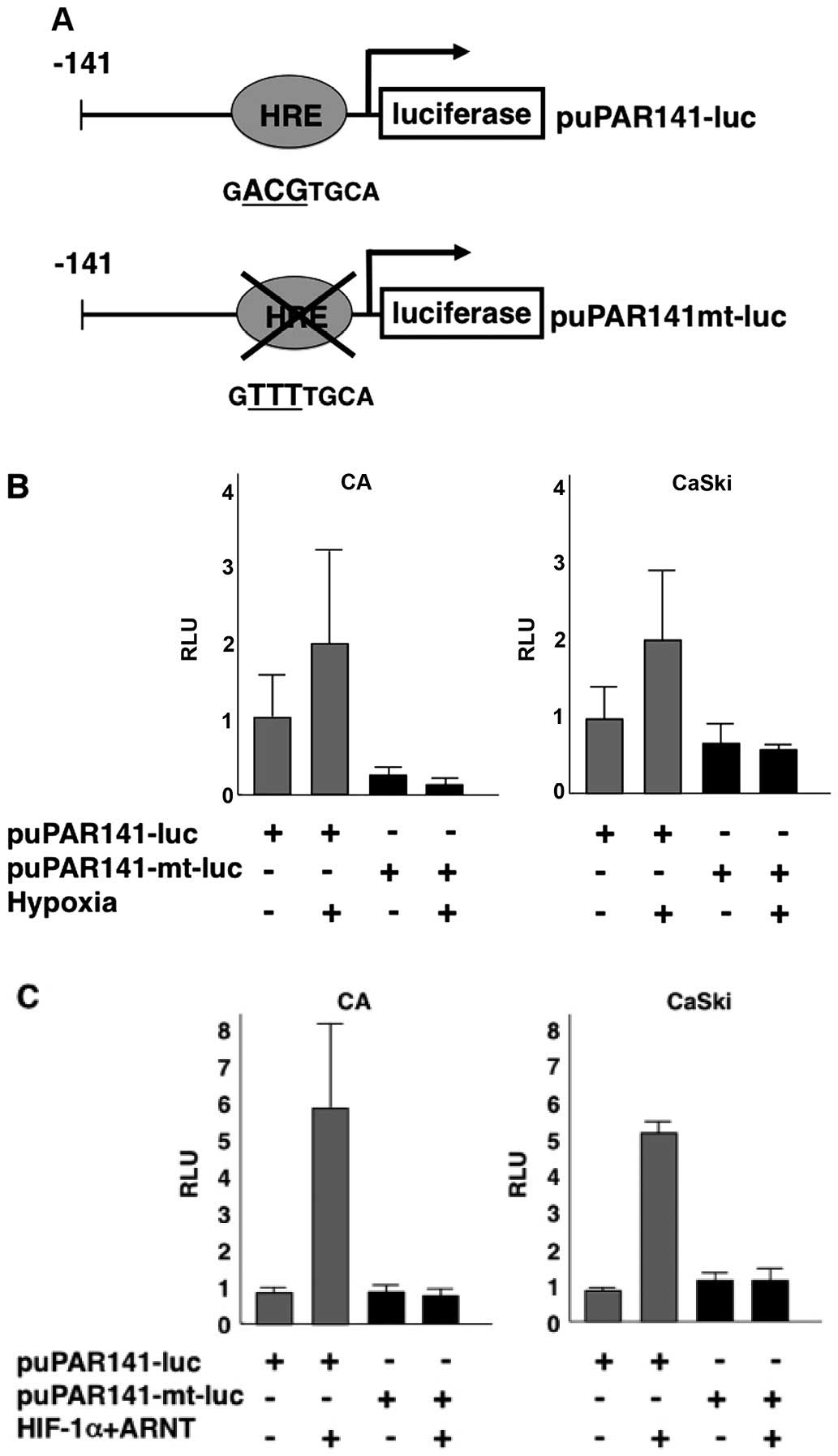
CA cells were transfected with puPAR-141-luc (Fig. 3A) and then exposed to 1%
O2 for 24 h. Hypoxia transactivated the uPAR promoter by
2-fold (Fig. 3B). Additional
cotransfection experiments were performed with puPAR-141-luc,
HIF-1α and ARNT or empty vectors. HIF-1 overexpression
transactivated the uPAR promoter by 5–6 fold (Fig. 3C).
The uPAR promoter contains a putative HRE. To
confirm whether the uPAR promoter regions are responsible for
transactivation by HIF-1, we prepared a reporter construct,
puPAR-141mt-luc, where the putative HRE is mutated by replacement
of AGG with TTT (Fig. 3A). Neither
hypoxia nor HIF-1 overexpression increased the activity of this
mutated promoter, indicating that this putative HRE (HRE/uPAR) is
essential for the regulation of uPAR by hypoxia and HIF-1 (Fig. 3B and C).
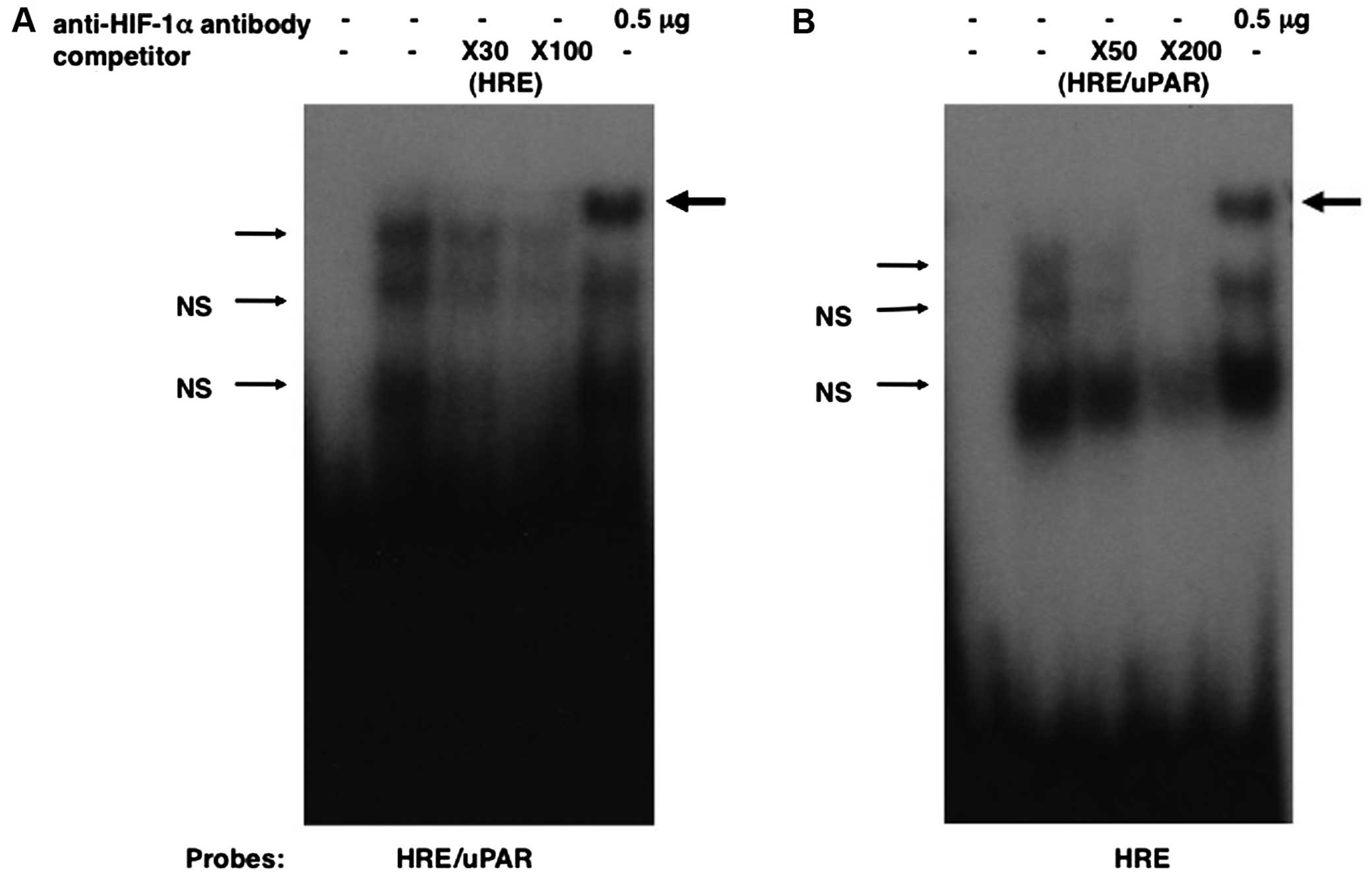
HIF-1 interacts with the putative HRE in
the uPAR promoter
To determine whether HIF-1 has direct interaction
with the HRE on the uPAR promoter, electrophoretic mobility shift
assays were performed. Oligonucleotide corresponding to nucleotides
−50 to −25 (HRE/uPAR) of the uPAR promoter was incubated with
HIF-1α and ARNT prepared by in vitro
transcription/translation and subjected to electrophoresis. A
DNA-protein complex was formed when HRE/uPAR was incubated with the
HIF-1-programmed rabbit reticulocyte lysate (Fig. 4A), but not with the unprogrammed
lysate (data not shown). This complex was specifically retarded by
anti-HIF-1α antibody (Fig. 4). The
addition of a 30-fold molar excess of cold HIF-1 consensus
oligonucleotide (HRE) markedly reduced binding (Fig. 4A). To additionally substantiate a
HIF-1 binding to this oligonucleotide, we performed competition
assays using end-labeled HIF-1 consensus oligonucleotide and cold
HRE/uPAR as a competitor. The unlabeled HRE/uPAR competed with
HIF-1 binding to the wild-type probe when present in the reaction
at 50- and 200-fold molar excess (Fig.
4B). These results confirm that HIF-1 binds to the HRE/uPAR and
indicate that the affinity of HIF-1 binding to the HRE/uPAR is
similar to that of the consensus HIF-1 sequence.
These data indicate that hypoxia-induced uPAR
expression is induced by the enhanced activity of HIF-1 on the
HRE/uPAR promoter. Taken together, the putative HIF-1 binding site
in the proximal region of the uPAR promoter is essential for the
regulation of uPAR.
Disruption of the uPAR expression by
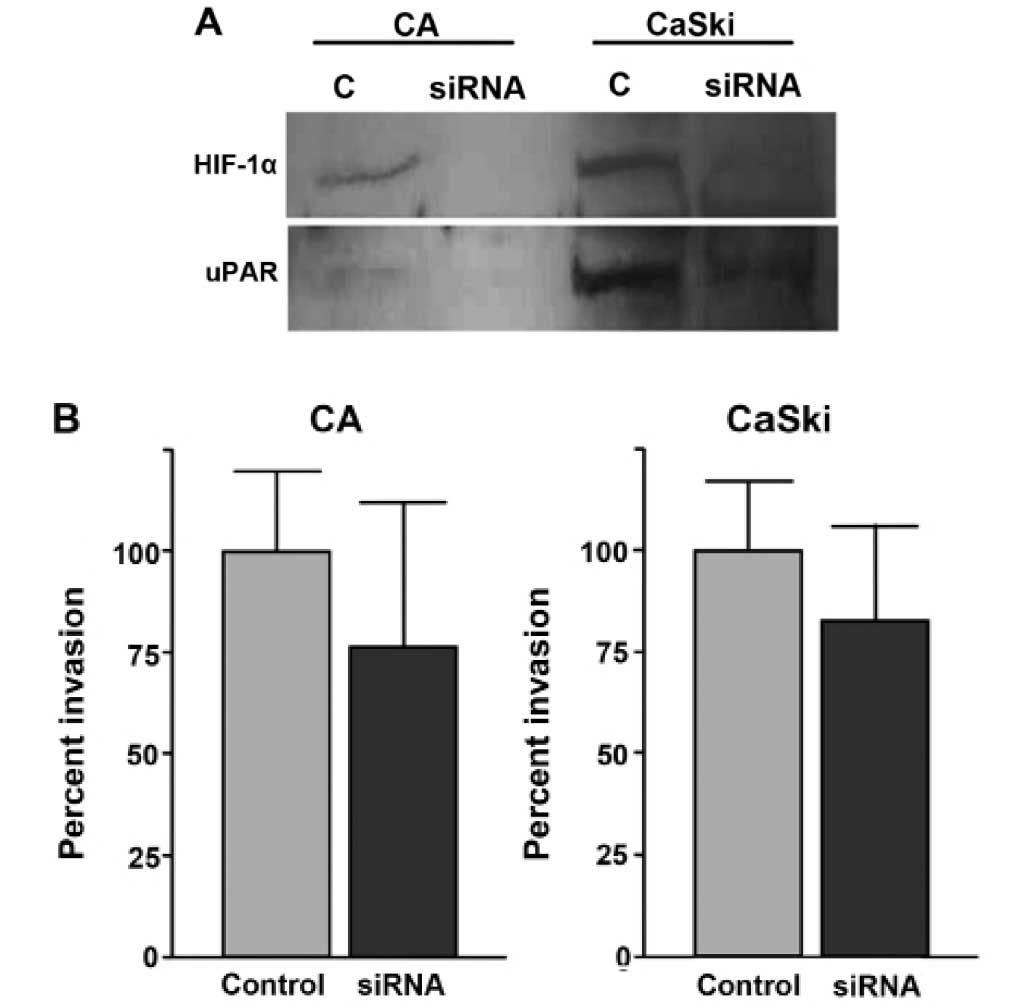
HIF-1 siRNA
HIF-1α siRNA was previously used to inhibit HIF-1α
expression (12). In the present
study, we used this HIF-1α siRNA to examine whether hypoxia-induced
uPAR expression is mediated by HIF-1 upregulation. When CaSki cells
were transfected with siRNA and exposed to hypoxic conditions, the
HIF-1α and uPAR expressions were downregulated compared with the
control siRNA transfected cells (Fig.
5A). The level of actin was monitored as a control and was
found to be unaltered by siRNA (Fig.
5A). Similar results were obtained using CA cells (Fig. 5A). These results indicate that
HIF-1α is required for hypoxia induced uPAR expression.
To examine the effect on invasiveness by HIF-1α
knockdown, we performed invasion assay in CaSki and CA cells after
HIF-1α siRNA tranfection. Downregulation of HIF-1 expression
significantly diminishes invasion in cervical cancer cells
(Fig. 5).
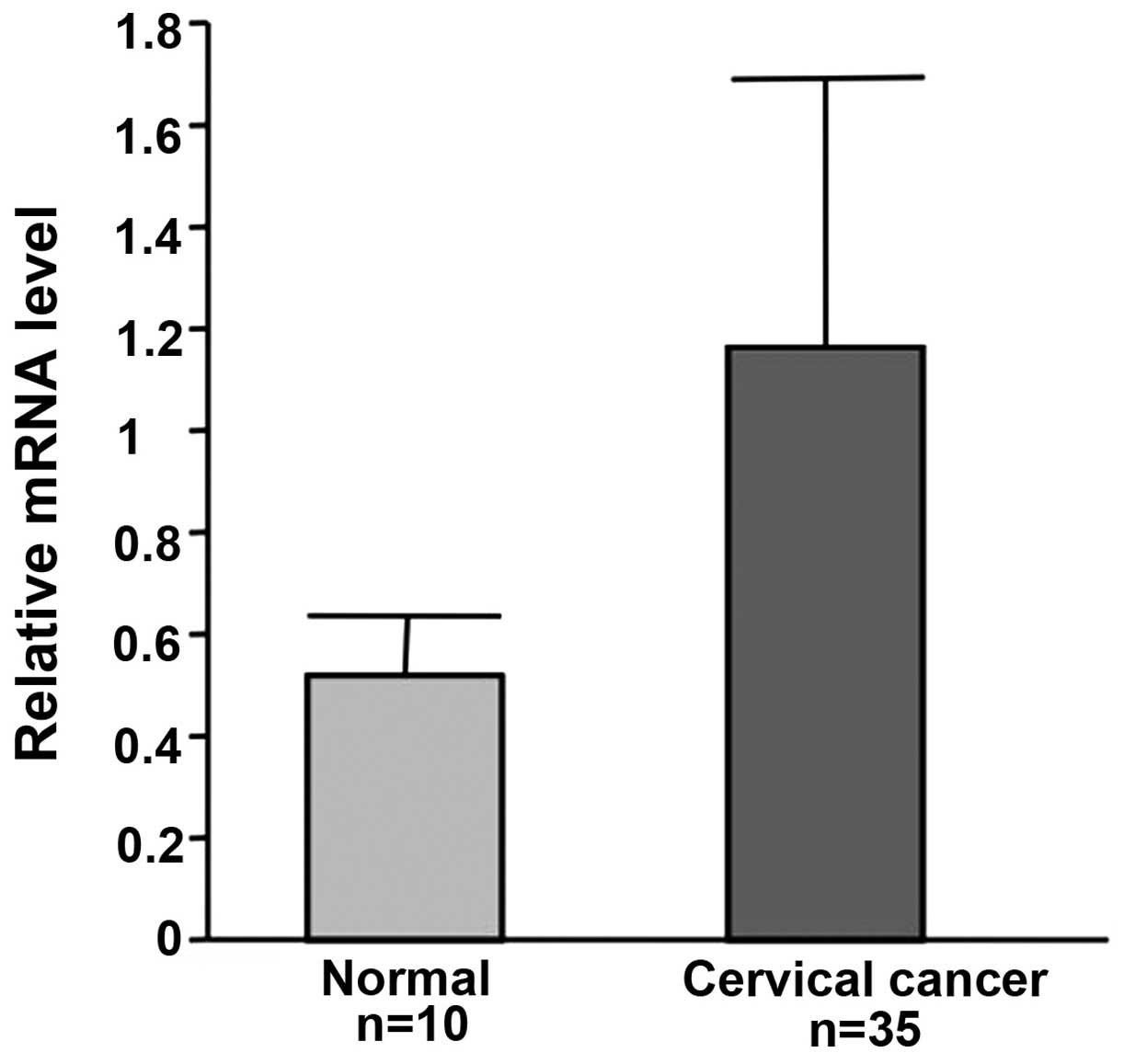
uPAR mRNA expression in cervical cancer
tissue
The expression of uPAR mRNA was examined in cervical
cancer from 35 patients and normal cervix from 10 patients who
suffered from uterine myoma or prolapse of the uterus. Triplicate
measurements were carried out. uPAR mRNA levels were much higher in
cervical cancer than in normal tissues (Fig. 6).
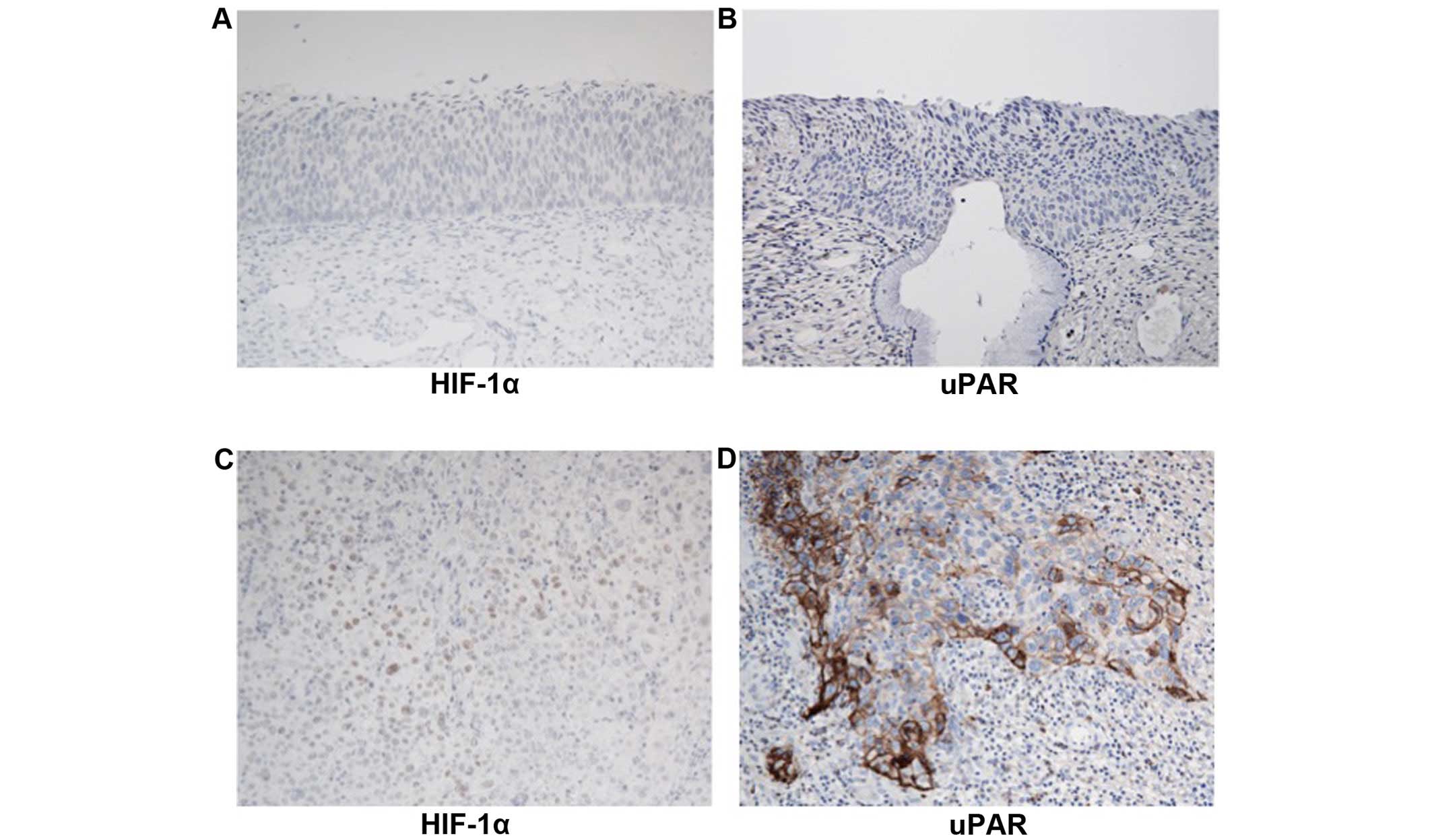
Localization of the uPAR expression
Immunohistochemical analysis of uPAR may lead to the
understanding of neoplastic progression of invasion in cervical
carcinogenesis. Increased nuclear staining of HIF-1α was observed
in invasive cervical cancer tissue samples (Fig. 7C). However, no or faint staining was
observed in CIN3 (Fig. 7A).
High-level cytoplasmic staining of uPAR was detected in invasive
cervical cancer, whereas not in CIN3 (Fig. 7B and D).
Discussion
Tumor cells need to migrate through the
extracellular matrix and invade blood vessels into systemic
circulation to be disseminated to metastatic organs (13,14).
Firstly, tumor cells detach from the primary tumor, invade blood
vessels and then metastasize in organs where they form the
secondary lesion. In spite of significant progress in cancer
biology, it is not known why only some but not all cells of a
clonal tumor population acquire the ability to migrate through the
tissue barriers (14). One
hypothesis is that specific transient regional tumor
micro-environment of low oxygen mediates a distinctive epigenetic
gene expression profile in a subset of cells, which likely causes
selection of more invasive cell clones (15,16).
Tumor hypoxia is one of the most critical situations during
development of tumor invasion.
Our data show that HIF-1 transactivates the uPAR
promoter and then mediates the induction of uPAR and invasion in
cervical cancer cells during hypoxia. We have also shown that the
protein level of uPAR in cervical cancer samples is higher than
that in normal cervix. The level of uPAR was increased under
hypoxic conditions, as determined by real-time RT-PCR analysis and
western blotting in cervical cancer cell lines. When siRNA was used
to reduce HIF-1α expression, not only uPAR expression but also
invasiveness was significantly diminished during hypoxia. According
to electrophoretic mobility shift assay, we identified the active
HRE in the uPAR core promoter. These findings may be relevant to
several biological processes in which cell invasion occurs. For
example, increased expression of uPAR in cancer may facilitate
their invasion through hypoxic regions of the stroma (17). In addition, increased expression of
uPAR is enhanced in migrating endothelial cells in vitro,
and the fact that neovascularization of transplanted tumors in
vivo is inhibited by uPAR antagonists suggests a role for
hypoxia-stimulated uPAR expression in tumor angiogenesis (18–20).
uPAR antagonists inhibit tumor metastasis, as well as local
invasion, perhaps explaining the association of elevated uPAR
levels in cervical, breast and other types of carcinomas with a
poor clinical outcome (21–23).
Correlation between HIF-1 and regulation of uPAR has
rarely been shown. The protease system of uPA/uPAR and
metalloproteinases participate in the metastatic disease
progression, which depends on hypoxia (13). A previous study showed that exposure
of MDA-MB231 human breast carcinoma cells to hypoxia upregulated
uPAR expression and increased invasion (17). It has also shown that these effects
were prevented by incubating the cells with a blocking antibody to
uPAR (17). It was hypothesized in
that previous study that HIF-1 may be involved in the
transcriptional activation of the uPAR gene (17). Also, Büchler et al has shown
that the effects of hypoxia on uPAR and invasion were mediated
directly by HIF-1 in pancreatic cancer cells (24).
The HRE (5′-RCGTG-3′) overlaps the E-box (CACGTG),
which is known to bind to several nuclear factors, such as c-Myc,
Max and Mad (25–28). It is unclear whether HIF-1 competes
with these factors for binding to these sites, whereas recent
findings showed that hypoxia downregulated the c-Myc expression
(29). Under hypoxic conditions,
HIF-1α may play a predominant role in regulating promoter activity
of uPAR. Taken together, these data strongly indicate that
hypoxia-induced uPAR expression is due to the enhanced activity of
HIF-1 on the uPAR promoter. Furthermore, the proximal HRE in the
uPAR promoter is essential for the upregulation of uPAR during
hypoxia since the uPAR promoter was no longer transactivated by
hypoxia when we used the puPAR-141mt-luc which had the mutated
putative HRE by replacement of AGG with TTT.
These data may be important for future therapeutics
since local tumor invasion and early metastatic progression are the
most challenging clinical features of cervical cancer. The
coordinated activation by HIF-1 of a large battery of target genes,
the protein products of which perform diverse but related functions
contributing to tumor invasion, suggests that inhibitors of HIF-1
activity may have therapeutic utility as anticancer agents
(30). In conclusion, our results
indicate that hypoxia-induced upregulation of uPAR is mediated by
HIF-1 in cervical cancer cells, and provide evidence that
regulation of uPAR expression by HIF-1 represents a mechanism for
invasive cervical cancer during hypoxia.
Acknowledgments
The present study was supported by Grants-in-Aids
from the Ministry of Education, Culture, Sports, Science and
Technology of Japan (MEXT).
References
|
1
|
Brizel DM, Scully SP, Harrelson JM,
Layfield LJ, Bean JM, Prosnitz LR and Dewhirst MW: Tumor
oxygenation predicts for the likelihood of distant metastases in
human soft tissue sarcoma. Cancer Res. 56:941–943. 1996.PubMed/NCBI
|
|
2
|
Brizel DM, Sibley GS, Prosnitz LR, Scher
RL and Dewhirst MW: Tumor hypoxia adversely affects the prognosis
of carcinoma of the head and neck. Int J Radiat Oncol Biol Phys.
38:285–289. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Vaupel P, Thews O, Mayer A, Höckel S and
Höckel M: Oxygenation status of gynecologic tumors: What is the
optimal hemoglobin level? Strahlenther Onkol. 178:727–731. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Duong HS, Le AD, Zhang Q and Messadi DV: A
novel 3-dimensional culture system as an in vitro model for
studying oral cancer cell invasion. Int J Exp Pathol. 86:365–374.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Petrella BL, Lohi J and Brinckerhoff CE:
Identification of membrane type-1 matrix metalloproteinase as a
target of hypoxia-inducible factor-2 alpha in von Hippel-Lindau
renal cell carcinoma. Oncogene. 24:1043–1052. 2005. View Article : Google Scholar
|
|
6
|
Maity A and Solomon D: Both increased
stability and transcription contribute to the induction of the
urokinase plasminogen activator receptor (uPAR) message by hypoxia.
Exp Cell Res. 255:250–257. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Montuori N, Mattiello A, Mancini A,
Santoli M, Taglialatela P, Caputi M, Rossi G and Ragno P:
Urokinase-type plasminogen activator up-regulates the expression of
its cellular receptor through a post-transcriptional mechanism.
FEBS Lett. 508:379–384. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Giatromanolaki A, Koukourakis MI, Sivridis
E, Turley H, Talks K, Pezzella F, Gatter KC and Harris AL: Relation
of hypoxia inducible factor 1 alpha and 2 alpha in operable
non-small cell lung cancer to angiogenic/molecular profile of
tumours and survival. Br J Cancer. 85:881–890. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Semenza GL: Hypoxia-inducible factors in
physiology and medicine. Cell. 148:399–408. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fukuda R, Zhang H, Kim JW, Shimoda L, Dang
CV and Semenza GL: HIF-1 regulates cytochrome oxidase subunits to
optimize efficiency of respiration in hypoxic cells. Cell.
129:111–122. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nishi H, Nakada T, Kyo S, Inoue M, Shay JW
and Isaka K: Hypoxia-inducible factor 1 mediates upregulation of
telomerase (hTERT). Mol Cell Biol. 24:6076–6083. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
McNally SJ, Harrison EM, Ross JA, Garden
OJ and Wigmore SJ: Curcumin induces heme oxygenase 1 through
generation of reactive oxygen species, p38 activation and
phosphatase inhibition. Int J Mol Med. 19:165–172. 2007.
|
|
13
|
Kim J, Yu W, Kovalski K and Ossowski L:
Requirement for specific proteases in cancer cell intravasation as
revealed by a novel semiquantitative PCR-based assay. Cell.
94:353–362. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yamano M, Fujii H, Takagaki T, Kadowaki N,
Watanabe H and Shirai T: Genetic progression and divergence in
pancreatic carcinoma. Am J Pathol. 156:2123–2133. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lal A, Peters H, St Croix B, Haroon ZA,
Dewhirst MW, Strausberg RL, Kaanders JH, van der Kogel AJ and
Riggins GJ: Transcriptional response to hypoxia in human tumors. J
Natl Cancer Inst. 93:1337–1343. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Koong AC, Denko NC, Hudson KM, Schindler
C, Swiersz L, Koch C, Evans S, Ibrahim H, Le QT, Terris DJ, et al:
Candidate genes for the hypoxic tumor phenotype. Cancer Res.
60:883–887. 2000.PubMed/NCBI
|
|
17
|
Graham CH, Forsdike J, Fitzgerald CJ and
Macdonald-Goodfellow S: Hypoxia-mediated stimulation of carcinoma
cell invasiveness via upregulation of urokinase receptor
expression. Int J Cancer. 80:617–623. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mazar AP, Henkin J and Goldfarb RH: The
urokinase plasminogen activator system in cancer: Implications for
tumor angiogenesis and metastasis. Angiogenesis. 3:15–32. 1999.
View Article : Google Scholar
|
|
19
|
Bauer TW, Liu W, Fan F, Camp ER, Yang A,
Somcio RJ, Bucana CD, Callahan J, Parry GC, Evans DB, et al:
Targeting of urokinase plasminogen activator receptor in human
pancreatic carcinoma cells inhibits c-Met- and insulin-like growth
factor-I receptor-mediated migration and invasion and orthotopic
tumor growth in mice. Cancer Res. 65:7775–7781. 2005.PubMed/NCBI
|
|
20
|
Reuning U, Sperl S, Kopitz C, Kessler H,
Krüger A, Schmitt M and Magdolen V: Urokinase-type plasminogen
activator (uPA) and its receptor (uPAR): Development of antagonists
of uPA/uPAR interaction and their effects in vitro and in vivo.
Curr Pharm Des. 9:1529–1543. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Degryse B, Sier CF, Resnati M, Conese M
and Blasi F: PAI-1 inhibits urokinase-induced chemotaxis by
internalizing the urokinase receptor. FEBS Lett. 505:249–254. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ramont L, Pasco S, Hornebeck W, Maquart FX
and Monboisse JC: Transforming growth factor-beta1 inhibits tumor
growth in a mouse melanoma model by down-regulating the plasminogen
activation system. Exp Cell Res. 291:1–10. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fox SB, Taylor M, Grøndahl-Hansen J,
Kakolyris S, Gatter KC and Harris AL: Plasminogen activator
inhibitor-1 as a measure of vascular remodelling in breast cancer.
J Pathol. 195:236–243. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Büchler P, Reber HA, Tomlinson JS,
Hankinson O, Kallifatidis G, Friess H, Herr I and Hines OJ:
Transcriptional regulation of urokinase-type plasminogen activator
receptor by hypoxia-inducible factor 1 is crucial for invasion of
pancreatic and liver cancer. Neoplasia. 11:196–206. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Montagne M, Naud JF, McDuff FO and Lavigne
P: Toward the elucidation of the structural determinants
responsible for the molecular recognition between Mad1 and Max.
Biochemistry. 44:12860–12869. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hu J, Banerjee A and Goss DJ: Assembly of
b/HLH/z proteins c-Myc, Max, and Mad1 with cognate DNA: Importance
of protein-protein and protein-DNA interactions. Biochemistry.
44:11855–11863. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Semov A, Marcotte R, Semova N, Ye X and
Wang E: Microarray analysis of E-box binding-related gene
expression in young and replicatively senescent human fibroblasts.
Anal Biochem. 302:38–51. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Vervoorts J and Lüscher B: DNA binding of
Myc/Max/Mad network complexes to oligonucleotides containing two E
box elements: c-Myc/Max heterodimers do not bind DNA cooperatively.
Biol Chem. 380:1121–1126. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Koshiji M, Kageyama Y, Pete EA, Horikawa
I, Barrett JC and Huang LE: HIF-1alpha induces cell cycle arrest by
functionally counteracting Myc. EMBO J. 23:1949–1956. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Powis G and Kirkpatrick L: Hypoxia
inducible factor-1alpha as a cancer drug target. Mol Cancer Ther.
3:647–654. 2004.PubMed/NCBI
|