Introduction
Colorectal cancer (CRC) is one of the most
aggressive cancers in the western countries. Despite the
development of new treatments, patients suffering from this disease
die of the spread of the primary cancer to other organs, mainly the
liver, rather than from the primary tumor. To date, the only
effective treatment is hepatic resection, and thus, a novel and
effective therapies are urgently needed to treat metastatic
disease, improving life expectancy and quality in CRC patients.
Several studies have included biologically active compounds due to
their low toxicity and excellent potential for cancer
treatment.
The relationship between inflammation and tumor
development is widely accepted (1).
During the progression of cancer inflammatory cells are recruited
into the tumor stroma by signals derived from the tumor, as well as
from the cells residing in the organ that is being colonized. This
inflammatory milieu results in the trigger for oxidative stress. In
this scenery, the presence of oxidative and inflammatory factors
play a crucial role regulating the expression of genes involved in
tumor progression and metastasis (2).
This off balance in the microenvironment of the
developing tumor affects the major cell types of the host organ in
the tumor, namely, the cancer associated fibroblasts (CAFs)
(3). The CAFs contribute to this
pro-tumoral microenvironment by further producing pro-inflammatory
and pro-angiogenic factors favoring the growth of the tumor,
invasion of the targeted organ, angiogenesis and finally metastasis
(3).
This inflammatory and oxidative millieu has
encouraged the validation of several nutritional and biological
components with anti-inflammatory and anti-oxidative properties, as
complements for antitumor therapies. Among others, extracts from
green tea and licorice, and vitamins has been reported to possess
immunomodulatory, anti-inflammatory and anti-oxidant properties
which make them excellent candidates for the validation of their
anti-tumorigenic and anti-metastatic effects in different types of
cancer (4–7). Moreover, the administration of several
of these compounds as nutrient mixtures has shown greater efficacy
than their administration as individual treatments (8,9).
OOS is a nutritional supplement with recognized
anti-oxidant, anti-inflammatory and immunomodulatory properties.
The solution is composed, among others, of green tea extract,
glyzyrrhicic acid, vitamin C, B6 and B12, minerals and aminoacids.
It was synthesized by combining two products, Viusid®
and Ocoxin®. The Viusid component has been tested in
different clinical trials showing beneficial effects in patients
suffering from chronic hepatitis C and cirrhosis (10,11).
Moreover, OOS antitumor effects have validated in vitro and
in vivo preclinical breast cancer models (12). Thus, we aim to study the effects of
OOS in the metastatic progression of CRC to the liver.
In the present study, we demonstrate the inhibitory
effect OOS in vitro and an in vivo pre-clinical model
of metastatic development of colorectal carcinoma to the liver. We
show that OOS exerts its effect by reducing tumor cell
proliferation and by increasing apoptosis in vivo.
Furthermore, this nutritional supplement shows not only a reduction
in the fibroblast recruitment to the tumor stroma, which has been
related to angiogenesis and tumor progression (13), but also to a modulation of the
inflammatory signature in the liver. Thus, OOS might be a novel and
effective complementary treatment which properties might help to
increase the efficacy of actual anti-metastatic therapies as well
as to improve the life-quality of patients suffering from this
disease.
Materials and methods
Animals
Balb/c mice (male, 8-weeks old) were obtained from
Charles River Laboratories España S.A. (Barcelona, Spain). The
animals were fed a standard chow and had access to water ad
libitum. All the proceedings were approved by the Basque
Country university ethics Committee for experimental Animal with
the reference number CEEA/357/2014/ARTETA RUIZ, in accordance with
the institutional, national and international guidelines regarding
the protection and care of animal use for scientific purposes.
Cell line
The murine colon cancer C26 cells, syngenic with
Balb/c mice (ATCC, LGC Standards S.L.U. Barcelona, Spain) were
used. Cells were grown under standard conditions in RPMI-1640
medium (Life Technologies, Madrid, Spain) supplemented with 10%
fetal bovine serum (FBS), penicillin (100 U/ml), streptomycin (100
µg/ml) and amphotericin B (0.25 µg/ml) all purchased
from Life Technologies.
Viability and cell cycle assays
PrestoBlue® reagent (Life Technologies)
was used for quantification of tumor cell viability following the
manufacturer's instructions. The C26 cells were cultured on
collagen (Sigma-Aldrich, St. Louis, MO, USA) precoated culture
plates at a concentration of 50,000 cells/ml in RPMI-1640
supplemented with 0.5% FBS and antibiotics-antimycotics. After 24-h
incubation in the presence of different OOS concentrations ranging
from 0 to a maximum of 1:200 (V/Vf) PrestoBlue was added for 2 h.
Finally, absorbance was measured with Fluoroskan Ascen (Thermo
Labsystems, Waltham, MA, USA). Results were calculated as the
average of three independent experiments. For cell cycle analyses,
cells were fixed after 72-h incubation in the presence of OOS
(1/100; V/Vf) and DNA stained with propidium iodide (PI; Life
Technologies) for 30 min. Then, cells were analyzed by flow
cytometry in a Beckman Coulter Gallios™ (Beckman Coulter Inc.,
Barcelona, Spain). The number of cells in each cell cycle phase was
quantified by means of weasel software.
In vitro migration assay
The migration assay was carried out on modified
Boyden chambers. Briefly, tumor cells, at a concentration of
2×105 cells/ml were cultured onto 8 µm-diameter
pore membrane (Greiner Bio-One, La Jolla, CA, USA) precoated with
collagen type I 10 µg/ml (Sigma-Aldrich). Tumor cells were
allowed to migrate for 18 h in the presence of OOS (1:100 V/Vf) in
culture medium supplemented with 0.5% FBS. Migrated cells were
quantified after 4% formalin fixation and crystal violet staining
(Sigma-Aldrich). Results were calculated from three independent
experiments and data were expressed as the mean of total migrated
cells per membrane.
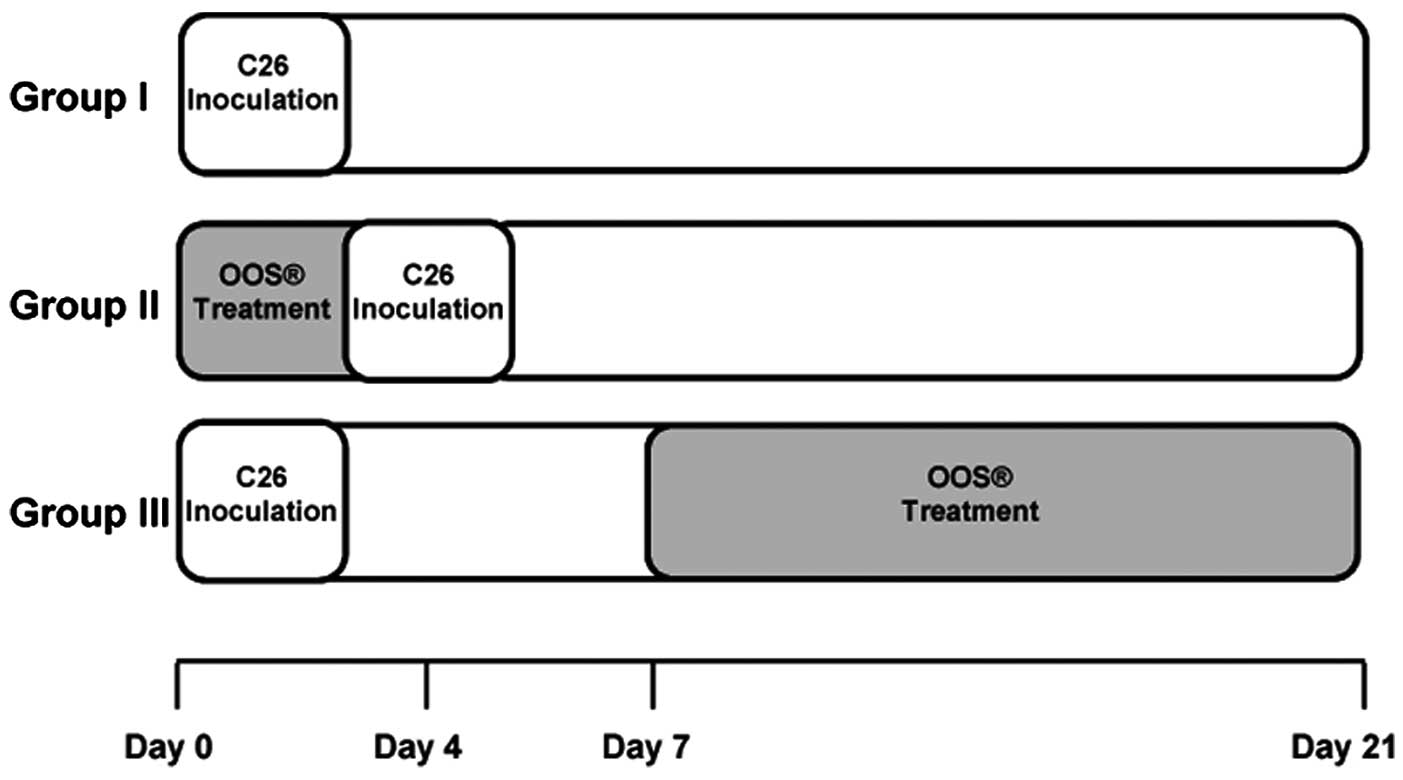
Experimental development of hepatic
metastasis
In vivo metastasis assay was carried out by
intrasplenical (i.s.) inoculation of tumor cells. The cells
(2×105) were inoculated in the inferior pole of the
spleen under anesthesia. The animals were treated with 100
µl of OOS and divided into 3 groups based on administration
pattern as follows: in group I, animals received no treatment; in
group II, mice were treated with OOS 4 days prior to tumor cell
inoculation; and in group III, mice were treated with OOS from the
7th day after tumor cell inoculation until the day of sacrifice
(Fig. 1). As control for tumor
development at the time of first doses of OOS in group III three
untreated animals were sacrificed the 7th day after tumor cell
inoculation. Animals from groups I, II and III were sacrificed 21
days after tumor cell inoculation, and livers were collected, fixed
in zinc solution (Sigma-Aldrich) and paraffin embedded for
histological analyses after H&E staining. Tumor occupied area
was quantified in three 7 µm-thick sections per liver,
separated 500 µm from each other and calculated as the area
occupied by tumor foci per section of liver tissue. At least 6 mice
per group were used per each experiment. Once the most effective
administration pattern was chosen by treating mice with 100
µl of OOS, the effectiveness of a lower OOS concentration
(50 µl) was tested under the same experimental
conditions.
Immunohistological analyses of liver
tissue
Liver tissue was analyzed for the expression of
Ki-67 (ab16667; 1:100; Abcam, Cambridge, UK) and caspase-3 (ab4051;
1:100; Abcam) by immunohistology. Mice from group I and III were
sacrificed and livers were collected for immunohistological
analyses in 7 µm tissue sections by staining with specific
antibodies. Additionally, the expression of α-smooth muscle actin
(ASMA) (MCA5781GA; 1:100; AbD Serotec, Raeigh, NC, USA) was also
quantified. Antigen retrieval was carried out in citrate buffer pH
6.0, then endogenous peroxidase and inspecific proteins were
blocked by incubating for 40 min with 3% of
H2O2 and 40 min with 3% FBS. Finally, liver
tissue was incubated with specific antibodies and antigen
expression was revealed by horseradish peroxidase (HRP)-conjugated
streptavidin (Life Technologies) and 2-Solution DAB kit (Life
Technologies) following the manufacturer's instructions. Antigen
expression levels were quantified by ImageJ software (NIH,
Bethesda, MD, USA). Results were expressed as the mean of at least
6 liver sections for each treatment.
RT-qPCR
Total RNA was extracted from paraffin embedded liver
tissues collected from mice treated with 100 µl of OOS
according to group I and III dosage pattern using Norgen FFPE RNA
Purification kit (Norgen Biotek Corp., Thorold, ON, Canada)
following the manufacturer's instructions. RNA concentration and
quality was assessed by NanoDrop spectrophotometer (ND-1000; Thermo
Fisher Scientific Inc., Rockford, IL, USA), and 2 µg RNA was
reverse transcribed into cDNA with recombinant moloney murine
leukemia virus reverse transcriptase and random primers (Life
Technologies). Quantification of cDNA template was performed with
real-time polymerase chain reaction (PCR) using SYBR-Green (Life
Technologies) as a fluorophore in ABI 7900HT (Life Technologies).
PCR primers (Life Technologies) were as follows: GAPDH
(housekeepeing) F, GTATGACTCCACTCACGGCAA and R,
CTTCCCATTCTCGGCCTTG; IL1β F, CTGTGACTCATGGGATGATGATG and R,
GCCTGTAGTGCAGTTGTCTAAT; INFγ F, TTCTTCAGCAACAGCAAGGC and R,
TGTGGGTTGTTGACCCTCAAA; COX2 F, TGCACTATGGTTACAAAAGCTGG and R,
TCGGAAGCTCCTTATTTCCCTT; TNFα F, CCAGTGTGGGAAGTGTCTT and R,
AAGCAAAAGAGGAGGCAACA; IL6 F, TCTATACCACTTCACAAGTCGGA and R,
GAATTGCCATTGCACAACTCTTT. Relative expression of target genes was
normalized to the internal control gene GAPDH by the ΔΔCt method.
Data were generated by the use of specific software (ABI Prism,
SDS2.0; life Technologies) after normalization. Results were
calculated at least from 6 livers per treatment and from two
independent experiments.
Statistical analysis
Statistical analysis was performed with the
Student's two-tailed unpaired t-test. Data are expressed as the
mean ± SD. The criterion for significance was P<0.05 for all
comparisons.
Results
OOS reduced the viability of C26 cell
line in vitro
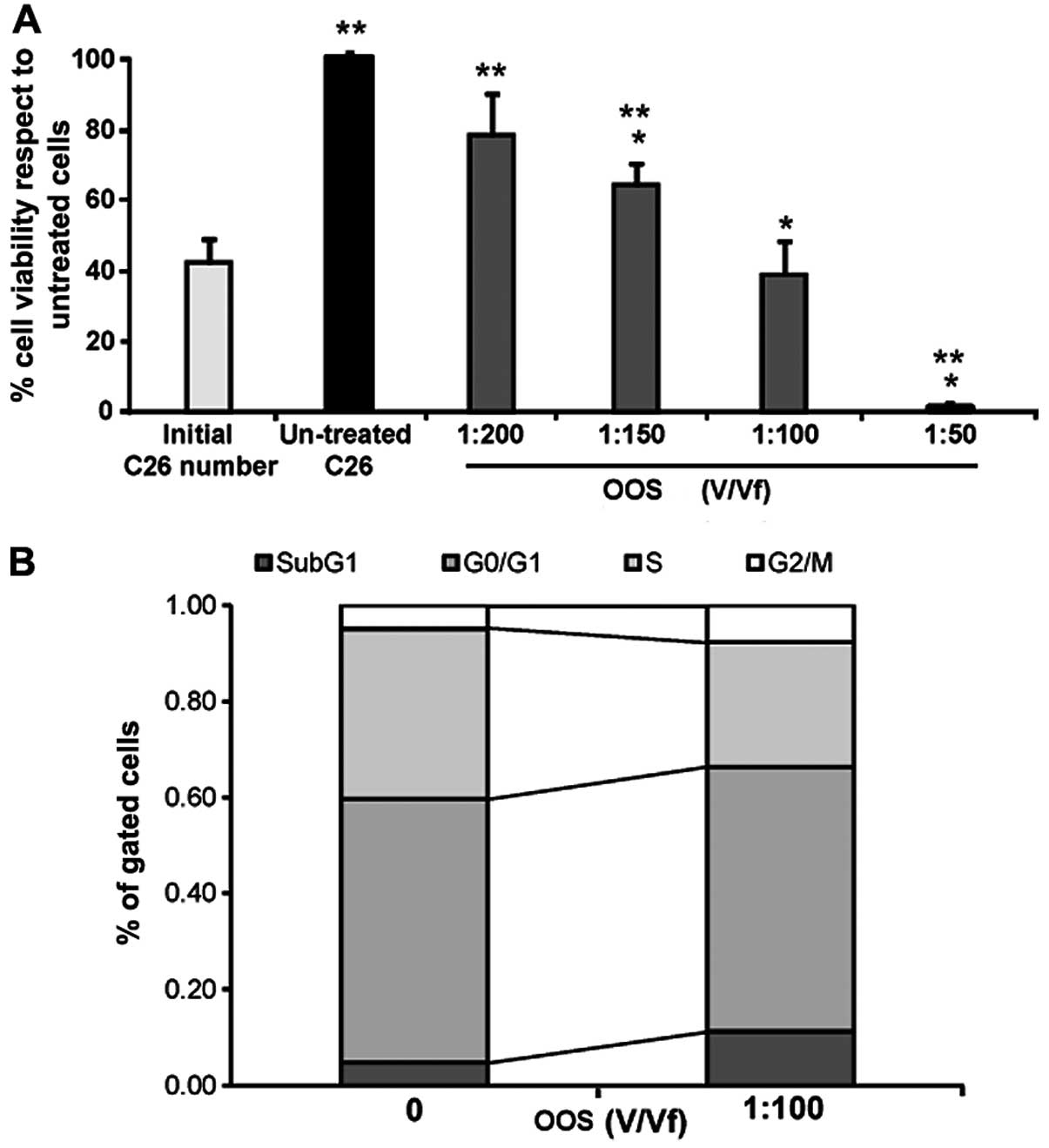
In order to test the effect of OOS in the viability
of colon carcinoma cells, C26 cells were treated with increasing
concentrations from 0 to 1:200 (V/Vf) of OOS for 24 h. As shown in
Fig. 2A, the viability of tumor
cells decreased in a dose-dependent manner from 20% at 1:200 (V/Vf)
of OOS to >90% at 1:50 (V/Vf) (Fig.
2A) in respect to untreated C26 cells. However, when compared
to the initial cell number, control untreated cells and every
treatment shows viability differences but 1:100 (V/Vf) resulted in
a significant non-proliferation.
Since the only concentration which inhibited
completely the proliferation of C26 cells was 1:100 OOS (V/Vf), we
next, analyzed the amount of cells in each phase of the cell cycle
by means of PI staining after the cuture for 72 h in the presence
of OOS. Then, the cells were analyzed by flow cytometry. As seen in
Fig. 2B, the number of cells in
phase G2/M and in the peak Sub G1 were
increased while the cell number ongoing DNA replication in phase S
showed a significant decrease compared to those cells cultured
under basal conditions.
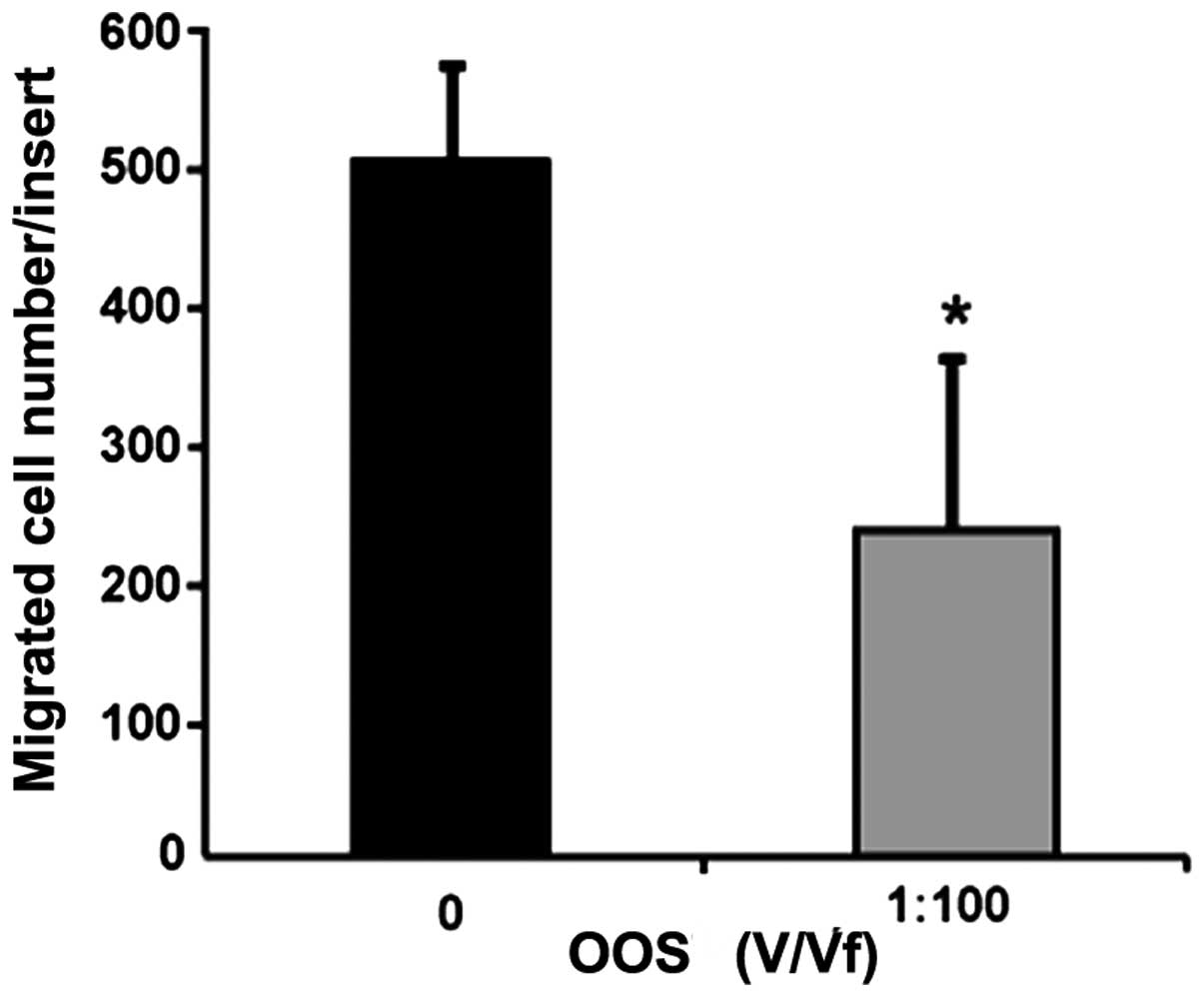
In vitro migratory potential of
C26 cells is reduced by OOS
To assess the effect of OOS in the migratory
potential of C26 cells through a collagen type I layer, we
incubated C26 cells on top of collagen type I-covered 8
µm-diameter pore membranes. The C26 cells were allowed to
migrate for 18-h in the presence of 1:100 of OOS (V/Vf). The amount
of migrated tumor cells was decreased by 50% in presence of 1:100
of OOS (V/Vf) (Fig. 3).
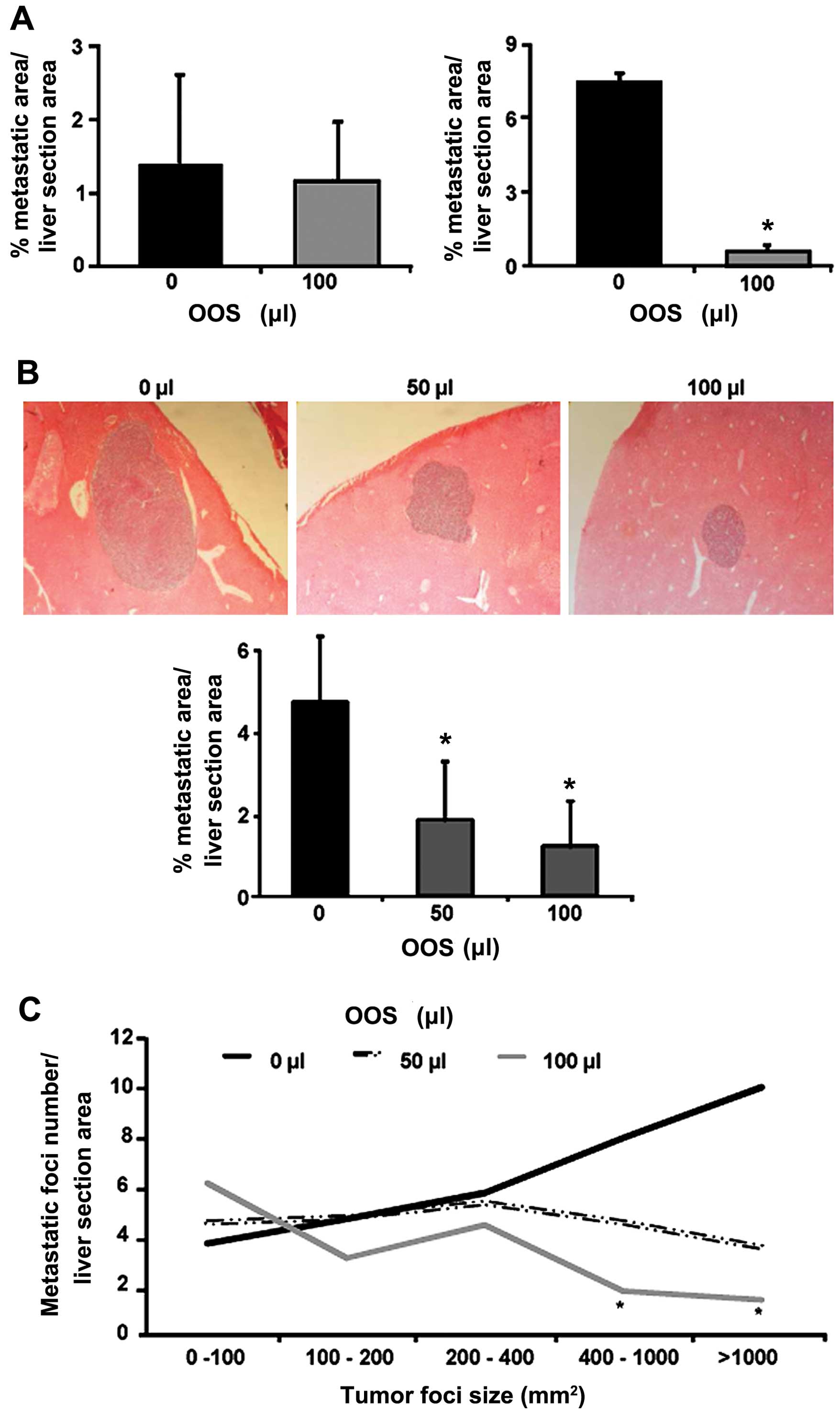
Effects of administration pattern in OOS
efficacy during the development of metastasis to the liver in
vivo
Next, the more efficient dosage pattern of OOS was
assessed. The tumor cells were inoculated i.s. and 100 µl of
OOS was administered once a day under the schedule described in
Materials and methods (Fig. 1),
until sacrifice. Under these dosage protocols, the weight of liver
and spleen did not show any change (data not shown) among the
experimental groups. The nutrient mixture only exerted significant
effects in those animals included in group III (Fig. 4A), while no effect was detected in
the preventive administration of OOS (group II) on the tumor burden
in the liver when compared to untreated group I (Fig. 4A). Even though, the metastatic
development was decreased at day 21 after tumor cell inoculation in
mice treated with OOS from day 7, the progression was not inhibited
but slowed down, since at day 7 after tumor cell inoculation only
few micrometastasis could be observed (data not shown).
Dose-dependent effect of OOS in the
metastasic development to the liver in vivo
Once the most effective pattern of administration
was established, a lower dose of OOS efficacy was tested under
these circumstances. Mice were treated with 50 and 100 µl of
OOS following the administration pattern of group III. After the
collection of livers and processing for histological analyses the
quantification of liver area occupied by tumor foci were carried
out. The results show, a reduction of >50% in the area of liver
tissue occupied by the tumor in those livers collected from 50 and
100 µl-treated mice (Fig.
4B). Even more, after the classification and quantification of
the tumor foci by their size, an decrease in the number of
metastatic foci larger than 400 mm2 of diameter were
detect in mice treated with 50 and 100 µl OOS compared to
those observed in the livers of untreated mice. Indeed, this
difference was only significant in livers collected from the
animals treated with 100 µl of OOS (Fig. 4C). However, no significant
difference was observed between the treatments.
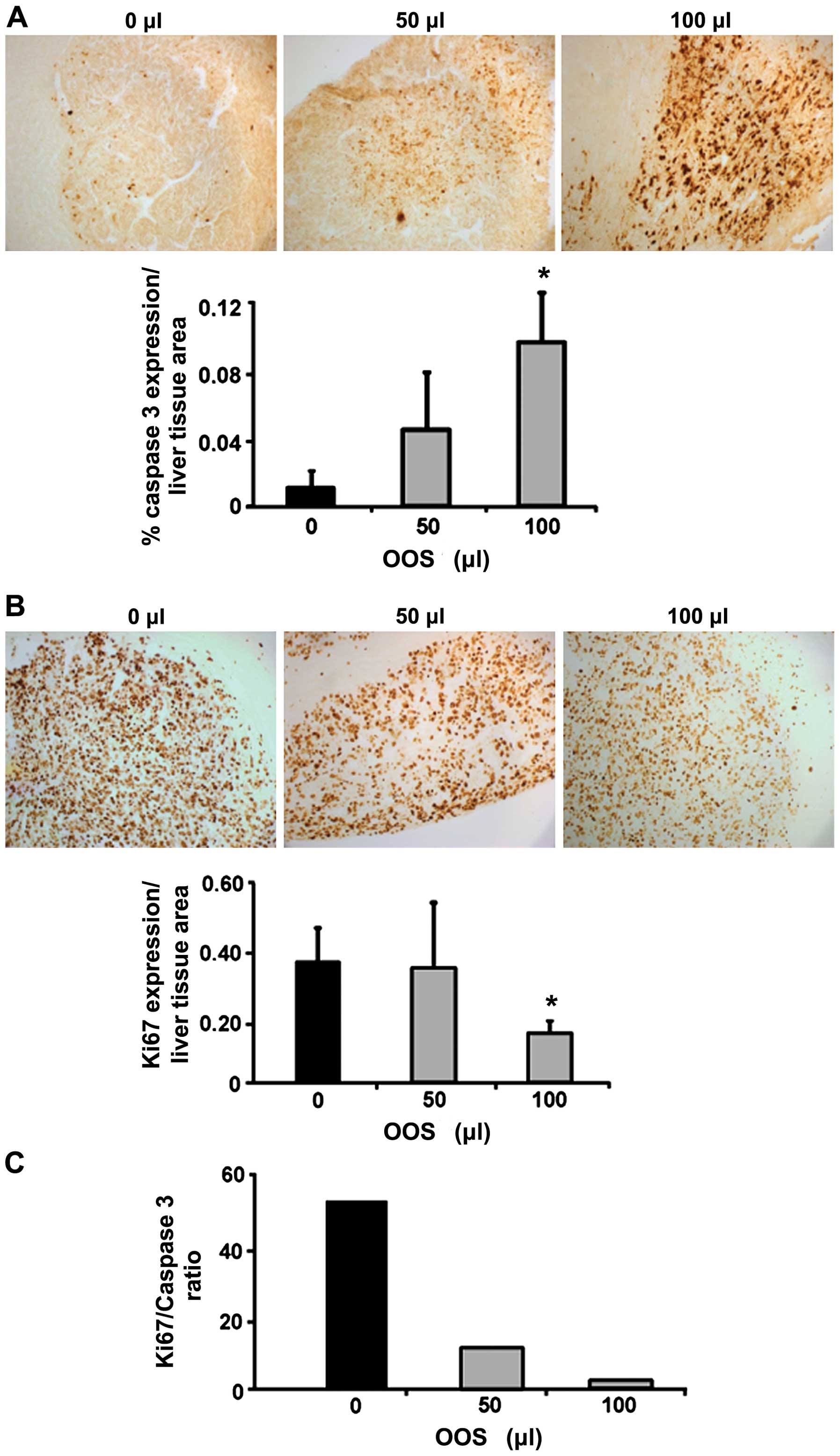
OOS affects tumor proliferation and
apoptosis in vivo
To quantify the expression levels for Ki-67 and
caspase-3, livers were collected either from untreated C26-bearing
mice and from C26-bearing mice treated with 50 or 100 µl of
OOS from day 7 since this dosage pattern was the only one showing
any kind of effectiveness. The livers were fixed and embedded in
paraffin and the expression levels of Ki-67 and caspase-3 were
analyzed by immunohistochemistry. As shown in Fig. 5A while the levels of caspase-3
expression were increased 10-fold after 100 µl OOS
administration compared to the untreated group. No significant
changes were observed in the animals treated with 50 µl of
OOS. Furthermore, the levels of Ki-67 showed a decrease in the
tumor foci developed in the livers of mice treated with 100
µl OOS compared to those observed in the livers from
untreated mice (Fig. 5B). However,
the difference was not observed in the livers collected from mice
treated with 50 µl of OOS. Even more, the ratio between
Ki-67 and caspase-3 expression was reduced in mice injected with
C26 cells and treated with both concentrations of OOS vs. Untreated
mice (Fig. 5C).
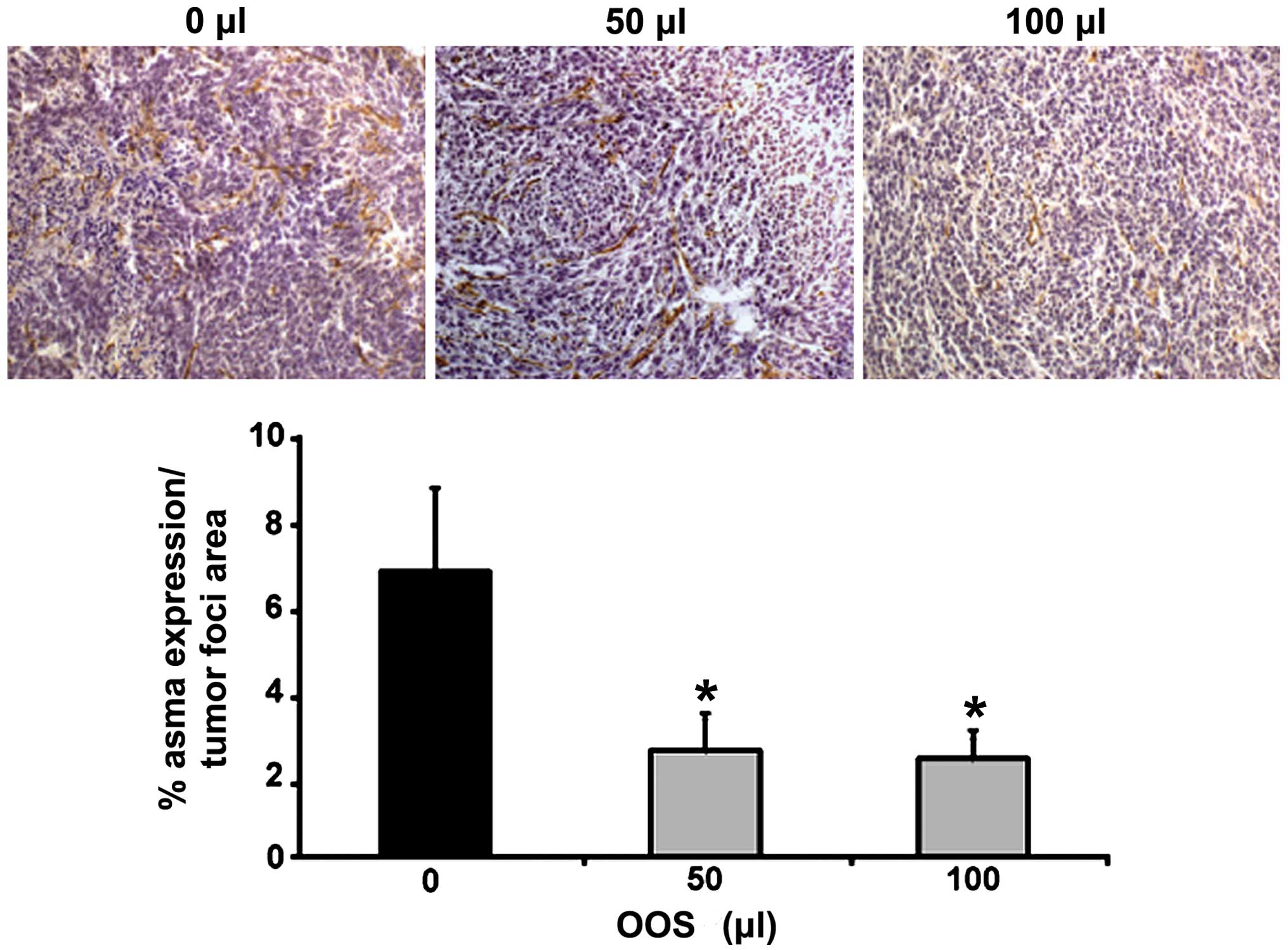
OOS treatment limits the infiltration of
myofibroblasts in the metastatic liver
Finally, the expression of ASMA was quantified in
the metastatic liver tissue in order to analyze the infiltration by
liver cancer-associated fibroblasts within the tumor foci. Sections
from those livers collected either from untreated C26-bearing mice
(group I) or from C26-bearing mice treated with 50 or 100 µl
of OOS following the dosage pattern for group III were stained with
specific antibodies for ASMA, an antigen expressed in CAFs. The
intratumoral levels of ASMA expression in liver tissue collected
from mice treated with 50 and 100 µl of OOS was reduced by
50% compared to the liver tumors obtained from untreated mice
(Fig. 6).
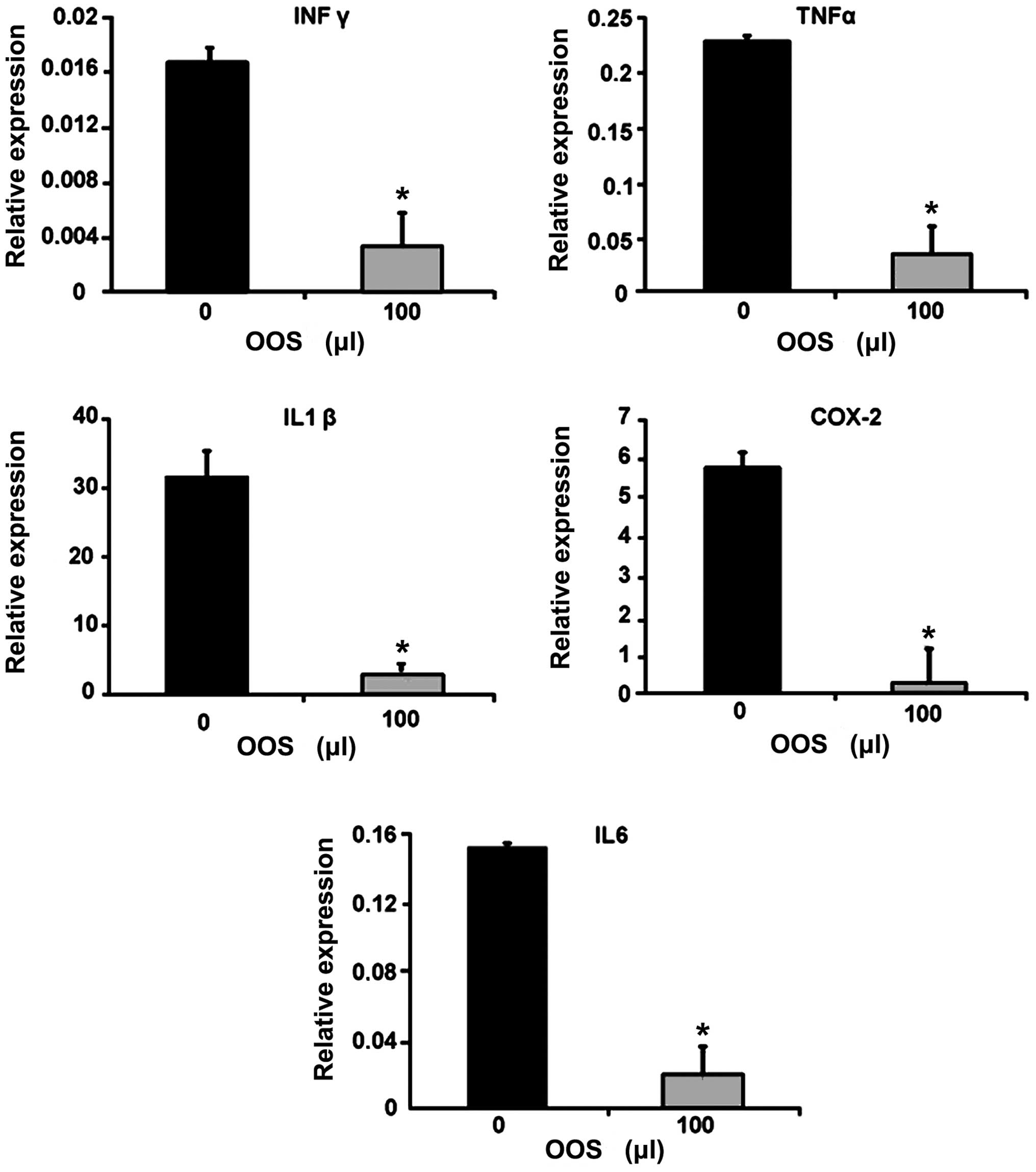
OOS modulates the expression of
inflammatory genes in metastatic livers
Since the tumor microenvironment is characterized by
an inflammatory signature, the gene expression of the inflammatory
molecules IFNγ, TNFα, COX-2, IL6 and IL1β was quantified in total
liver tissue by qPCR. As shown in Fig.
7, the expression of genes coding for the aforementioned
inflammatory molecules was significantly reduced in the livers
collected from mice treated with 100 µl of OOS according to
group III compared to those obtained from untreated mice (Fig. 7).
Discussion
CRC is one of the leading causes of cancer-related
deaths in the world, due mainly to the metastatic spread to distant
organs, specially the liver. Even though great advances have been
made in the development of therapies to treat CRC, they are often
aggressive and with limited efficacy. Thus, new complementary
therapies are being developed consisting in biological compound
mixtures. Certain nutrient mixtures have been proved to be
effective in several preclinical in vivo and in vitro
models such as pulmonary metastasis of melanoma and cervical cancer
(8,14). However, there is no report
describing the efficacy of these nutrient mixtures in the in
vivo metastatic spread of CRC to the liver. Thus, in the
present study, we aimed to investigate the effectiveness of OOS, a
complex mixture containing licorice extract, vitamins and minerals
which have proven anti-oxidant and anti-inflammatory properties in
other diseases (15,16) in the metastatic development of colon
carcinoma cells. Here, we highlight the anti-metastatic effect of
this nutrient mixture in vivo by means of reducing tumor
cell proliferation, migration and recruitment of CAFs, along with
an increase in apoptosis. Additionally, the mixture induced a
decrease in the expression of RNA levels for pro-inflammatory and
pro-angiogenic factors in metastasized liver provoking a slow down
of the metastatic development of colorectal cancer C26 cells to the
liver. Thus, these results support the need for further studies for
the use of OOS as a nutritional complement during the treatment of
colorectal cancer liver metastatic disease.
According to our results, the effect of OOS on the
metastatic progression of C26 cells to the liver depends on the
dosage pattern. Because a significant reduction in the tumor burden
of the liver was observed only in the mice treated with OOS 7 days
after tumor cell inoculation, but not when the solution was
administered days before the tumor inoculation. Lode et al
(17) have reported the inefficacy
of a nutrient mixture against metastasis in a neuroblastoma model.
However, as stated by Niedzwiecki (18) these observations might be due to
several reasons. On the one hand, the excessive amount of cells
inoculated by the authors or the need for longer and continuous
periods of treatment may account for this lack of effect. On the
other hand, antioxidants have been proved to exert their functions
mainly in the presence of chronic inflammation and in a highly
hypoxic environment (1). In the
absence of a highly inflammatory and pro-oxidant microenvironment
biologically active compounds might lack their anti-tumoral
activity (19), such as the one in
mice before the tumor inoculation. Thus, this could explain the
lack of effect of OOS in the early stage treatment. Even so,
further studies are needed to fully establish the mechanisms of OOS
when it is administered at early stages.
Dietary agents are believed to suppress, among
others, the hyper-proliferative processes during the development of
a tumor. Several phytochemicals have been shown to exert
suppressive effects on AP-1 and NF-κB interfering with growth and
proliferative signals (20,21). In accordance to this,
supplementation with dietary nutrient mixture OOS significantly
slows down murine colon carcinoma C26 tumor growth in vivo
in immune competent mice related to a decrease in the intratumoral
expression of Ki-67, an antigen strictly associated with cell
proliferation. The in vitro studies supported these findings
since the viability of tumor cells was significantly decreased and
a slight reduction in the number of cells in phase S was induced by
OOS treatment. Along with these results, an increase in the amount
of cells in phase G2/M was observed. In fact, cathechins
and licorice components cause cell cycle arrest by downregulating
cyclin D1 and E, key molecules in G1 to S transition and
completion of the latter one (22,23).
Ultimately, the cell cycle arrest will trigger cell cycle
deregulation and apoptotic processes via upregulation of p21 and
p27, both well-known tumor suppressor proteins downregulated in
numerous cancers (24). In addition
to the effect in the cell cycle progression, OOS breaks the
characteristic resistance of tumor cells to enter apoptosis.
Consistent with these observations, an increase in cell number in
subG1 in vitro and the intratumoral increase of caspase-3
in vivo after treatment with the complex nutrient mixture
points to the induction of programmed cell death within tumor foci
by this compound. The hypoxic and inflammatory tumor
microenvironment induces the activation of anti-apoptotic pathways
supporting the effect of anti-inflammatory or anti-oxidative drugs
which have been shown to increase the sensitivity of tumor cells to
pro-apoptotic signals (25). In
addition to its effects in tumor cell proliferation and apoptosis,
OOS reduced the migratory potential of C26 colon carcinoma cells
in vitro. Migration of tumor cells is a key step not only
for invasion of adjacent tissue in the primary organ but also for
extravasation and colonization of the secondary target organ during
the metastasic process. It is well-known that chronic inflammation
is linked to oxidative stress and both of them are related to the
invasive and migratory potential of cancer cells during tumor
progression (1). Thus, the
migratory capacity of tumor cells is also a potential target for
nutrient mixtures with anti-inflammatory and anti-oxidant
properties. Roomi et al (9,14) have
shown that in vitro, a nutrient mixture consisting in
lysine, ascorbic acid, proline and green tea, among others,
significantly reduced the migratory potential of a selected set of
different tumor cell types related to a decrease in the levels of
pro-migratory and pro-angiogenic factors such as proteases and VEGF
(9,14).
In the hepatic tumor microenvironment,
myofibroblasts are the most prominent CAFs. In the liver, CAFs are
mainly originated from transdifferentiated hepatic stellate cells
in the tumor stroma, and such tumor activated-hepatic stellate
cells promote tumor growth and invasiveness (26). Additionally, they contribute not
only to create a pro-tumoral stroma but also to trigger the
angiogenic switch allowing one step forward from an avascular to a
vascular state during the metastasic tumor growth (26). Furthermore, these tumor associated
fibroblasts participate actively in the production of
pro-inflammatory and pro-oxidant factors taking active part in
tumor expansion by inducing, among others, the trigering of an
angiogenic response. Moreover, recruited fibroblasts within a tumor
have been positively correlated with tumor progression and poor
prognosis in colorectal cancer (27). In our model, the nutrient mixture
OOS reduced the recruitment of myofibroblasts within the tumor
foci, as shown by a decreased amount of ASMA expressing cells
within the tumor foci which might also account for the reduced
angiogenesis. This reduction in myofibroblast infiltration in
vivo was related with an impaired migratory potential of 3T3
fibroblast in vitro by OOS (data not shown). The impaired
recruitment of CAFs might account as an additional mechanism by
which the nutrient mixture might exert its antitumoral effects.
Indeed, several compounds with anti-oxidant and anti-inflammatory
properties reduce the angiogenic response in different types of
tumors and show a significant impact in the tumor stroma formation
(28).
The antioxidant properties of several components of
OOS, such as polyphenols, cannot fully explain their antitumoral
activity. Cathechins modulate inflammatory pathways since they act
as signaling agents by interfering with NF-κB and AP-1 which, in
turn, results in inhibition of pro-inflammatory and pro-angiogenic
factors, including IL6, IL-1β, TNF-α and VEGF (29). Among them, TNFα or IL-1β induce the
expression of growth factors, stimulate epithelial tumor motility
and tumor angiogenesis (30,31).
Another proinflammatory molecule commonly upregulated in colon
cancer, COX-2, has been implicated in the growth and progression of
colorectal cancer via multiple pathways (32–34)
including the acquisition of resistance to apoptosis and promoting
angiogenesis through VEGF production (35,36).
Moreover, a decrease in COX-2 expression at RNA levels in total
liver caused by OOS administration might be related to a reduction
in VEGF levels (36). Additionally,
the reduction in Il-1β, and TNFα RNA levels after OOS
administration is consistent with the downregulation in the levels
of COX-2 RNA which is commonly stimulated by cytokines such as
Il-1β and TNFα as a result of the interaction between colon
carcinoma cells and the cells of the liver microenvironment
(36,37).
In summary, OOS slows down the metastatic
progression of CRC to the liver. Thus, the anti-oxidant and
anti-inflammatory properties of this nutrient mixture induced an
inhibition of the proliferative and migratory potential of tumor
cells which together with an increase in the sensitivity to
apoptotic signals might modulate the metastatic development of
colorectal cancer cells to the liver. Besides, OOS limits tumor
infiltration by CAFs and inhibits the production of inflammatory
and angiogenic factors within the tumor microenvironment.
Collectively, this creates an unfavorable and non-permissive
microenvironment for tumor growth suppressing the final steps of
tumor progression. Therefore, OOS may constitute a
pharmacologically safe complementary compound for the treatment of
cancer and its metastasis slowing down the tumor growth, and, thus,
increasing the life time and quality for patients suffering from
CRC liver metastasis. The above justifies further characterization
and validation of OOS in the metastatic development and their
combination with actual therapies.
Acknowledgments
We thank Evangelina Garcia and Maria Jesus Fernandez
for the excellent technical assistance. Additionally, we greatly
appreciate the support of the Genomics and Proteomics Unit, the
Analytical and High-Resolution Microscopy Unit and the Animal
Facilities from the Advance Research Facilities (SGIker) of the
University of Basque Country.
References
|
1
|
Reuter S, Gupta SC, Chaturvedi MM and
Aggarwal BB: Oxidative stress, inflammation, and cancer: How are
they linked? Free Radic Biol Med. 49:1603–1616. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mamlouk S and Wielockx B:
Hypoxia-inducible factors as key regulators of tumor inflammation.
Int J Cancer. 132:2721–2729. 2013. View Article : Google Scholar
|
|
3
|
Zhang Y, Tang H, Cai J, Zhang T, Guo J,
Feng D and Wang Z: Ovarian cancer-associated fibroblasts contribute
to epithelial ovarian carcinoma metastasis by promoting
angiogenesis, lymphangiogenesis and tumor cell invasion. Cancer
Lett. 303:47–55. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shankar S, Suthakar G and Srivastava RK:
Epigallocatechin-3-gallate inhibits cell cycle and induces
apoptosis in pancreatic cancer. Front Biosci. 12:5039–5051. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Goldberg ED, Amosova EN, Zueva EP, Razina
TG, Krylova SG and Zorikov PS: Licorice preparations improve
efficiency of chemotherapy and surgical treatment of transplanted
tumors. Bull Exp Biol Med. 145:252–255. 2008. View Article : Google Scholar
|
|
6
|
Ma YC, Li C, Gao F, Xu Y, Jiang ZB, Liu JX
and Jin LY: Epigallocatechin gallate inhibits the growth of human
lung cancer by directly targeting the EGFR signaling pathway. Oncol
Rep. 31:1343–1349. 2014.
|
|
7
|
Maruyama T, Murata S, Nakayama K, Sano N,
Ogawa K, Nowatari T, Tamura T, Nozaki R, Fukunaga K and Ohkohchi N:
(−)-Epigallocatechin-3-gallate suppresses liver metastasis of human
colorectal cancer. Oncol Rep. 31:625–633. 2014.
|
|
8
|
Roomi MW, Roomi N, Ivanov V, Kalinovsky T,
Niedzwiecki A and Rath M: Inhibition of pulmonary metastasis of
melanoma b16fo cells in C57BL/6 mice by a nutrient mixture
consisting of ascorbic Acid, lysine, proline, arginine, and green
tea extract. Exp Lung Res. 32:517–530. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Roomi MW, Kalinovsky T, Niedzwiecki A and
Rath M: Modulation of uPA, MMPs and their inhibitors by a novel
nutrient mixture in human colorectal, pancreatic and hepatic
carcinoma cell lines. Int J Oncol. 47:370–376. 2015.PubMed/NCBI
|
|
10
|
Gomez EV, Perez YM, Sanchez HV, Forment
GR, Soler EA, Bertot LC, Garcia AY, del Rosario Abreu vazquez M and
Fabian LG: Antioxidant and immunomodulatory effects of viusid in
patients with chronic hepatitis C. World J Gastroenterol.
16:2638–2647. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Vilar Gomez E, Sanchez Rodriguez Y, Torres
Gonzalez A, Calzadilla Bertot L, Arus Soler E, Martinez Perez Y,
Yasells Garcia A and Abreu Vazquez MR: Viusid, a nutritional
supplement, increases survival and reduces disease progression in
HCV-related decompensated cirrhosis: A randomised and controlled
trial. BMJ Open. 1:e0001402011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hernández-García S, González V, Sanz E and
Pandiella A: Effect of oncoxin oral solution in HER2-overexpressing
breast cancer. Nutr Cancer. 67:1159–1169. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tommelein J, Verset L, Boterberg T,
Demetter P, Bracke M and De Wever O: Cancer-associated fibroblasts
connect metastasis-promoting communication in colorectal cancer.
Front Oncol. 5(63)2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Roomi MW, Kalinovsky T, Cha J, Roomi NW,
Niedzwiecki A and Rath M: Effects of a nutrient mixture on
immunohistochemical localization of cancer markers in human
cervical cancer Hela cell tumor xenografts in female nude mice. Exp
Ther Med. 9:294–302. 2015.PubMed/NCBI
|
|
15
|
Lee SK, Park KK, Park JH, Lim SS and Chung
WY: The inhibitory effect of roasted licorice extract on human
metastatic breast cancer cell-induced bone destruction. Phytother
Res. 27:1776–1783. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu Y, Yu QY, Zhu ZL, Tang PY and LI K:
Vitamin B2 intake and the risk of colorectal cancer: A
meta-analysis of observational studies. Asian Pac J Cancer Prev.
16:909–913. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lode HN, Huebener N, Strandsby A and
Gaedicke G: Nutrient mixture including vitamin C, L-lysine,
L-proline, and epigal-locatechin is ineffective against tumor
growth and metastasis in a syngeneic neuroblastoma model. Pediatr
Blood Cancer. 50:284–288. 2008. View Article : Google Scholar
|
|
18
|
Niedzwiecki A: Micronutrient mixture in
tumor growth and metastasis. Pediatr Blood Cancer. 50:422–423;
author reply. 424–425. 2008. View Article : Google Scholar
|
|
19
|
Srividhya R, Jyothilakshmi V, Arulmathi K,
Senthilkumaran V and Kalaiselvi P: Attenuation of
senescence-induced oxidative exacerbations in aged rat brain by
(−)-epigallocatechin-3-gallate. Int J Dev Neurosci. 26:217–223.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Eferl R and Wagner EF: AP-1: A
double-edged sword in tumori-genesis. Nat Rev Cancer. 3:859–868.
2003. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Takada Y and Aggarwal BB: Flavopiridol
inhibits NF-kappaB activation induced by various carcinogens and
inflammatory agents through inhibition of IkappaBalpha kinase and
p65 phosphorylation: Abrogation of cyclin D1, cyclooxygenase-2, and
matrix metalloprotease-9. J Biol Chem. 279:4750–4759. 2004.
View Article : Google Scholar
|
|
22
|
Gupta S, Hussain T and Mukhtar H:
Molecular pathway for (−)-epigallocatechin-3-gallate-induced cell
cycle arrest and apoptosis of human prostate carcinoma cells. Arch
Biochem Biophys. 410:177–185. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hsia SM, Yu CC, Shih YH, Yuanchien Chen M,
Wang TH, Huang YT and Shieh TM: Isoliquiritigenin as a cause of DNA
damage and inhibitor of ataxia-telangiectasia mutated expression
leading to G2/M phase arrest and apoptosis in oral squamous cell
carcinoma. Head Neck. Jan 10–2015, View Article : Google Scholar : Epub ahead of
print.
|
|
24
|
He Y, Zhang X, Zeng X, Huang Y, Wei JA,
Han L, Li CX and Zhang GW: HuR-mediated posttranscriptional
regulation of p21 is involved in the effect of Glycyrrhiza
uralensis licorice aqueous extract on polyamine-depleted intestinal
crypt cells proliferation. J Nutr Biochem. 23:1285–1293. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lee JS, Ahn JH, Cho YJ, Kim HY, Yang YI,
Lee KT, Jang DS and Choi JH: α-Terthienylmethanol, isolated from
Eclipta prostrata, induces apoptosis by generating reactive oxygen
species via NADPH oxidase in human endometrial cancer cells. J
Ethnopharmacol. 169:426–434. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Olaso E, Salado C, Egilegor E, Gutierrez
V, Santisteban A, Sancho-Bru P, Friedman SL and Vidal-Vanaclocha F:
Proangiogenic role of tumor-activated hepatic stellate cells in
experimental melanoma metastasis. Hepatology. 37:674–685. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Berdiel-Acer M, Sanz-Pamplona R, Calon A,
Cuadras D, Berenguer A, Sanjuan X, Paules MJ, Salazar R, Moreno V,
Batlle E, et al: Differences between CAFs and their paired NCF from
adjacent colonic mucosa reveal functional heterogeneity of CAFs,
providing prognostic information. Mol Oncol. 8:1290–1305. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hassan LE, Ahamed MB, Majid AS, Baharetha
HM, Muslim NS, Nassar ZD and Majid AM: Correlation of
antiangiogenic, antioxidant and cytotoxic activities of some
Sudanese medicinal plants with phenolic and flavonoid contents. BMC
Complement Altern Med. 14(406)2014.PubMed/NCBI
|
|
29
|
Fassina G, Venè R, Morini M, Minghelli S,
Benelli R, Noonan DM and Albini A: Mechanisms of inhibition of
tumor angiogenesis and vascular tumor growth by
epigallocatechin-3-gallate. Clin Cancer Res. 10:4865–4873. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Leibovich SJ, Polverini PJ, Shepard HM,
Wiseman DM, Shively V and Nuseir N: Macrophage-induced angiogenesis
is mediated by tumour necrosis factor-alpha. Nature. 329:630–632.
1987. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Saijo Y, Tanaka M, Miki M, Usui K, Suzuki
T, Maemondo M, Hong X, Tazawa R, Kikuchi T, Matsushima K, et al:
Proinflammatory cytokine IL-1 beta promotes tumor growth of lewis
lung carcinoma by induction of angiogenic factors: In vivo analysis
of tumor-stromal interaction. J Immunol. 169:469–475. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tsujii M, Kawano S and DuBois RN:
Cyclooxygenase-2 expression in human colon cancer cells increases
metastatic potential. Proc Natl Acad Sci USA. 94:3336–3340. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tsujii M, Kawano S, Tsuji S, Sawaoka H,
Hori M and DuBois RN: Cyclooxygenase regulates angiogenesis induced
by colon cancer cells. Cell. 93:705–716. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sun Y, Tang XM, Half E, Kuo MT and
Sinicrope FA: Cyclooxygenase-2 overexpression reduces apoptotic
susceptibility by inhibiting the cytochrome c-dependent apoptotic
pathway in human colon cancer cells. Cancer Res. 62:6323–6328.
2002.PubMed/NCBI
|
|
35
|
Han YD, Hong YK, Kang JG, Choi YJ and Park
CH: Relation of the expression of cyclooxygenase-2 in colorectal
adenomas and adenocarcinomas to angiogenesis and prognosis. J
Korean Soc Coloproctol. 26:339–346. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Valcárcel M, Arteta B, Jaureguibeitia A,
Lopategi A, Martínez I, Mendoza L, Muruzabal FJ, Salado C and
Vidal-Vanaclocha F: Three-dimensional growth as multicellular
spheroid activates the proangiogenic phenotype of colorectal
carcinoma cells via LFA-1-dependent VEGF: Implications on hepatic
micrometastasis. J Transl Med. 6(57)2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Arteta B, Lasuen N, Lopategi A,
Sveinbjörnsson B, Smedsrød B and Vidal-Vanaclocha F: Colon
carcinoma cell interaction with liver sinusoidal endothelium
inhibits organ-specific antitumor immunity through
interleukin-1-induced mannose receptor in mice. Hepatology.
51:2172–2182. 2010. View Article : Google Scholar : PubMed/NCBI
|