Introduction
Cullin-5 (CUL5) is a member of the cullin-RING E3
ubiquitin ligase (CRL) family. CUL5 has been shown to be involved
in numerous important cellular processes including the cell cycle
and proliferation (1). Recognition
of the role of CUL5 in cancer cell growth and invasiveness began
with the demonstration that overexpression of CUL5 in T47D breast
cancer cells led to significant suppression of proliferation
(2). Subsequent studies involving
CUL5 expression in breast cancer as well as in cervical and
hepatocellular cancer have confirmed the antiproliferative effect
of increased CUL5 expression (3–5).
Endometrial adenocarcinoma is the most common
gynecologic cancer and one of the most common cancers in women
worldwide (6). The American Cancer
Society estimates that there will be nearly 55,000 new cases of
endometrial cancer in the US alone in 2015 with more than 10,000
deaths. Indeed, while patient outcomes for most cancers have
improved over the past two decades, overall survival among women
diagnosed with endometrial cancer has worsened (7). Thus, more effective therapeutic
intervention in endometrial cancer is needed.
More than 95% of endometrial cancers are either the
less aggressive type I, or endometrioid adenocarcinomas (~80%), or
the more aggressive type II, or serous, adenocarcinomas (~10–15%).
Using a screening panel composed of primary endometrial
endometrioid and endometrial serous tumors we found that CUL5 was
significantly underexpressed, −2.92-fold (p<0.001) in the former
and −4.33-fold (p<0.001) in the latter, compared with benign
endometrium. We also found that overexpression of CUL5 in Ishikawa
H endometrial cancer cells significantly slowed growth.
Furthermore, we showed that microRNA miR-182, which is
significantly overexpressed in this cancer (8) and which is associated with poor
outcome in colorectal adenocarcinoma (9), targets CUL5 in both Ishikawa H and
Hec50co endometrial cancer cells. Finally, overexpression of
wild-type CUL5 in Ishikawa H cells led to diminished cell growth
compared with untransfected cells and lower expression of potential
downstream client proteins such as JAK2 and FAS-L.
Taken together, miR-182, CUL5 and downstream clients
including JAK2 and FAS-L represent a potentially important and
useful set of new therapeutic targets in endometrial cancers.
Materials and methods
Tumor tissue procurement
Primary tumor tissues from 18 primary endometrial
endometrioid adenocarcinomas and 16 primary endometrial serous
adenocarcinomas along with 6 benign endometrium tissues were
obtained under informed consent through the Gynecologic
Malignancies Tissue Repository, housed in the University of Iowa
Hospitals and Clinics, Department of Obstetrics and Gynecology
(IRB#200209010). All tissues are flash frozen specimens for which a
clear histologic diagnosis has been assigned.
Cell culture
Endometrial cancer cells Ishikawa H and Hec50co were
grown and maintained under optimal conditions of Dulbecco's
modified Eagle's medium (DMEM) with 10% fetal bovine serum and 1%
penicillin/streptomycin. These cells were chosen since Ishikawa H
cells are a model for endometrial endometrioid adenocarcinoma
(ER+, PR+, p53 wt, PTEN mut) and Hec50co
cells are a model for endometrial serous adenocarcinoma
(ER−, PR−, p53 mut and PTEN wt) (10).
For the purpose of examining the effects of CUL5
over-expression, we chose the type I cancer cell line Ishikawa H
which represents the great majority of endometrial cancers. A CUL5
expression vector, CMV6-CUL5 (OriGene), was purchased and prepared
for transfection. A stable and reliable CUL5-overexpressing
Ishikawa H subline, CUL5wt35, was produced by selection on G418
(Geneticin), and the identity of the integrated plasmid was
verified by direct sequencing of cellular gDNA.
In order to assess CUL5 targeting by miR-182, both
Ishikawa H and Hec50co cells were transiently transfected with a
miR-182 mimic (Life Technologies). Transfections were carried out
three separate times and total cellular RNA was purified from
untransfected, mock-transfected and transfected cells from each
replicate.
Expression of CUL5 protein as well as other relevant
proteins was assessed via standard western blotting hybridizations.
Anti-CUL5 antibody was purchased from Abcam (ab97280), antibodies
for JAK2 and FAS-L were purchased from Cell Signaling (#3230 and
#4273, respectively) and the anti-β-actin loading control was
purchased from Sigma (A1978).
RNA purification and qPCR assays
Whether from primary tissues or cultured cells,
total cellular RNA was purified using the mirVana miRNA Isolation
kit following the manufacturer's recommendations (Life
Technologies). RNA concentration and purity were determined using a
NanoDrop 1000 spectrophotometer (Thermo Scientific).
Quantitative PCR CUL5 expression assays were
performed on both primary tissue RNAs, and cultured cells following
SuperScript III (Invitrogen) reverse transcription of fixed total
RNA mass (200 ng for tissues and 100 ng for cells). Resulting cDNAs
were amplified using primer sets: CUL5for,
5′-CTCAAGTCAACTCACCCAAAGA-3′ and CUL5rev,
5-GCTGCAAACGTCCAATCAAG-3′; and 18S rRNAfor,
5′-AACTTTCGATGGTAGTCGCCG-3′ and 18S rRNArev,
5′-CCTTGGATGTGGTAGCCGTTT-3′. Amplicons were detected in a Power
SYBR-Green qPCR assay carried out on an Applied Biosystems 7900HT
Real-Time PCR System in the Genomics Division of the Iowa Institute
of Human Genetics (IIHG).
Quantitative PCR miR-182 expression assays were
performed on both primary tissue RNAs and cultured cells following
MultiScribe MuLV reverse transcription (TaqMan MicroRNA Reverse
Transcription kit) of fixed total RNA mass (200 ng for tissues and
100 ng for cells) in the presence of miRNA-specific RT primers
(both from Life Technologies). Resulting cDNAs were amplified using
miRNA-specific TaqMan primers (Life Technologies) also on an
Applied Biosystems 7900HT Real-Time PCR system in the Genomics
Division of the IIHG.
Raw cycle threshold (Ct) values for both CUL5 and
miR-182 were normalized (ΔCt) against standard normalizers; 18S
rRNA for CUL5 and RNU48 for miR-182. Fold-change differences for
all analyses were calculated as 2−ΔΔCt where ΔΔCt =
(Mean ΔCtexperiment − Mean ΔCtcontrol)
(11,12). Statistical significance of
fold-changes was assessed via a standard t-test with unequal
variances (13).
Cell proliferation assay
Cell proliferation was determined by measuring
(3H)-thymidine incorporation into Ishikawa H cells
mock-transfected or overexpressing Cul5 (CUL5wt35) as previously
described (14).
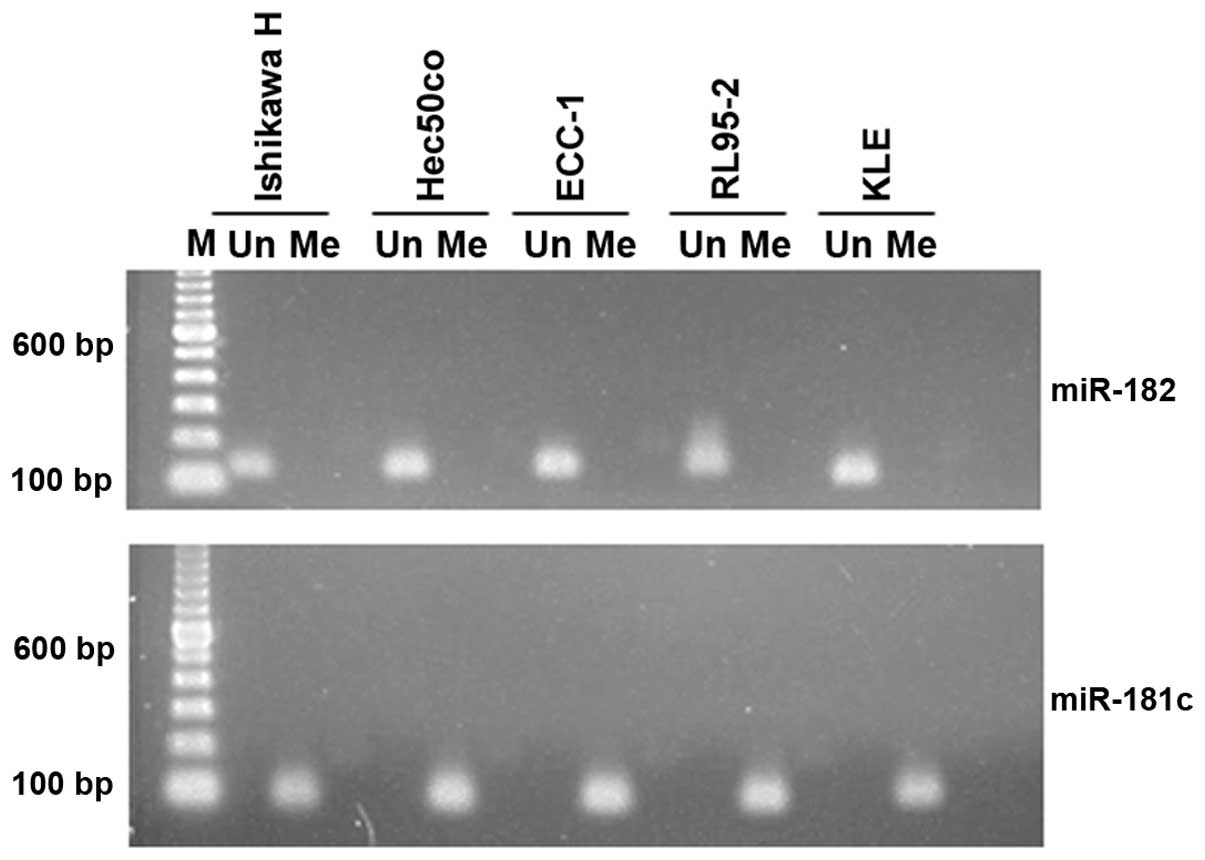
Methylation-specific PCR
In order to assess the methylation state of the
miR-182 promoter, bisulfite conversion of genomic DNA from five
endometrial cancer cell lines (Ishikawa H, Hec50co, ECC-1, RL95-2
and KLE) was carried out using the EZ DNA Methylation-Direct kit
(Zymo Research) according to the manufacturer's recommendations.
Methylation-specific PCR primer sets for the miR-182 promoter were
designed using the MethPrimer package (15). A well-documented hyper-methylated
miRNA promoter (miR-181c) was chosen as a positive control
(16). Previously validated
methylation-specific PCR primers for miR-181c were used following
MethPrimer in silico validation. PCR reactions were carried
out using ZymoTaq™ PreMix (Zymo Research).
Results
CUL5 is significantly underexpressed in
endometrial cancers
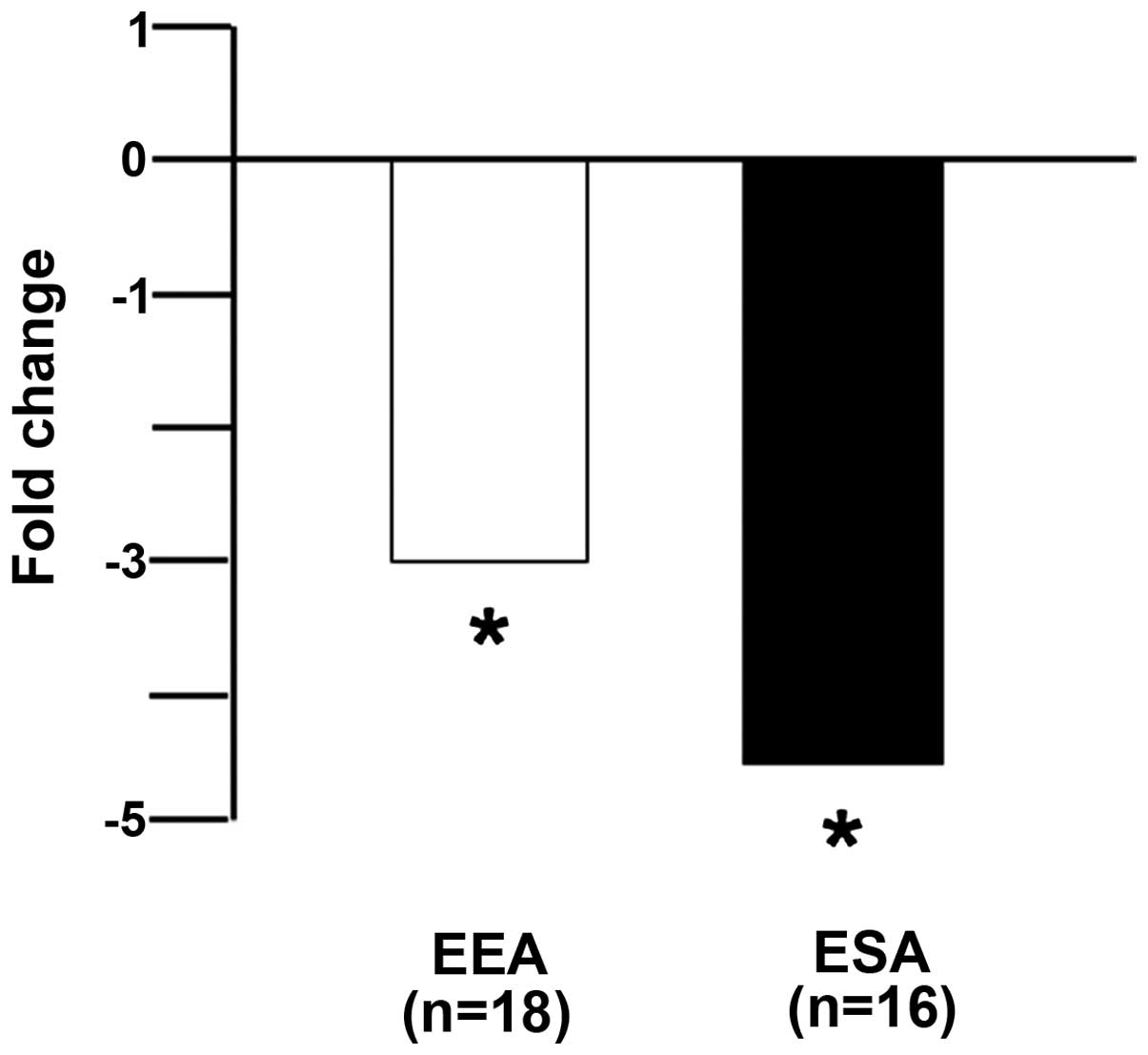
Our primary tumor screening panel was composed of 18
endometrial endometrioid adenocarcinomas, 16 endometrial serous
adenocarcinomas and 6 benign endometrium. The CUL5 qPCR assay
results presented in Fig. 1 showed
that both endometrial endometrioid and endometrial serous
adenocarcinomas displayed significant underexpression (−2.9-fold,
p<0.01; and −4.3-fold, p<0.01, respectively) compared with
the benign endometrium.
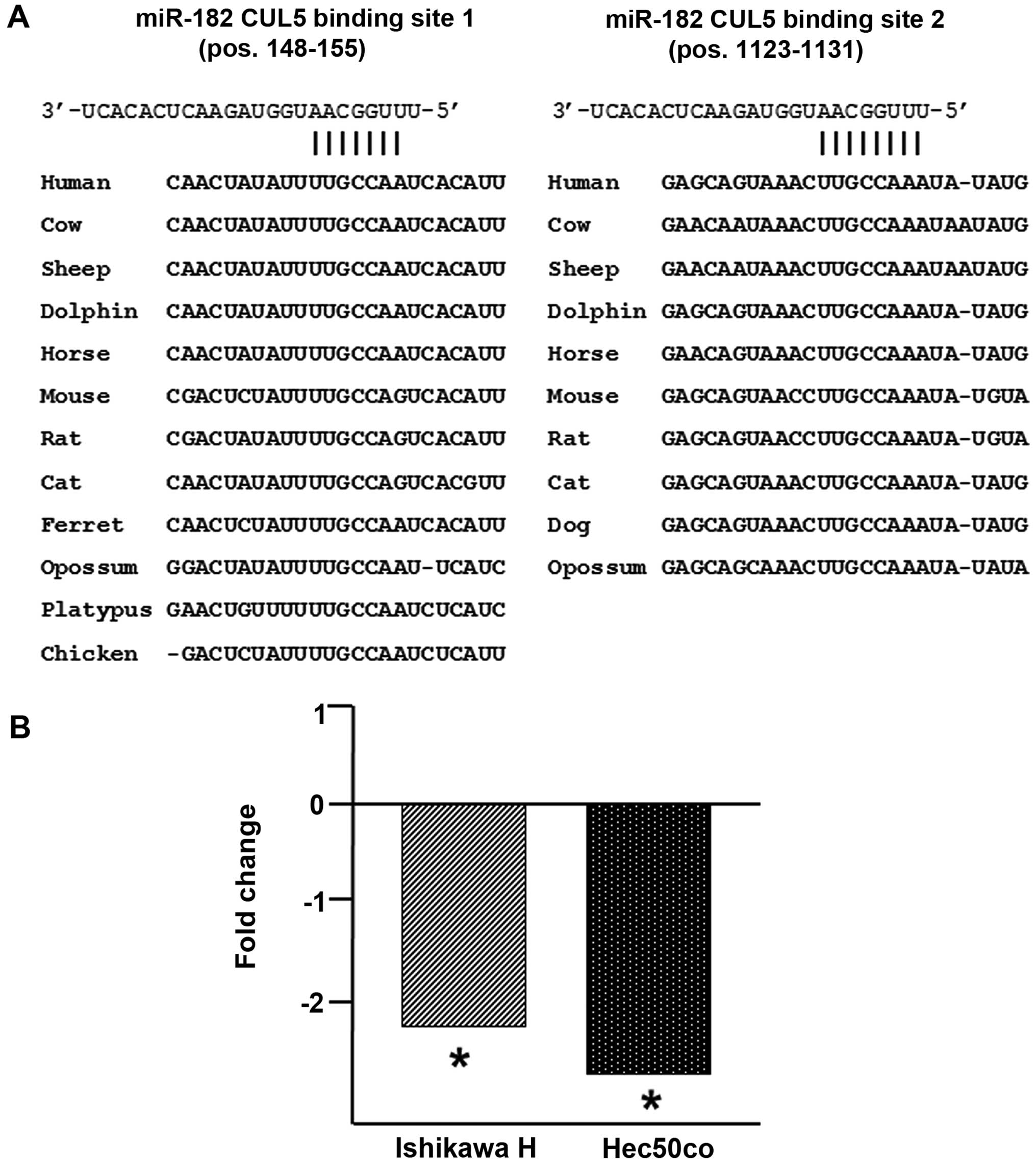
miR-182 targets CUL5
Two prior cancer studies have shown that CUL5 is a
validated target of microRNAs miR-19a/b and miR-7, the former in
cervical and the latter in hepatocellular cancer (4,5). Our
previous study of miRNA expression in endometrial cancers showed
that, while both miR-19a/b and miR-7 display increased expression
relative to benign endometrium, neither miRNA achieves statistical
significance (8). In addition, both
TargetScan 5.2 and PicTar gave high confidence predictions that
CUL5 would be a target of miR-182. This prediction was supported by
an evolutionarily deep conservation of two miR-182 binding sites in
the CUL5 3′-UTR (Fig. 2A).
Furthermore, our previous study (8)
showed that miR-182 was significantly overexpressed in both
endometrial endometrioid and endometrial serous adenocarcinomas
compared with benign endometrium (27.2-fold, p<0.01; and
11.5-fold, p<0.01, respectively) which is consistent with the
significant underexpression of CUL5 in these cancers. We
transiently transfected miR-182 into the type I and II endometrial
cell lines Ishikawa H and Hec50co (10) and observed that CUL5 was
significantly underexpressed (−2.2-fold, p<0.01; and −2.6-fold,
p<0.01, respectively) in these cells when compared with the
mock-transfected cells (Fig. 2B).
Thus, at least in these two cancers, CUL5 is a target of miR-182
such that CUL5 downregulation is achieved, in part, through
modulation of miR-182 expression.
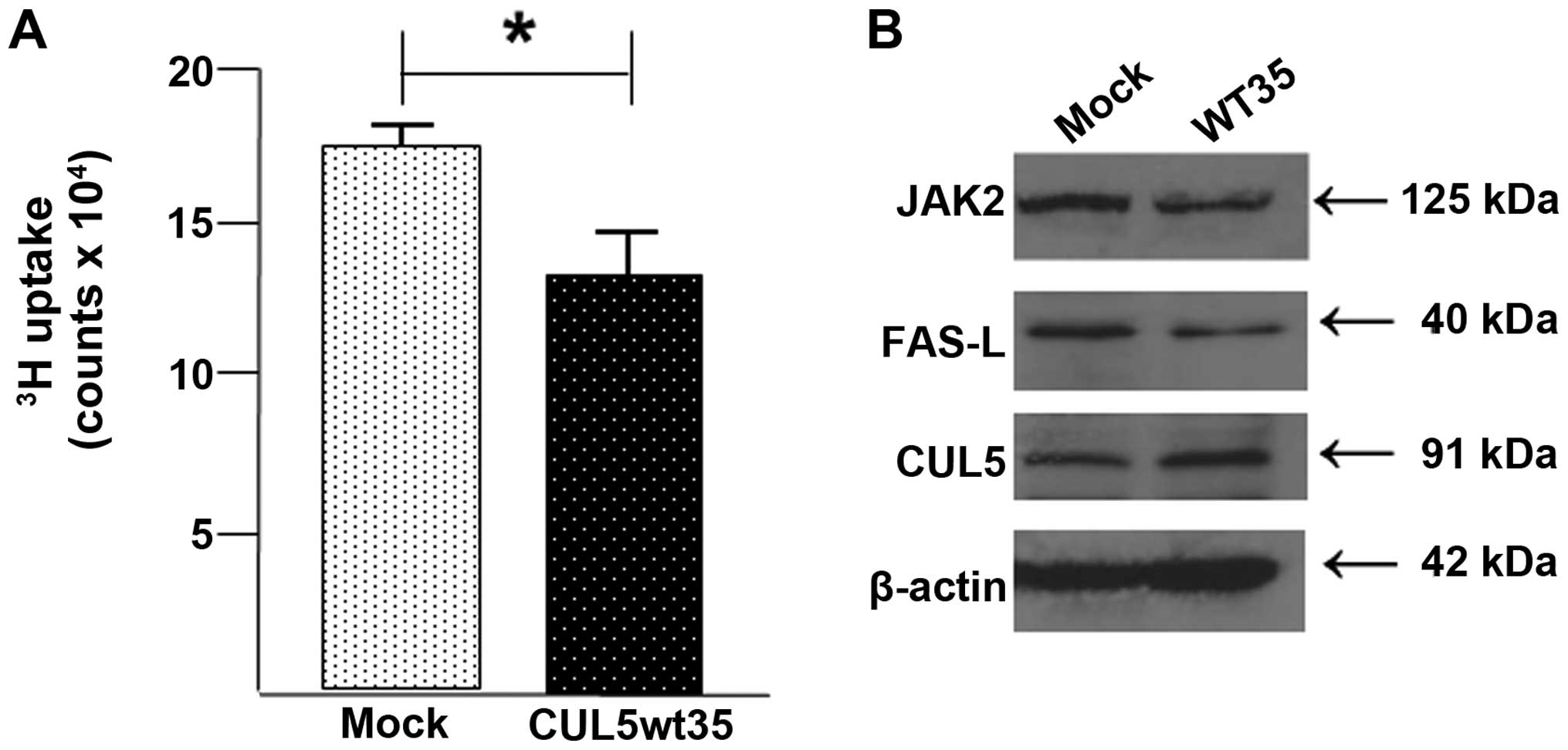
Overexpression of CUL5 inhibits cell
growth
In order to assess the effect of CUL5 on endometrial
cancer cell behavior, we created a stable CUL5-overexpressing
Ishikawa H subcell line, CUL5wt35, using a CMV6 entry vector
containing the CUL5 coding region. Cells were selected on G418. We
measured cell proliferation with a tritiated (3H)
thymidine uptake assay. Results, as presented in Fig. 3A, showed that overexpression of CUL5
significantly reduced proliferation compared with the untransformed
Ishikawa H cells.
Overexpression of CUL5 affects known
downstream 'client' proteins
In order to confirm that both CUL5 mRNA and protein
are overexpressed in CUL5wt35 cells, mRNA levels in the CUL5wt35
and mock-transfected Ishikawa H cells were assayed by SYBR-Green
qPCR and CUL5 protein levels were assessed via western blotting.
The SYBR-Green CUL5 mRNA expression assay showed that there was a
32-fold increase in CUL5 mRNA in the CUL5wt35 cells as compared
with the mock-transfected cells (data not shown). CUL5 protein
expression was also increased in the CUL5wt35 cells compared with
the mock-transfected cells although the increase was more modest
(Fig. 3B). Also shown in Fig. 3B are expression levels of three
proteins proposed to be affected by CUL5 expression.
Discussion
The cullin-RING ligase (CRL) ubiquitin family, first
reported in the late 1990s (1), has
taken a central position in regulatory biology. One of seven
members of the human cullin gene family, the ubiquitin ligase
scaffold protein cullin-5 (CUL5) has been shown to be an important
element in cardiovascular biology (17) and has been implicated in several
types of cancers including breast cancer (2,3). We
examined CUL5 expression in two histologic forms of endometrial
(uterine) cancers; the less aggressive and better prognosis
endometrioid adenocarcinoma and the more aggressive, poorer
prognosis serous adenocarcinoma. We reported in the present study,
that CUL5 expression was significantly less in both histologic
types and that the loss of CUL5 expression was greater in the
serous histologic type than that in the endometrioid histologic
type. This finding suggests that an increased loss of CUL5
expression in endometrial cancers could be a marker for prognosis.
Such a connection will obviously require study of more endometrial
tumors than the 34 tumors we had available for the present study
but it is a question worth pursuing.
In addition to the important mechanistic studies of
CUL5 that have been reported from the laboratory of Dr
Burnatowska-Hledin since their initial discovery of CUL5 in 2000
(18–20), two other studies have demonstrated a
regulatory role for two miRNAs, one, for miR-19a and miR-19b, in
cervical cancer and another, for miR-7, in hepatocellular cancer
(4,5). We searched miRNA target prediction
algorithms for CUL5 and discovered that several, including
TargetScan and PicTar, indicated that miR-182 was an even higher
confidence prediction than either of the others. We confirmed that
there are two highly conserved miR-182 binding sites in the 3′-UTR
of the CUL5 gene and demonstrated in the present study via
quantitative PCR assay that miR-182 was overexpressed in the two
model endometrial cancer cell lines Ishikawa H and Hec50co
(10) and that CUL5 is indeed a
target of this miRNA at least in endometrial cancers. This
conclusion is supported by our previously reported finding that
miR-182 is significantly overexpressed in endometrial
adenocarcinomas (8). In addition,
it has been demonstrated in colorectal adenocarcinomas that high
miR-182 expression is a reliable predictor of a poor prognosis
(9,21), and possibly in prostate cancer as
well (22). Notably, the opposite
relationship has been implied in cervical cancers (23).
Given the proliferative effects of low CUL5
expression reported elsewhere (4,19), we
overexpressed CUL5 in the Ishikawa H endometrial cancer cell line
by creating a stable subcell line, CUL5wt35. We validated the
increased mRNA and protein levels in the CUL5wt35 cells relative to
the mock-transfected Ishikawa H cells and showed that CUL5
over-expression significantly slowed cell proliferation by
measuring 3H-thymidine uptake. This confirmed that CUL5
downregulation, achieved at least in part by miR-182 upregulation,
is a circumstance favorable to endometrial cancer proliferation. As
to the mechanism of miR-182 downregulation, it has been shown in
highly metastatic C8161.9 and WM266-4 melanoma cells that
demethylation using 5-aza-2′-deoxycytidine (DAC) and tricostatin A
resulted in significant miR-182 expression increases (24). We examined the methylation status of
the miR-182 promoter in 5 endometrial cancer cell lines (Ishikawa
H, Hec50co, ECC-1, RL95-2 and KLE) by methylation-specific PCR. All
5 cell lines indicate that the miR-182 promoter is in fact
unmethylated (Fig. 4). While this
mechanism obviously requires additional exploration, it does
suggest that epigenetic modulation to lower miR-182 expression
could be an avenue through which to promote higher CUL5 presence in
endometrial cancers.
Finally, we observed that overexpression of CUL5 was
coupled with modestly lowered expression of two potential CUL5
ubiquitin ligase client proteins, JAK2 and FAS-L. The exact
relationships among these and other possible client proteins are as
yet undetermined but, clearly, loss of CUL5 ubiquitin ligase would
be beneficial in that pro-growth clients would escape targeting.
Several studies have concluded that dysregulation of the
ubiquitination process could be carcinogenic (25). Although CUL5-RING-ligases may not
have a large number of clients, the fact that JAK2 is a client does
implicate the highly pro-growth JAK-STAT pathway (26) in the cascade that starts with
miR-182 promoter demethylation and ends with highly proliferative
endometrial cancer cells. These results present a series of
potential interventional modalities that should be explored,
particularly in more aggressive endometrial cancers.
Acknowledgments
The present study was supported in part by NIH Grant
RO1CA99908 to K.-K.L., NIH Grant K12HD000849 to H.-D.R., the
University of Iowa Carver, College of Medicine, Department of
Obstetrics and Gynecology Research Development Fund, and DOD
Collaborative Undergraduate HBCU Student Summer training program
Awards W81XWH-12-1-0117 and W81XWH-13-1-0178 to B.L. We acknowledge
the Department of Obstetrics and Gynecology, Women's Health Tissue
Repository and Gynecologic Malignancy Repository (Dr Donna
Santillan, Director) and the continued invaluable assistance of the
University of Iowa Institute of Human Genetics, Genomics Facility
(in particular Garry Hauser and Mary Boes).
References
|
1
|
Petroski MD and Deshaies RJ: Function and
regulation of cullin-RING ubiquitin ligases. Nat Rev Mol Cell Biol.
6:9–20. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Burnatowska-Hledin MA, Kossoris JB, Van
Dort CJ, Shearer RL, Zhao P, Murrey DA, Abbott JL, Kan CE and
Barney CC: T47D breast cancer cell growth is inhibited by
expression of VACM-1, a cul-5 gene. Biochem Biophys Res Commun.
319:817–825. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Johnson AE, Le IP, Buchwalter A and
Burnatowska-Hledin MA: Estrogen-dependent growth and estrogen
receptor (ER)-alpha concentration in T47D breast cancer cells are
inhibited by VACM-1, a cul 5 gene. Mol Cell Biochem. 301:13–20.
2007. View Article : Google Scholar
|
|
4
|
Xu XM, Wang XB, Chen MM, Liu T, Li YX, Jia
WH, Liu M, Li X and Tang H: MicroRNA-19a and -19b regulate cervical
carcinoma cell proliferation and invasion by targeting CUL5. Cancer
Lett. 322:148–158. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ma C, Qi Y, Shao L, Liu M, Li X and Tang
H: Downregulation of miR-7 upregulates cullin 5 (CUL5) to
facilitate G1/S transition in human hepatocellular carcinoma cells.
IUBMB Life. 65:1026–1034. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
American Cancer Society Cancer facts &
figures 2014. http://www.cancer.org/acs/groups/content/@research/documents/document/acspc-041770.pdf.
Accessed February 10, 2014.
|
|
7
|
Howlader N, Noone AM, Krapcho M, Garshell
J, Miller D, Altekruse SF, Kosary CL, Yu M, Ruhl J, Tatalovich Z,
et al: SEER Cancer Statistics Review, 1975–2010. National Cancer
Institute; Bethesda, MD, USA: http://seer.cancer.gov/csr/1975_2011/,
based on November 2013 SEER data submission, posted to the SEER
website, April 2014.
|
|
8
|
Devor EJ, Hovey AM, Goodheart MJ,
Ramachandran S and Leslie KK: microRNA expression profiling of
endometrial endometrioid adenocarcinomas and serous adenocarcinomas
reveals profiles containing shared, unique and differentiating
groups of microRNAs. Oncol Rep. 26:995–1002. 2011.PubMed/NCBI
|
|
9
|
Rapti SM, Kontos CK, Papadopoulos IN and
Scorilas A: Enhanced miR-182 transcription is a predictor of poor
overall survival in colorectal adenocarcinoma patients. Clin Chem
Lab Med. 52:1217–1227. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Albitar L, Pickett G, Morgan M, Davies S
and Leslie KK: Models representing type I and type II human
endometrial cancers: Ishikawa H and Hec50co cells. Gynecol Oncol.
106:52–64. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2-ΔΔCT method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
12
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative Ct method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar
|
|
13
|
Snedecor GW and Cochran WG: Statistical
Methods. 8th edition. Iowa State University Press; Ames, IA: pp.
158–160. 1989
|
|
14
|
Stanic B, Katsuyama M and Miller FJ Jr: An
oxidized extracellular oxidation-reduction state increases Nox1
expression and proliferation in vascular smooth muscle cells via
epidermal growth factor receptor activation. Arterioscler Thromb
Vasc Biol. 30:2234–2241. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li LC and Dahiya R: MethPrimer: Designing
primers for methylation PCRs. Bioinformatics. 18:1427–1431. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hashimoto Y, Akiyama Y, Otsubo T, Shimada
S and Yuasa Y: Involvement of epigenetically silenced microRNA-181c
in gastric carcinogenesis. Carcinogenesis. 31:777–784. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Burnatowska-Hledin M and Barney CC: New
insights into the mechanism for VACM-1/cul5 expression in vascular
tissue in vivo. Int Rev Cell Mol Biol. 313:79–101. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Burnatowska-Hledin M, Zhao P, Capps B,
Poel A, Parmelee K, Mungall C, Sharangpani A and Listenberger L:
VACM-1, a cullin gene family member, regulates cellular signaling.
Am J Physiol Cell Physiol. 279:C266–C273. 2000.PubMed/NCBI
|
|
19
|
Van Dort C, Zhao P, Parmelee K, Capps B,
Poel A, Listenberger L, Kossoris J, Wasilevich B, Murrey D, Clare P
and Burnatowska-Hledin M: VACM-1, a cul-5 gene, inhibits cellular
growth by a mechanism that involves MAPK and p53 signaling
pathways. Am J Physiol Cell Physiol. 285:C1386–C1396. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bradley SE, Johnson AE, Le IP, Oosterhouse
E, Hledin MP, Marquez GA and Burnatowska-Hledin M: Phosphorylation
of VACM-1/Cul5 by protein kinase A regulates its neddylation and
antiproliferative effect. J Biol Chem. 285:4883–4895. 2010.
View Article : Google Scholar :
|
|
21
|
Zhang Y, Wang X, Wang Z, Tang H, Fan H and
Guo Q: miR-182 promotes cell growth and invasion by targeting
forkhead box F2 transcription factor in colorectal cancer. Oncol
Rep. 33:2592–2598. 2015.PubMed/NCBI
|
|
22
|
Li Y, Zhang D, Wang X, Yao X, Ye C, Zhang
S, Wang H, Chang C, Xia H, Wang YC, et al: Hypoxia-inducible
miR-182 enhances HIF1α signaling via targeting PHD2 and FIH1 in
prostate cancer. Sci Rep. 5:124952015. View Article : Google Scholar
|
|
23
|
Sun J, Ji J, Huo G, Song Q and Zhang X:
miR-182 induces cervical cancer cell apoptosis through inhibiting
the expression of DNMT3a. Int J Clin Exp Pathol. 8:4755–4763.
2015.PubMed/NCBI
|
|
24
|
Liu S, Howell PM and Riker AI:
Up-regulation of miR-182 expression after epigenetic modulation of
human melanoma cells. Ann Surg Oncol. 20:1745–1752. 2013.
View Article : Google Scholar
|
|
25
|
Zhou MJ, Chen FZ and Chen HC:
Ubiquitination-involved enzymes and cancer. Med Oncol. 31:932014.
View Article : Google Scholar
|
|
26
|
Yu H, Lee H, Herrmann A, Buettner R and
Jove R: Revisiting STAT3 signalling in cancer: New and unexpected
biological functions. Nat Rev Cancer. 14:736–746. 2014. View Article : Google Scholar : PubMed/NCBI
|