Introduction
Colon cancer is one of the most common malignancies
of the digestive system and is the leading cause of mortality from
cancer worldwide (1). Treatment of
colon cancer includes surgery resection, chemotherapy, radiotherapy
and targeted therapy, alone or in combination. To date,
chemotherapy is still the major treatment for colon cancer, yet the
serious side-effects associated with chemotherapy drugs greatly
reduce the life quality of the patients (2). Hence, there is a great clinical need
to explore new chemotherapy drugs for colon cancer treatment with
low toxicity. Natural products or their derivates are one of the
major sources of chemotherapy drugs, and some have been used for
cancer treatment for decades, such as vincristine, taxol and
camptothecin (3,4). Oridonin (ORI) is extracted from the
Chinese herb Rabdosia rubescens and/or related species
(5). Increasing evidence suggests
that ORI is capable of inhibiting growth and inducing apoptosis in
several types of cancer cells, such as breast and colon cancer,
leukemia, lymphoma, pancreatic cancer and osteosarcoma (5,6). Our
previous study also confirmed the anticancer activity of ORI in
colon cancer cells. However, the exact mechanisms underlying this
effect remain unclear.
Bone morphogenetic proteins (BMPs), belonging to the
TGF-β superfamily, consist of one of the important pathways to
regulate development, and aberrant signaling transduction is one of
the main causes for colon cancer (7–9). It
has been reported that BMP2 can inhibit the proliferation of
colorectal cancer cells (10), and
BMP4 can induce differentiation in colorectal cancer stem cells, as
well as increase their response to chemotherapy drugs (11). BMP7, another important member of the
BMP family, has been approved for the treatment of bone fracture
healing and spine surgery due to its excellent osteogenic activity
(12). Apart from the osteogenic
differentiation activity, BMP7 is involved in brown fat cell
development and thermogenesis (13). Other studies indicated that BMP7 is
also implicated in cancer (14),
although its role in cancer needs to be further investigated.
Mitogen-activated protein kinases (MAPKs) are critical mediators
for signaling transduction, and respond to a wide range of
extracellular stresses such as UV radiation, hypoxia, and oxidative
stress (15). p38 MAPK, one class
of MAPKs, is involved in regulating cell proliferation, apoptosis,
and autophagy (15). Studies have
indicated that p38 MAPK activation is involved in the anticancer
effect of ORI in pancreatic cancer (16,17).
Various functions of BMP7 are also mediated by p38 MAPK,
upregulation or suppression (18,19).
To date, it remains unknown whether BMP7 is associated with the
anticancer activity of ORI in colon cancer cells.
In the present study, we investigated the anticancer
effect of ORI in colon cancer, and dissected the possible
mechanisms underlying this anticancer activity of ORI in human
colon cancer cells.
Materials and methods
Chemicals and drug preparations
ORI was obtained from Hao-Xuan Bio-Tech Co., Ltd.
(Xi'an, China). The HCT116 cell line was obtained from the American
Type Culture Collection (ATCC; Manassas, VA, USA). ORI was prepared
with dimethyl sulfoxide (DMSO) or 0.4% carboxymethylcellulose
sodium (CMC-Na) as a suspension for the in vitro or in
vivo experiments, respectively. The p38 MAPK inhibitor SB203580
(#S1076) was obtained from Selleckchem (Houston, TX, USA). All
antibodies were purchased from Santa Cruz Biotechnology (Santa
Cruz, CA, USA). Cells were cultured in Dulbecco's modified Eagle's
medium (DMEM) with 10% fetal bovine serum (FBS), 100 U/ml of
penicillin and 100 µg/ml of streptomycin at 37°C in 5%
CO2.
Cell proliferation and viability
assays
Cell proliferation and viability were assessed with
the crystal violet staining assay. HCT116 cells were plated in a
24-well plate and then treated with different concentrations of
ORI; DMSO was used as a solvent control. The cells were washed
carefully with cold (4°C) phosphate-buffered saline (PBS) and
stained with 0.5% crystal violet formalin solution at room
temperature. For quantification, the crystal violet was dissolved
with 1 ml 20% acetic acid at room temperature for 20 min with
shaking. The absorbance at 570 nm was then measured. Each assay was
carried out in triplicate.
Construction of the BMP7 recombinant
adenovirus
Recombinant adenoviruses expressing BMP7 were
constructed with the AdEasy system (20), tagged with green fluorescence
protein (GFP) and designated as AdBMP7. The recombinant adenovirus
expressing GFP only was used as a vector control.
Flow cytometric analysis of apoptosis and
cell cycle distribution
HCT116 cells were plated in a 6-well plate. The
cells were treated with different concentrations of ORI or DMSO.
Then, the cells were harvested and washed with cold PBS, followed
by incubating with Annexin V-EGFP and propidium iodide (PI)
following the instructions in the kits (KeyGen Biotech, Nanjing,
China). Then, the cells were analyzed by fluorescence-activated
cell sorting (FACS) for apoptosis. For the cell cycle assay, HCT116
cells were treated with different concentrations of ORI or DMSO. At
48 h after treatment, the cells were collected and washed with PBS,
fixed with cold (4°C) 70% ethanol, and washed with 50%, 30% ethanol
and PBS, successively. Then, the cells were stained with 1 ml of PI
(20 mg/ml) containing RNase (1 mg/ml) in PBS for 30 min, and
subjected to FACS analysis. Each assay was carried out in
triplicate.
Reverse transcription and polymerase
chain reaction analysis (RT-PCR)
Subconfluent HCT116 cells were plated in T25 flasks
and treated with different concentrations of ORI or DMSO. Total RNA
was extracted with TRIzol reagent (Invitrogen, USA) and subjected
to RT reaction to generate cDNA. Then, the cDNA was used as a
template for detecting the expression level of target genes with
PCR. The primer sequences are available upon request. Each assay
was carried out in triplicate.
Western blot assay
HCT116 cells were plated in a 6-well plate and
treated with different concentrations of ORI or DMSO. The medium
was discarded, and the cells were washed with cold PBS. The cells
were lysed with 300 µl lysis buffer in each well, and the lysate
was boiled for 10 min. Then, the samples were subjected to SDS-PAGE
separation and transferred to polyvinylidene fluoride (PVDF)
membranes, which were immunoblotted with primary antibodies and
HRP-conjugated secondary antibodies sequentially. The target
proteins were developed with SuperSignal West Pico Substrate
(Pierce, Rockford, IL, USA). Each assay was carried out in
triplicate.
Statistical analysis
All quantitative data are expressed as mean ± SD.
Statistical significance between vehicle vs. drug treatment was
determined by the Student's t-test. A value of p<0.05 was
considered to indicate a statistically significant result.
Results
ORI inhibits the proliferation of HCT116
cells
Evidence indicates that ORI shows an
antiproliferative effect in various types of cancer cells. Thus, we
tested this effect of ORI on HCT116 cells to validate whether ORI
could be used as a chemotherapeutic drug for human colon cancer.
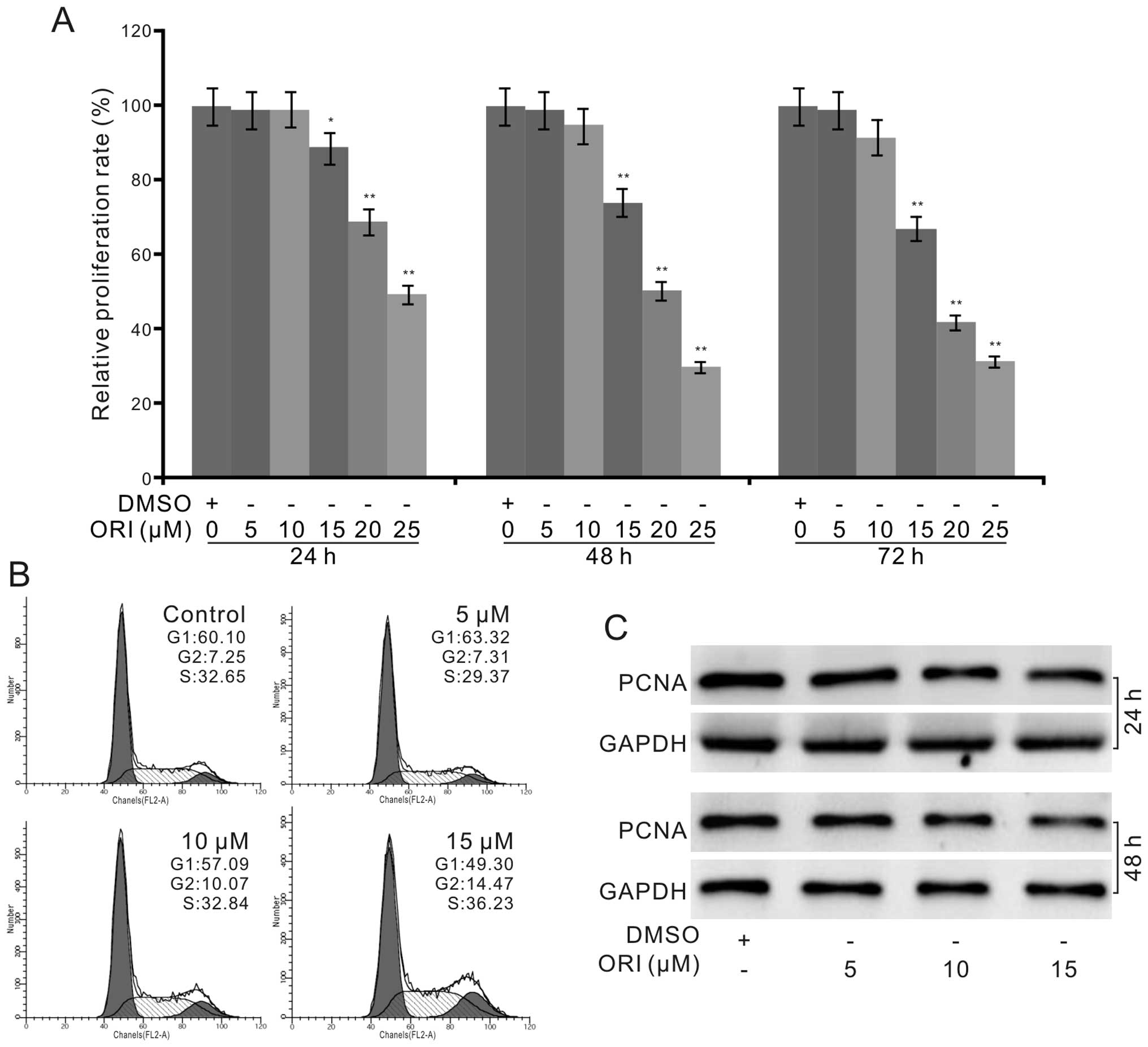
The crystal violet staining assay results showed that ORI
effectively inhibited the proliferation of the HCT116 cells in a
time- and concentration-dependent manner (Fig. 1A). Cell cycle analysis results
showed that ORI induced cell cycle arrest at the G2 phase in the
HCT116 cells (Fig. 1B). Western
blot assay results showed that ORI also decreased the level of
proliferating cell nuclear antigen (PCNA) in the HCT116 cells
(Fig. 1C). These data indicate that
ORI can inhibit the proliferation of HCT116 cells, and may be a
potential chemotherapeutic agent for colon cancer.
ORI induces HCT116 cells to undergo
apoptosis
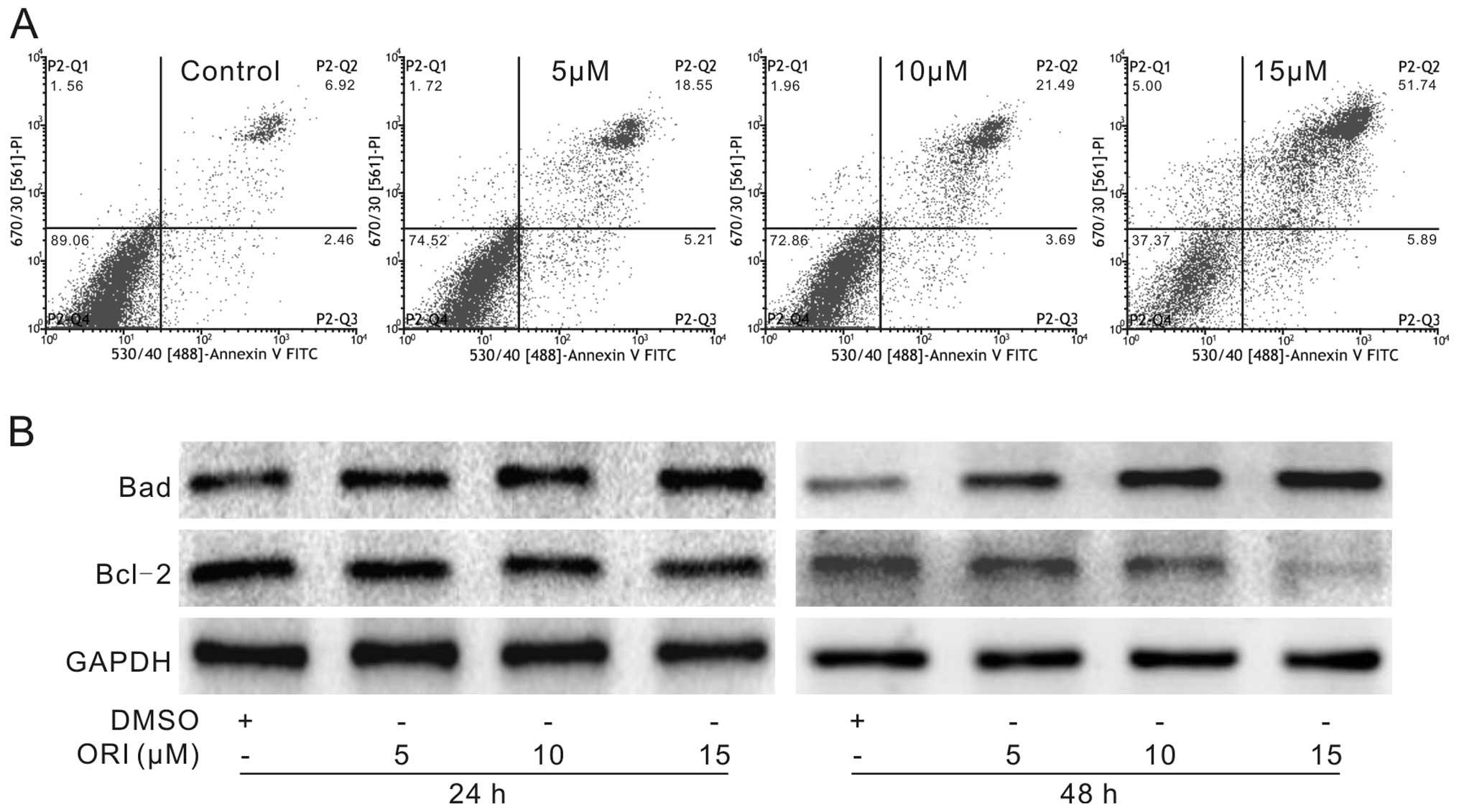
Next, we analyzed whether ORI induces apoptosis in
HCT116 cells. HCT116 cells were seeded in a 6-well plate and were
treated with different concentrations of ORI or DMSO. Then, the
cells were harvested for flow cytometric analysis or lysed for
western blot assay. The results showed that ORI increased the
percentage of apoptotic cells in a concentration-dependent manner
(Fig. 2A). Western blot assay
results showed that ORI notably increased the level of Bad and
reduced the level of Bcl-2 in the HCT116 cells (Fig. 2B). These findings demonstrate that
ORI may be an effective inducer of apoptosis in colon cancer
cells.
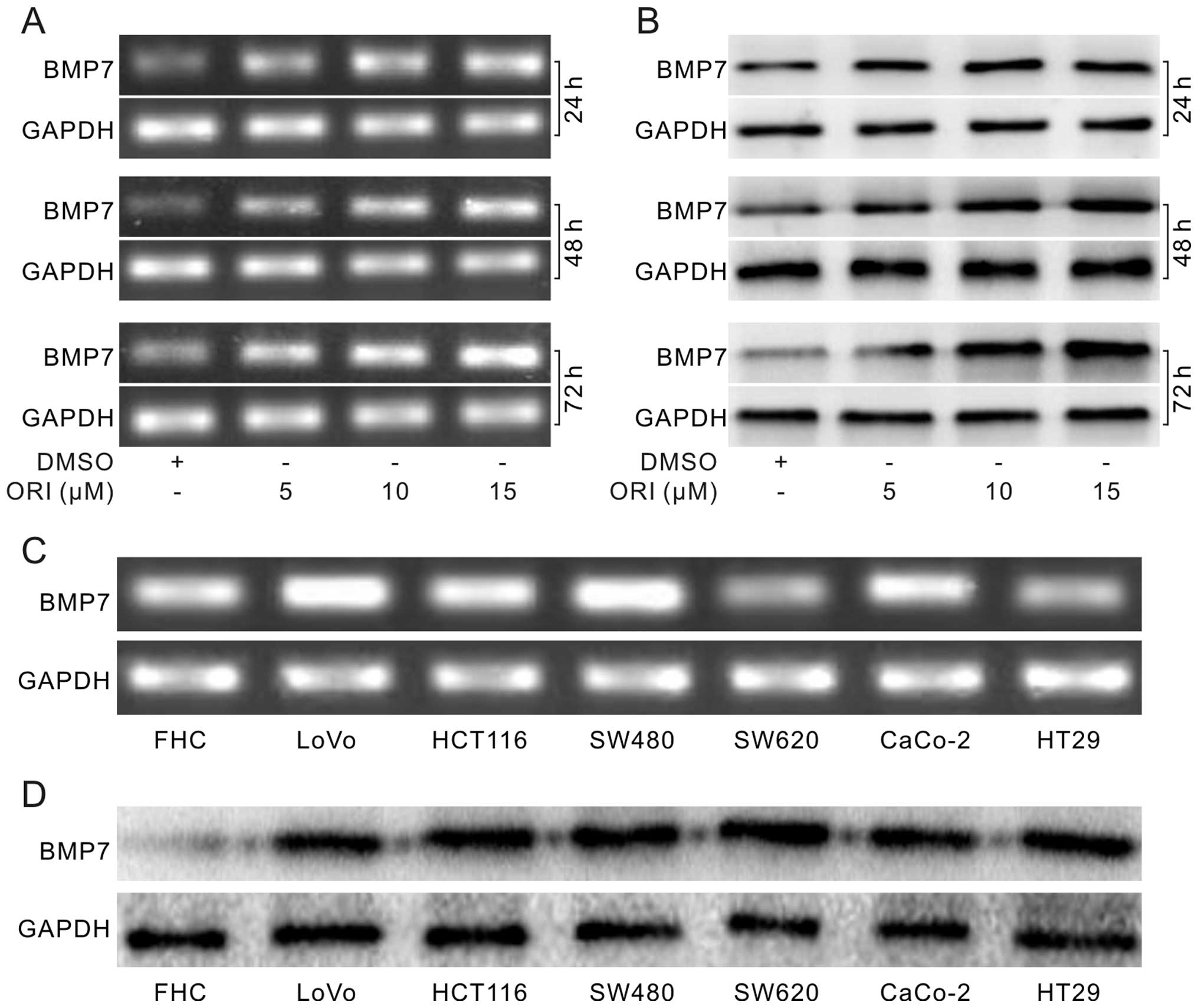
ORI upregulates BMP7 in HCT116 cells
The TGF-β signaling pathway is important for the
development of the digestive system, and aberrant TGF-β signaling
is one of the main etiologies of colon cancer (21). The PCR assay results showed that ORI
increased the mRNA level of BMP7 substantially (Fig. 3A). Western blot assay results
confirmed that ORI increased the level of BMP7 in a concentration-
and time-dependent manner (Fig.
3B). Furthermore, PCR (Fig. 3C)
and western blot assay results (Fig.
3D) showed that endogenous expression of BMP7 in colon cancer
cell lines was higher than that in the FHC cells. These data
demonstrate that ORI can upregulate BMP7, and it may play an
essential role in the antiproliferative effect of ORI in colon
cancer cells.
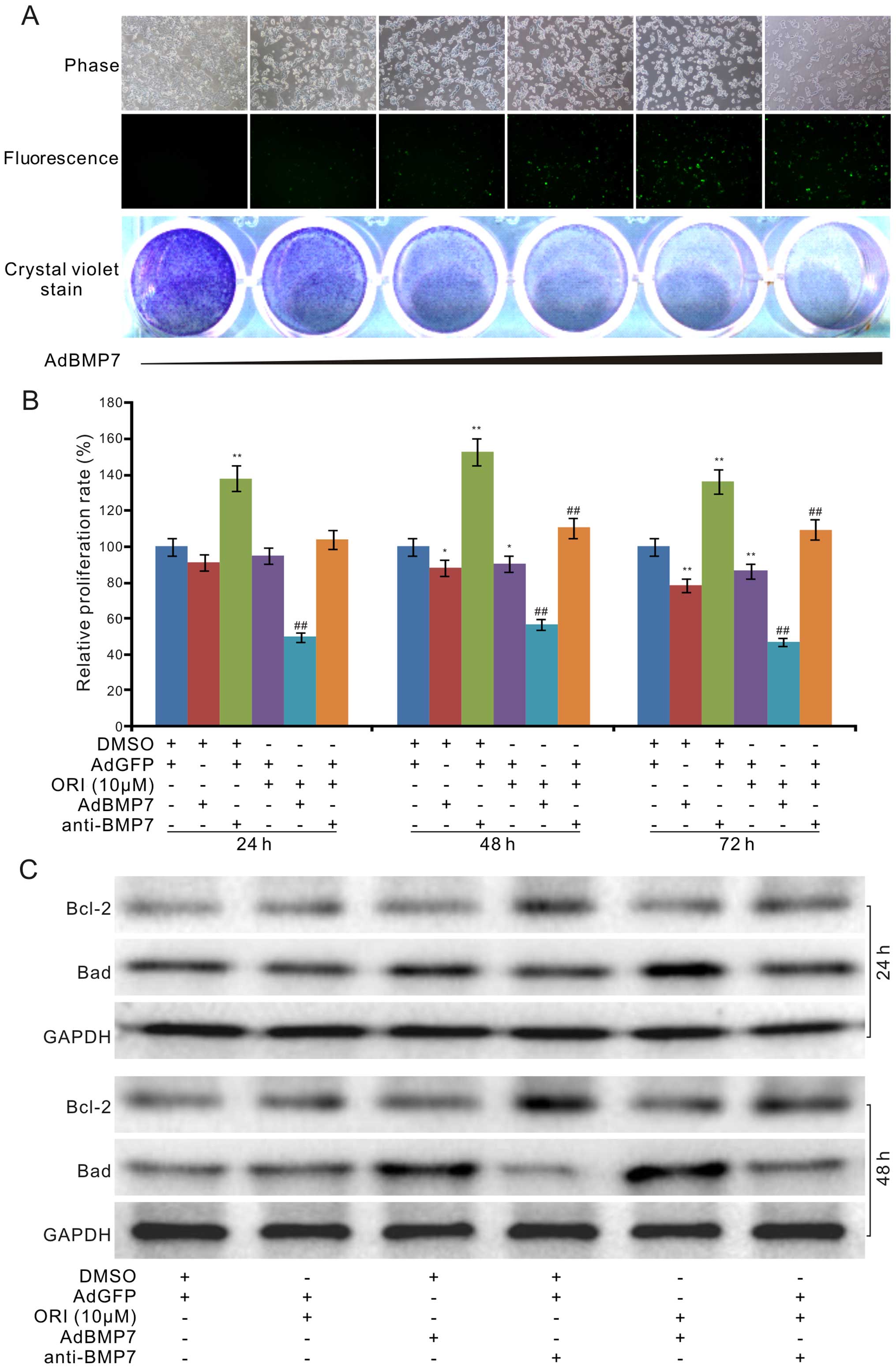
BMP7 partly mediates the
antiproliferative effect of ORI in HCT116 cells
We next investigated the influence of BMP7 on the
antiproliferative effect of ORI in colon cancer cells. We
introduced recombinant adenoviruses of BMP7 (AdBMP7) for exogenous
expression of BMP7 and the BMP7-specific antibody for BMP7
immunodepletion. The results showed that exogenous expression of
BMP7 inhibited the proliferation of the HCT116 cells (Fig. 4A). Crystal violet staining analysis
results indicated that ORI combined with exogenous expression of
BMP7 substantially enhanced the antiproliferative effect of ORI,
while the BMP7 antibody (4 ng/ml) antibody almost reversed the
antiproliferative effect of ORI (10 µM) in the HCT116 cells
(Fig. 4B). Further results showed
that exogenous expression of BMP7 increased the protein level of
Bad upregulated by ORI, but potentiated the effect of ORI on
decreasing the level of Bcl-2; on the contrary, the BMP7 antibody
attenuated the effects of ORI on Bad and Bcl-2, respectively
(Fig. 4C). These results indicate
that upregulation of BMP7 may mediate the antiproliferative effect
of ORI in colon cancer cells.
p38 MAPK mediates the antiproliferative
effect of BMP7 in HCT116 cells
Although exogenous expression of BMP7 enhanced the
antiproliferative effect of ORI in HCT116 cells, the mechanisms
which mediated this effect remain unknown. Western blot analysis
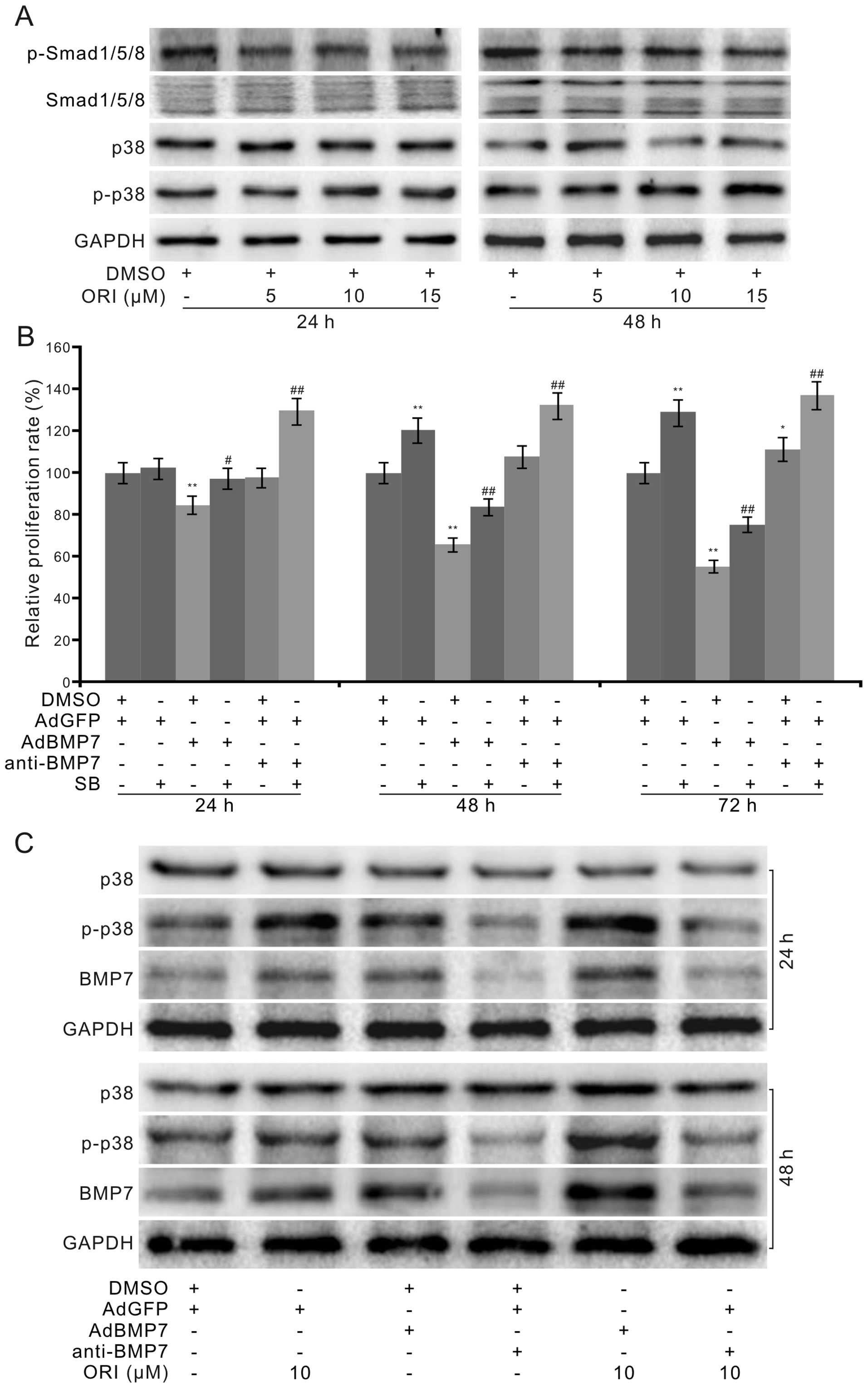
showed that ORI exerted no substantial effect on the
phosphorylation of Smad1/5/8 (p-Smad1/5/8), but increased the
phosphorylation of p38 MAPK (p-p38) in a concentration-dependent
manner (Fig. 5A). These results
indicate that the antiproliferative effect of ORI may not be
mediated by BMP7 through canonical BMP/Smad signaling at least.
Further crystal violet staining assay results showed that the p38
MAPK inhibitor (SB203580, SB) promoted the proliferation of HCT116
cells, attenuated the antiproliferative effect of BMP7, and
synergistically enhanced the proliferation of HCT116 cells when
combined with the BMP7 antibody (Fig.
5B). These data imply that p38 MAPK may be critical for BMP7 to
mediate the antiproliferative effect of ORI in HCT116 cells. The
subsequent western blot assay results showed that BMP7 increased
p38 MAPK phosphorylation, and the BMP7 antibody apparently
inhibited p38 MAPK phosphorylation. The combination of BMP7 and ORI
markedly increased the level of p38 MAPK phosphorylation, while the
BMP7 antibody combined with ORI decreased it apparently (Fig. 5C). These results indicated that BMP7
may mediate the antiproliferative effect of ORI in part by
activating p38 MAPK signaling.
Discussion
In the present study, we demonstrated that ORI may
be an excellent candidate chemotherapeutic agent for colon cancer
treatment. Mechanistically, we found that the anticancer activity
of ORI may be mediated though the upregulation of BMP7 to activate
p38 MAPK signaling.
ORI is extracted from the Chinese traditional
medicinal herb Rabdosia rubescens. As a diterpenoid
compound, ORI exhibits various pharmacological functions and has
been used for many years as an antitumor, anti-microbial,
anti-inflammatory and antioxidant agent (5). Studies indicate that ORI exhibits
antiproliferation and apoptosis-inducing effects in many types of
cancer cells, such as lymphoma, breast cancer, leukemia, colon and
lung cancer (22–26). The present study also demonstrated
this anticancer effect of ORI in colon cancer cells. Several
molecular mechanisms have been reported to be involved in this
effect, such as blockage of extracellular signal-regulated kinase
(ERK) and PI3K/Akt, activation of p53 and p38 MAPK, increase in
hydrogen peroxide and suppression of Wnt/β-catenin signaling
(6,16, 26–28).
However, to date, the precise mechanisms mediating the anticancer
activity of ORI remain unclear.
The pathogenesis of colon cancer includes the
aberrant transduction of various signaling pathways, such as TGF-β,
Wnt/β-catenin, PI3K/Akt (21,29).
BMP7 belongs to the TGF-β superfamily and is also known as
osteogenic protein-1 (OP-1). It was discovered by Marshal Urist in
1965. As one of the members of the BMPs, BMP7 can effectively
commit the precursor cells to osteoblastic lineage. The FDA has
approved the clinical usage of BMP7 for the treatment of
bone-related disease, such as bone fracture healing and spine
surgery (12). Besides osteogenic
differentiation, BMP7 has many other physiological functions, such
as the regulation of brown fat cells (13) and appetite (30). In fact, it has been reported that
the expression of BMP7 is associated with cancer development,
metastasis and invasion (31–35).
BMP7 inhibits TGFβ1-related epithelial-mesenchymal transition (EMT)
in breast cancer cells (36), but
induces EMT in prostate cancer cells (37). Expression of BMP7 was found to be
higher in cancerous tissues than that in normal tissues, and a high
level of BMP7 is related with the recurrence of bladder cancer
(31), poor prognosis in
hepatocellular carcinoma (32,38),
secondary drug-resistance in mantle cell lymphoma (39), and bone metastasis of lung cancer
(34). BMP7 may also be a target
for MYC to promote cell survival in childhood medulloblastoma
(40). BMP7 was detectable in about
50% of gastric cancer, and may be used as a strong marker for tumor
recurrence (33). Hence, BMP7 may
be a potential target for cancer treatment. However, it was also
reported that BMP7 exerts a protective effect in intestinal
epithelial cells from lesions of precancerous and inflammatory
bowel diseases (14), and
suppresses the growth of colon cancer cells in a
Smad4-dependent-manner (9).
Therefore, the role of BMP7 in colon cancer is controversial, and
may be different in specific cell types and the microenvironment.
Our data showed that the expression of BMP7 in colon cancer cells
was higher than that in FHC cells (Fig.
3C and D). ORI increased the level of BMP7, and exogenous
expression of BMP7 enhanced the antiproliferative effect of ORI in
HCT116 cells. The BMP7 antibody partly reversed the inhibitory
effect of ORI on cell proliferation (Fig. 4B). Thus, our findings suggest that
elevated BMP7 may mediate the antiproliferative effect of ORI in
colon cancer cells.
BMP7 exerts its physiological function through the
BMP/Smad pathway (41), namely
canonical BMP/Smad signaling pathway, or non-canonical BMP/Smad
signaling pathway, such as PI3K/Akt and MAPKs (18,37,42).
We found that ORI did not increase the phosphorylation of Smad1/5/8
(Fig. 5A), although BMP7 was
upregulated. This finding implied that the proliferation inhibitory
effect of ORI may not be mediated through the BMP/Smad signaling
pathway, which is consistent with the report that BMP7 exerts the
antiproliferative effect in a Smad4-independent pathway (14). It has been reported that BMP7 can
activate or suppress p38 MAPK signaling (18,19),
and the anticancer activity of ORI may contribute to the activation
of p38 MAPK (16). Thus, we
speculated that the effect of BMP7 on the antiproliferative
activity of ORI in colon cancer cells may be associated with p38
MAPK activation. Western blot assay showed that ORI activated p38
MAPK in the HCT116 cells (Fig. 5A).
The p38 MAPK inhibitor partly reversed the growth inhibitory effect
of ORI, but synergistically promoted the proliferation-enhancing
effect of the BMP7 antibody in the HCT116 cells (Fig. 5B). Therefore, activation of p38 MAPK
may be essential for BMP7 to mediate the anticancer effect of ORI
in colon cancer cells. Our further investigation demonstrated that
BMP7 increased the level of p-p38 MAPK, and synergistically
elevated the ORI-induced p-p38 MAPK, while the BMP7 antibody
markedly decreased the level of p-p38 MAPK affected by ORI in
HCT116 cells. All these data suggest that BMP7 may partly mediate
the antiproliferative effect of ORI by activating p38 MAPK in colon
cancer cells.
In summary, our findings demonstrated that ORI may
be an excellent candidate chemotherapy drug for colon cancer. The
anticancer activity of ORI may be mediated by upregulating BMP7 to
activate p38 MAPK in colon cancer cells. However, the molecular
mechanisms of how ORI upregulate BMP7 need to be further
investigated.
Acknowledgments
We thank Professor Tong-Chuan He of the University
of Chicago Medical Center (Chicago, IL, USA) for his kind provision
of the recombinant adenoviruses. The present study was partly
supported by a research grant from the Natural Science Foundation
of China (grants no. NSFC 81372120 and 81572226 to Bai-Cheng
He).
References
|
1
|
Applegate CC and Lane MA: Role of
retinoids in the prevention and treatment of colorectal cancer.
World J Gastrointest Oncol. 7:184–203. 2015.PubMed/NCBI
|
|
2
|
Gustavsson B, Carlsson G, Machover D,
Petrelli N, Roth A, Schmoll HJ, Tveit KM and Gibson F: A review of
the evolution of systemic chemotherapy in the management of
colorectal cancer. Clin Colorectal Cancer. 14:1–10. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Banjerdpongchai R, Chanwikruy Y,
Rattanapanone V and Sripanidkulchai B: Induction of apoptosis in
the human leukemic U937 cell line by Kaempferia parviflora
Wall.ex.Baker extract and effects of paclitaxel and camptothecin.
Asian Pac J Cancer Prev. 10:1137–1140. 2009.PubMed/NCBI
|
|
4
|
Weaver BA: How Taxol/paclitaxel kills
cancer cells. Mol Biol Cell. 25:2677–2681. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Owona BA and Schluesener HJ: Molecular
insight in the multifunctional effects of oridonin. Drugs R D.
15:233–244. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu Y, Liu YZ, Zhang RX, Wang X, Meng ZJ,
Huang J, Wu K, Luo JY, Zuo GW, Chen L, et al: Oridonin inhibits the
proliferation of human osteosarcoma cells by suppressing
Wnt/β-catenin signaling. Int J Oncol. 45:795–803. 2014.PubMed/NCBI
|
|
7
|
Voorneveld PW, Kodach LL, Jacobs RJ, Liv
N, Zonnevylle AC, Hoogenboom JP, Biemond I, Verspaget HW, Hommes
DW, de Rooij K, et al: Loss of SMAD4 alters BMP signaling to
promote colorectal cancer cell metastasis via activation of Rho and
ROCK. Gastroenterology. 147:196–208.e13. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bertrand FE, Angus CW, Partis WJ and
Sigounas G: Developmental pathways in colon cancer: Crosstalk
between WNT, BMP, Hedgehog and Notch. Cell Cycle. 11:4344–4351.
2012. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Beck SE, Jung BH, Fiorino A, Gomez J,
Rosario ED, Cabrera BL, Huang SC, Chow JY and Carethers JM: Bone
morphogenetic protein signaling and growth suppression in colon
cancer. Am J Physiol Gastrointest Liver Physiol. 291:G135–G145.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang Y, Chen X, Qiao M, Zhang BQ, Wang N,
Zhang Z, Liao Z, Zeng L, Deng Y, Deng F, et al: Bone morphogenetic
protein 2 inhibits the proliferation and growth of human colorectal
cancer cells. Oncol Rep. 32:1013–1020. 2014.PubMed/NCBI
|
|
11
|
Lombardo Y, Scopelliti A, Cammareri P,
Todaro M, Iovino F, Ricci-Vitiani L, Gulotta G, Dieli F, de Maria R
and Stassi G: Bone morphogenetic protein 4 induces differentiation
of colorectal cancer stem cells and increases their response to
chemotherapy in mice. Gastroenterology. 140:297–309. 2011.
View Article : Google Scholar
|
|
12
|
Kamiya N, Ye L, Kobayashi T, Lucas DJ,
Mochida Y, Yamauchi M, Kronenberg HM, Feng JQ and Mishina Y:
Disruption of BMP signaling in osteoblasts through type IA receptor
(BMPRIA) increases bone mass. J Bone Miner Res. 23:2007–2017. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Richard D and Picard F: Brown fat biology
and thermogenesis. Front Biosci. 16:1233–1260. 2011. View Article : Google Scholar
|
|
14
|
Grijelmo C, Rodrigue C, Svrcek M, Bruyneel
E, Hendrix A, de Wever O and Gespach C: Proinvasive activity of
BMP-7 through SMAD4/src-independent and ERK/Rac/JNK-dependent
signaling pathways in colon cancer cells. Cell Signal.
19:1722–1732. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tormos AM, Taléns-Visconti R, Nebreda AR
and Sastre J: p38 MAPK: A dual role in hepatocyte proliferation
through reactive oxygen species. Free Radic Res. 47:905–916. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bu HQ, Liu DL, Wei WT, Chen L, Huang H, Li
Y and Cui JH: Oridonin induces apoptosis in SW1990 pancreatic
cancer cells via p53- and caspase-dependent induction of p38 MAPK.
Oncol Rep. 31:975–982. 2014.
|
|
17
|
Bu HQ, Luo J, Chen H, Zhang JH, Li HH, Guo
HC, Wang ZH and Lin SZ: Oridonin enhances antitumor activity of
gemcitabine in pancreatic cancer through MAPK-p38 signaling
pathway. Int J Oncol. 41:949–958. 2012.PubMed/NCBI
|
|
18
|
Guan J, Li H, Lv T, Chen D, Yuan Y and Qu
S: Bone morphogenic protein-7 contributes to cerebral ischemic
preconditioning induced-ischemic tolerance by activating p38
mitogen-activated protein kinase signaling pathway. Inflammation.
37:1289–1296. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Takahashi M, Otsuka F, Miyoshi T, Otani H,
Goto J, Yamashita M, Ogura T, Makino H and Doihara H: Bone
morphogenetic protein 6 (BMP6) and BMP7 inhibit estrogen-induced
proliferation of breast cancer cells by suppressing p38
mitogen-activated protein kinase activation. J Endocrinol.
199:445–455. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Luo J, Deng ZL, Luo X, Tang N, Song WX,
Chen J, Sharff KA, Luu HH, Haydon RC, Kinzler KW, et al: A protocol
for rapid generation of recombinant adenoviruses using the AdEasy
system. Nat Protoc. 2:1236–1247. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mishra L, Shetty K, Tang Y, Stuart A and
Byers SW: The role of TGF-beta and Wnt signaling in
gastrointestinal stem cells and cancer. Oncogene. 24:5775–5789.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu YQ, Mu ZQ, You S, Tashiro S, Onodera S
and Ikejima T: Fas/FasL signaling allows extracelluar-signal
regulated kinase to regulate cytochrome c release in
oridonin-induced apoptotic U937 cells. Biol Pharm Bull.
29:1873–1879. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hsieh TC, Wijeratne EK, Liang JY,
Gunatilaka AL and Wu JM: Differential control of growth, cell cycle
progression, and expression of NF-κB in human breast cancer cells
MCF-7, MCF-10A, and MDA-MB-231 by ponicidin and oridonin,
diterpenoids from the Chinese herb Rabdosia rubescens. Biochem
Biophys Res Commun. 337:224–231. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhou GB, Kang H, Wang L, Gao L, Liu P, Xie
J, Zhang FX, Weng XQ, Shen ZX, Chen J, et al: Oridonin, a
diterpenoid extracted from medicinal herbs, targets AML1-ETO fusion
protein and shows potent antitumor activity with low adverse
effects on t(8;21) leukemia in vitro and in vivo. Blood.
109:3441–3450. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhu Y, Xie L, Chen G, Wang H and Zhang R:
Effects of oridonin on proliferation of HT29 human colon carcinoma
cell lines both in vitro and in vivo in mice. Pharmazie.
62:439–444. 2007.PubMed/NCBI
|
|
26
|
Li D, Wu LJ, Tashiro S, Onodera S and
Ikejima T: Oridonin-induced A431 cell apoptosis partially through
blockage of the Ras/Raf/ERK signal pathway. J Pharmacol Sci.
103:56–66. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hu HZ, Yang YB, Xu XD, Shen HW, Shu YM,
Ren Z, Li XM, Shen HM and Zeng HT: Oridonin induces apoptosis via
PI3K/Akt pathway in cervical carcinoma HeLa cell line. Acta
Pharmacol Sin. 28:1819–1826. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gao FH, Liu F, Wei W, Liu LB, Xu MH, Guo
ZY, Li W, Jiang B and Wu YL: Oridonin induces apoptosis and
senescence by increasing hydrogen peroxide and glutathione
depletion in colorectal cancer cells. Int J Mol Med. 29:649–655.
2012.PubMed/NCBI
|
|
29
|
Markowitz SD and Bertagnolli MM: Molecular
origins of cancer: Molecular basis of colorectal cancer. N Engl J
Med. 361:2449–2460. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Saini S, Duraisamy AJ, Bayen S, Vats P and
Singh SB: Role of BMP7 in appetite regulation, adipogenesis, and
energy expenditure. Endocrine. 48:405–409. 2015. View Article : Google Scholar
|
|
31
|
Kuzaka B, Janiak M, Włodarski KH,
Radziszewski P and Włodarski PK: Expression of bone morphogenetic
protein-2 and -7 in urinary bladder cancer predicts time to tumor
recurrence. Arch Med Sci. 11:378–384. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li W, Cai HX, Ge XM, Li K, Xu WD and Shi
WH: Prognostic significance of BMP7 as an oncogene in
hepatocellular carcinoma. Tumour Biol. 34:669–674. 2013. View Article : Google Scholar
|
|
33
|
Aoki M, Ishigami S, Uenosono Y, Arigami T,
Uchikado Y, Kita Y, Kurahara H, Matsumoto M, Ueno S and Natsugoe S:
Expression of BMP-7 in human gastric cancer and its clinical
significance. Br J Cancer. 104:714–718. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu Y, Chen J, Yang Y, Zhang L and Jiang
WG: Molecular impact of bone morphogenetic protein 7, on lung
cancer cells and its clinical significance. Int J Mol Med.
29:1016–1024. 2012.PubMed/NCBI
|
|
35
|
Alarmo EL, Pärssinen J, Ketolainen JM,
Savinainen K, Karhu R and Kallioniemi A: BMP7 influences
proliferation, migration, and invasion of breast cancer cells.
Cancer Lett. 275:35–43. 2009. View Article : Google Scholar
|
|
36
|
Ying X, Sun Y and He P: Bone morphogenetic
protein-7 inhibits EMT-associated genes in breast cancer. Cell
Physiol Biochem. 37:1271–1278. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lim M, Chuong CM and Roy-Burman P: PI3K,
Erk signaling in BMP7-induced epithelial-mesenchymal transition
(EMT) of PC-3 prostate cancer cells in 2- and 3-dimensional
cultures. Horm Cancer. 2:298–309. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Motoyama K, Tanaka F, Kosaka Y, Mimori K,
Uetake H, Inoue H, Sugihara K and Mori M: Clinical significance of
BMP7 in human colorectal cancer. Ann Surg Oncol. 15:1530–1537.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Camara-Clayette V, Koscielny S, Roux S,
Lamy T, Bosq J, Bernard M, Fest T, Lazar V, Lenoir G and Ribrag V:
BMP7 expression correlates with secondary drug resistance in mantle
cell lymphoma. PLoS One. 8:e739932013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fiaschetti G, Castelletti D, Zoller S,
Schramm A, Schroeder C, Nagaishi M, Stearns D, Mittelbronn M,
Eggert A, Westermann F, et al: Bone morphogenetic protein-7 is a
MYC target with prosur-vival functions in childhood
medulloblastoma. Oncogene. 30:2823–2835. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Itoh F, Asao H, Sugamura K, Heldin CH, ten
Dijke P and Itoh S: Promoting bone morphogenetic protein signaling
through negative regulation of inhibitory Smads. EMBO J.
20:4132–4142. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Greenblatt MB, Kim JM, Oh H, Park KH, Choo
MK, Sano Y, Tye CE, Skobe Z, Davis RJ, Park JM, et al: p38α MAPK is
required for tooth morphogenesis and enamel secretion. J Biol Chem.
290:284–295. 2015. View Article : Google Scholar :
|