Introduction
Angiogenesis is indispensable in cancer progression,
and therefore a promising target for the development of cancer
treatments. Endothelial cells (ECs) that line the tumor vasculature
are ideal target cells for cancer therapeutics, since they are
easily accessible to agents delivered by blood and are genetically
stable without being easily mutated into drug-resistant variants
(1,2). Anginex, an antiparallel
β-sheet-forming 33-mer peptide, is structured and synthesized based
on the basic folding principles and incorporation of short
sequences from the β-sheet domains of natural anti-angiogenic
agents including platelet factor-4 (PF4), interleukin-8 (IL-8) and
bactericidal-permeability increasing protein 1 (BP1) (3–6).
Anginex has been shown to prevent the proliferation, adhesion and
migration of activated ECs, and induce apoptosis in these cells.
Furthermore, it has been demonstrated that anginex significantly
reduces tumor growth and angiogenesis in mouse models (7–9). These
advances suggest that anginex is a very promising agent for the
treatment of various solid tumors regardless of the heterogeneity
of cancer cells.
As a small artificial synthetic peptide, anginex has
the characteristics of hypotoxicity, hyperosmolality and high
selectivity; but it can easily be hydrolyzed by endopeptidase with
poor stability and short half-life. Recombinant anginex has been
proven to have comparable angiostatic properties as the synthetic
peptide (10). We have previously
constructed the AdNT4-anginex virus, which secrete anginex and has
shown apparent anti-angiogenesis and antitumor effects in
vitro and in vivo (11,12).
However, adenovirus transduction is transient with a relatively
strong host immune response (13).
In contrast to adenovirus, adeno-associated virus (AAV) can deliver
genes to cells to provide a stable expression with no immune
response, and preferentially integrate into a specific site at the
q arm of chromosome 19 (14–16).
Therefore, AAV may be a more suitable vector to deliver anginex for
the treatment of cancers. Previous studies have shown that anginex
inhibits migration and proliferation, and induces apoptosis in ECs;
however, its precise underlying molecular mechanism remains to be
determined.
In order to achieve a stable expression of anginex
for cancer therapy, we constructed the recombinant adeno-associated
virus (rAAV2)-anginex; and assessed the effects of rAAV2-anginex on
the proliferation of human umbilical vein endothelial cells
(HUVECs) in vitro, as well as its anti-angiogenesis and
antitumor effects in mouse tumor xenografts. Furthermore, we
explored the mechanism of anti-angiogenesis and antitumor effects
of rAAV2-anginex.
Materials and methods
Materials
The empty adeno-associated virus (AAV2) was
purchased from Bo Miao Biotechnology (Beijing, China). The
rAAV2-anginex virus was designed with NT4 secretion signal sequence
at the N-terminal side and the 6xHis-tag sequence at the C-terminal
side of anginex. Titers of rAAV2-anginex and AAV2 virus were
4.4×1011 and 5.0×1011 vg/ml, respectively.
Four-week-old female athymic BALB/c nude mice were purchased from
the Animal Center of the Medical College of Xi'an Jiaotong
University. White leghorn eggs were purchased from Xi'an Chicken
Breeding Station (Xi'an, China). Fetal bovine serum (FBS),
Dulbecco's modified Eagle's medium (DMEM) and Rosewell Park
Memorial Institute (RPMI)-1640 medium were purchased from HyClone
(Logan, UT, USA). Vascular endothelial growth factor 165
(VEGF165; 100-20) was purchased from PeproTech (Rocky
Hill, NJ, USA). NF-κB pathway sampler kit (#9936), rabbit
anti-His-tag monoclonal antibody (#12698), rabbit
anti-extracellular signal regulated kinase monoclonal antibody
(ERK, #4695), rabbit anti-phospho-ERK monoclonal antibody (p-ERK,
#4370), rabbit anti-phospho-Akt monoclonal antibody (p-Akt, #4060),
rabbit anti-Akt monoclonal antibody (#4685), rabbit
anti-phospho-c-Jun N-terminal kinase monoclonal antibody (p-JNK,
#4668), horseradish peroxidase (HRP)-conjugated anti-mouse (#7046)
and anti-rabbit (#7044) IgG antibodies were purchased from Cell
Signaling Technology (Danvers, MA, USA). Rabbit anti-CD31
polyclonal antibody (ab28364), rabbit anti-Bcl-2 monoclonal
antibody (ab136285), rabbit anti-Bax polyclonal antibody (ab7977),
rabbit anti-Fas poly-clonal antibody (ab82419) and rabbit
anti-cyclin D1 polyclonal antibody (ab7958) were purchased from
Abcam (Cambridge, UK). GAPDH monoclonal antibody (HRP-60004),
rabbit anti-VEGF165 polyclonal antibody (19003-1-AP) and
rabbit anti-JNK polyclonal antibody (51151-1-AP) were purchased
from the Proteintech Group (Wuhan, China).
Cell culture
HUVECs and human ovarian cancer cell line SKOV3 were
kindly provided by the Translational Medical Center of the Medical
College of Xi'an Jiaotong University. HUVECs and SKOV3 cells were
cultured at 37°C with 5% CO2 in DMEM and RPMI-1640
medium, respectively, containing 10% FBS, 100 U/ml of penicillin
and 100 µg/ml of streptomycin (Life Technologies, Grand
Island, NY, USA). HUVECs were transduced for 72 h with
rAAV2-anginex or AAV2 virus at a multiplicity of infection (MOI) of
106 for further experiments. Cells were passaged every
2–3 days in 100-mm dishes and harvested at the end of the treatment
period for further analysis.
MTT assay
HUVECs were plated into 96-well plates. The next
day, cells were separately treated with rAAV2-anginex and AAV2
virus, and medium as blank control. Each group with 7 repeats was
incubated for 7 days. Cell viability was evaluated daily with
[3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide
(MTT)] (5 mg/ml) assay (Promega, Shanghai, China). Briefly, 20
µl of MTT was added and incubated in the dark for an
additional 4 h at 37°C. Then, the supernatant was discarded and 100
µl of dimethyl sulfoxide (DMSO; Sigma-Aldrich, St. Louis,
MO, USA) was added into each well to dissolve the formazan product.
The absorbance of the enzyme was measured at 490 nm excitation
emission wavelength using a microplate reader (Bio-Rad, Hercules,
CA, USA).
EdU incorporation assay
Cell proliferation was assessed with
5-ethynyl-2′-deoxyuridine (EdU) incorporation assay (C10310-1;
RiboBio, Guangzhou, China), according to the manufacturer's
instructions. Briefly, HUVECs transduced with rAAV2-anginex and
empty AAV2 virus, and untreated HUVECs were plated in triplicate in
96-well plates at a density of 3×103 cells/well, and
cultured to logarithmic phase. Then, 25 µM of EdU was added
into each well and cells were cultured for an additional 1.5 h at
37°C. Cells were fixed with 4% formaldehyde for 30 min at room
temperature and treated with 0.5% Triton X-100 for 10 min at room
temperature for permeabilization. After washing with
phosphate-buffered saline (PBS) 3 times, 100 µl of 1X
Apollo® reaction cocktail was added into each well; and
cells were incubated in the dark for 30 min at room temperature.
Then, cells were stained with 100 µl of Hoechst 33342 in the
dark for 30 min and visualized under a fluorescent microscope
(Nikon-Eclipse; Nikon, Tokyo, Japan). EdU-positive (red) and
Hoechst 33342-positive cells (blue) were counted using Image Pro
Plus (IPP) 6.0 software (Media Cybernetics, Bethesda, MD, USA). EdU
incorporation rate was expressed as the ratio of EdU-positive cells
to total Hoechst 33342-positive cells.
Flow cytometric analysis for cell cycle
and apoptosis
For cell cycle analysis, cells were trypsinized into
single cell suspensions and fixed with 70% ethanol for 2 h at 4°C.
After washing with PBS, cells were treated with RNase A (50
µg/ml), and stained with 25 µg/ml of propidium iodide
(PI) for 30 min at 37°C in the dark. Samples were analyzed using a
flow cytometer (BD FACSCalibur; BD Biosciences, Franklin Lakes, NJ,
USA) and data were analyzed with the ModiFit LT v2.0 software. For
analysis of apoptosis, cells were harvested by trypsinization and
washed with PBS, followed by double staining with PE Annexin V and
7-AAD, according to the manufacturer's instructions (559763; BD
Biosciences).
Transwell invasion and migration
assays
Each Transwell chamber with polycarbonate membrane
filters with 24-well inserts and an 8-µm pore size (Corning,
Corning, NY, USA) was coated with 50 µl of Matrigel (1:6
dilution; BD, San Diego, CA, USA), and the gel was polymerized at
37°C for 8 h. Then, the prepared cells (4×104) were
plated onto the upper chambers with serum-free medium (200
µl/well), while the bottom chambers were filled with DMEM
containing 10% FBS (600 µl/well). After incubation for 48 h
at 37°C in 5% CO2, non-invading cells in the upper
chamber were removed; and cells that invaded to the lower surface
of the filter were fixed with 4% paraformaldehyde for 15 min and
stained with crystal violet for 30 min. The number of invasive
cells were visualized under a microscope (Nikon-Eclipse) and
quantified by counting the number of cells in five randomly chosen
fields with 200-fold magnification. Migration assay was performed
without adding Matrigel, and experimental steps were the same as
the invasion assay; but the number of plated cells was
2×104.
Tube formation assay
Each well of the pre-chilled 96-well plate was
bottom-coated with 50 µl of Matrigel (BD) and incubated at
37°C for 30 min to polymerize. HUVECs trans-duced with the
rAAV2-anginex and empty AAV2 virus, and untreated HUVECs were
seeded into each well and incubated at 37°C with 5% CO2
for 6 h. Then, images were captured (magnification, ×40) to
evaluate tube formation using IPP 6.0 software (Media
Cybernetics).
Chick chorioallantoic membrane (CAM)
assay
CAM assay was performed as previously described
(11). Briefly, fresh fertilized
white leghorn eggs were incubated at 37°C with 60% relative
humidity, and flipped every 4 h with an automatic hatching machine
(Science Incubator, Shandong, China). On day 3, a 1×2 cm
rectangular window was made to continually observe the angiogenesis
of the chick embryo of the CAM, and this window was covered with
tape to prevent dehydration during the non-observation time. On day
7, a piece of gelatin sponge was placed on the CAM for the
convenience of intervention. On day 13, 1 µg of
VEGF165 + 100 µl of PBS, 1 µg of
VEGF165 + 5.0×1010 vg of rAAV2-anginex and 1
µg of VEGF165 + 5.0×1010 vg of AAV2
were added, respectively, to the CAM. On day 18, CAM angiogenesis
was monitored and photographed.
RT-PCR
Total RNAs were isolated using a Fast 200 RNA
extraction kit (Feijie Biological Technology, Shanghai, China),
while cDNA was synthesized with a reverse transcription kit
(#RR036A; Takara, Dalian, China), according to the manufacturer's
instructions. PCR was performed using a PCR amplifying system
(Bio-Rad). Primers were designed and synthesized by Sangon Biotech
(Shanghai, China). Primer sequences of rAAV2-anginex were as
follows: forward, 5′-GTA CGGTGGGAGGTCTATATAAGCA-3′ and reverse,
5′-GGT CCCGGTGTCTTCTATGGA-3′; for an internal standard, primer
sequences of GAPDH were as follows: forward, 5′-AGG
TCCACCACTGACACGTT-3′ and reverse, 5′-GCCTCAAG ATCAGCAAT-3′. The
reaction consisted of a Premix Taq (12.5 µl), forward
and reverse primers (0.4 µM, 1 µl), cDNA (1
µl) and ddH2O (10.5 µl). Reaction
conditions were as follows: 94°C for 3 min, 25 cycles of 94°C for
30 sec, 59.6°C for 30 sec and 72°C for 30 sec. PCR products were
electrophoretically separated on 2% agarose gel, and bands were
visualized by ethidium bromide staining. Band intensity was
evaluated using an agarose gel imaging system (Bio-Rad).
Western blotting
HUVECs were lysed at 4°C for 30 min in cell lysis
buffer containing protease and phosphatase inhibitors (Roche,
Indianapolis, IN, USA). Lysates were centrifuged at 16,000 x g for
20 min at 4°C. Protein concentration of the supernatant was
measured using a Pierce BCA protein assay kit (23227; Thermo Fisher
Scientific, Waltham, MA, USA), according to the manufacturer's
instructions. A total of 80 µg of each cell lysate was
separated by 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) polyacrylamide gels and transferred onto
a polyvinylidene difluoride (PVDF) membrane (Millipore Corp.,
Billerica, MA, USA). After blocking with 5% skim milk in
Tris-buffered saline (TBS) containing 0.1% Tween-20 at room
temperature for 2 h, the membranes were incubated with primary
antibodies (diluted at 1:1,000) overnight at 4°C. GAPDH (diluted at
1:5,000) was used as a loading control. After washing 4 times in
TBS/Tween-20, the membranes were incubated for 1 h at room
temperature with goat anti-rabbit or horse anti-mouse
HRP-conjugated antibody at 1:2,000 dilution in TBS/Tween-20
containing 5% skim milk. After extensive washing in TBS/Tween-20,
blots were visualized with an enhanced chemiluminescence kit (ECL;
Millipore Corp.).
Xenograft model
A total of 1×107 SKOV3 cells in 100
µl of PBS were subcutaneously injected into the left flank
of female nude mice. When tumors reached an average size of ~200
mm3, tumor-bearing mice were randomly divided into 3
groups (n=6), and respectively treated with rAAV2-anginex
(1.8×1011 vg) and rAAV2 (1.8×1011 vg), with
PBS (100 µl) as blank control. The two maximum vertical
diameters (a, long diameter; b, short diameter) of the tumor were
measured and recorded every other day. Mice were sacrificed after
30 days. Then, the final size of the tumor was measured and
photographed. Tumor volumes were calculated by 0.5xab2.
All experimental procedures were carried out according to protocols
approved by the Ethics Committee for Animal Experimentation of the
Medical College of Xi'an Jiaotong University, in accordance with
the National Institutes of Health Guide for the Care and Use of
Laboratory Animals.
Immunohistochemistry (IHC)
Transplanted tumor tissues were formalin fixed,
paraffin embedded and cut into 4-µm serial sections.
Expression of VEGF165 and microvessel density (MVD;
CD31) was measured using IHC by a streptavidin-biotin peroxidase
(SP) kit (SP-9001; Beijing Zhongshan Golden Bridge Biotechnology,
Beijing, China), according to the manufacturer's instructions.
Briefly, antigens were retrieved using a microwave at high power
for 5 min, followed by 13 min at middle-low power in 10 mM of
citrate buffer (pH 6.0). Then, the sections were treated with 3%
hydrogen peroxide in methanol for 10 min at room temperature to
quench endogenous peroxidase activity, followed by incubation with
reagent A for 15 min at room temperature. Subsequently, the
sections were incubated with anti-CD31 polyclonal antibody
(dilution, 1:50) and anti-VEGF165 polyclonal antibody
(dilution, 1:200) in a humidified chamber at 4°C overnight. After
washing in PBS, the sections were, respectively, incubated with
reagent B and C for 15 min at 37°C. Then, diaminobenzidine
(ZLI-9018; Beijing Zhongshan Golden Bridge Biotechnology) was added
to the sections, according to the manufacturer's instructions.
Finally, the sections were rinsed with water and counterstained
with Harris hematoxylin. For negative control, PBS was used instead
of the primary antibody. The sections were observed and separately
scored by two independent investigators.
The MVD (CD31)-positivity judgment was based on the
standard Weidner et al correction methods (17). Briefly, cytoplasmic staining for
brown is positive staining; and both single cell and
primmorph-positive staining, whether or not a lumen formation
exists, are considered capillaries. Three concentrated areas as
'hot spots' at low magnification (×10) were randomly selected, and
the number of capillaries in every hot area was counted at high
magnification (×200). Results are expressed as mean ± standard
error (mean ± SEM).
The expression intensity of VEGF165 was
quantified as the sum of the integrated optical densities (IODs) of
threshold pixels for all signals measured in each image. Then, the
average value for each group was calculated. All imaging analysis
was performed using IPP 6.0 software (Media Cybernetics). Results
are expressed as mean ± SEM.
Statistical analysis
Statistical analysis was performed using SPSS
software (Statistical Package for the Social Sciences version 21.0;
SPPS, Inc., Chicago, IL, USA). Differences between groups were
analyzed by one-way ANOVA and LSD analysis. All statistical tests
were two-sided. P<0.05 was considered to indicate a
statistically significant result.
Results
rAAV2-anginex inhibits HUVEC
proliferation
Anginex targets activate ECs and potent antitumor
and angiogenesis effects have been shown in mouse models (7–9). In
order to achieve a stable expression of anginex for cancer therapy,
a recombinant anginex adeno-associated virus, rAAV2-anginex, was
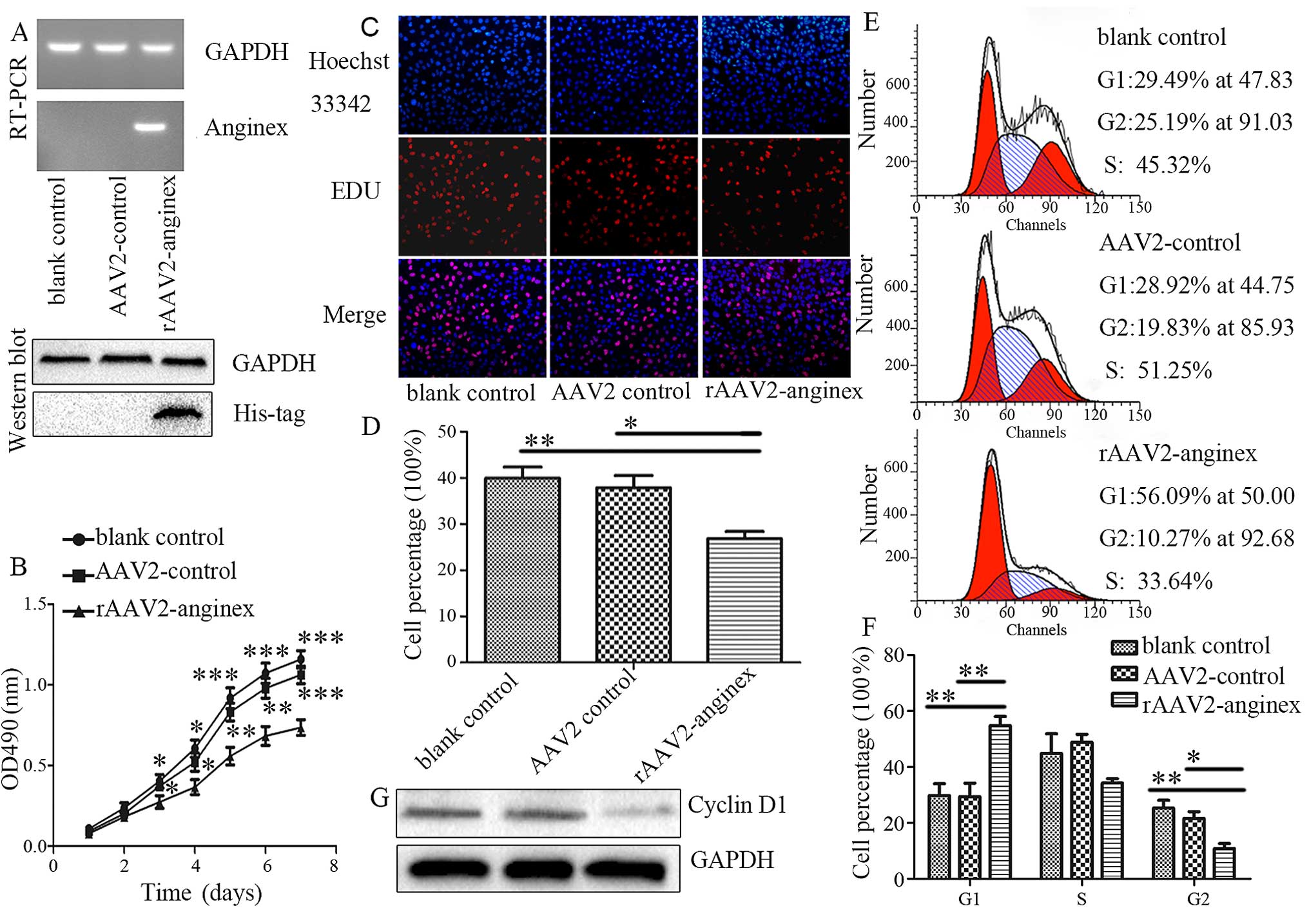
constructed. The expression of rAAV2-anginex was confirmed by
RT-PCR (24 h) and western blotting (72 h), respectively, after
transduction of the rAA2-anginex and AAV2 virus in HUVECs. As shown
in Fig. 1A, both the mRNA and
protein of anginex were detected in rAAV2-anginex virus-transduced
cells. Next, the effect of anginex on HUVEC proliferation was
assessed by MTT assay. HUVECs treated with the rAAV2-anginex virus
grew slower from the third day (P=0.048), compared with the blank
control group and empty virus AAV2. The inhibition effect was more
obvious with increased treatment time, with a growth inhibition
rate of 30.77% (P<0.001; Fig.
1B). EDU incorporation assay further verified that
rAAV2-anginex significantly inhibited HUVEC proliferation (P=0.012;
Fig. 1C and D). After incubation
with EDU for 90 min, EDU incorporation in the rAAV2-anginex, blank
control and AAV2 control group was 26.79±1.61% (mean ± SEM),
39.62±2.13% (P<0.01) and 37.85±2.67% (P<0.05), respectively.
These results clearly show that rAAV2-anginex significantly
inhibited HUVEC proliferation.
PF4, which is also an anti-angiogenic agent that
contain β-sheet domains, has been shown to inhibit the cell cycle
progression of ECs by blocking the transition from G1 to S phase
and the progression of S phase (18). In order to test whether
rAAV2-anginex alters cell cycle progression, flow cytometric
analysis was carried out on HUVEC following the transduction of
rAAV2-anginex. Compared with controls, rAAV2-anginex transduced
cells displayed an increased percentage of cells in the G1 phase
(P<0.01) and fewer cells in the G2 phase (P<0.05), but there
was no significant difference in S phase distribution (P>0.05);
implying that anginex may block the G1-S transition (Fig. 1E and F). Furthermore, western blot
analysis revealed that rAAV2-anginex reduced cyclin D1 protein
levels, compared with AAV2 (Fig.
1G). Taken together, these results demonstrate that
rAAV2-anginex inhibited HUVEC proliferation by reducing the G1/S
phase transition of HUVECs.
rAAV2-anginex induces HUVEC
apoptosis
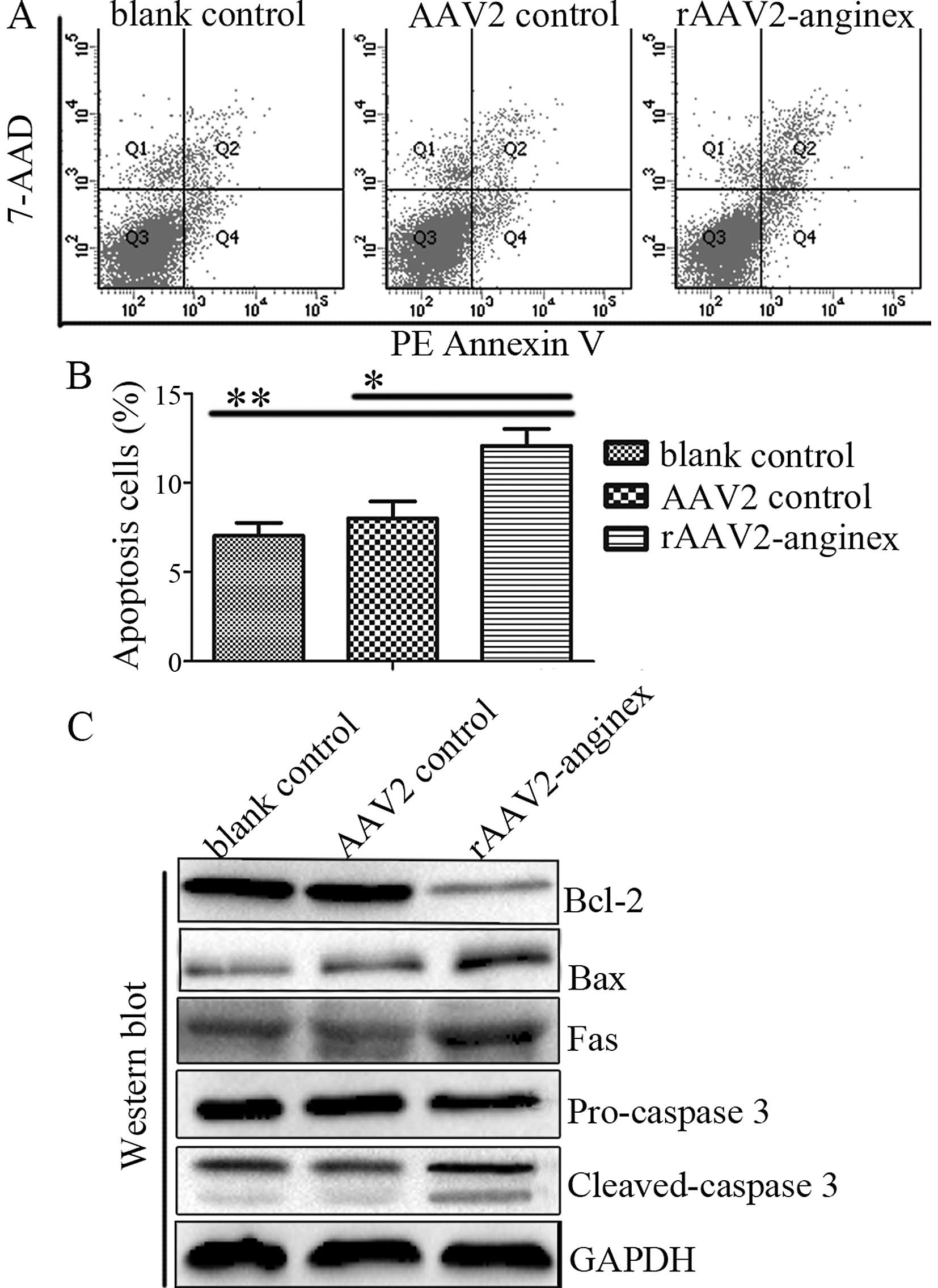
In order to explore whether rAAV2-anginex induces
apoptosis, HUVECs were stained with Annexin V, followed by flow
cytometry. The transduction of rAAV2-anginex resulted in an
elevated number of apoptotic cells, compared with the control
groups (P<0.05; Fig. 2A and B).
Further immunoblotting revealed that rAAV2-anginex decreased Bcl-2
protein levels and increased protein expression levels of Bax, Fas
and cleaved-caspase-3 (Fig. 2C).
These data indicate that rAAV2-anginex induced apoptosis in
HUVECs.
rAAV2-anginex inhibits HUVEC migration
and invasion
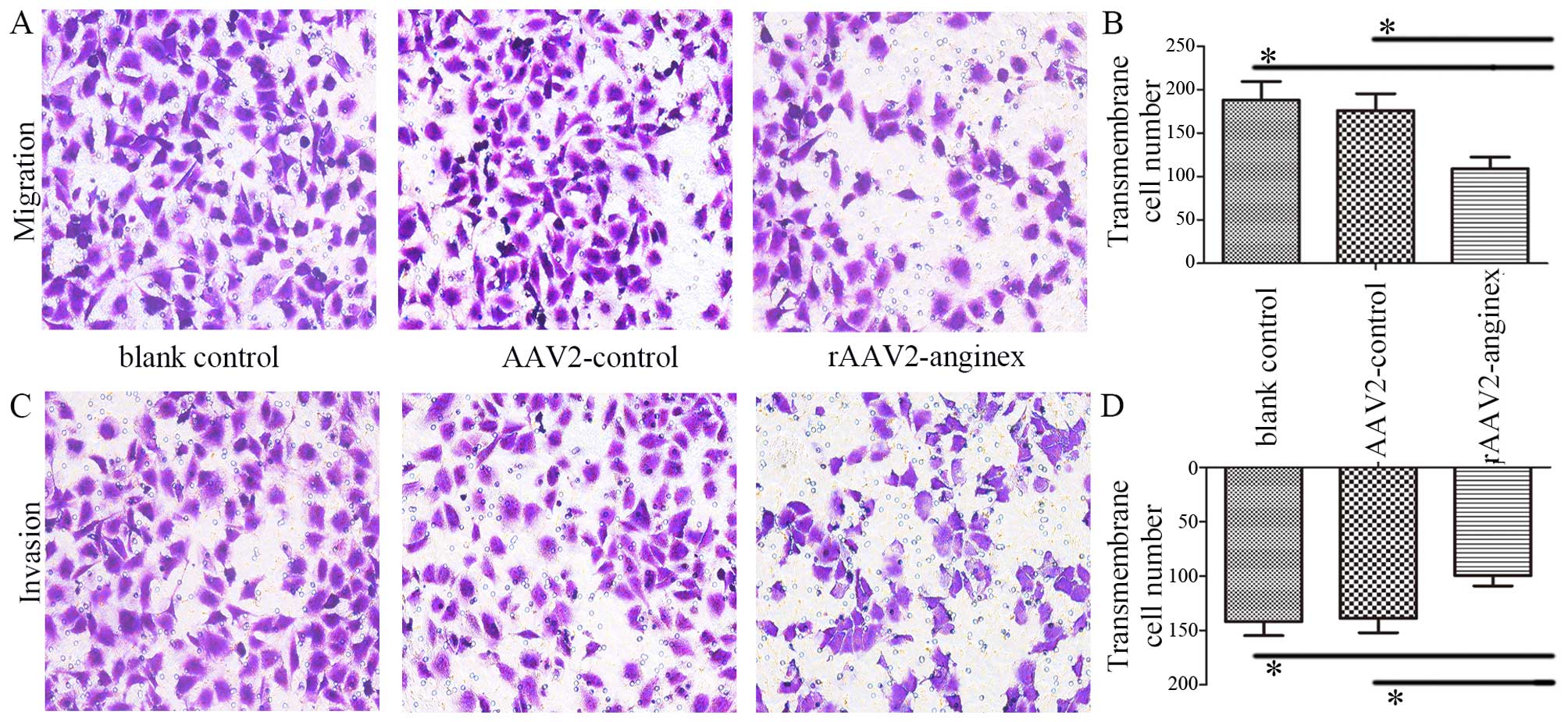
Previous studies have shown that anginex inhibited
the adhesion and migration of ECs (19). Accordingly, the migration and
invasion ability of HUVECs transduced with rAAV2-anginex were
examined by Transwell migration and Matrigel invasion assays.
Transwell migration assay revealed that rAAV2-anginex significantly
decreased the migration of HUVECs, compared to the empty AAV2 and
blank groups (P<0.05; Fig. 3A and
B). In addition, Matrigel invasion assay demonstrated that
rAAV2-anginex significantly reduced HUVEC invasion, compared to the
empty AAV2 and blank group (P<0.05; Fig. 3C and D). Thus, rAAV2-anginex had a
remarkable inhibitory effect on HUVEC migration and invasion.
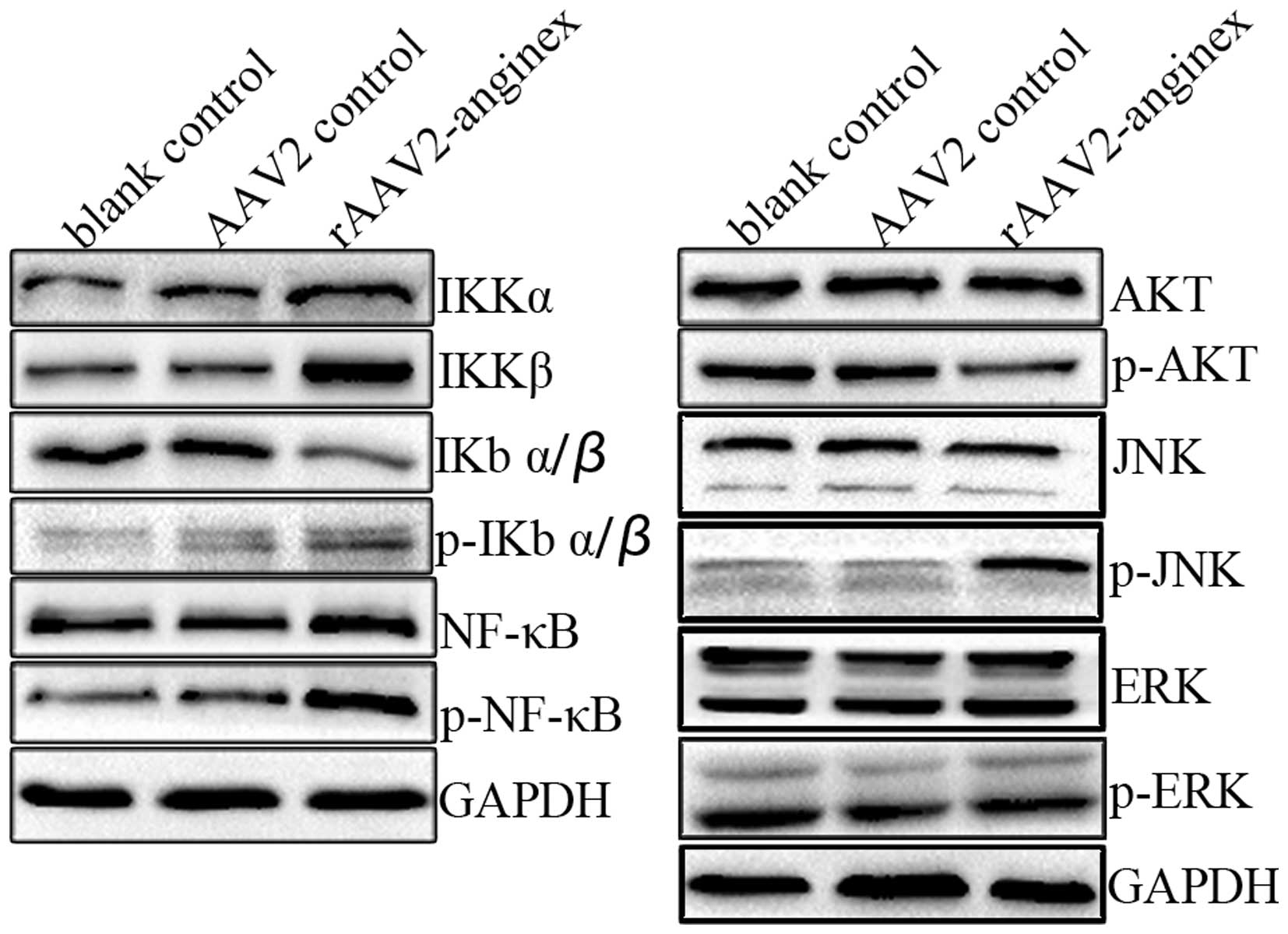
Modulation of Akt, JNK and NF-κB
signaling by rAAV2-anginex in HUVECs
Akt, MAPK and NF-κB signaling pathways play an
important role in the regulation of cell proliferation, survival,
migration and invasion. In order to explore the potential mechanism
for inhibiting cell proliferation, migration and invasion during
rAAV2-anginex-induced HUVEC apoptosis, the activity of Akt, MAPK
and the NF-κB signaling pathway was assessed by immunoblotting. As
shown in Fig. 4, compared with the
control group, p-Akt in the rAAV2-anginex treated group markedly
decreased. For the MAPK signaling pathway, p-JNK increased in the
rAAV2-anginex treated group, while p-ERK had no obvious change
compared to the control group. In the NF-κB signal pathway, for the
expression of p-NF-κB, p-IKBα/β and IKKα, IKKβ increased and total
IKBα/β decreased in the rAAV2-anginex group, suggesting that the
NF-κB signaling pathway was activated by rAAV2-anginex. Thus,
rAAV2-anginex treatment resulted in the inhibition of Akt and
activation of the JNK and NF-κB signaling pathways, but not ERK
signaling in HUVECs.
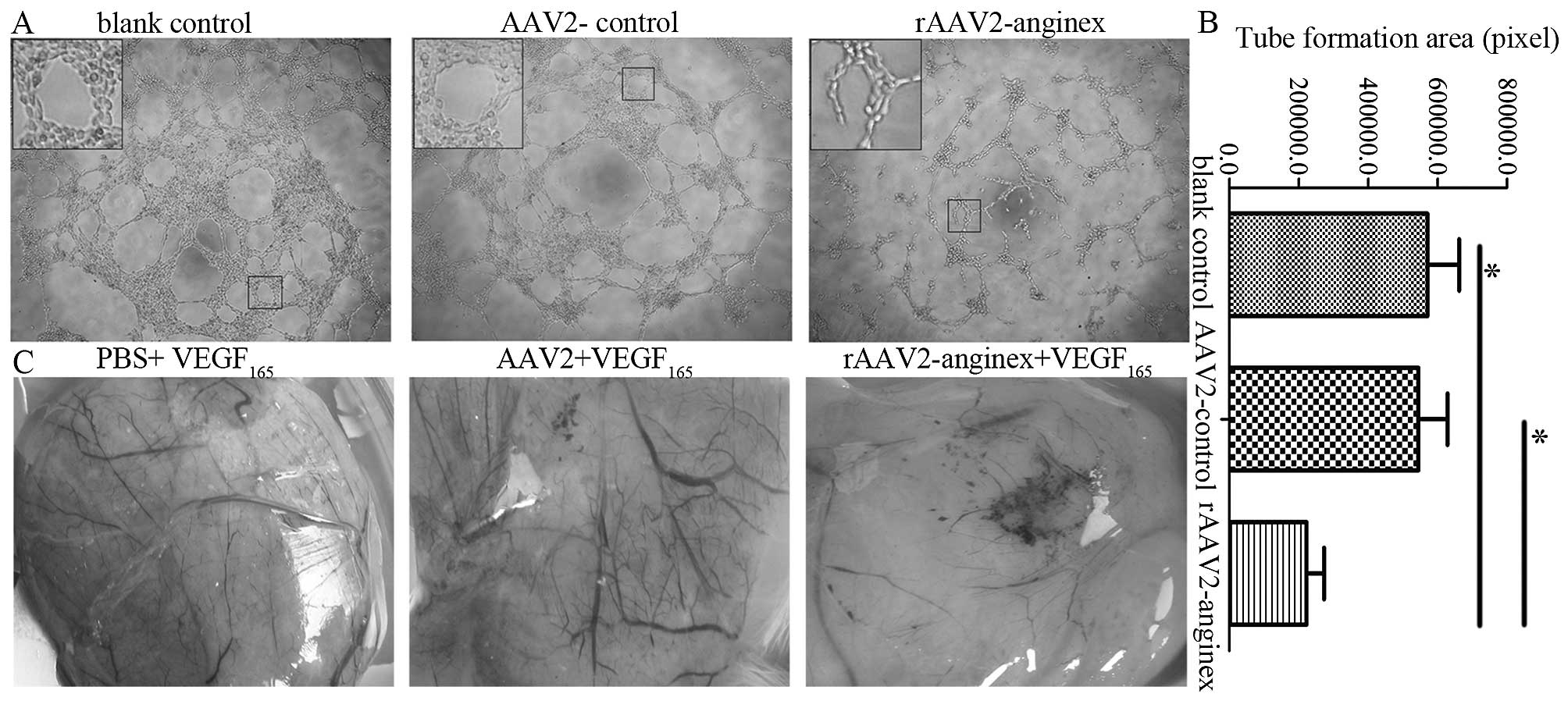
rAAV2-anginex inhibits angiogenesis in
HUVECs and chick embryo
In order to assess the anti-angiogenesis effect of
rAAV2-anginex, tube formation assay in vitro and CAM assay
in vivo, were performed. Compared with blank control and
AAV2 control, rAAV2-anginex-treated HUVECs formed incomplete
tube-like structures; and the extent of the tube formations of
HUVECs significantly decreased (P<0.05; Fig. 5A and B). In the in vivo CAM
assay, rAAV2-anginex treatment significantly resulted in a
decreased number of blood vessels and even the disappearance of
blood vessels in some areas. Compared with vessels in the PBS group
and AAV2 group, the rAAV2-anginex-treated group revealed that the
caliber, distribution and length of vessels were reduced. Moreover,
in the rAAV2-anginex treated group, coarse vessels became
discontinued and obliterated, and the vascular net resembling leaf
vein vanished and was replaced by confused arrangement and
disfiguration (Fig. 5C). Therefore,
rAAV2-anginex significantly inhibited angiogenesis both in HUVECs
in vitro and chick embryo in vivo.
rAAV2-anginex reduces angiogenesis and
inhibits tumor growth in ovarian cancer SKOV3 cell nude mice
xenograft model
In order to further explore the anti-angiogenesis
and antitumor effects of rAAV2-anginex, an ovarian cancer SKOV3
cell nude mice xenograft model was applied to determine the
anti-angiogenesis and antitumor effects of rAAV2-anginex. When the
subcutaneous tumor volume reached ~200 mm3, mice were
treated with PBS, AAV2 and rAAV2-anginex. On day 8 after treatment,
transplanted tumor volume in the rAAV2-anginex group was
significantly lower than in the control groups (P<0.05). On the
last day of observation (day 30), inhibition rates of tumor growth
in the rAAV2-anginex group were 43.20% (P<0.01) and 46.56%
(P<0.01) relative to the PBS and AAV2 groups, respectively
(Fig. 6A). However, there were no
significant differences in food and water intake, exercise scope
and intensity, and response sensitivity in mice among the groups,
suggesting the safety of the adeno-associated virus.
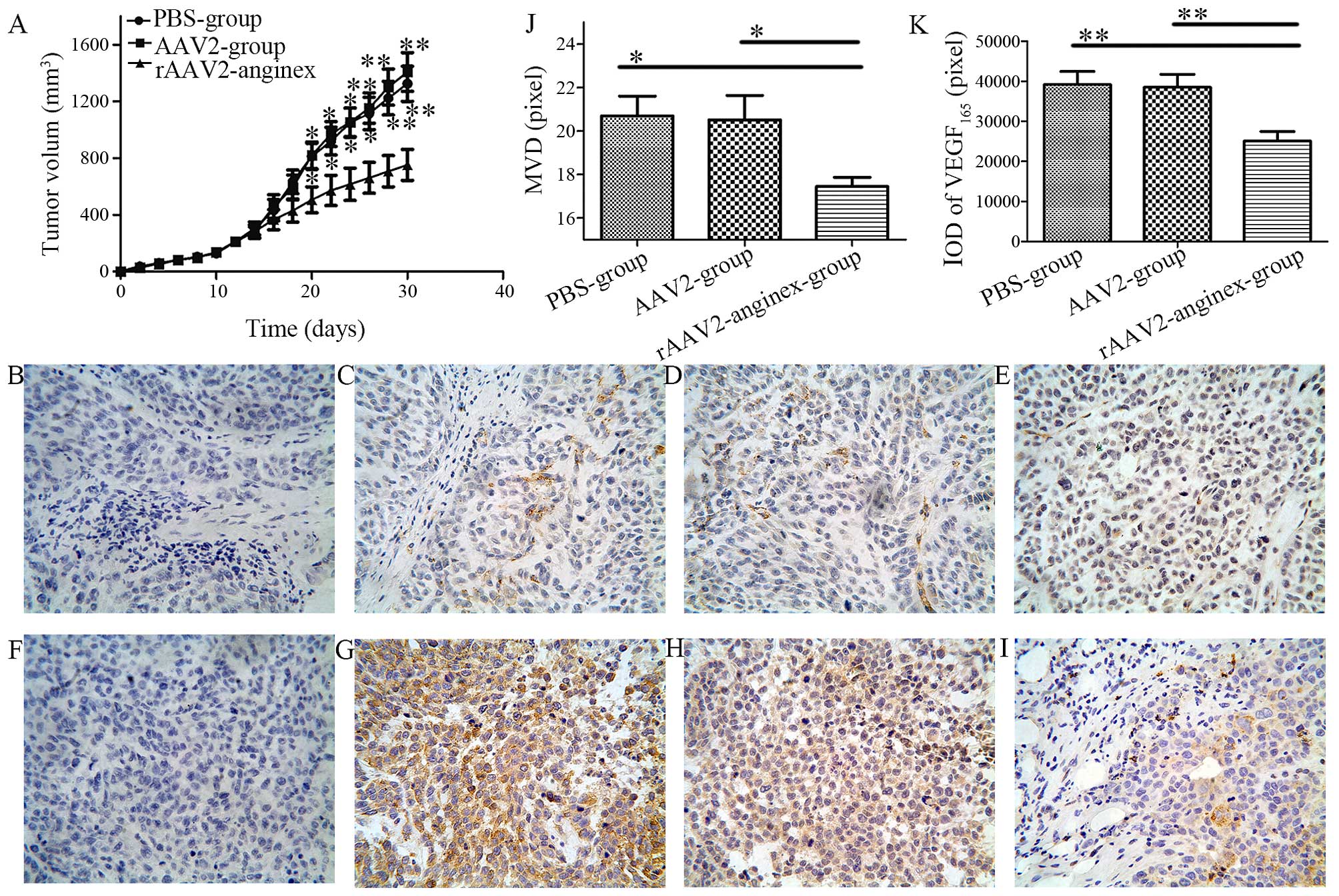 | Figure 6Anti-angiogenesis and antitumor
effects of rAAV2-anginex in vivo. (A) The transplantation of
tumor volumes was monitored regularly in SKOV3 xenograft models.
Data are expressed as mean ± SD; *P<0.05 and
**P<0.01, respectively, compared with the
rAAV2-anginex group. (B–G) Representative immu-nohistochemical
staining of MVD (CD31) (upper, B–E) and VEGF165 (lower,
F–I) in nude mouse xenograft tissues of human ovarian cancer cell
line SKOV3. (B) Immunostaining for PBS (magnification, ×400). (C)
Immunostaining for CD31 in the PBS-treated group (magnification,
×400). (D) Immunostaining for CD31 in the AAV2-treated group
(magnification, ×400). (E) Immunostaining for CD31 in the
rAAV2-anginex treated group (magnification, 400). (F)
Immunostaining for PBS (magnification, 400). (G) Immunostaining for
VEGF165 in the PBS-treated group (magnification, 400).
(H) Immunostaining for VEGF165 in the AAV2-treated group
(magnification, 400). (I) Immunostaining for VEGF165 in
the rAAV2-anginex-treated group (magnification, 400). (J) Qualified
of MVD (CD31). (K) Qualified IOD of VEGF165. |
In order to test the anti-angiogenesis effect of
rAAV2-anginex in mice, VEGF165 and MVD were assessed by
IHC. CD31 is marker of vascular endothelial cells, and the IHC of
CD31 was used for MVD (Fig. 6B–E).
According to the Weidner et al (17) correction method, MVD values were
significantly different among the 3 groups (P<0.05, Fig. 6J). Mean MVD value in the
rAAV2-anginex group was 17.45±0.42 (mean ± SEM), which
significantly decreased compared with the AAV2 group (20.52±1.12;
P=0.024) and PBS group (20.70±0.91; P=0.018); and there was no
statistically significant difference in MVD values between the AAV2
and PBS group (P=0.885). These results revealed that rAAV2-anginex
significantly inhibited the microvascular density of ovarian
transplantation tumor tissues.
Immunostaining of VEGF165 was observed on
the plasma membrane and in the cytoplasm (Fig. 6F–I). Mean IOD of paraffin sections
in the rAAV2-anginex group was 25,118.33±2,351.43 (mean ± SEM),
which was significantly lower than the PBS group
(39,208.00±3,256.43; P<0.01) and AAV2 group (38,521.17±3,260.06;
P<0.01; Fig. 6I). There was no
statistically significant difference in the IOD of paraffin
sections between the AAV2 and PBS group (P=0.873). Thus, the
rAAV2-anginex virus significantly inhibited the protein expression
level of VEGF165 in ovarian transplantation tumors.
Discussion
In the present study, we demonstrated that the
rAAV2-anginex virus significantly inhibited the proliferation,
migration and invasion of HUVECs, accompanied with cell cycle
arrest at the G1/S phase and induction of apoptosis. Moreover,
rAAV2-anginex treatment led to the inhibition of Akt and activation
of the JNK and NF-κB signaling pathways in HUVECs. Furthermore,
rAAV2-anginex significantly inhibited angiogenesis both in HUVECs
in vitro and chick embryo in vivo. Importantly,
rAAV2-anginex inhibited angiogenesis and tumor growth in the
ovarian cancer SKOV3 cell nude mouse xenograft model.
Anginex is a small artificial cytokine-like
β-sheet-forming peptide with anti-angiogenesis and antitumor
effects. However, its poor stability and short half-life limits its
further development for cancer therapy. In the present study, we
applied the recombinant adeno-associated virus (rAAV) as a delivery
method for anginex (rAAV2-anginex). We found that rAAV2-anginex
significantly inhibited G1/S phase transition, accompanied by the
downregulation of cyclin D1. Cyclin D1 is a downstream target of
PI3K-Akt signaling, which is a predominant cell growth-promoting
signaling pathway (20). Notably,
rAAV2-anginex significantly reduced the phosphorylation of Akt.
Furthermore, Akt signaling indirectly and directly regulates
pro-apoptosis and anti-apoptosis proteins, and ultimately blocks
the apoptosis process (21–23). In the present study, we found that
rAAV2-anginex significantly induced apoptosis, which was
accompanied by the increase in pro-apoptosis proteins Bax and Fas
and decrease of anti-apoptosis protein Bcl-2. These results
indicate that the inhibition of Akt signaling is one of the
mechanisms by which rAAV2-anginex inhibited proliferation and
induced apoptosis in HUVECs. Akt signaling had been reported to
activate NF-κB signaling (24). The
activation of NF-κB signaling in endothelial cells is critical for
the activity of angiostatic agents including anginex (25). However, in the present study,
rAAV2-anginex inhibited Akt signaling and activated the NF-κB
signaling pathway, suggesting that rAAV2-anginex may induce
apoptosis and inhibit angiogenesis through different signaling
pathways. Surprisingly, we found that rAAV2-anginex induced
phosphorylation of JNK. Since NF-κB and JNK signaling are two key
regulators of the pathophysiology of cells and a wide range of
cross-talk occurs between these two signaling pathways (26–28),
it is possible that the activation of the JNK/NF-κB signaling
pathway may be involved in the antitumor and anti-angiogenesis
effects of rAAV2-anginex. Additionally, galectin-1 is a cell
surface receptor that promotes H-Ras signaling to the
Raf/mitogen-activated protein kinase/extracellular signal regulated
kinase (Erk) kinase (Mek)/Erk cascade. It has been reported that
synthetic anginex could bind galectin-1 in activated ECs and
disrupt the adhesion and migration of tumor ECs, thereby resulting
in the inhibition of tumor angiogenesis (29). However, in the present study,
rAAV2-anginex did not significantly change the total expression of
Erk and phospho-Erk, suggesting that rAAV2-anginex may inhibit
angiogenesis independent of Erk signaling.
Considering the potent effects of rAAV2-anginex on
HUVECs in vitro, we further explored the anti-angiogenesis
and antitumor growth effects of rAAV2-anginex in vivo; and
found that rAAV2-anginex significantly inhibited angiogenesis in
chick embryo. Importantly, the xenograft model further confirmed
the safety and efficiency of rAAV2-anginex for further clinical
trials. Compared with the PBS and AAV2 group, inhibition rates of
tumor growth in the rAAV2-anginex group were 43.20 and 46.56% on
the day 30; which is less than that reported in the (73%) B16F10
xenograft (7) and (80%) MA148
xenograft models (9) of synthetic
anginex. These different inhibitory effects may be due to the
different xenograft models chosen, and the observation times. In
addition, the synthetic anginex was continuously injected for 21 or
28 days; while in the present study, the rAAV2-anginex virus was
injected only once, which may have influenced tumor growth to some
extent. In the study of Brandwijk et al (30), recombinant and synthetic anginex
injected every 3 days inhibited tumor growth by ~80 and 55%,
respectively; which demonstrated the remarkable antitumor effect of
recombinant anginex. Surprisingly, the decreased expression of
VEGF165 protein in the rAAV2-anginex virus-treated group
may provide a new understanding of the anti-angiogenesis of
rAAV2-anginex.
In summary, the present study demonstrates that
rAAV2-anginex inhibited the proliferation, migration and invasion
of HUVECs, as well as promoted apoptosis of HUVECs, ultimately
leading to the inhibition of angiogenesis and tumor growth.
Additionally, the xenograft model revealed that the
adeno-associated virus is safe and has a long-lasting effect,
providing a new treatment option for cancer therapy. More
importantly, the present study indicates that the regulation of the
Akt, JNK and NF-κB signaling pathways are potential mechanisms of
the effects of rAAV2-anginex on HUVECs.
Acknowledgments
The present study was supported by grants from the
National Natural Science Foundation of China (no. 81172169).
References
|
1
|
Griffioen AW and Molema G: Angiogenesis:
Potentials for pharmacologic intervention in the treatment of
cancer, cardiovascular diseases, and chronic inflammation.
Pharmacol Rev. 52:237–268. 2000.PubMed/NCBI
|
|
2
|
Folkman J: Angiogenesis in cancer,
vascular, rheumatoid and other disease. Nat Med. 1:27–31. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mayo KH, Haseman J, Ilyina E and Gray B:
Designed beta-sheet-forming peptide 33mers with potent human
bactericidal/permeability increasing protein-like bactericidal and
endotoxin neutralizing activities. Biochim Biophys Acta.
1425:81–92. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Griffioen AW, van der Schaft DW,
Barendsz-Janson AF, Cox A, Struijker Boudier HA, Hillen HF and Mayo
KH: Anginex, a designed peptide that inhibits angiogenesis. Biochem
J. 354:233–242. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ilyina E, Roongta V and Mayo KH: NMR
structure of a de novo designed, peptide 33mer with two distinct,
compact beta-sheet folds. Biochemistry. 36:5245–5250. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dings RP, Arroyo MM, Lockwood NA, van Eijk
LI, Haseman JR, Griffioen AW and Mayo KH: Beta-sheet is the
bioactive conformation of the anti-angiogenic anginex peptide.
Biochem J. 373:281–288. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
van der Schaft DW, Dings RP, de Lussanet
QG, van Eijk LI, Nap AW, Beets-Tan RG, Bouma-Ter Steege JC,
Wagstaff J, Mayo KH and Griffioen AW: The designer anti-angiogenic
peptide anginex targets tumor endothelial cells and inhibits tumor
growth in animal models. FASEB J. 16:1991–1993. 2002.PubMed/NCBI
|
|
8
|
Mayo KH, van der Schaft DW and Griffioen
AW: Designed beta-sheet peptides that inhibit proliferation and
induce apoptosis in endothelial cells. Angiogenesis. 4:45–51. 2001.
View Article : Google Scholar
|
|
9
|
Dings RP, van der Schaft DW, Hargittai B,
Haseman J, Griffioen AW and Mayo KH: Anti-tumor activity of the
novel angiogenesis inhibitor anginex. Cancer Lett. 194:55–66. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Brandwijk RJ, Nesmelova I, Dings RP, Mayo
KH, Thijssen VL and Griffioen AW: Cloning an artificial gene
encoding angiostatic anginex: From designed peptide to functional
recombinant protein. Biochem Biophys Res Commun. 333:1261–1268.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dong DF, Li EX, Wang JB, Wu YY, Shi F, Guo
JJ, Wu Y, Liu JP, Liu SX and Yang GX: Anti-angiogenesis and
anti-tumor effects of AdNT4-anginex. Cancer Lett. 285:218–224.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li EX, Liu SX, Yang GX, Wang QY, Wu YY and
Shi F: Construction and identification of the recombinant
prokaryotic expression plasmid containingbetapep-25 peptide. Xi Bao
Yu Fen Zi Mian Yi Xue Za Zhi. 22:154–160. 2006.In Chinese.
|
|
13
|
Berns KI and Giraud C: Adenovirus and
adeno-associated virus as vectors for gene therapy. Ann NY Acad
Sci. 772:95–104. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Samulski RJ, Zhu X, Xiao X, Brook JD,
Housman DE, Epstein N and Hunter LA: Targeted integration of
adeno-associated virus (AAV) into human chromosome 19. EMBO J.
10:3941–3950. 1991.PubMed/NCBI
|
|
15
|
Kotin RM, Menninger JC, Ward DC and Berns
KI: Mapping and direct visualization of a region-specific viral DNA
integration site on chromosome 19q13-qter. Genomics. 10:831–834.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kotin RM, Linden RM and Berns KI:
Characterization of a preferred site on human chromosome 19q for
integration of adeno-associated virus DNA by non-homologous
recombination. EMBO J. 11:5071–5078. 1992.PubMed/NCBI
|
|
17
|
Weidner N, Semple JP, Welch WR and Folkman
J: Tumor angiogenesis and metastasis - correlation in invasive
breast carcinoma. N Engl J Med. 324:1–8. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gupta SK and Singh JP: Inhibition of
endothelial cell proliferation by platelet factor-4 involves a
unique action on S phase progression. J Cell Biol. 127:1121–1127.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang JB, Wang MD, Li EX and Dong DF:
Advances and prospects of anginex as a promising anti-angiogenesis
and anti-tumor agent. Peptides. 38:457–462. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Muise-Helmericks RC, Grimes HL, Bellacosa
A, Malstrom SE, Tsichlis PN and Rosen N: Cyclin D expression is
controlled post-transcriptionally via a phosphatidylinositol
3-kinase/Akt-dependent pathway. J Biol Chem. 273:29864–29872. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xin M and Deng X: Nicotine inactivation of
the proapoptotic function of Bax through phosphorylation. J Biol
Chem. 280:10781–10789. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Henshall DC, Araki T, Schindler CK, Lan
JQ, Tiekoter KL, Taki W and Simon RP: Activation of
Bcl-2-associated death protein and counter-response of Akt within
cell populations during seizure-induced neuronal death. J Neurosci.
22:8458–8465. 2002.PubMed/NCBI
|
|
23
|
Datta SR, Brunet A and Greenberg ME:
Cellular survival: A play in three Akts. Genes Dev. 13:2905–2927.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pianetti S, Arsura M, Romieu-Mourez R,
Coffey RJ and Sonenshein GE: Her-2/neu overexpression induces
NF-kappaB via a PI3-kinase/Akt pathway involving calpain-mediated
degradation of IkappaB-alpha that can be inhibited by the tumor
suppressor PTEN. Oncogene. 20:1287–1299. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tabruyn SP, Mémet S, Avé P, Verhaeghe C,
Mayo KH, Struman I, Martial JA and Griffioen AW: NF-kappaB
activation in endothelial cells is critical for the activity of
angiostatic agents. Mol Cancer Ther. 8:2645–2654. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Karin M: Nuclear factor-kappaB in cancer
development and progression. Nature. 441:431–436. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen F, Castranova V and Shi X: New
insights into the role of nuclear factor-kappaB in cell growth
regulation. Am J Pathol. 159:387–397. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu J and Lin A: Wiring the cell signaling
circuitry by the NF-kappa B and JNK1 crosstalk and its applications
in human diseases. Oncogene. 26:3267–3278. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Thijssen VL, Postel R, Brandwijk RJ, Dings
RP, Nesmelova I, Satijn S, Verhofstad N, Nakabeppu Y, Baum LG,
Bakkers J, et al: Galectin-1 is essential in tumor angiogenesis and
is a target for antiangiogenesis therapy. Proc Natl Acad Sci USA.
103:15975–15980. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Brandwijk RJ, Dings RP, van der Linden E,
Mayo KH, Thijssen VL and Griffioen AW: Anti-angiogenesis and
anti-tumor activity of recombinant anginex. Biochem Biophys Res
Commun. 349:1073–1078. 2006. View Article : Google Scholar : PubMed/NCBI
|