Introduction
Gastric cancer (GC) is one of the most common human
cancers. Better knowledge of the changes in gene expression that
occur during gastric carcinogenesis may lead to improvements in
diagnosis, treatment and prevention of GC (1,2). We
previously performed serial analysis of gene expression (SAGE) on
four primary GC tissues and identified several genes whose
expression was either upregulated or downregulated in GC (3). Of these genes, REG4 and
OLFM4 were found to encode secreted proteins and serve as
highly sensitive serum markers for GC. Although detailed functions
of REG4 and OLFM4 are unclear, it has been reported
that REG4 is upregulated in aldehyde dehydrogenase 1
(ALDH1)-positive gastric cancer stem cells (CSCs) (4). OLFM4 is a highly specific and
robust marker for Lgr5-positive stem cells of small intestine
(5), suggesting that both
REG4 and OLFM4 play important roles in stem cell
function. In the past decade, cancer has been recognized as a stem
cell disease (6). CSCs have been
described in numerous solid tumors and have been characterized by
specific cell surface marker expression including CD44, CD133 and
ALDH1 (7). However, gastric CSCs
have not been completely characterized. To characterize CSCs,
spheroid colony formation is a useful methods (8). To form spheroid colony, cells are
cultured in culture dishes specially coated for non-attachment with
serum-free media. The spheroid colonies have characteristics of CSC
phenotype (8). However, gene
expression profile of spheroid colonies from the GC cells remains
unclear.
In the present study, we analyzed gene expression
profile of spheroid colonies from the GC cell lines by microarray
analysis, and found that expression of KIFC1 was upregulated
in gastric spheroid colonies. KIFC1 (also known as HSET) is a
C-type kinesin of the kinesin-14 family (9), and is assumed to be a minus
end-directed motor protein (10).
Kinesins are a family of molecular motors and play important roles
in intracellular transport or cell division (11). Alteration of several types of
kinesins have been reported in human cancers (11). Among them, upregulation of
KIFC1 has been shown in human glioblastoma, lung, breast and
colon cancer (12). In breast
cancer cells, forced expression of KIFC1 inhibits
docetaxel-mediated apoptosis (13).
However, expression of KIFC1 in GC has not been
investigated. Therefore, we analyzed the expression and
distribution of KIFC1 in human GC by immunohistochemistry, and
examined the relationship between KIFC1 staining and
clinicopathologic characteristics. We also analyzed the effect of
inhibiting KIFC1 expression by RNA interference (RNAi) on the
spheroid formation in GC cells.
Materials and methods
Cell lines
Two cell lines derived from human GC (MKN-45 and
MKN-74) were used. All cell lines were purchased from the Japanese
Collection of Research Bioresources Cell Bank (Osaka, Japan). All
cell lines were maintained in RPMI-1640 medium (Nissui
Pharmaceutical Co., Ltd., Tokyo,. Japan) containing 10% fetal
bovine serum (BioWhittaker, Walkersville, MD, USA) in a humidified
atmosphere of 5% CO2 and 95% air at 37°C.
Spheroid colony formation
For the generation of spheres, 2,000 cells were
plated on a 24-well ultra-low attachment plate (Corning, Corning,
NY, USA). Cells were grown in mTeSR medium (StemCell Technologies
Inc.). The plates were incubated at 37°C in a 5% CO2
incubator for 15 days. Sphere number and size were determined and
counted under a microscope.
Microarray analysis
Total RNA from MKN-45 spheroid body-forming and
parental cells and MKN-74 spheroid body-forming and parental cells
was isolated by lysing cells in Isogen lysis buffer (Nippon Gene,
Tokyo, Japan) followed by isopropanol precipitation. We performed
microarray analyses using the Human Genome U133 Plus 2.0 Array
(Affymetrix, Santa Clara, CA, USA) as previously described
(14). The arrays were scanned with
a GeneChip Scanner 3000 (Affymetrix), and the data were analyzed by
Microarray Suite version 5.0 with Affymetrix default analysis
settings and global scaling as normalization method. The trimmed
mean target intensity of each array was arbitrarily set to
1,000.
Quantitative reverse
transcription-polymerase chain reaction (qRT-PCR) analysis
Total RNA was extracted with an RNeasy Mini kit
(qiagen, Valencia, CA, USA), and 1 µg of total RNA was
converted to cDNA using the First Strand cDNA Synthesis kit
(Amersham Biosciences, Piscataway, NJ, USA). quantitation of
KIF2C, KIF4A, KIF11, KIF15,
KIF20A, KIF20B, KIF22, KIF23 and
KIFC1 mRNA levels was performed by real-time fluorescence
detection as previously described (15). PCR was conducted using the
SYBR-Green PCR Core Reagents kit (Applied Biosystems). Real-time
detection of the emission intensity of SYBR-Green bound to
double-stranded DNA was performed with the ABI PRISM 7700 Sequence
Detection System (Applied Biosystems). ACTB-specific PCR products
were amplified from the same RNA samples and served as an internal
control.
RNAi and cell growth assay
Short interfering RNA (siRNA) oligonucleotides
targeting KIFC1 and a negative control were purchased from
Invitrogen (Carlsbad, CA, USA). We used three independent BST2
siRNA oligonucleotide sequences. Transfection was performed using
Lipofectamine RNAiMAX (Invitrogen) as previously described
(16). Briefly, 60 pmol of siRNA
and 10 µl of Lipofectamine RNAiMAX were mixed in 1 ml of
RPMI-1640 medium (10 nmol/l final siRNA concentration). After 20
min of incubation, the mixture was added to cells, and then cells
were plated in culture dishes. Forty-eight hours after transfection
cells were analyzed.
To examine cell growth,
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay was performed. The cells were seeded at a density of 2,000
cells/well into 96-well plates. Cell growth was monitored after 1,
2 and 4 days. Three independent experiments were performed. Mean ±
SE was calculated for each of the experiments.
Western blot analysis
Cells were lysed as previously described (17). The lysates (40 µg) were
solubilized in Laemmli sample buffer by boiling and then subjected
to 10% SDS-polyacrylamide gel electrophoresis followed by
electro-transfer onto a nitrocellulose filter. Anti-KIFC1
monoclonal antibody was purchased from Abnova.
Peroxidase-conjugated anti-mouse IgG was used in the secondary
reaction. Immunocomplexes were visualized with an ECL Western Blot
Detection system (Amersham Biosciences). β-actin (Sigma, St. Louis,
MO, USA) was also stained as a loading control.
Tissue samples
In a retrospective study design, 114 primary tumors
were collected from patients diagnosed with GC who underwent
surgery between 2003 and 2007 at Hiroshima University Hospital
(Hiroshima, Japan). The present study was approved by the Ethics
Committee for Human Genome Research of Hiroshima University
(Hiroshima, Japan). All patients underwent curative resection. Only
patients without preoperative radiotherapy or chemotherapy and
without clinical evidence of distant metastasis were enrolled in
the study. Operative mortality was defined as death within 30 days
of patients leaving the hospital, and these patients were removed
from the analysis. Postoperative follow-up was scheduled every one,
two or three months during the first two years after surgery and
every six months thereafter unless more frequent follow-up was
deemed necessary. Chest X-ray, chest computed tomography scan and
serum chemistries were performed at every follow-up visit. Patients
were followed by the patients physician until their death or the
date of the last documented contact.
Immunohistochemistry
One or two representative tumor blocks, including
the tumor center, invading front and tumor-associated
non-neoplastic mucosa, was examined from each patient by
immunohistochemistry. In cases of large, late-stage tumors, two
different sections were examined to include representative areas of
the tumor center as well as of the lateral and deep tumor invasive
front. Immunohistochemical analysis was performed with a Dako
EnVision+ Mouse Peroxidase Detection System (Dako Cytomation,
Carpinteria, CA, USA). Antigen retrieval was performed by microwave
heating in citrate buffer (pH 6.0) for 30 min. Peroxidase activity
was blocked with 3% H2O2-methanol for 10 min,
and sections were incubated with normal goat serum (Dako
Cytomation) for 20 min to block non-specific antibody binding
sites. Sections were incubated with a mouse monoclonal anti-KIFC1
(1:50; Abnova) or anti-Ki-67 antibodies (Dako Cytomation) for 1 h
at room temperature, followed by incubation with EnVision+
anti-mouse peroxidase for 1 h. For color reaction, sections were
incubated with the DAB substrate-chromogen solution (Dako
Cytomation) for 10 min. Sections were counterstained with 0.1%
hematoxylin. Negative controls were created by omission of the
primary antibody.
Expression of KIFC1 was scored in all tumors as
positive or negative. When >10% of tumor cells were stained, the
immunostaining was considered positive for KIFC1. Using these
definitions, two surgical pathologists (N.O. and K.S.), without
knowledge of the clinical and pathologic parameters or the patients
outcomes, independently reviewed immunoreactivity in each specimen.
Interobserver differences were resolved by consensus review at a
double-headed microscope after independent review. For the
Ki-67-index, a total of 1,000 nuclei were counted to evaluate the
percentage of positive nuclei. The Ki-67-index was considered to
reflect the proliferative index. Immunostaining of CD44 and ALDH1
was also performed as previously described (7).
Statistical methods
Associations between clinicopathological parameters
and KIFC1 expression were analyzed by Fisher's exact test.
Kaplan-Meier survival curves were constructed for KIFC1-positive
and KIFC1-negative patients. Survival rates were compared between
KIFC1-positive and KIFC1-negative groups. Differences between
survival curves were tested for statistical significance by a
log-rank test. Differences in the sphere number and size between
the two groups were tested by the Student t-test.
Results
Gene expression profile of the spheroid
body-forming and the parental cells
To characterize spheroid colonies from the GC cells,
we performed microarray analyses by Human Genome U133 Plus 2.0
Array in the spheroid body-forming and the parental cells from
MKN-45 cells. We found 797 genes whose expression was significantly
higher in the spheroid body-forming cells than in the parental
cells, and 646 genes whose expression was significantly lower the
spheroid body-forming cells than in the parental cells. Microarray
analyses were also performed in the spheroid body-forming and the
parental cells from MKN-74 cells. We found 822 genes whose
expression was significantly higher in the spheroid body-forming
cells than in the parental cells, and 599 genes whose expression
was significantly lower than spheroid body-forming cells in the
parental cells. To identify ideal biomarkers for gastric CSC, we
focused on genes whose expression was high in the spheroid
body-forming cells from both MKN-45 and MKN-74 cells, and
identified 255 genes. Among these genes, 9 KIF genes (KIF11,
KIF15, KIF2C, KIF20A, KIF20B,
KIF22, KIF23, KIFC1 and KIF4A) were
upregulated in the spheroid body-forming cells from both MKN-45 and
MKN-74 cells. Kinesins are a family of molecular motors and play
important roles in intracellular transport or cell division
(11). Alteration of several types
of kinesins have been reported in human cancers (11), however, to the best of our
knowledge, significance of KIF genes in CSC and GC has not
yet been studied. Therefore, we decided to investigate KIF
expression in GC.
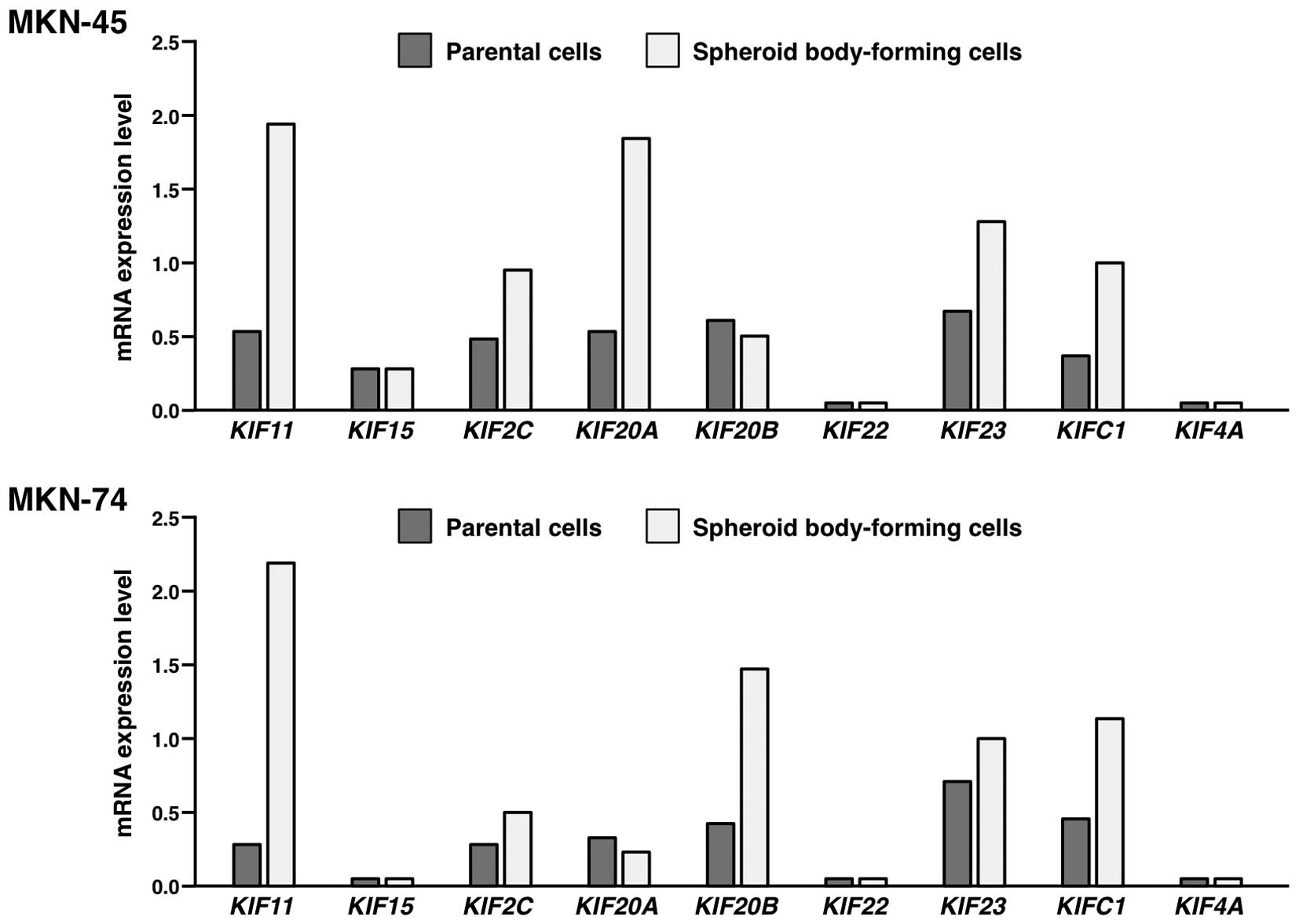
Expression of KIF genes in the spheroid
body-forming cells compared to the parental cells
To confirm upregulation of KIF genes in the
spheroid body-forming cells, expression of KIF11,
KIF15, KIF2C, KIF20A, KIF20B,
KIF22, KIF23, KIFC1 and KIF4A mRNA was
measured by qRT-PCR in MKN-45 and MKN-74 cells (Fig. 1). Among the 9 KIF genes, only
KIF11 and KIFC1 genes were expressed more than twice
higher in the spheroid body-forming cells than in the parental
cells in both MKN-45 and MKN-74 cells. In the present study, we
focused on KIFC1 since an antibody against KIFC1 protein is
commercially available. KIFC1, also known as HSET, is a minus
end-directed motor protein that promotes microtubule cross-linking,
sliding, bundling and spindle pole focusing, has been identified as
an essential mediator of supernumerary centrosome clustering in
cancer cells (18). KIFC1 is
essential for the survival of cancer cells with extra centrosomes.
In contrast, KIFC1 is not essential for mitosis in normal cells,
indicating that KIFC1 is a cancer-selective therapeutic target
(18). In contrast, significance of
KIFC1 in the spheroid body-forming cells has not been studied.
Therefore, function of KIFC1 was further analyzed in GC
cells.
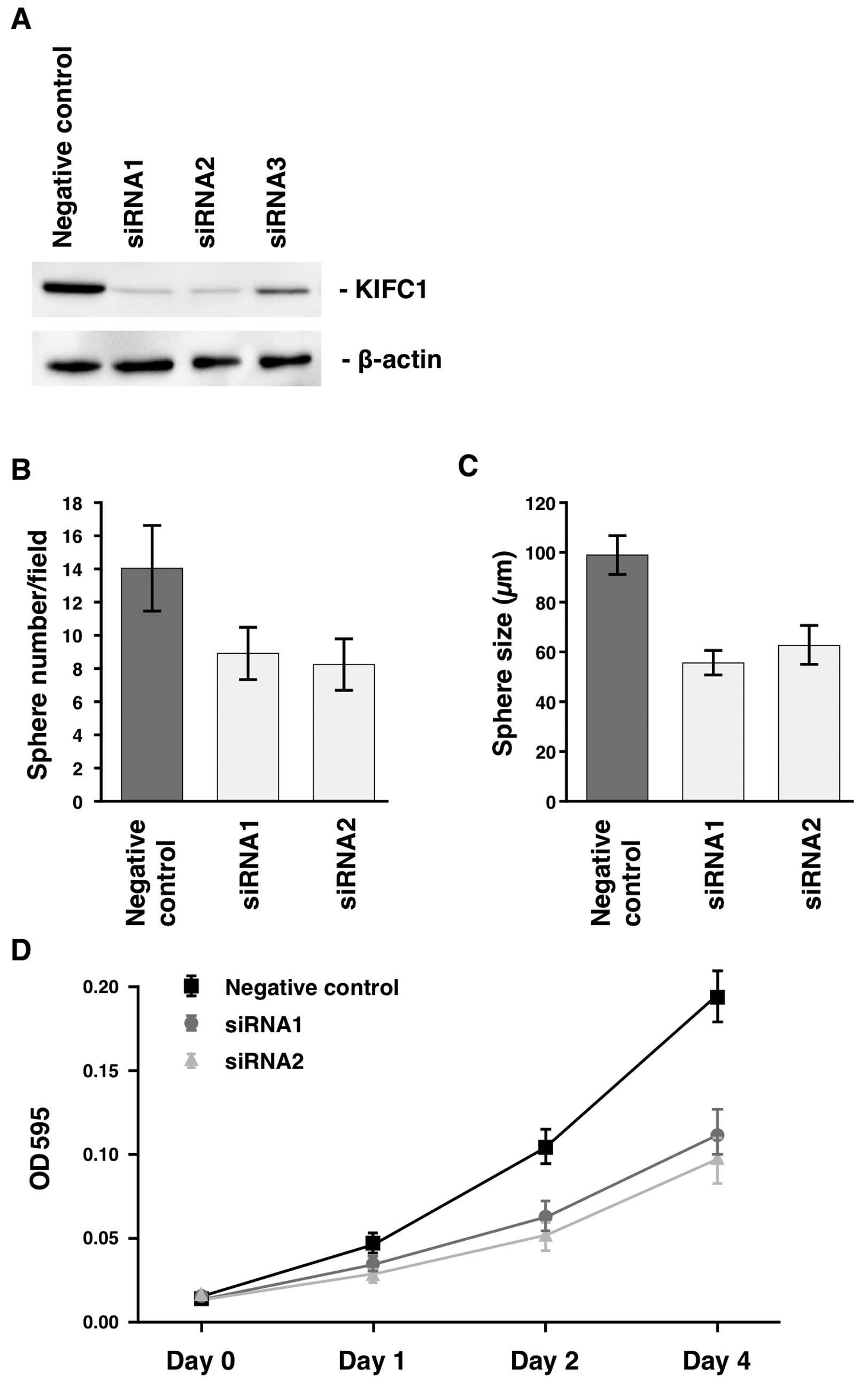
Effect of inhibition of KIFC1 on sphere
number and size
Effect of KIFC1 inhibition on sphere number
and size was investigated. First, MKN-45 GC cells, in which obvious
KIFC1 protein was detected, were selected for these experiments. We
examined transition of KIFC1 expression by western blot analysis of
cell extracts of MKN-45 transfected with KIFC1-specific
siRNAs. Three types of siRNAs (siRNA1, siRNA2 and siRNA3) were
transfected into MKN-45. The anti-KIFC1 antibody detected a band of
~74 kDa in western blot analyses of cell extracts from MKN-45 GC
cells, and the expression of KIFC1 was substantially suppressed by
treatment with siRNA1 and siRNA2 (Fig.
2A). Therefore, to knock down the endogenous KIFC1, we
used siRNA1 and siRNA2 in the following experiments. We analyzed
sphere number and size 15 days after siRNA transfection. The number
of spheres from MKN-45 cells was significantly reduced in
KIFC1 siRNA1-trasfected and KIFC1 siRNA2-transfected
MKN-45 cells than in negative control siRNA-transfected cells
(Fig. 2B). The size of sphere from
MKN-45 cells was significantly reduced in KIFC1
siRNA1-trasfected and KIFC1 siRNA2-transfected MKN-45 cells
compared to negative control siRNA-transfected cells (Fig. 2C). We also analyzed the number and
the size of spheres from MKN-74 cells, and similar results were
obtained (data not shown). These results suggest that KIFC1 is
required for sphere formation in GC cells.
Next, we performed an MTT assay 4 days after siRNA
transfection to investigate the antiproliferative effects of
KIFC1 inhibition. KIFC1 siRNA1-trasfected and
KIFC1 siRNA2-transfected MKN-45 cells showed significantly
reduced cell growth relative to negative control siRNA-transfected
MKN-45 cells (Fig. 2D). We also
performed an MTT assay 4 days after siRNA transfection in MKN-74
cells, and similar results were obtained (data not shown). These
results indicate that KIFC1 is involved in GC cell
growth.
Immunohistochemical analysis of KIFC1 in
GC
We found that KIFC1 was upregulated in
spheroid body-forming cells. In addition, the number and size of
spheres were significantly reduced in KIFC1 inhibition.
However, expression and distribution of KIFC1 in GC have not been
investigated. Therefore, immunohistochemistry was performed on 114
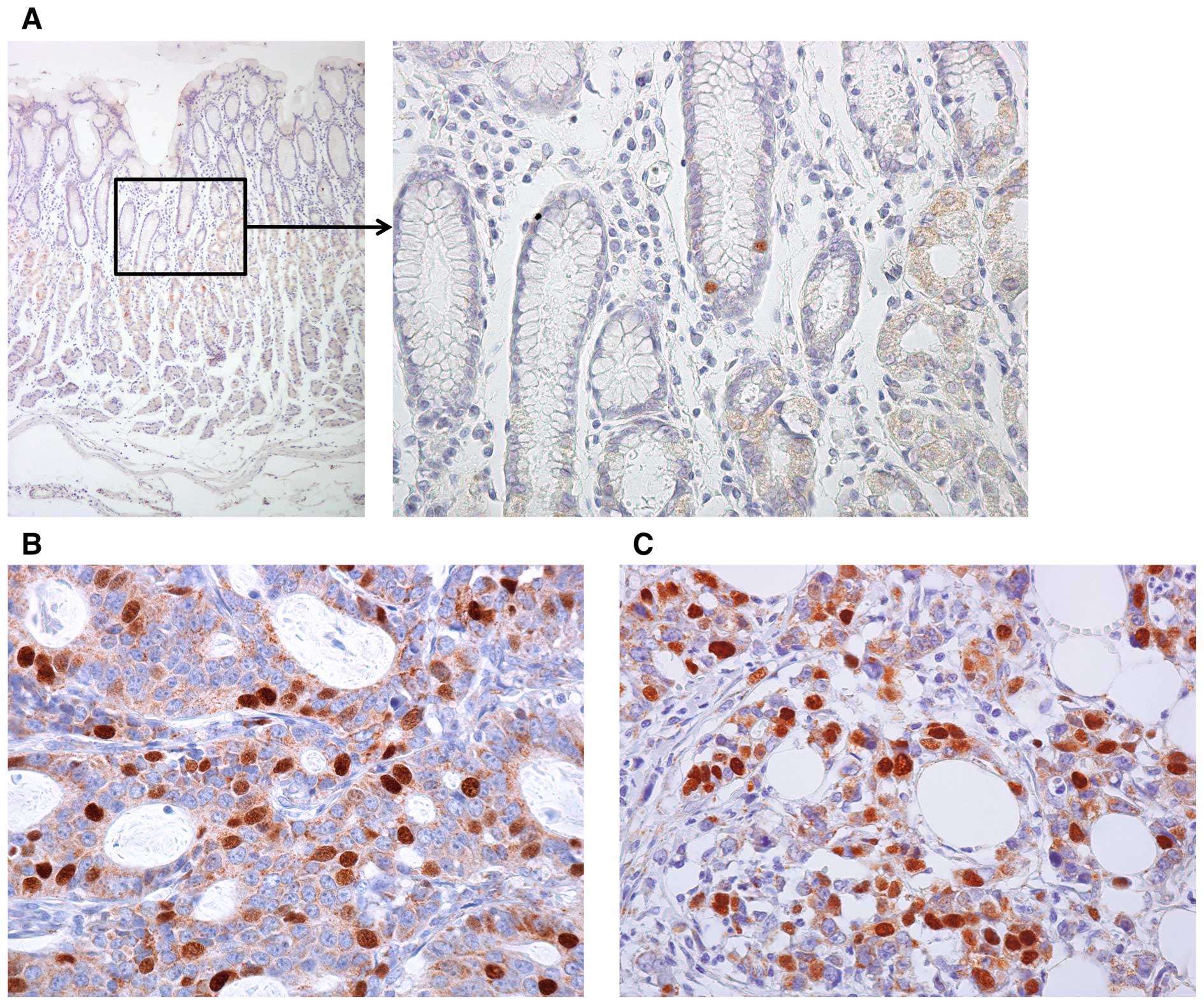
GC tissue samples. In non-neoplastic gastric mucosa, weak or no
staining of KIFC1 was observed in the foveolar epithelium or
stromal cells. In some cases, nuclear staining was observed in the
epithelial cells of gastric isthmus (Fig. 3A), which contains gastric stem cells
(19). In contrast, GC tissue
showed stronger, more extensive staining. Expression of KIFC1 was
frequently observed in intestinal type GC (Fig. 3B). GC cells of diffuse type GC were
also stained by KIFC1 (Fig. 3C).
Staining of KIFC1 was observed mainly in the nucleus. Many GC cases
showed heterogeneity of KIFC1 staining and the percentage of
KIFC1-stained GC cells ranged from 0 to 60%. A tendency for
upregulation of KIFC1 at the invasive front was not observed. When
>10% of tumor cells were stained, the immunostaining was
considered positive for KIFC1. In total, 42 (37%) of 114 GC cases
were positive for KIFC1.
We next examined the relationship of KIFC1 staining
to clinicopathological characteristics (Table I). GC cases positive for KIFC1 were
found more frequently in stage III/IV cases than in stage I/II
cases. GC cases positive for KIFC1 were found more frequently in
intestinal type GC cases than in diffuse type GC cases.
Furthermore, KIFC1-positive GC cases showed high Ki-67 labeling
index. In contrast, Kaplan-Meier analysis demonstrated that KIFC1
expression was not associated with survival (P=0.0922). Univariate
and multivariate Cox proportional hazards analysis also showed that
KIFC1 expression was not a prognostic predictor for survival in
patients with GC (data not shown).
 | Table IRelationship between KIFC1 expression
and clinico-pathological characteristics. |
Table I
Relationship between KIFC1 expression
and clinico-pathological characteristics.
| KIFC1 expression
| P-value |
|---|
| Positive (%) | Negative |
|---|
| Age (years) | | | 0.2456 |
| <66 | 17 (31) | 38 | |
| ≥66 | 25 (42) | 34 | |
| Gender | | | 0.3280 |
| Male | 28 (41) | 41 | |
| Female | 14 (31) | 31 | |
| T
classification | | | 0.0527 |
| T1 | 13 (27) | 36 | |
| T2/3/4 | 29 (45) | 36 | |
| N
classification | | | 0.0800 |
| N0 | 16 (28) | 41 | |
| N1/2/3 | 26 (46) | 31 | |
| M
classification | | | 0.6372 |
| M0 | 32 (36) | 58 | |
| M1 | 10 (42) | 14 | |
| Stage | | | 0.0346 |
| I | 14 (26) | 39 | |
| II/III/IV | 28 (46) | 33 | |
| Histological
classification | | | 0.0004 |
| Intestinal | 29 (55) | 24 | |
| Diffuse | 13 (21) | 48 | |
| Ki-67 labeling
index (%) | | | 0.0064 |
| <40 | 13 (24) | 42 | |
| ≥40 | 29 (49) | 30 | |
Association between KIFC1 expression and
CSC marker expression
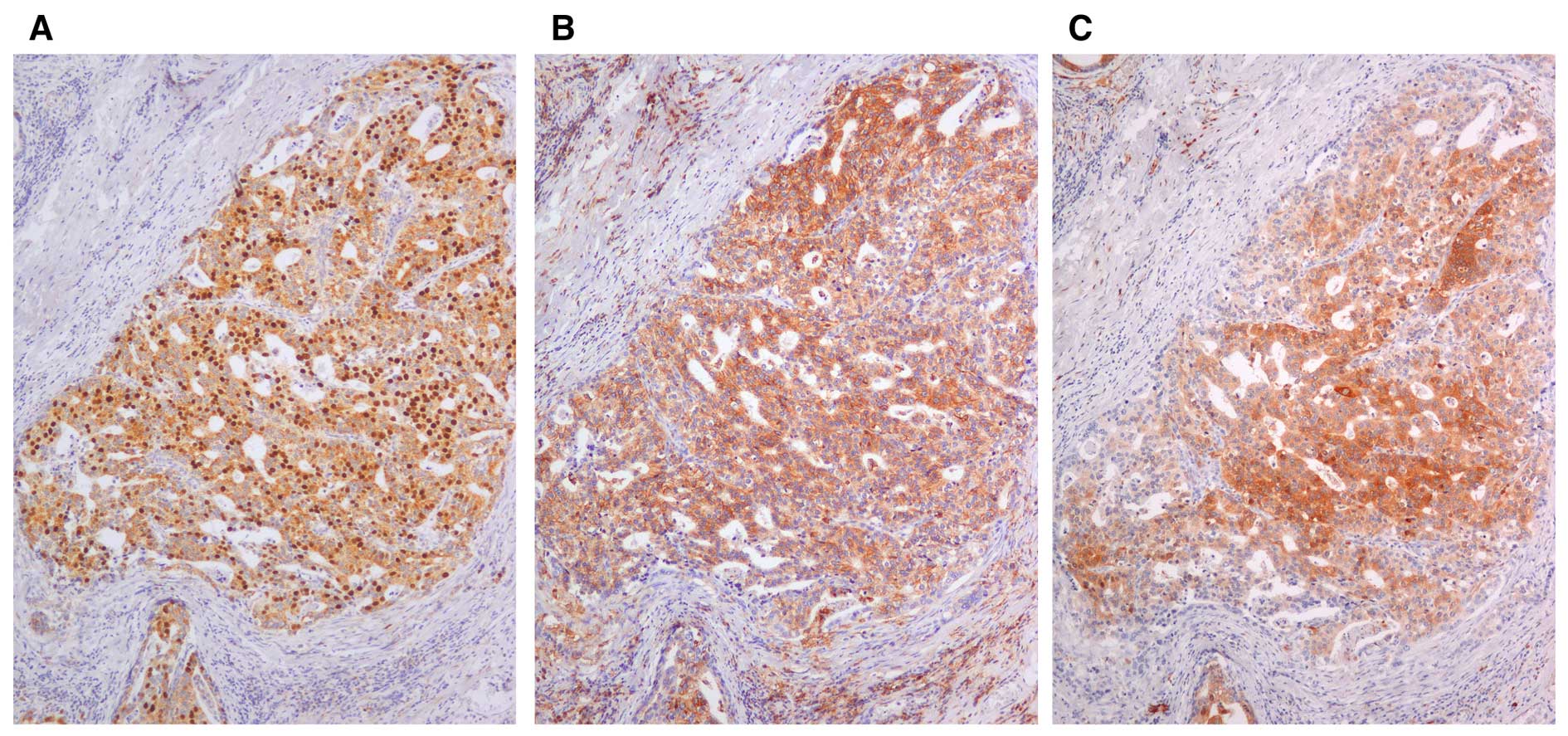
Overexpression of KIFC1 was observed in GC. However,
it remains unclear whether KIFC1 is associated with gastric CSC.
CSC markers include CD133, CD44, CD24, CD166 and ALDH1. Among them,
CD44 and ALDH1 are widely used for gastric CSC markers (7). Therefore, immunostaining of CD44 and
ALDH1 was also performed in 114 GC cases. As previously reported,
in non-neoplastic gastric mucosa, staining of CD44 was observed in
lymphocytes and stromal cells but not epithelial cells. Staining of
ALDH1 was detected in the cytoplasm of parietal cells. In GC
samples, staining of CD44 and ALDH1 was observed mainly in the
membrane and the cytoplasm of the GC cells, respectively. Of 114 GC
cases, 53 (46%) GC cases were positive for CD44 and 46 GC cases
(40%) were positive for ALDH1. As shown in Fig. 4, expression of KIFC1 was observed in
CD44-positive GC and ALDH1-positive GC cells. KIFC1-positive GC
cases were significantly frequently found in CD44-positive GC cases
(P=0.0001; Fisher's exact test; Table
II). In addition, KIFC1-positive GC cases were significantly
frequently observed in ALDH1-positive GC cases (P=0.0007; Fisher's
exact test; Table II).
| Table IIRelationship between KIFC1 expression
and CD44 or ALDH1. |
Table II
Relationship between KIFC1 expression
and CD44 or ALDH1.
| KIFC1 expression
| P-value |
|---|
| Positive (%) | Negative |
|---|
| CD44 | | | |
| Positive | 30 (57) | 23 | 0.0001 |
| Negative | 12 (20) | 49 | |
| ALDH1 | | | |
| Positive | 26 (57) | 20 | 0.0007 |
| Negative | 16 (24) | 52 | |
Discussion
To characterize CSCs, one useful method is spheroid
colony formation (8). The spheroid
colonies have characteristics of CSC phenotype (8). It has been reported that CD44 is
upregulated in spheroid formation (20) and CD44 has been identified as one of
the cell surface markers associated with CSC (21). These results indicate that genes
whose expression is upregulated in spheroid formation are markers
for CSC. In the present study, 9 KIF genes (KIF11,
KIF15, KIF2C, KIF20A, KIF20B,
KIF22, KIF23, KIFC1 and KIF4A) were
upregulated in the spheroid body-forming cells by microarray
analysis. Among the 9 KIF genes, KIF11 and
KIFC1 were expressed more than twice higher in the spheroid
body-forming cells than in the parental cells in both MKN-45 and
MKN-74 cells. Furthermore, we focused on KIFC1 since an
antibody against KIFC1 protein is commercially available. KIFC1
manages spindle length both in mitosis and meiosis using its
sliding activity along microtubules. In addition, KIFC1 mediates
proper cytokinesis by organizing and stabilizing spindles (22). However, association between KIFC1
and CSC has not been analyzed. In the present study, we showed that
both the number and size of spheres from GC cell lines were
significantly reduced in KIFC1 siRNA-trasfected cells than
in negative control siRNA-transfected cells. These results suggest
that KIFC1 is required for sphere formation in GC cells.
In non-neoplastic gastric mucosa, nuclear staining
of KIFC1 was observed in the epithelial cells of gastric isthmus,
which contains gastric stem cells (19). It has been reported that
ASPM, which encodes abnormal spindle microtubule assembly,
is also expressed in gastric isthmus (23). ASPM is a microtubule minus
end-associated protein, and is associated with controlling
self-renewal and symmetrical cell division in stem/progenitor cells
(24), suggesting that spindle
organization by KIFC1 and ASPM plays an important role in gastric
stem cells. It is well known that one of characteristics of stem
cells is drug-resistance (25). It
has been reported that expression of endogenous microtubule
depolymerizing factors may favor the development of
docetaxel-resistance (26). In
fact, overexpression of KIFC1 increases resistance to docetaxel
(13). Therefore, expression of
KIFC1 can confer resistance to docetaxel in the epithelial cells of
gastric isthmus.
In summary, we found that KIFC1 is overexpressed in
GC. Since CSC was characterized as a minority population (<5%)
of cells (27), KIFC1 is not a
specific marker for gastric CSC. However, since knockdown of KIFC1
by RNAi inhibits sphere formation, KIFC1 likely plays an important
role in CSC.
Acknowledgments
We thank Mr. Shinichi Norimura for excellent
technical assistance and advice. The present study was carried out
with the kind cooperation of the Research Center for Molecular
Medicine, Faculty of Medicine, Hiroshima University. We thank the
Analysis Center of Life Science, Hiroshima University, for the use
of their facilities. The present study was supported in part by
Grants-in-Aid for Cancer Research from the Ministry of Education,
Culture, Sports, Science and Technology of Japan and from the
Ministry of Health, Labour and Welfare of Japan.
References
|
1
|
Oue N, Sentani K, Sakamoto N and Yasui W:
Clinicopathologic and molecular characteristics of gastric cancer
showing gastric and intestinal mucin phenotype. Cancer Sci.
106:951–958. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Aung PP, Oue N, Mitani Y, Nakayama H,
Yoshida K, Noguchi T, Bosserhoff AK and Yasui W: Systematic search
for gastric cancer-specific genes based on SAGE data: Melanoma
inhibitory activity and matrix metalloproteinase-10 are novel
prognostic factors in patients with gastric cancer. Oncogene.
25:2546–2557. 2006. View Article : Google Scholar
|
|
3
|
Oue N, Hamai Y, Mitani Y, Matsumura S,
Oshimo Y, Aung PP, Kuraoka K, Nakayama H and Yasui W: Gene
expression profile of gastric carcinoma: Identification of genes
and tags potentially involved in invasion, metastasis, and
carcinogenesis by serial analysis of gene expression. Cancer Res.
64:2397–2405. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Katsuno Y, Ehata S, Yashiro M, Yanagihara
K, Hirakawa K and Miyazono K: Coordinated expression of REG4 and
aldehyde dehydrogenase 1 regulating tumourigenic capacity of
diffuse-type gastric carcinoma-initiating cells is inhibited by
TGF-β. J Pathol. 228:391–404. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
van der Flier LG, Haegebarth A, Stange DE,
van de Wetering M and Clevers H: OLFM4 is a robust marker for stem
cells in human intestine and marks a subset of colorectal cancer
cells. Gastroenterology. 137:15–17. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bessède E, Dubus P, Mégraud F and Varon C:
Helicobacter pylori infection and stem cells at the origin of
gastric cancer. Oncogene. 34:2547–2555. 2015. View Article : Google Scholar
|
|
7
|
Wakamatsu Y, Sakamoto N, Oo HZ, Naito Y,
Uraoka N, Anami K, Sentani K, Oue N and Yasui W: Expression of
cancer stem cell markers ALDH1, CD44 and CD133 in primary tumor and
lymph node metastasis of gastric cancer. Pathol Int. 62:112–119.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Takaishi S, Okumura T and Wang TC: Gastric
cancer stem cells. J Clin Oncol. 26:2876–2882. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ando A, Kikuti YY, Kawata H, Okamoto N,
Imai T, Eki T, Yokoyama K, Soeda E, Ikemura T, Abe K, et al:
Cloning of a new kinesin-related gene located at the centromeric
end of the human MHC region. Immunogenetics. 39:194–200. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
DeLuca JG, Newton CN, Himes RH, Jordan MA
and Wilson L: Purification and characterization of native
conventional kinesin, HSET, and CENP-E from mitotic HeLa cells. J
Biol Chem. 276:28014–28021. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rath O and Kozielski F: Kinesins and
cancer. Nat Rev Cancer. 12:527–539. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pannu V, Rida PC, Ogden A, Turaga RC,
Donthamsetty S, Bowen NJ, Rudd K, Gupta MV, Reid MD, Cantuaria G,
et al: HSET overexpression fuels tumor progression via centrosome
clustering-independent mechanisms in breast cancer patients.
Oncotarget. 6:6076–6091. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
De S, Cipriano R, Jackson MW and Stark GR:
Overexpression of kinesins mediates docetaxel resistance in breast
cancer cells. Cancer Res. 69:8035–8042. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fujita T, Chiwaki F, Takahashi RU, Aoyagi
K, Yanagihara K, Nishimura T, Tamaoki M, Komatsu M, Komatsuzaki R,
Matsusaki K, et al: Identification and characterization of
CXCR4-positive gastric cancer stem cells. PLoS One.
10:e01308082015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kondo T, Oue N, Yoshida K, Mitani Y, Naka
K, Nakayama H and Yasui W: Expression of POT1 is associated with
tumor stage and telomere length in gastric carcinoma. Cancer Res.
64:523–529. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sakamoto N, Oue N, Sentani K, Anami K,
Uraoka N, Naito Y, Oo HZ, Hinoi T, Ohdan H, Yanagihara K, et al:
Liver-intestine cadherin induction by epidermal growth factor
receptor is associated with intestinal differentiation of gastric
cancer. Cancer Sci. 103:1744–1750. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yasui W, Ayhan A, Kitadai Y, Nishimura K,
Yokozaki H, Ito H and Tahara E: Increased expression of p34cdc2 and
its kinase activity in human gastric and colonic carcinomas. Int J
Cancer. 53:36–41. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kwon M, Godinho SA, Chandhok NS, Ganem NJ,
Azioune A, Thery M and Pellman D: Mechanisms to suppress multipolar
divisions in cancer cells with extra centrosomes. Genes Dev.
22:2189–2203. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fukaya M, Isohata N, Ohta H, Aoyagi K,
Ochiya T, Saeki N, Yanagihara K, Nakanishi Y, Taniguchi H, Sakamoto
H, et al: Hedgehog signal activation in gastric pit cell and in
diffuse-type gastric cancer. Gastroenterology. 131:14–29. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mayer B, Klement G, Kaneko M, Man S, Jothy
S, Rak J and Kerbel RS: Multicellular gastric cancer spheroids
recapitulate growth pattern and differentiation phenotype of human
gastric carcinomas. Gastroenterology. 121:839–852. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ishimoto T, Nagano O, Yae T, Tamada M,
Motohara T, Oshima H, Oshima M, Ikeda T, Asaba R, Yagi H, et al:
CD44 variant regulates redox status in cancer cells by stabilizing
the xCT subunit of system xc− and thereby promotes tumor
growth. Cancer Cell. 19:387–400. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kim N and Song K: KIFC1 is essential for
bipolar spindle formation and genomic stability in the primary
human fibroblast IMR-90 cell. Cell Struct Funct. 38:21–30. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vange P, Bruland T, Beisvag V, Erlandsen
SE, Flatberg A, Doseth B, Sandvik AK and Bakke I: Genome-wide
analysis of the oxyntic proliferative isthmus zone reveals ASPM as
a possible gastric stem/progenitor cell marker overexpressed in
cancer. J Pathol. 237:447–459. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Higgins J, Midgley C, Bergh AM, Bell SM,
Askham JM, Roberts E, Binns RK, Sharif SM, Bennett C, Glover DM, et
al: Human ASPM participates in spindle organisation, spindle
orientation and cytokinesis. BMC Cell Biol. 11:852010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Stojnev S, Krstic M, Ristic-Petrovic A,
Stefanovic V and Hattori T: Gastric cancer stem cells: Therapeutic
targets. Gastric Cancer. 17:13–25. 2014. View Article : Google Scholar
|
|
26
|
Jordan MA and Wilson L: Microtubules as a
target for anticancer drugs. Nat Rev Cancer. 4:253–265. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Al-Hajj M, Wicha MS, Benito-Hernandez A,
Morrison SJ and Clarke MF: Prospective identification of
tumorigenic breast cancer cells. Proc Natl Acad Sci USA.
100:3983–3988. 2003. View Article : Google Scholar : PubMed/NCBI
|