Introduction
Hepatocellular carcinoma (HCC) is one of the most
common and aggressive malignancies worldwide (1). The major risk factors contributing to
HCC include infection with hepatitis B or C virus, non-alcoholic
steatohepatitis disease, aflatoxin B exposure and chronic
alcoholism (2,3). It is widely accepted that
hepatocarcinogenesis is a multi-step process of serial genetic and
epigenetic abnormalities (4–6). DNA
methylation, the best-characterized epigenetic mechanism, has been
reported as one of the pivotal alterations in HCC development and
progression (7,8). Global genomic hypomethylation and gene
specific hypermethylation are two main patterns of aberrant DNA
methylation, and these alterations coexist in HCC. Growing evidence
showed that aberrant methylation presents potential clinical
applications for HCC detection, diagnosis and prognosis (9,10).
Therefore, evaluating the status of DNA methylation could aid in
identifying molecular biomarkers that may have potential
applications in clinical practice for HCC.
TRIM58 (tripartite motif containing 58) is a member
of the tripartite motif-containing family. The TRIM proteins
frequently possess E3 ubiquitin ligase activities and participate
in a broad range of physiological processes and disease, including
innate immunity, development process, genetic diseases, and cancer
(11). It has been reported that
TRIM58 regulated terminal erythroid cell cycles and enucleation
(12). However, the association
between TRIM58 and HCC has not been well documented.
Recently, hypermethylation of TRIM58 has been detected in
hepatocellular carcinoma using methylation microarrays, and further
validated in 10 paired tumor and adjacent liver tissues using
combined bisulfite restriction analysis (COBRA) (13). Additionally, Tao et al also
found that TRIM58 mRNA expression was upregulated after
5-aza-dc treatment in HCC cell lines, which indicated that
TRIM58 methylation was tightly associated with its
downregulation in HCC cell lines. However, this observation based
on cell lines was not validated in clinical specimens. Therefore,
it is necessary to revaluate the methylation and expression of
TRIM58 in a larger cohort of clinical samples and explore
its potential clinical implication.
In the present study we measured the methylation
level and mRNA expression of TRIM58 in HCC and adjacent
non-tumor tissues by MSRE-qPCR and quantitative real-time PCR.
Correlations between the methylation and mRNA expression, and
clinicopathological features were evaluated.
Materials and methods
Tissue samples
A total of 181 HCC tissues and 172 adjacent
non-tumor tissues (including 172 paired samples) were obtained from
HCC patients who underwent surgical resections from May 2011 to
November 2014 at Zhongnan Hospital of Wuhan University and Hubei
Cancer Hospital. In addition, 13 normal tissues from patients with
hepatic hemangiomas were included as control. Among these samples,
76 pairs of HCC tissues and matched adjacent non-tumor tissues were
fresh-frozen tissues that were stored at −80°C immediately after
surgery, whereas the others were formalin-fixed paraffin-embedded
(FFPE) samples.
All patients were diagnosed by ultrasonography or
computed tomography, and confirmed by histology of liver biopsy.
None of the patients had an additional history of a solid organ
tumor or preoperative therapy. This study was approved by the
Medical Ethical Committee of Zhongnan Hospital of Wuhan University
and followed the tenets of the Declaration of Helsinki and its
later amendments. Informed consent was obtained from all
individuals before the study carried out. The demographic
clinicopathological information was obtained from the electronic
medical records of patients, which are shown in Table I.
 | Table IClinicopathological characteristics
of patients with HCC. |
Table I
Clinicopathological characteristics
of patients with HCC.
|
Characteristics | No. (%) |
|---|
| Total case
number | 181 |
| Gender | |
| Male | 154 (85.08%) |
| Female | 27 (14.92%) |
| Age at
diagnosis | |
| <55 years | 96 (53.04%) |
| ≥55 years | 83 (45.86%) |
| Missing | 2 (1.10%) |
| Mean ± SD | 52.82±11.06 |
| Median
(interquartile range) | 54 (45–61) |
| Etiology | |
| HBV (+) HCV
(−) | 166 (91.71%) |
| HBV (−) HCV
(+) | 2 (1.10%) |
| HBV (+) HCV
(+) | 6 (3.31%) |
| HBV (−) HCV
(−) | 2 (1.10%) |
| Missing | 5 (2.76%) |
| Child-Pugh
class | |
| A | 167 (92.27%) |
| B | 9 (4.97%) |
| C | 0 (0.0%) |
| Missing | 5 (2.76%) |
| Serum AFP
(ng/ml) | |
| Negative
(<20) | 59 (32.60%) |
| Positive
(≥20) | 114 (62.98%) |
| Missing | 8 (4.42%) |
| Negative
(<200) | 96 (53.04%) |
| Positive
(≥200) | 77 (42.54%) |
| Cirrhosis | |
| Negative | 36 (19.89%) |
| Positive | 139 (76.80%) |
| Missing | 6 (3.31%) |
| Size of tumor
(cm) | |
| <5.0 | 62 (34.25%) |
| ≥5.0 | 114 (62.98%) |
| Missing | 5 (2.76%) |
| Number of
tumor(s) | |
| Single | 142 (78.45%) |
| Multiple | 34 (18.78%) |
| Missing | 5 (2.76%) |
| Tumor embolus | |
| Negative | 140 (77.35%) |
| Positive | 35 (19.34%) |
| Missing | 6 (3.31%) |
| Differentiation
(Edmondson) | |
| I | 16 (8.84%) |
| II | 109 (60.22%) |
| III | 50 (27.62%) |
| IV | 2 (1.10%) |
| Missing | 4 (2.21%) |
| TNM stage | |
| I | 105 (58.01%) |
| II | 17 (9.39%) |
| III | 54 (29.83%) |
| IV | 0 (0.0%) |
| Missing | 5 (2.76%) |
Bisulfite genomic sequencing (BGS) and
methylation-sensitive restriction enzyme digestion and quantitative
PCR (MSRE-qPCR)
DNA from FFPE samples and fresh-frozen tissues was
isolated using QIAamp DNA FFPE Tissue kit (Qiagen) and standard
phenol/chloroform extraction, respectively. DNA was quantified by
the NanoDrop-2000c (Thermo Scientific, Rockford, IL, USA) and
stored at −20°C until use. Bisulphite conversion and bisulfite
genomic sequencing (BGS) were performed as previously described
(14). The methylation level of
TRIM58 was calculated as follows: 100% × methylated CG /
(methylated CG + unmethylated CG). MSRE-qPCR was performed to
detect the methylation level of TRIM58 as our previous study
(14). ACTB without
methylation-sensitive restriction enzyme recognition site was used
as an internal control. The methylation level of TRIM58 was
measured by 100% × 2−ΔCq (undigested − digested) after
normalization to the ACTB (15). The primers used for BGS and
MSRE-qPCR were listed in Table
II.
| Table IIThe list of primer sequences. |
Table II
The list of primer sequences.
| Primer name | Forward primer
(5′→3′) | Reverse primer
(5′→3′) | Amplicon length
(bp) | Tm (°C) | Assay |
|---|
| TRIM58-BGS1 |
AATAAAGATGGAAGGAAGG |
AAACAAAAACTATAACCGC | 444 | Touch-down | BGS |
| TRIM58-BGS2 |
GAGGAGGGATTTTAGTTAGAAATGTTTA |
ACTCCTACAAAAAATCCAAACACAC | 343 | Touch-down | BGS |
| TRIM58-MSRE |
TGTGCAGACCGCGAGGG |
GAAATCCAGGCACACCGGG | 129 | 64.5 | MSRE-qPCR |
| ACTB-MSRE |
TGGCACCACACCTTCTACAA |
CCACTCACCTGGGTCATCTT | 116 | 64.5 | MSRE-qPCR |
| TRIM58-RT |
GGCTGCTGAAAGGGAATGAGT |
CCGAAGAAGACCAGAGAAGAGTTG | 182 | 65.5 | RT-PCR |
| ACTB-RT |
CAGAGCCTCGCCTTTGCC |
CATGCCGGAGCCGTTGTC | 102 | 65.5 | RT-PCR |
RNA isolation and gene expression
analysis
Total RNA was isolated from fresh-frozen tissues
using TRIzol Reagent (Life Technologies, Gaithersburg, MD, USA)
according to the manufacturer's instructions. The cDNA was
synthesized from 1 µg RNA by PrimeScript™ RT reagent kit
with gDNA Eraser (Takara, Dalian, China). The expression level of
TRIM58 was performed in duplicate by quantitative PCR using
Bio-Rad CFX96™ real-time PCR detection system (Bio-Rad, Hercules,
CA, USA) in accordance with the manufacturer's protocol. Each assay
was carried out in a total volume of 20 µl, including 1X
SYBR® Premix Ex Taq™ GC (Takara), 0.5 µM of
forward and reverse primers (Table
II), 2 µl cDNA template. Non-template control was used
as negative control, and melting curve analysis was employed to
verify the specificity of PCR product. All reactions were
normalized to ACTB as an internal control, and the relative
expression level of TRIM58 was determined by using the
comparative Cq method (2−ΔCq).
Statistical analysis
The comparison of TRIM58 methylation level
between various subgroups was conducted using Wilcoxon matched
pairs test or Mann-Whitney U test where necessary. Paired-samples
T-test was used to compare the expression of TRIM58 (log10
transformation) between HCC tissues and adjacent non-tumor tissues.
The correlation between two continuous variables was evaluated by
Spearman's rank correlation. For survival analysis, Kaplan-Meier
method with log-rank test and COX regression model were performed
to elucidate the prognostic significance of TRIM58
methylation. The statistical analyses were conducted with the SPSS
16.0 and GraphPad Prism 6.0, all p-values were two-sided and
p<0.05 was considered statistically significant.
Results
Hypermethylation of TRIM58 in HCC
To investigate the methylation status of
TRIM58 in HCC, we firstly evaluated the methylation level in
30 paired HCC tissues and adjacent non-tumor tissues using BGS. A
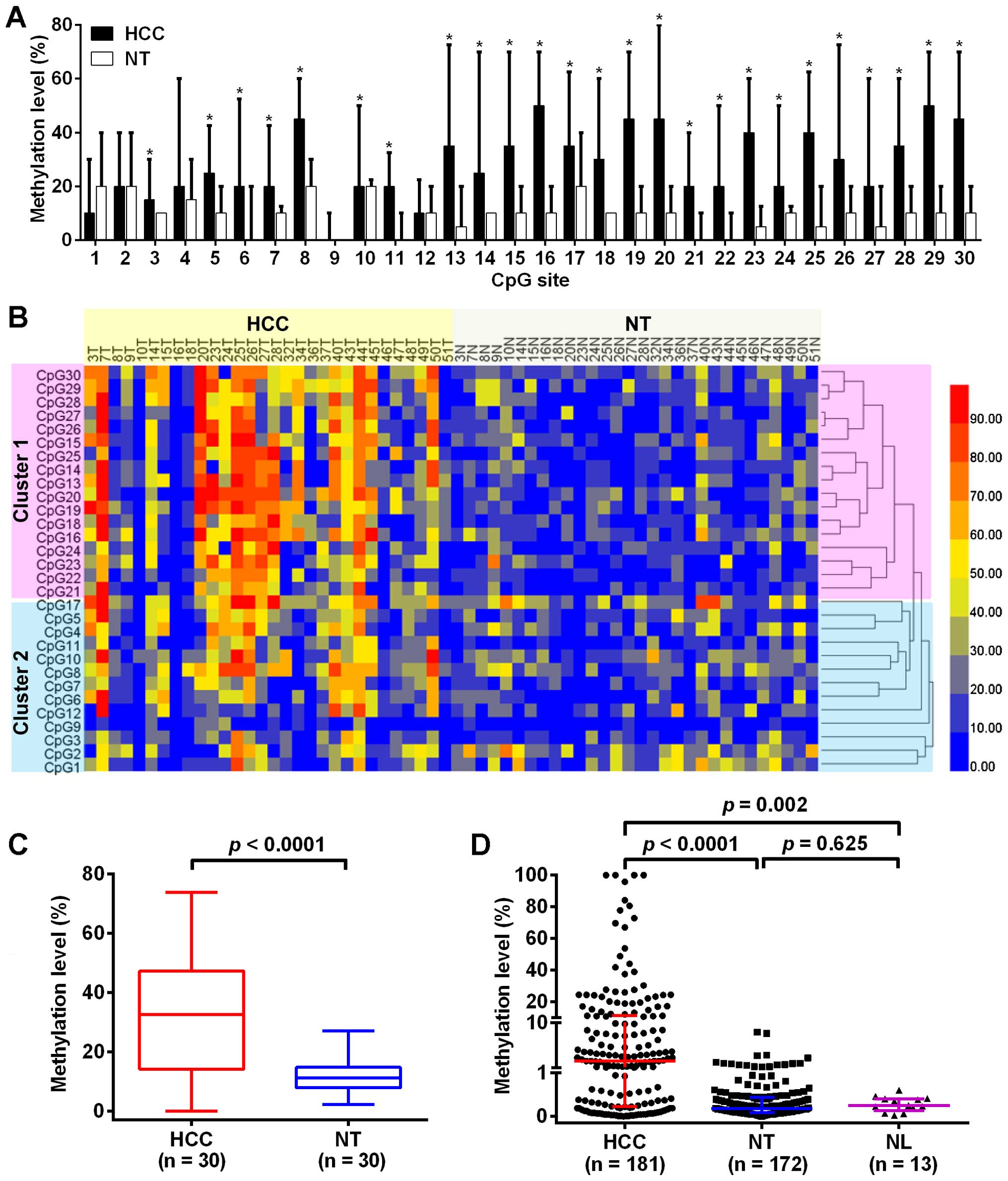
total of 30 CpG sites were detected and the methylation levels were
varied in different CpG sites (Fig.
1A). The methylation level of TRIM58 in HCC tissues were
significantly higher than that in paired adjacent non-tumor tissues
at each CpG site (p<0.05), excepting the CpG sites at 1, 2, 4, 9
and 12 (p>0.05). A hierarchical clustering analysis was
performed according to the methylation level of each CpG site in 60
liver tissues, and the 30 CpG sites were divided into two distinct
subclasses, cluster 1 and cluster 2 (Fig. 1B). The methylation differences
between HCC and adjacent non-tumor tissues at CpG sites in cluster
1 were more obvious than in cluster 2. The overall methylation of
TRIM58 in HCC tissues was also significantly higher than
that in matched adjacent non-tumor tissues [32.50% (14.17%–47.17%)
vs. 11.17% (7.92%–14.75%), p<0.0001, Fig. 1C].
To further explore the association between TRIM58
methylation and clinicopathological characteristics, 181 HCC
tissues, 172 matched adjacent non-tumor tissues and 13 normal liver
tissues were used to verify the methylation level of TRIM58 in the
region between CpG site 13 and 30 (cluster 1) using MSRE-qPCR.
Representative results of BGS and MSRE-qPCR are displayed in
Fig. 2. The result from MSRE-qPCR
showed that the methylation level of TRIM58 in HCC tissues
was significantly higher than that in adjacent non-tumor tissues
and normal liver tissues (p<0.0001 and p=0.002, respectively,
Fig. 1D). However, there was no
significant difference in methylation level of TRIM58
between the adjacent non-tumor tissues and normal liver tissues
(p=0.625, Fig. 1D). With a 10%
hypermethylation level threshold established (16,17),
28.18% (51/181) of HCC specimens showed increased hypermethylation,
as opposed to 0% in the adjacent non-tumor tissues and normal liver
tissues. Overall, both data from BGS and MSRE-qPCR indicated a
significant increase in the methylation level of TRIM58 in
HCC tissues compared with the adjacent non-tumor tissues.
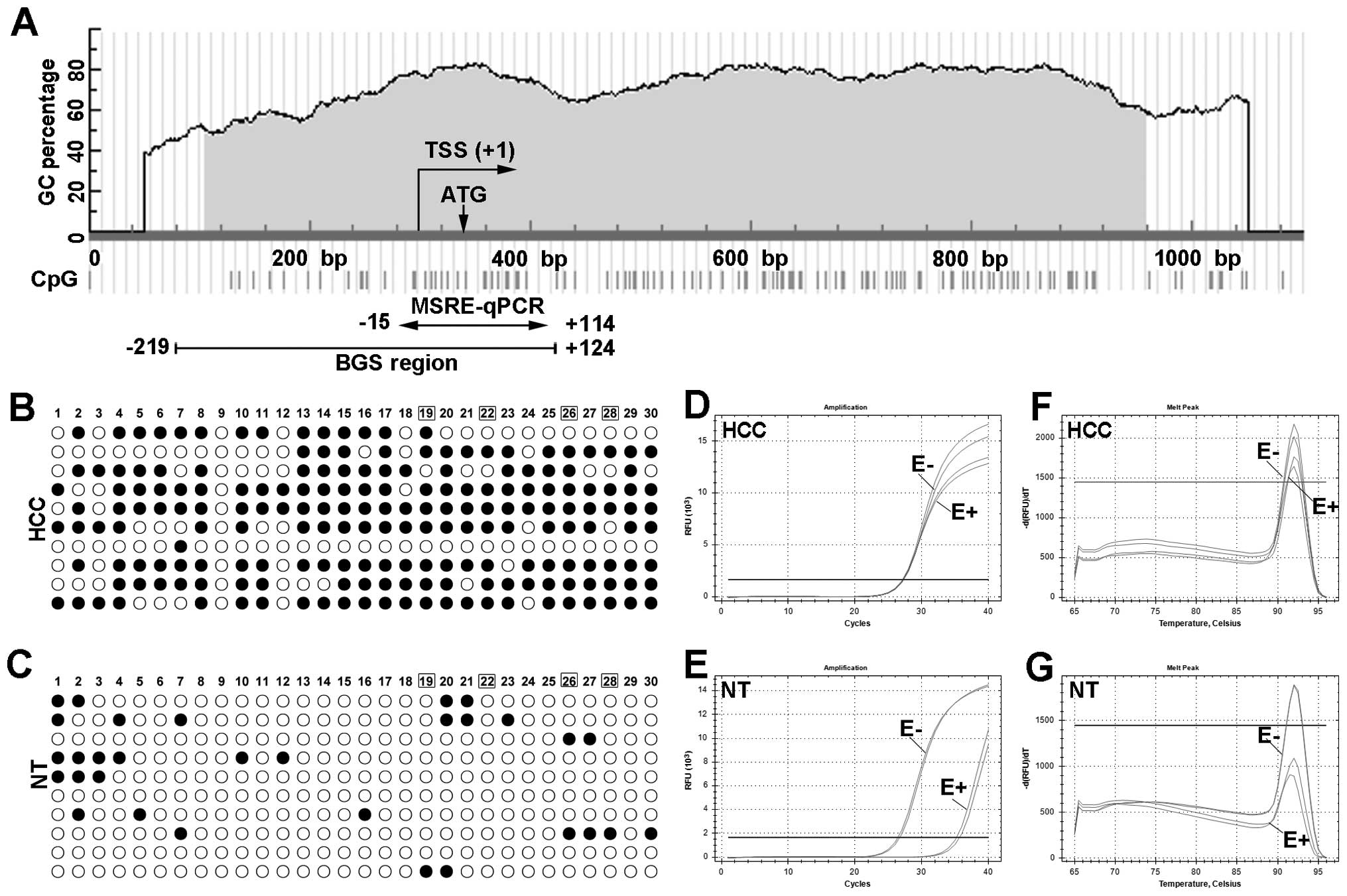 | Figure 2Representative results of
TRIM58 methylation in HCC and adjacent non-tumor liver
tissues using BGS and MSRE-qPCR. (A) Schematic structure of
TRIM58 CGI, including the transcript start site (TSS),
translation initiation site (ATG), BGS and MSRE-qPCR region. The
position of BGS and MSRE-qPCR are marked with arrows, and the
number indicates the relative position to TSS (TSS was considered
as +1). (B and C) TRIM58 methylation in HCC and adjacent
non-tumor liver tissues using BGS. Each row represents the
sequencing result from one clone, and the number represents the
position of CpG sites in target region (number 1 indicate the first
CpG site in the target region). Open and closed circles represent
methylated and unmethylated CG dinucleotides, respectively. The CpG
sites within recognition sites of MSRE are marked in box (19, 22,
26 and 28). MSRE-qPCR results for the same sample of BGS are
displayed as amplification curve (D and E) and melt curve (F and
G). The line marked as E+ and E- indicate digested sample and
undigested sample for TRIM58, respectively. HCC,
hepatocellular carcinoma tissues; NT, adjacent non-tumor
tissues. |
Correlations between TRIM58 methylation
and clinico-pathological features
The correlation between TRIM58 methylation
and clinicopathological features of 181 HCC samples are summarized
in Table III. The methylation
level of TRIM58 was increased in patients with serum AFP
≥200 ng/ml (p= 0.015), tumor embolus (p= 0.0026) and advanced
tumor-node-metastasis (TNM) stage (p=0.046), respectively. No other
correlations were observed between the methylation of TRIM58
and clinicopathological parameters, such as patient gender, age,
cirrhosis, tumor number and size, as well as tumor differentiation
(p>0.05). Spearman's rank order correlation coefficient was
calculated to further analyze associations between TRIM58
methylation and AFP, tumor number and size as continuous variables.
The results indicated that the methylation of TRIM58 had a
positive significant correlations with AFP level (p=0.037,
rs=0.159). However, no correlations between
TRIM58 methylation and tumor number and size were
observed.
| Table IIIAssociations between TRIM58
methylation and clinicopathological parameters in 181 HCC
patients. |
Table III
Associations between TRIM58
methylation and clinicopathological parameters in 181 HCC
patients.
|
Characteristics | No. (%) | Methylation level
(%)a | p-valueb |
|---|
| Gender | | | 0.992 |
| Male | 154 (85.08%) | 2.54
(0.21–12.37) | |
| Female | 27 (14.92%) | 1.38
(0.32–7.57) | |
| Age at
diagnosis | | | 0.073 |
| <55 years | 96 (53.04%) | 3.00
(0.38–18.82) | |
| ≥55 years | 83 (45.86%) | 1.04
(0.14–10.86) | |
| Missing | 2 (1.10%) | | |
| Serum AFP
(ng/ml) | | | 0.397 |
| Negative
(<20) | 59 (32.60%) | 2.14
(0.19–7.57) | |
| Positive
(≥20) | 114 (62.98%) | 2.73
(0.22–15.6) | |
| Missing | 8 (4.42%) | | |
| Serum AFP
(ng/ml) | | | 0.015 |
| Negative
(<200) | 96 (53.04%) | 17.98
(0.84–58.83) | |
| Positive
(≥200) | 77 (42.54%) | 25.12
(2.21–56.08) | |
| Missing | 8 (4.42%) | | |
| Cirrhosis | | | 0.444 |
| Negative | 36 (19.89%) | 0.99
(0.08–19.59) | |
| Positive | 139 (76.80%) | 2.83
(0.25–10.86) | |
| Missing | 6 (3.31%) | | |
| Size of tumor
(cm) | | | 0.229 |
| <5.0 | 62 (34.25%) | 1.33
(0.24–7.00) | |
| ≥5.0 | 114 (62.98%) | 3.22
(0.19–17.00) | |
| Missing | 5 (2.76%) | | |
| Number of
tumor(s) | | | 0.326 |
| Single | 142 (78.45%) | 2.20
(0.21–10.92) | |
| Multiple | 34 (18.78%) | 3.4
(0.36–11.98) | |
| Missing | 5 (2.76%) | | |
| Tumor embolus | | | 0.026 |
| Negative | 140 (77.35%) | 1.92
(0.18–10.58) | |
| Positive | 35 (19.34%) | 3.51
(1.12–19.29) | |
| Missing | 6 (3.31%) | | |
| Differentiation
(Edmonson) | | | 0.439 |
| I + II | 125 (69.06%) | 2.14
(0.22–10.18) | |
| III + IV | 52 (28.73%) | 3.85
(0.22–18.82) | |
| Missing | 4 (2.21%) | | |
| TNM stage | | | 0.046 |
| I + II | 122 (67.40%) | 1.31
(0.19–9.03) | |
| III + IV | 54 (29.83%) | 3.65
(0.86–17.54) | |
| Missing | 5 (2.76%) | | |
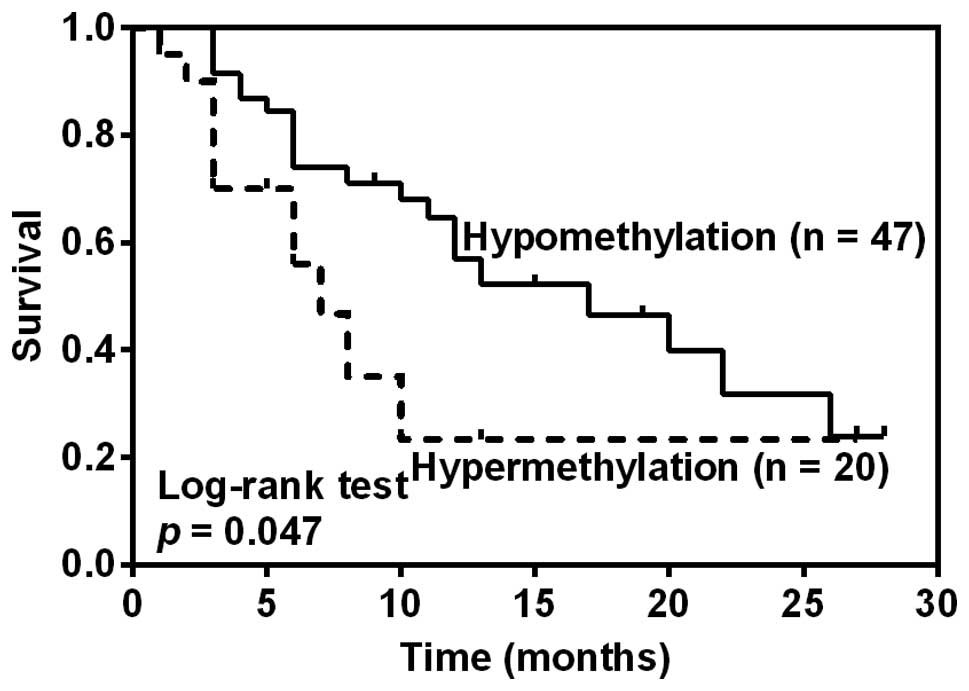
TRIM58 hypermethylation tend to be
associated with worse disease-free survival
The follow-up began with the date of HCC resection
and ended with the date of death or the last clinical review before
June 30, 2015. Survival analysis was finally conducted in 67 HCC
patients with the follow-up time more than 3 months. The median
follow-up time was 10 months (range, 3–33 months). The median
disease-free survival time (DFS, defined as survival without any
clinical evidence of recurrence or metastasis) was 12.33 months and
32 patients developed recurrence or metastasis. The overall
survival analysis was not performed due to the death of only two
patients. To explore the association between TRIM58
methylation and DFS, patients were divided into groups of
hypomethylation or hypermethylation according to the threshold of
TRIM58 methylation level established (10%). Kaplan-Meier
analysis revealed that HCC patients with TRIM58
hypermethylation showed a significantly shorter median DFS after
hepatectomy than those with TRIM58 hypomethylation (7 months
vs. 17 months, p=0.047, Fig. 3).
Furthermore, COX regression model showed that larger tumor size
(p=0.0002, HR=1.113, 95% CI: 1.059–1.205) and advanced
differentiation (p=0.013, HR=2.448, 95% CI: 1.206–4.972), but not
TRIM58 hypermethylation (p>0.05), were independent
prognostic predictors for unfavorable DFS (data not shown).
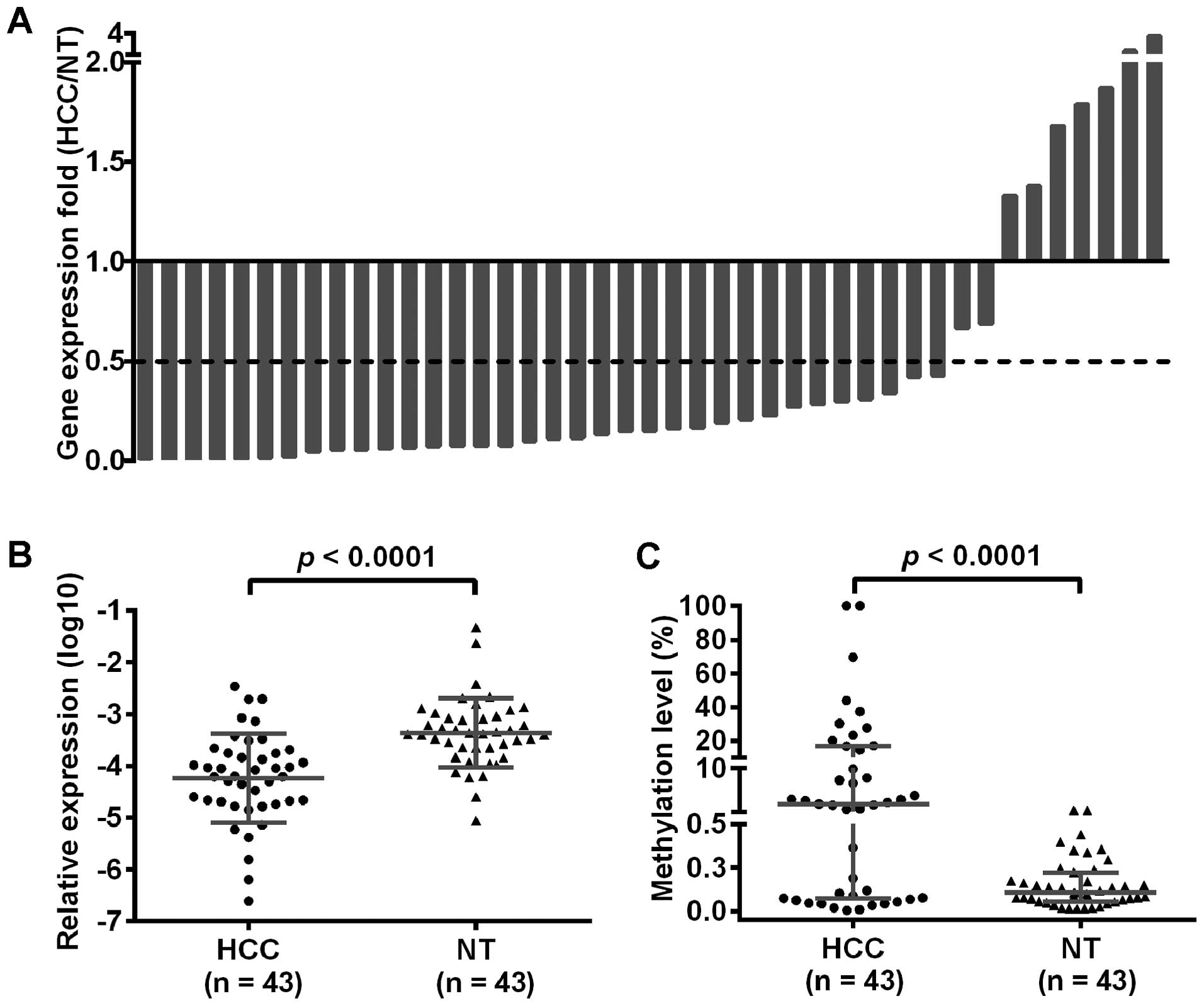
TRIM58 mRNA expression was downregulated
in HCC and inversely associated with its methylation
To further study the relationship between
TRIM58 methylation and mRNA expression, we measured the mRNA
expression of TRIM58 in 43 of the 172 paired HCC and
adjacent non-tumor tissues by quantitative real-time PCR.
Considering 0.5 as the cut-off value, TRIM58 mRNA expression
was downregulated in about 79% (34/43) of HCC tissues (Fig. 4A) and the difference was
statistically significant (p<0.0001, Fig. 4B). Moreover, TRIM58
methylation was significantly higher in HCC specimens (p<0.0001,
Fig. 4C) and had an inverse
correlation with its mRNA expression, despite a lower correlation
coefficient (p=0.015, rs= −0.260).
Discussion
It is well known that aberrant methylation plays a
pivotal role in hepatocarcinogenesis (7). TRIM58 methylation was first
reported in HCC by methylation microarrays, and validated in only
10 paired primary tumor and adjacent liver tissues (13). However, the authors did not perform
any clinical associations with their data due to the limited
samples. In this study, detailed TRIM58 methylation of 30
CpG sites at a 343 bp CGI (CpG islands) region was initially
analyzed by BGS in a pilot cohort of 30 paired HCC and adjacent
non-tumor tissues. The result indicated that the TRIM58
methylation increased significantly in HCC tissues compared with
adjacent non-tumor tissues, and the methylation difference of CpG
sites in cluster 1 (−6 ~ +99) was more severe, which was in
accordance with previous study from Tao et al (13). Furthermore, this observation was
further validated in a larger cohort of 181 HCC tissues, 172
adjacent non-tumor tissues and 13 normal liver tissues using
MSRE-qPCR. Additionally, the methylation of TRIM58 was
inversely associated with its mRNA expression, and tended to
correlate with tumor embolus, advanced TNM stage and unfavorable
DFS after hepatectomy.
Both results of BGS and MSRE-qPCR showed an
increased methylation level of TRIM58 in HCC tissues
compared with non-tumor tissues and normal liver tissues. It
further validated that gene specific hypermethylation was
considered as a common epigenetic event in HCC (8). However, the significant difference
between adjacent non-tumor tissues and normal liver tissues was not
observed in this study, and hypermethylation of TRIM58
appeared in HCC tissues, but not in adjacent non-tumor tissues and
normal liver tissues. The results indicated that hypermethylation
of TRIM58 (>10%) was one of the specific events
associated with tumorigenesis rather than other benign pathological
processes, such as inflammation and cirrhosis (18).
It is well documented that aberrant methylation
showed the potential value of prognostic prediction for patients
with HCC, which might assist in making therapeutic schedule after
hepatectomy (19,20). Consistently, we found that
hypermethylation of TRIM58 was associated with worse DFS,
although it was not an independent prognostic predictor for
unfavourable DFS. This may be due to the relative short follow-up
time and limited sample size. Nevertheless, it implied that
TRIM58 hypermethylation was not conducive to the patient's
DFS. In accordance with previous study, larger tumor size and
high-grade differentiation (Edmondson) were independent unfavorable
factors for DFS (21). In addition,
a positive correlation between TRIM58 methylation and tumor
embolus, advanced TNM stage, the characteristics of disease
progression and unfavorable prognosis (22,23),
were observed in patients with HCC. These results suggested that
TRIM58 methylation closely associated with aggressive
biological behavior of HCC and might be served as a potential
prognostic marker for HCC patients. Thus, hypermethylation of
TRIM58 might play a crucial role in hepatocarcinogenesis,
especially for the progressive and aggressive nature of this
disease.
Gene specific hypermethylation frequently mediated
transcriptional silencing of the associated gene and played an
important role in tumorigenesis (24,25).
Numerous genes, such as SOCS1 (17), GSTP1 (26), SFRP1 (27), FOXD3 (28), SYK (29), FAM43B (30), which are involved in tumor
suppression, cell cycle regulation, apoptosis, and DNA repair have
been shown to be suppressed by DNA hypermethylation in HCC. In our
study TRIM58 mRNA expression was significantly reduced in
HCC tissues comparing with adjacent non-tumor tissues, while the
assessment of DNA methylation by MSRE-qPCR demonstrated that
TRIM58 methylation was significantly increased in HCC
tissues. Furthermore, 72 h treatment with demethylation agent
(5-aza-dc) in HCC cell lines was able to upregulate TRIM58
mRNA expression (13). These
results indicated that TRIM58 methylation was inversely
correlated with the downregulation of its expression. Herein, we
speculated that TRIM58 methylation might be involved in the
mechanism of gene silencing and then participated in the
development of HCC, although it needed further investigation.
Of note, the following limitations should be
considered in the present study. Firstly, although we detected
TRIM58 methylation in HCC tissues, adjacent non-tumor
tissues and normal liver tissues, the methylation levels in tissues
both from patients of chronic hepatitis and liver cirrhosis without
HCC were unclear, which will assist in stating the continuous
changes of methylation in hepatocarcinogenesis. Secondly, the mRNA
expression analysis was conducted in a small sample size as our
study was mainly based on FFPE specimens. Finally, our follow-up
time was relatively short with limited sample size, resulting in
low power to evaluate the association between TRIM58
methylation and DFS. Thus, further validation in a prospective and
large-scale clinical study is needed.
In conclusion, our data showed that hypermethylation
of TRIM58 is one of the specific events in HCC, and may
contribute to the downregulation of its mRNA expression. Moreover,
hypermethylation of TRIM58 tend to be associated with worse
DFS after hepatectomy. However, the clinical application of
TRIM58 needs to be further assessed with more samples with
different pathophysiological processes and histological
characteristics.
Acknowledgments
This research was supported by the National Basic
Research Program of China (973 Program, 2012CB720605), the Science
and Technology Research Plan of Wuhan City (2015060101010057).
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
El-Serag HB: Epidemiology of viral
hepatitis and hepatocellular carcinoma. Gastroenterology.
142:1264–1273.e1. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Forner A, Llovet JM and Bruix J:
Hepatocellular carcinoma. Lancet. 379:1245–1255. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shibata T and Aburatani H: Exploration of
liver cancer genomes. Nat Rev Gastroenterol Hepatol. 11:340–349.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pogribny IP and Rusyn I: Role of
epigenetic aberrations in the development and progression of human
hepatocellular carcinoma. Cancer Lett. 342:223–230. 2014.
View Article : Google Scholar :
|
|
6
|
You JS and Jones PA: Cancer genetics and
epigenetics: Two sides of the same coin? Cancer Cell. 22:9–20.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Um TH, Kim H, Oh BK, Kim MS, Kim KS, Jung
G and Park YN: Aberrant CpG island hypermethylation in dysplastic
nodules and early HCC of hepatitis B virus-related human multistep
hepato-carcinogenesis. J Hepatol. 54:939–947. 2011. View Article : Google Scholar
|
|
8
|
Raggi C and Invernizzi P: Methylation and
liver cancer. Clin Res Hepatol Gastroenterol. 37:564–571. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mikeska T, Bock C, Do H and Dobrovic A:
DNA methylation biomarkers in cancer: Progress towards clinical
implementation. Expert Rev Mol Diagn. 12:473–487. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sceusi EL, Loose DS and Wray CJ: Clinical
implications of DNA methylation in hepatocellular carcinoma. HPB
Oxf. 13:369–376. 2011. View Article : Google Scholar
|
|
11
|
Napolitano LM and Meroni G: TRIM family:
Pleiotropy and diversification through homomultimer and
heteromultimer formation. IUBMB Life. 64:64–71. 2012. View Article : Google Scholar
|
|
12
|
Thom CS, Traxler EA, Khandros E, Nickas
JM, Zhou OY, Lazarus JE, Silva AP, Prabhu D, Yao Y, Aribeana C, et
al: Trim58 degrades Dynein and regulates terminal erythropoiesis.
Dev Cell. 30:688–700. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tao R, Li J, Xin J, Wu J, Guo J, Zhang L,
Jiang L, Zhang W, Yang Z and Li L: Methylation profile of single
hepatocytes derived from hepatitis B virus-related hepatocellular
carcinoma. PLoS One. 6:e198622011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Qiu X, Hu B, Huang Y, Deng Y, Wang X and
Zheng F: Hypermethylation of ACP1, BMP4, and TSPYL5 in
hepato-cellular carcinoma and their potential clinical
significance. Dig Dis Sci. 61:149–157. 2016. View Article : Google Scholar
|
|
15
|
Zhang X, Liu S, Shen C, Wu Y, Zhang L,
Chen X and Lu F: DNA methylation consistency implicates the primary
tumor cell origin of recurrent hepatocellular carcinoma.
Epigenomics. 7:581–592. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang Y, Cheng J, Xu C, Liu S, Jiang S, Xu
Q, Chen X, Zhuang H and Lu F: Quantitative methylation analysis
reveals gender and age differences in p16INK4a hypermethylation in
hepatitis B virus-related hepatocellular carcinoma. Liver Int.
32:420–428. 2012.
|
|
17
|
Zhang X, Wang J, Cheng J, Ding S, Li M,
Sun S, Zhang L, Liu S, Chen X, Zhuang H, et al: An integrated
analysis of SOCS1 down-regulation in HBV infection-related
hepatocellular carcinoma. J Viral Hepat. 21:264–271. 2014.
View Article : Google Scholar :
|
|
18
|
Gao XD, Qu JH, Chang XJ, Lu YY, Bai WL,
Wang H, Xu ZX, An LJ, Wang CP, Zeng Z, et al: Hypomethylation of
long interspersed nuclear element-1 promoter is associated with
poor outcomes for curative resected hepatocellular carcinoma. Liver
Int. 34:136–146. 2014. View Article : Google Scholar :
|
|
19
|
Villanueva A, Portela A, Sayols S,
Battiston C, Hoshida Y, Méndez-González J, Imbeaud S, Letouzé E,
Hernandez-Gea V, Cornella H, et al HEPTROMIC Consortium: DNA
methylation-based prognosis and epidrivers in hepatocellular
carcinoma. Hepatology. 61:1945–1956. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mah WC, Thurnherr T, Chow PK, Chung AY,
Ooi LL, Toh HC, Teh BT, Saunthararajah Y and Lee CG: Methylation
profiles reveal distinct subgroup of hepatocellular carcinoma
patients with poor prognosis. PLoS One. 9:e1041582014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yan T, Zhao JJ, Bi XY, Zhao H, Huang Z, Li
ZY, Zhou JG, Li Y, Li C, Cai JQ, et al: Prognosis of hepatocellular
carcinoma: A study of 832 cases. Zhonghua Zhong Liu Za Zhi.
35:54–58. 2013.In Chinese. PubMed/NCBI
|
|
22
|
Tandon P and Garcia-Tsao G: Prognostic
indicators in hepato-cellular carcinoma: a systematic review of 72
studies. Liver Int. 29:502–510. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lu W, Dong J, Huang Z, Guo D, Liu Y and
Shi S: Comparison of four current staging systems for Chinese
patients with hepatocellular carcinoma undergoing curative
resection: Okuda, CLIP, TNM and CUPI. J Gastroenterol Hepatol.
23:1874–1878. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fukushige S and Horii A: DNA methylation
in cancer: A gene silencing mechanism and the clinical potential of
its biomarkers. Tohoku J Exp Med. 229:173–185. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gan L, Chen S, Zhong J, Wang X, Lam EK,
Liu X, Zhang J, Zhou T, Yu J, Si J, et al: ZIC1 is downregulated
through promoter hypermethylation, and functions as a tumor
suppressor gene in colorectal cancer. PLoS One. 6:e169162011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhong S, Tang MW, Yeo W, Liu C, Lo YM and
Johnson PJ: Silencing of GSTP1 gene by CpG island DNA
hypermethylation in HBV-associated hepatocellular carcinomas. Clin
Cancer Res. 8:1087–1092. 2002.PubMed/NCBI
|
|
27
|
Kaur P, Mani S, Cros MP, Scoazec JY,
Chemin I, Hainaut P and Herceg Z: Epigenetic silencing of sFRP1
activates the canonical Wnt pathway and contributes to increased
cell growth and proliferation in hepatocellular carcinoma. Tumour
Biol. 33:325–336. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
He G, Hu S, Zhang D, Wu P, Zhu X, Xin S,
Lu G, Ding Y and Liang L: Hypermethylation of FOXD3 suppresses cell
proliferation, invasion and metastasis in hepatocellular carcinoma.
Exp Mol Pathol. 99:374–382. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shin SH, Lee KH, Kim BH, Lee S, Lee HS,
Jang JJ and Kang GH: Downregulation of spleen tyrosine kinase in
hepatocellular carcinoma by promoter CpG island hypermethylation
and its potential role in carcinogenesis. Lab Invest. 94:1396–1405.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xu X, Liu RF, Wan BB, Xing WM, Huang J and
Han ZG: Expression of a novel gene FAM43B repressing cell
proliferation is regulated by DNA methylation in hepatocellular
carcinoma cell lines. Mol Cell Biochem. 354:11–20. 2011. View Article : Google Scholar : PubMed/NCBI
|